
Nutritional Composition and Estimated Iron and Zinc Bioavailability of Meat Substitutes Available on the Swedish Market

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection and Preparation of Samples
2.2. Mineral Analyses
2.3. Phytate Analysis
2.4. Estimation of Iron and Zinc Bioavailability
2.5. Determination of Total Phenolic Content
2.6. Analysis of Total Fat and Fatty Acid Composition
2.7. Determination of Salt Content
2.8. Determination of Protein Content
2.9. Analysis of Amino Acid Profiles
2.10. Analysis of Dietary Fiber
2.11. Statistics
3. Results and Discussion
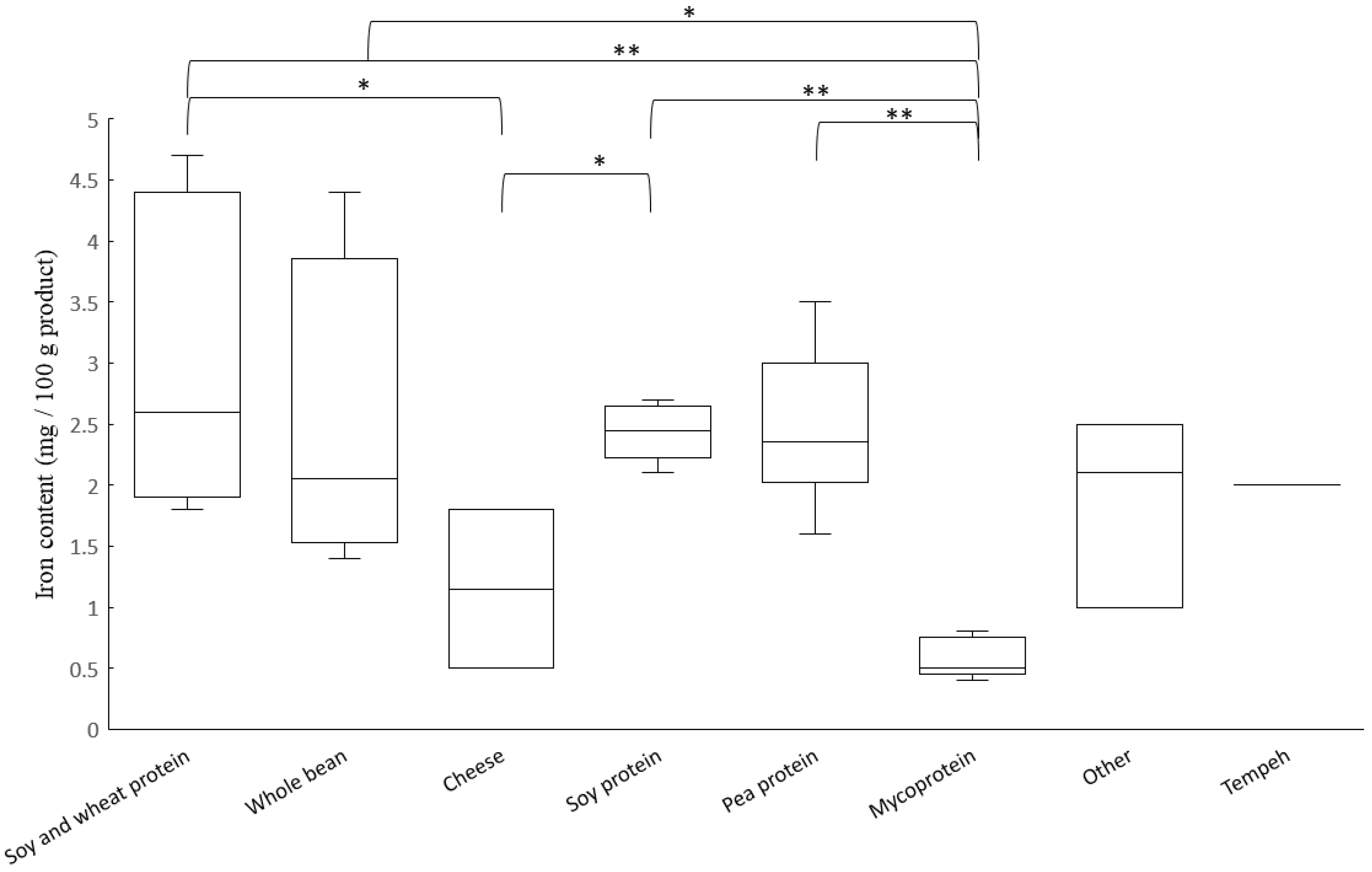
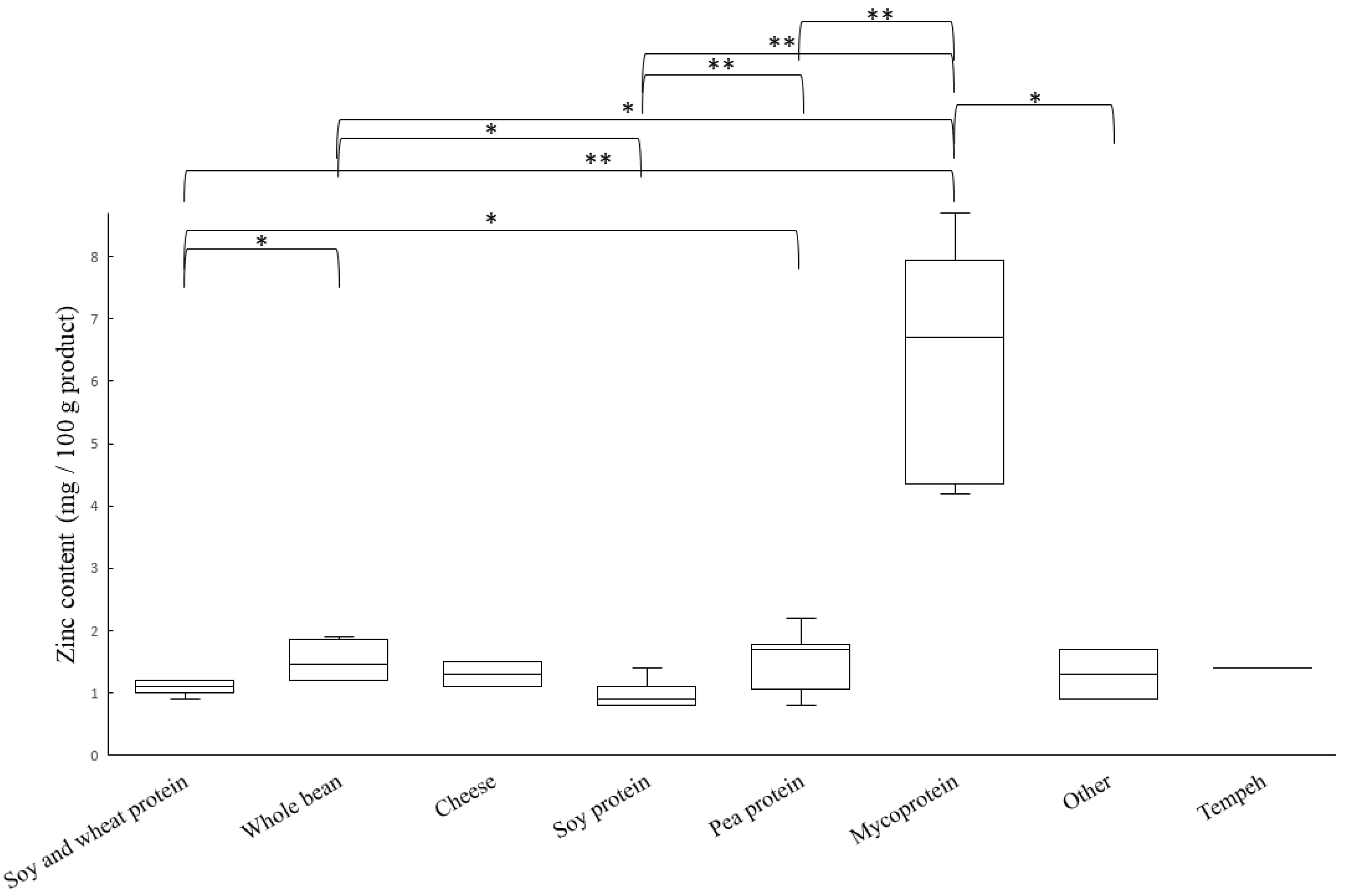
3.1. Content of Iron and Zinc
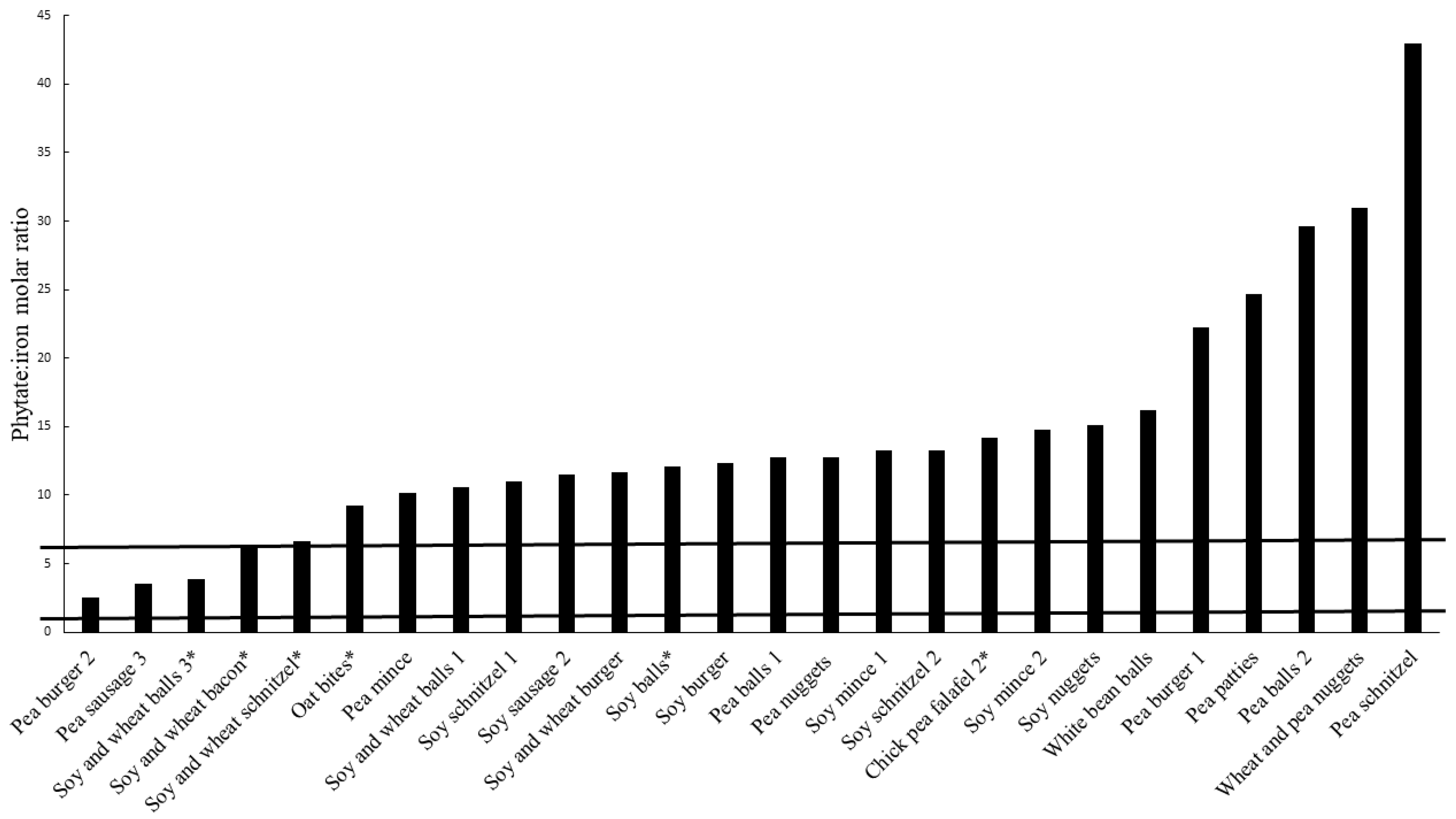
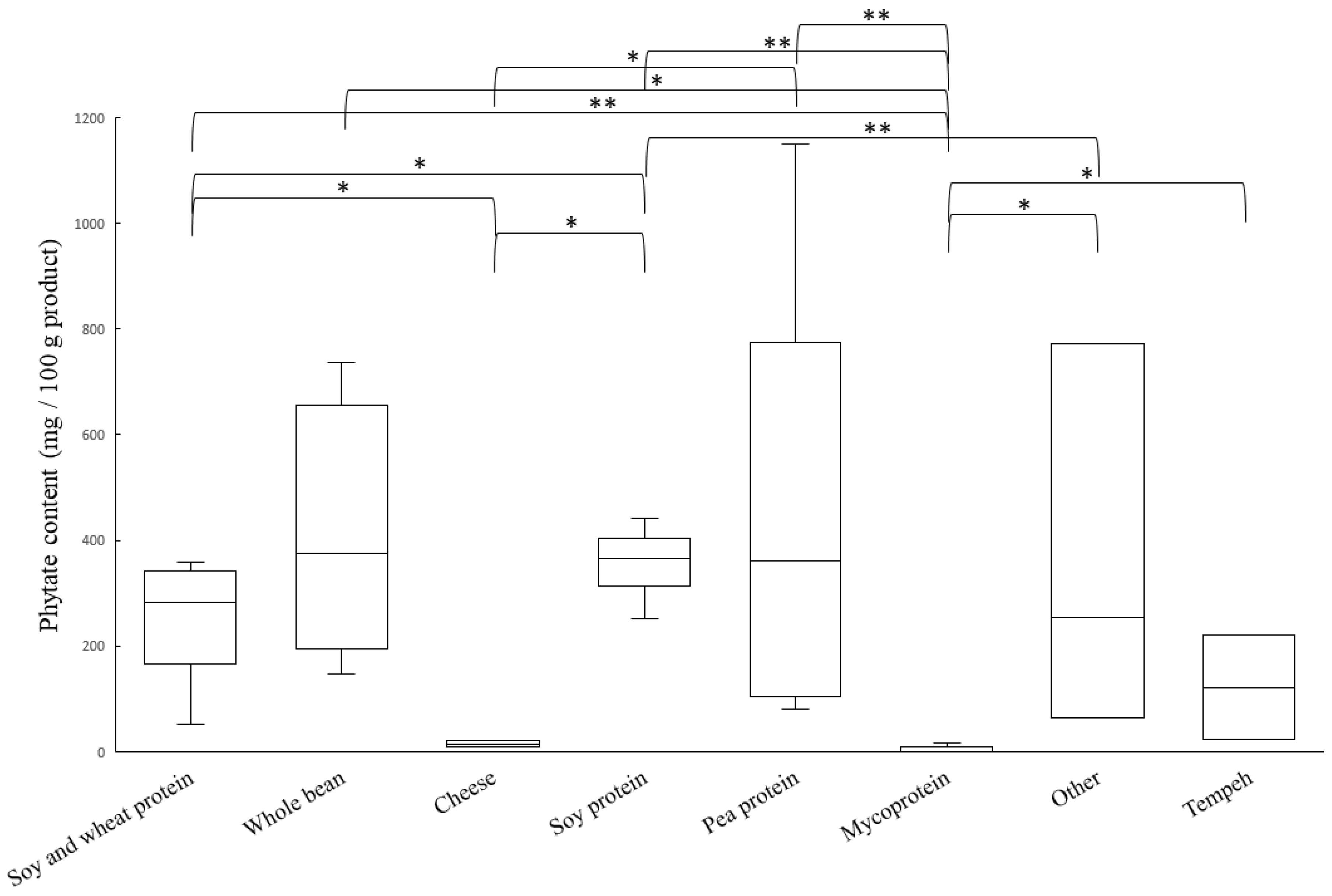
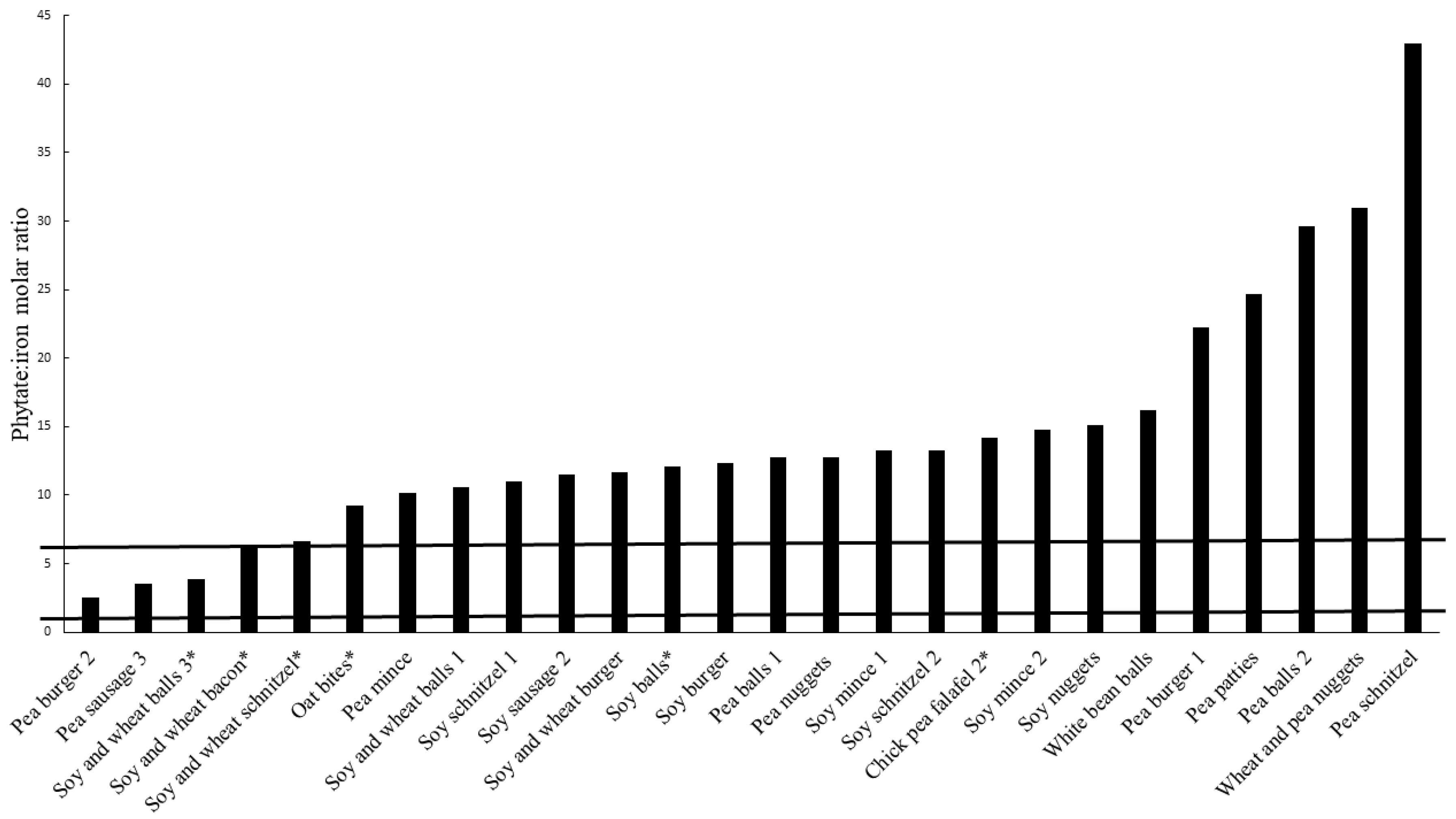
3.2. Phytate Content
3.3. Estimated Bioavailability of Iron and Zinc
3.4. Estimated Bioavailability of Zinc
3.5. Total Phenolic Content
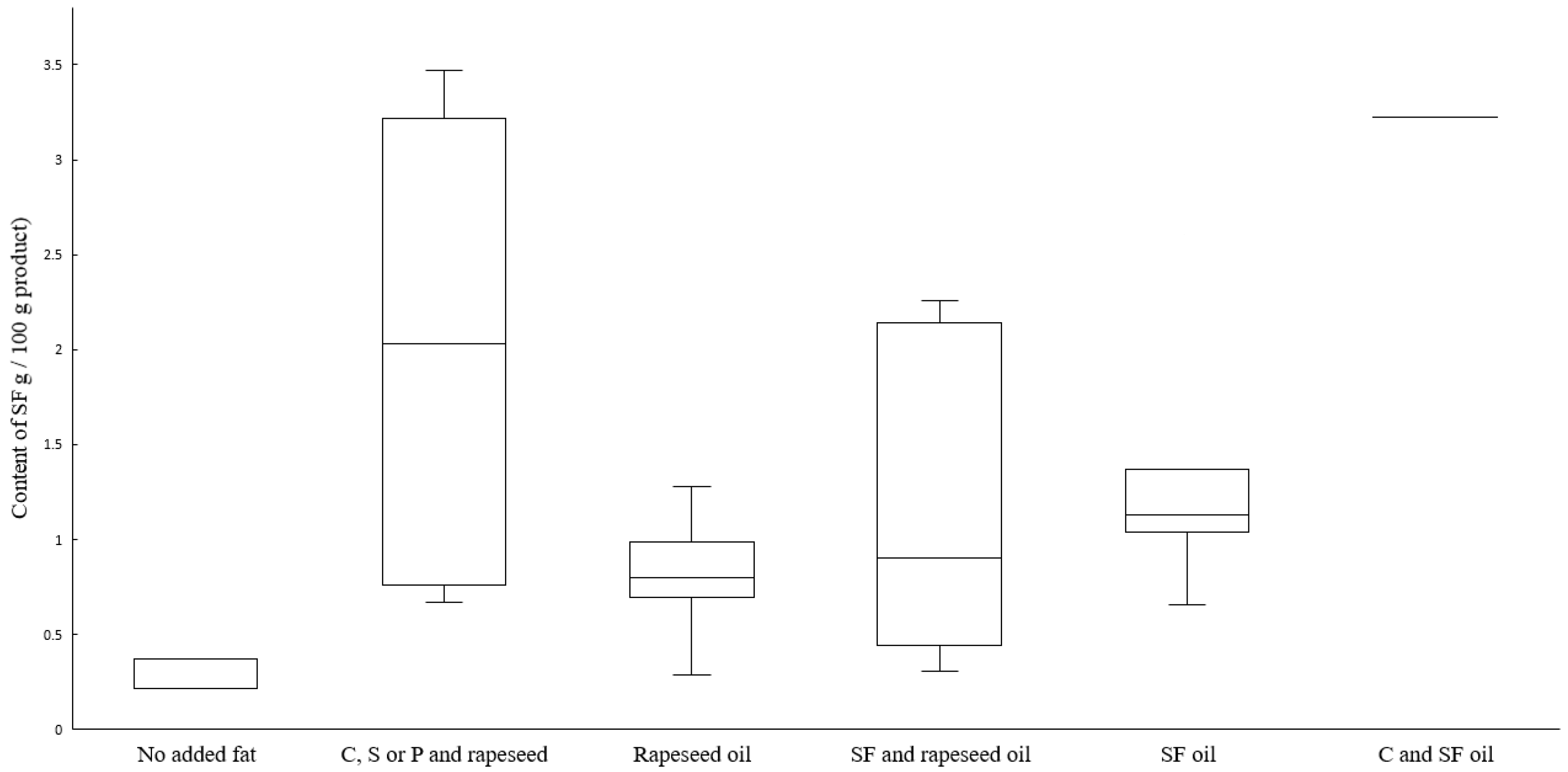
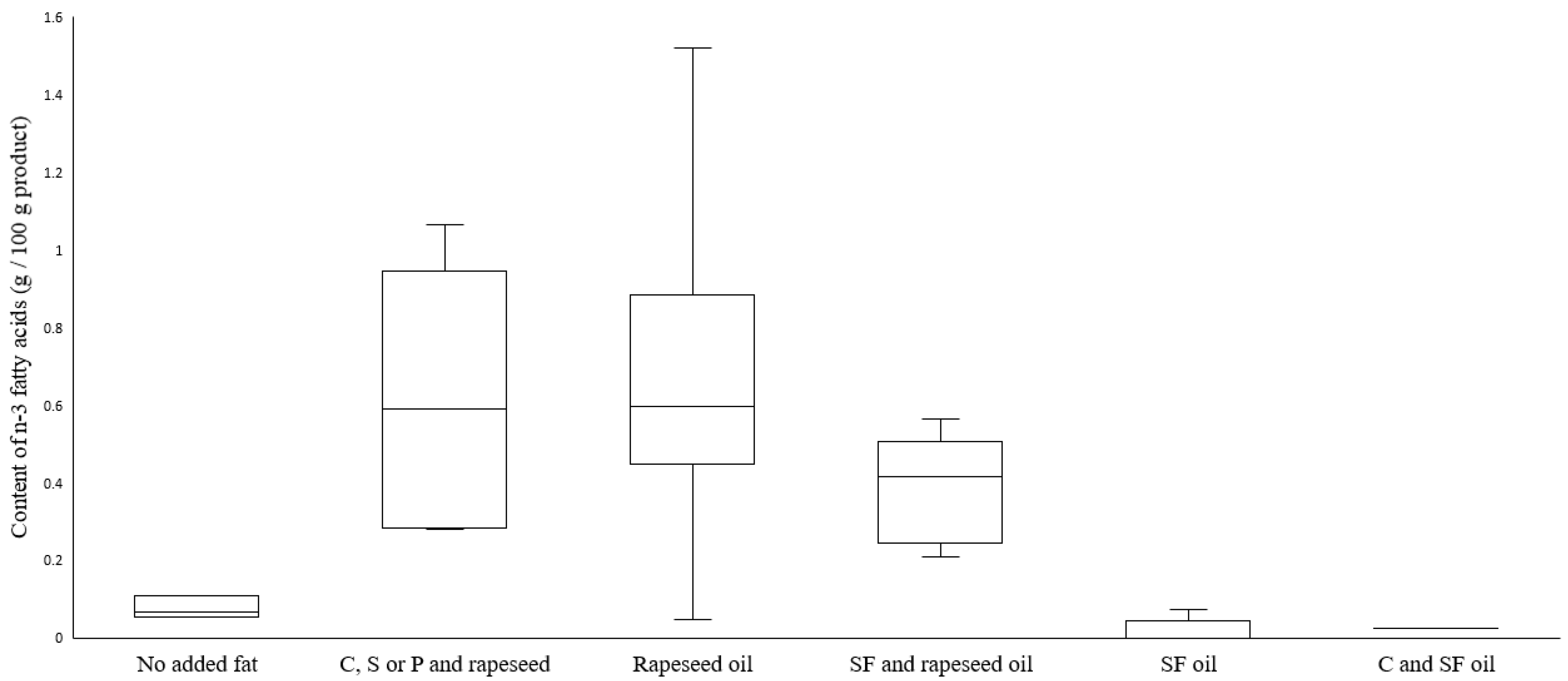
3.6. Total Fat and Fatty Acid Composition
3.7. Salt Content
3.8. Protein Content
3.9. Amino Acid Profiles
3.10. Dietary Fiber
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Clark, M.A.; Springmann, M.; Hill, J.; Tilman, D. Multiple health and environmental impacts of foods. Proc. Natl. Acad. Sci. USA 2019, 116, 23357–23362. [Google Scholar] [CrossRef]
- Springmann, M.; Godfray, H.C.J.; Rayner, M.; Scarborough, P. Analysis and valuation of the health and climate change cobenefits of dietary change. Proc. Natl. Acad. Sci. USA 2016, 113, 4146–4151. [Google Scholar] [CrossRef] [PubMed]
- Poore, J.; Nemecek, T. Reducing food’s environmental impacts through producers and consumers. Science 2018, 360, 987–992. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Sabate, R.; Sabaté, J. Consumer attitudes towards environmental concerns of meat consumption: A systematic review. Int. J. Environ. Res. Public Health 2019, 16, 1220. [Google Scholar] [CrossRef] [PubMed]
- De Koning, W.; Dean, D.; Vriesekoop, F.; Aguiar, L.K.; Anderson, M.; Mongondry, P.; Oppong-Gyamfi, M.; Urbano, B.; Luciano, C.A.G.; Jiang, B.; et al. Drivers and Inhibitors in the Acceptance of Meat Alternatives: The Case of Plant and Insect-Based Proteins. Foods 2020, 9, 1292. [Google Scholar] [CrossRef] [PubMed]
- Willett, W.; Rockström, J.; Loken, B.; Springmann, M.; Lang, T.; Vermeulen, S.; Garnett, T.; Tilman, D.; DeClerck, F.; Wood, A.; et al. Food in the Anthropocene: The EAT–Lancet Commission on healthy diets from sustainable food systems. Lancet 2019, 393, 447–492. [Google Scholar] [CrossRef]
- Li, H.; Zeng, X.; Wang, Y.; Zhang, Z.; Zhu, Y.; Li, X.; Hu, A.; Zhao, Q.; Yang, W. A prospective study of healthful and unhealthful plant-based diet and risk of overall and cause-specific mortality. Eur. J. Nutr. 2022, 61, 387–398. [Google Scholar] [CrossRef]
- Oncina-Cánovas, A.; Vioque, J.; González-Palacios, S.; Martínez-González, M.Á.; Salas-Salvadó, J.; Corella, D.; Zomeño, D.; Martínez, J.A.; Alonso-Gómez, Á.M.; Wärnberg, J.; et al. Pro-vegetarian food patterns and cardiometabolic risk in the PREDIMED-Plus study: A cross-sectional baseline analysis. Eur. J. Nutr. 2022, 61, 357–372. [Google Scholar] [CrossRef]
- Kouvari, M.; Tsiampalis, T.; Chrysohoou, C.; Georgousopoulou, E.; Skoumas, J.; Mantzoros, C.S.; Pitsavos, C.S.; Panagiotakos, D.B. Quality of plant-based diets in relation to 10-year cardiovascular disease risk: The ATTICA cohort study. Eur. J. Nutr. 2022, 61, 2639–2649. [Google Scholar] [CrossRef]
- Kim, J.; Kim, H.; Giovannucci, E.L. Plant-based diet quality and the risk of total and disease-specific mortality: A population-based prospective study. Clin. Nutr. 2021, 40, 5718–5725. [Google Scholar] [CrossRef]
- Tziva, M.; Negro, S.O.; Kalfagianni, A.; Hekkert, M.P. Understanding the protein transition: The rise of plant-based meat substitutes. Environ. Innov. Soc. Transit. 2020, 35, 217–231. [Google Scholar] [CrossRef]
- Ritchie, H.; Reay, D.S.; Higgins, P. Potential of meat substitutes for climate change mitigation and improved human health in high-income markets. Front. Sustain. Food Syst 2018, 2, 16. [Google Scholar] [CrossRef]
- Good Foods Institute. 2021 U.S. Retail Market Insights, Plant-Based Foods. 2022. Available online: https://gfi.org/wp-content/uploads/2022/03/2021-U.S.-retail-market-insights_Plant-based-foods-GFI.pdf (accessed on 17 August 2022).
- Barclays Investment Bank. Carving up the Alternative Meat Market. 2019. Available online: https://www.investmentbank.barclays.com/our-insights/carving-up-the-alternative-meat-market.html (accessed on 8 July 2022).
- Mintel Global New Products Database (GNPD). UK Meat-Free Foods Market Report 2019. Available online: https://store.mintel.com/report/uk-meat-free-foods-market-report (accessed on 12 July 2022).
- Apostolidis, C.; McLeay, F. Should we stop meating like this? Reducing meat consumption through substitution. Food Policy 2016, 65, 74–89. [Google Scholar] [CrossRef]
- Penna Franca, P.A.; Duque-Estrada, P.; da Fonseca e Sá, B.F.; van der Goot, A.J.; Pierucci, A.P.T.R. Meat substitutes—past, present, and future of products available in Brazil: Changes in the nutritional profile. Future Foods 2022, 5, 100133. [Google Scholar] [CrossRef]
- Alessandrini, R.; Brown, M.K.; Pombo-Rodrigues, S.; Bhageerutty, S.; He, F.J.; MacGregor, G.A. Nutritional Quality of Plant-Based Meat Products Available in the UK: A Cross-Sectional Survey. Nutrients 2021, 13, 4225. [Google Scholar] [CrossRef]
- Bryngelsson, S.; Moshtaghian, H.; Bianchi, M.; Hallström, E. Nutritional assessment of plant-based meat analogues on the Swedish market. Int. J. Food Sci. Nutr. 2022, 6, 1–13. [Google Scholar] [CrossRef]
- Estell, M.; Hughes, J.; Grafenauer, S. Plant Protein and Plant-Based Meat Alternatives: Consumer and Nutrition Professional Attitudes and Perceptions. Sustainability 2021, 13, 1478. [Google Scholar] [CrossRef]
- Švarc, P.L.; Jensen, M.B.; Langwagen, M.; Poulsen, A.; Trolle, E.; Jakobsen, J. Nutrient content in plant-based protein products intended for food composition databases. J. Food Compos. Anal. 2022, 106, 104332. [Google Scholar] [CrossRef]
- Sandberg, A.-S. Developing Functional Ingredient: A Case Study of Pea Protein. In Functional Foods: Concept to Product, 2nd ed.; Saarela, M., Ed.; Woodhead Publishing Ltd.: Abington, PA, USA; CRC Press LLC: Boca Raton, FL, USA, 2011; Chapter 15; pp. 358–383. [Google Scholar]
- Wang, N.; Maximiuk, L. Effect of air classification processing variables on yield, composition, and certain antinutrients of air-classified fractions from field peas by response surface methodology. J. Food Process. Preserv. 2019, 43, e13999. [Google Scholar] [CrossRef]
- Silventoinen, P.; Kortekangas, A.; Ercili-Cura, D.; Nordlund, E. Impact of ultra-fine milling and air classification on biochemical and techno-functional characteristics of wheat and rye bran. Food Res. Int. 2021, 139, 109971. [Google Scholar] [CrossRef]
- Silventoinen, P.; Rommi, K.; Holopainen-Mantila, U.; Poutanen, K.; Nordlund, E. Biochemical and Techno-Functional Properties of Protein- and Fibre-Rich Hybrid Ingredients Produced by Dry Fractionation from Rice Bran. Food Bioprocess Technol. 2019, 12, 1487–1499. [Google Scholar] [CrossRef]
- De Angelis, D.; Pasqualone, A.; Allegretta, I.; Porfido, C.; Terzano, R.; Squeo, G.; Summo, C. Antinutritional factors, mineral composition and functional properties of dry fractionated flours as influenced by the type of pulse. Heliyon 2021, 7, e06177. [Google Scholar] [CrossRef]
- Elzerman, J.E.; van Boekel, M.A.J.S.; Luning, P.A. Exploring meat substitutes: Consumer experiences and contextual factors. Br. Food J. 2013, 115, 700–710. [Google Scholar] [CrossRef]
- Fredriksson, M.; Carlsson, N.-G.; Almgren, A.; Sandberg, A.-S. Simultaneous and sensitive analysis of Cu, Ni, Zn, Co, Mn, and Fe in food and biological samples by ion chromatography. JAFC 2002, 50, 59–65. [Google Scholar] [CrossRef]
- Carlsson, N.-G.; Bergman, E.-L.; Skoglund, E.; Hasselblad, K.; Sandberg, A.-S. Rapid analyzes of inositol phosphates. J. Agric. Food Chem. 2001, 49, 1695–1701. [Google Scholar] [CrossRef]
- Siegenberg, D.; Baynes, R.D.; Bothwell, T.H.; Macfarlane, B.J.; Lamparelli, R.D.; Car, N.G.; MacPhail, P.; Schmidt, U.; Tal, A.; Mayet, F. Ascorbic acid prevents the dose-dependent inhibitory effects of polyphenols and phytates on nonheme-iron absorption. Am. J. Clin. Nutr. 1991, 53, 537–541. [Google Scholar] [CrossRef]
- Hallberg, L.; Brune, M.; Rossander, L. Iron absorption in man: Ascorbic acid and dose-dependent inhibition by phytate. Am. J. Clin. Nutr. 1989, 49, 140–144. [Google Scholar] [CrossRef]
- Fredlund, K.; Isaksson, M.; Rossander-Hulthén, L.; Almgren, A.; Sandberg, A.-S. Absorption of zinc and retention of calcium: Dose-dependent inhibition by phytate. J. Trace Elem. Med. Biol. 2006, 20, 49–57. [Google Scholar] [CrossRef]
- European Food Safety Authority. EFSA Panel on dietetic products, nutrition and allergies (NDA) Scientific Opinion on Dietary Reference Values for Zinc; EFSA Journal: 12(2014). Available online: https://efsa.onlinelibrary.wiley.com/doi/pdf/10.2903/j.efsa.2014.3844 (accessed on 17 August 2022).
- Hurrell, R.; Egli, I. Iron bioavailability and dietary reference values. Am. J. Clin. Nutr. 2010, 91, 1461S–1467S. [Google Scholar] [CrossRef]
- Dahdouh, S.; Grande, F.; Espinosa, S.N.; Vincent, A.; Gibson, R.; Bailey, K.; King, J.; Rittenschober, D.; Charrondière, U.R. Development of the FAO/INFOODS/IZINCG Global Food Composition Database for Phytate. J. Food Compos. Anal. 2019, 78, 42–48. [Google Scholar] [CrossRef]
- Howard, L.; Clark, J.; Brownmiller, C. Antioxidant capacity and phenolic content in blueberries as affected by genotype and growing season. J. Sci. Food Agric. 2003, 83, 1238–1247. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Lepage, G.; Roy, C.C. Direct transesterification of all classes of lipids in a one-step reaction. J. Lipid. Res. 1986, 27, 114–120. [Google Scholar] [CrossRef]
- Cavonius, L.R.; Carlsson, N.-G.; Undeland, I. Quantification of total fatty acids in microalgae: Comparison of extraction and transesterification methods. Anal. Bioanal. Chem. 2014, 406, 7313–7322. [Google Scholar] [CrossRef]
- Mariotti, F.; Tomé, D.; Mirand, P.P. Converting Nitrogen into Protein—Beyond 6.25 and Jones’ Factors. Crit. Rev. Food Sci. Nutr. 2008, 48, 177–184. [Google Scholar] [CrossRef]
- Özcan, S.; Şenyuva, H.Z. Improved and simplified liquid chromatography/atmospheric pressure chemical ionization mass spectrometry method for the analysis of underivatized free amino acids in various foods. J. Chromatogr. A 2006, 1135, 179–185. [Google Scholar] [CrossRef]
- Knudsen, K.E.B. Carbohydrate and lignin contents of plant materials used in animal feeding. Anim. Feed. Sci. Technol. 1997, 67, 319–338. [Google Scholar] [CrossRef]
- Dunlop, M.V.; Kilroe, S.P.; Bowtell, J.L.; Finnigan, T.J.; Salmon, D.L.; Wall, B.T. Mycoprotein represents a bioavailable and insulinotropic non-animal-derived dietary protein source: A dose–response study. Br. J. Nutr. 2017, 118, 673–685. [Google Scholar] [CrossRef]
- Derbyshire, E.; Ayoob, K.-T. Mycoprotein: Nutritional and Health Properties. Nutr. Today 2019, 54, 7–15. [Google Scholar] [CrossRef]
- Neufingerl, N.; Eilander, A. Nutrient Intake and Status in Adults Consuming Plant-Based Diets Compared to Meat-Eaters: A Systematic Review. Nutrients 2022, 14, 29. [Google Scholar] [CrossRef]
- Eklund-Jonsson, C.; Sandberg, A.-S.; Larsson Alminger, M. Reduction of phytate content while preserving minerals during whole grain cereal tempe fermentation. J. Cereal Sci. 2006, 44, 154–160. [Google Scholar] [CrossRef]
- Sutardi; Buckle, K.A. Reduction in Phytic Acid Levels in Soybeans During Tempeh Production, Storage and Frying. J. Food Sci. 1985, 50, 260–263. [Google Scholar] [CrossRef]
- Fredrikson, M.; Biot, P.; Alminger, M.L.; Carlsson, N.-G.; Sandberg, A.-S. Production Process for High-Quality Pea-Protein Isolate with Low Content of Oligosaccharides and Phytate. J. Agric. Food Chem. 2001, 49, 1208–1212. [Google Scholar] [CrossRef]
- Mayer Labba, I.-C.; Frøkiær, H.; Sandberg, A.-S. Nutritional and antinutritional composition of fava bean (Vicia faba L., var. minor) cultivars. Food Res. Int. 2021, 140, 110038. [Google Scholar] [CrossRef]
- Helstad, A.; Forsén, E.; Ahlström, C.; Mayer Labba, I.C.; Sandberg, A.S.; Rayner, M.; Purhagen, J.K. Protein extraction from cold-pressed hempseed press cake: From laboratory to pilot scale. J. Food Sci. 2022, 87, 312–325. [Google Scholar] [CrossRef]
- Deak, N.A.; Johnson, L.A. Fate of Phytic Acid in Producing Soy Protein Ingredients. J. Am. Oil Chem. Soc. 2007, 84, 369. [Google Scholar] [CrossRef]
- The European Parliament and the Council of the European Union. Regulation (EC) No 1924/2006 of the European Parliament and of the Council, on Nutrition and Health Claims Made on Foods. 2006. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX:02006R1924-20141213 (accessed on 17 August 2022).
- The European Parliament and the Council of the European Union. Regulation (EC) No 1924/2006 of the European Parliament and of the Council, on Nutrition and Health Claims Made on Foods; Article 5. 2006. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX:02006R1924-20141213 (accessed on 17 August 2022).
- Umbreit, J. Iron deficiency: A concise review. Am. J. Hematol. 2005, 78, 225–231. [Google Scholar] [CrossRef]
- Cooper, M.; Greene-Finestone, L.; Lowell, H.; Levesque, J.; Robinson, S. Iron sufficiency of Canadians. Health Rep. 2012, 23, 41–48. [Google Scholar]
- Hercberg, S.; Preziosi, P.; Galan, P. Iron deficiency in Europe. Public Health Nutr. 2001, 4, 537–545. [Google Scholar] [CrossRef]
- Lahti-Koski, M.; Valsta, L.M.; Alfthan, G.; Tapanainen, H.; Aro, A. Iron status of adults in the capital area of Finland. Eur. J. Nutr. 2003, 42, 287–292. [Google Scholar] [CrossRef]
- Modlinska, K.; Adamczyk, D.; Maison, D.; Pisula, W. Gender Differences in Attitudes to Vegans/Vegetarians and Their Food Preferences, and Their Implications for Promoting Sustainable Dietary Patterns–A Systematic Review. Sustainability 2020, 12, 6292. [Google Scholar] [CrossRef]
- Mayer Labba, I.-C.; Hoppe, M.; Gramatkovski, E.; Hjellström, M.; Abdollahi, M.; Undeland, I.; Hulthén, L.; Sandberg, A.-S. Lower Non-Heme Iron Absorption in Healthy Females from Single Meals with Texturized Fava Bean Protein Compared to Beef and Cod Protein Meals: Two Single-Blinded Randomized Trials. Nutrients 2022, 14, 3162. [Google Scholar] [CrossRef]
- Stoffel, F.; Santana, W.d.O.; Gregolon, J.G.N.; Kist, T.B.L.; Fontana, R.C.; Camassola, M. Production of edible mycoprotein using agroindustrial wastes: Influence on nutritional, chemical and biological properties. Innov. Food Sci. Emerg. Technol. 2019, 58, 102227. [Google Scholar] [CrossRef]
- Barzee, T.J.; Cao, L.; Pan, Z.; Zhang, R. Fungi for future foods. J. Future Foods 2021, 1, 25–37. [Google Scholar] [CrossRef]
- Bowman, S.M.; Free, S.J. The structure and synthesis of the fungal cell wall. Bioessays 2006, 28, 799–808. [Google Scholar] [CrossRef] [PubMed]
- Delgado-Licon, E.; Ayala, A.L.; Rocha-Guzman, N.E.; Gallegos-Infante, J.A.; Atienzo-Lazos, M.; Drzewiecki, J.; Martínez-Sánchez, C.E.; Gorinstein, S. Influence of extrusion on the bioactive compounds and the antioxidant capacity of the bean/corn mixtures. Int. J. Food Sci. Nutr. 2009, 60, 522–532. [Google Scholar] [CrossRef] [PubMed]
- El-Hady, E.-S.A.A.; Habiba, R.A.A. Effect of soaking and extrusion conditions on antinutrients and protein digestibility of legume seeds. Lwt-Food Sci. Technol. 2003, 36, 285–293. [Google Scholar] [CrossRef]
- Yağcı, S.; Göğüş, F. Effect of Incorporation of Various Food By-products on Some Nutritional Properties of Rice-based Extruded Foods. Food Sci. Technol. Int. 2009, 15, 571–581. [Google Scholar] [CrossRef]
- NNR. Nordic Nutrition Recommentations 2012; Nordic Council of Ministers: Copenhagen, Denmark, 2012; pp. 249–279. [Google Scholar]
- NNR. Nordic Nutrition Recommendations 2012; Nordic Council of Ministers: Copenhagen, Denmark, 2012; pp. 281–310. [Google Scholar]
- Boye, J.; Zare, F.; Pletch, A. Pulse proteins: Processing, characterization, functional properties and applications in food and feed. Food Res. Int. 2010, 43, 414–431. [Google Scholar] [CrossRef]
- Singh, S.; Gamlath, S.; Wakeling, L. Nutritional aspects of food extrusion: A review. Int. J. Food Sci. Technol. 2007, 42, 916–929. [Google Scholar] [CrossRef]
- Chawla, R.; Patil, G. Soluble dietary fiber. Compr. Rev. Food Sci. Food Saf. 2010, 9, 178–196. [Google Scholar] [CrossRef]
- Scheppach, W.; Luehrs, H.; Melcher, R.; Gostner, A.; Schauber, J.; Kudlich, T.; Weiler, F.; Menzel, T. Antiinflammatory and anticarcinogenic effects of dietary fibre. Clin. Nutr. Suppl. 2004, 1, 51–58. [Google Scholar] [CrossRef]
- Bosscher, D.; Van Caillie-Bertrand, M.; Van Cauwenbergh, R.; Deelstra, H. Availabilities of calcium, iron, and zinc from dairy infant formulas is affected by soluble dietary fibers and modified starch fractions. Nutrition 2003, 19, 641–645. [Google Scholar] [CrossRef]
- Debon, S.J.; Tester, R.F. In vitro binding of calcium, iron and zinc by non-starch polysaccharides. Food Chem. 2001, 73, 401–410. [Google Scholar] [CrossRef]
- Miyada, T.; Nakajima, A.; Ebihara, K. Iron bound to pectin is utilised by rats. Br. J. Nutr. 2011, 106, 73–78. [Google Scholar] [CrossRef]
- Turnlund, J.; King, J.; Keyes, W.; Gong, B.; Michel, M. A stable isotope study of zinc absorption in young men: Effects of phytate and a-cellulose. Am. J. Clin. Nutr. 1984, 40, 1071–1077. [Google Scholar] [CrossRef]
- Van den Heuvel, E.; Schaafsma, G.; Muys, T.; van Dokkum, W. Nondigestible oligosaccharides do not interfere with calcium and nonheme-iron absorption in young, healthy men. Am. J. Clin. Nutr. 1998, 67, 445–451. [Google Scholar] [CrossRef]
- Wickramasinghe, K.; Breda, J.; Berdzuli, N.; Rippin, H.; Farrand, C.; Halloran, A. The shift to plant-based diets: Are we missing the point? Glob. Food Secur. 2021, 29, 100530. [Google Scholar] [CrossRef]
- Choudhury, D.; Singh, S.; Seah, J.S.H.; Yeo, D.C.L.; Tan, L.P. Commercialization of Plant-Based Meat Alternatives. Trends Plant Sci. 2020, 25, 1055–1058. [Google Scholar] [CrossRef]
- Tso, R.; Forde, C.G. Unintended Consequences: Nutritional Impact and Potential Pitfalls of Switching from Animal- to Plant-Based Foods. Nutrients 2021, 13, 2527. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Name | Main Source of Protein | Category |
|---|---|---|
| Soy and wheat bacon | Soy and wheat protein 85% | Soy and wheat protein |
| Soy and wheat balls 1 | Soy protein 47%, Wheat protein 14% | Soy and wheat protein |
| Soy and wheat balls 2 | Soy and wheat * | Soy and wheat protein |
| Soy and wheat balls 3 | Soy and wheat protein, 43% | Soy and wheat protein |
| Soy and wheat burger | Soy protein 58.5%, Wheat protein 13% | Soy and wheat protein |
| Soy and wheat nuggets | Soy 13%, Wheat 5% | Soy and wheat protein |
| Soy and wheat sausage | Soy protein, 3.5%; Wheat protein, 10.1% | Soy and wheat protein |
| Soy and wheat schnitzel | Soy and wheat protein 58% | Soy and wheat protein |
| Chick pea falafel 1 | Chick pea 61% | Whole bean |
| Chick pea falafel 2 | Chick pea, 77% | Whole bean |
| Farm bean sausage | Farm bean 60% | Whole bean |
| White bean balls | White beans 51% | Whole bean |
| Cheese patties 1 | Cheese 17% | Cheese |
| Cheese patties 2 | Cheese * | Cheese |
| Soy balls | Soy protein, 18.5% | Soy protein |
| Soy burger | Soy protein, 21% | Soy protein |
| Soy mince 1 | Soy protein 29% | Soy protein |
| Soy mince 2 | Soy protein 23% | Soy protein |
| Soy nuggets | Soy 45% | Soy protein |
| Soy sausage 1 | Soy protein, 17% | Soy protein |
| Soy sausage 2 | Soy protein, 14.9% | Soy protein |
| Soy schnitzel 1 | Soy protein 48% | Soy protein |
| Soy schnitzel 2 | Soy protein, 15% | Soy protein |
| Pea balls 1 | Pea protein * | Pea protein |
| Pea balls 2 | Pea protein * | Pea protein |
| Pea burger 1 | Pea protein * | Pea protein |
| Pea burger 2 | Pea protein 53% | Pea protein |
| Pea mince | Pea 21% | Pea protein |
| Pea nuggets | Pea protein * | Pea protein |
| Pea patties | Pea protein 4% | Pea protein |
| Pea sausage 1 | Pea protein * | Pea protein |
| Pea sausage 2 | Pea protein * | Pea protein |
| Pea sausage 3 | Pea protein, 51% | Pea protein |
| Pea sausage 4 | Pea protein * | Pea protein |
| Pea schnitzel | Pea protein * | Pea protein |
| Mycoprotein bites | Mycoprotein 95% | Mycoprotein |
| Mycoprotein burger | Mycoprotein 40% | Mycoprotein |
| Mycoprotein Filets | Mycoprotein 85% | Mycoprotein |
| Mycoprotein mince | Mycoprotein 88% | Mycoprotein |
| Mycoprotein schnizel | Mycoprotein 38% | Mycoprotein |
| Oat, pea and bean bites | Oat protein, 17%; Pea protein, 16%, fava bean protein, 16% | Other |
| Wheat and pea nuggets | Wheat and pea protein * | Other |
| Wheat fish sticks | Wheat 33% | Other |
| Tempeh bites | Fermented peas ** | Tempeh |
| Tempeh Burger | Fermented peas ** | Tempeh |
| Group nr | Category | Median |
|---|---|---|
| 1 | Soy and wheat protein, n = 8 | 10.6 (9.6–10.2) |
| 2 | Whole bean, n = 4 | 19.3 (7.0–37.7) |
| 3 | Cheese, n = 2 | 8.2 (6.9–9.6) |
| 4 | Soy protein, n = 9 | 11.7 (9.7–14.9) |
| 5 | Pea protein, n = 12 | 10.2 (9.4–26.5) |
| 6 | Mycoprotein, n = 5 | 8.5 (7.4–9.8) |
| 7 | Other, n = 3 | 8.7 (6.7–14.0) |
| 8 | Tempeh 1, n = 1 | 21.2 ± 0.9 |
| Group Category | Source of Added Fat | Total Fat | Sig. | MUFA | Sig. | PUFA | Sig. | Essential FA 1 | Sig. |
|---|---|---|---|---|---|---|---|---|---|
| 1 | No fat (n = 3) | 2.4 (2.2–2.7) | 2, 3 **, 4, 5, 6 | 0.4 (0.31–0.42) | 2, 3, 4, 5 | 1.00 (0.93–1.32) | 2, 3, 4, 5 | 1.0 (0.93–1.32) | 2, 3, 4, 5 |
| 2 | C, S or P and rapeseed oil (n = 5) | 16.3 (10.5–17.5) | 1, 4, 5 | 5.76 (3.49–6.76) | 1, 3, 4 | 2.82 (1.53–4.58) | 1, 5 ** | 2.82 (1.52–4.58) | 1, 5 |
| 3 | Rapeseed oil (n = 22) | 12.8 (9.1–16.2) | 1 | 5.37 (4.06–7.61) | 1, 5 | 3.15 (2.23–4.07) | 1, 5 ** | 3.15 (2.23–4.07) | 1, 5 ** |
| 4 | SF and rapeseed oil (n = 6) | 10.9 (7.2–12.1) | 1, 2 | 3.99 (2.69–4.69) | 1, 2 | 2.16 (1.63–2.89) | 1 | 2.16 (1.63–2.89) | 1, 5 ** |
| 5 | SF oil (n = 7) | 13.2 (11.0–14.2) | 1, 2 | 3.15 (2.44–3.21) | 1, 2, 3 | 5.89 (5.65–9.64) | 1, 2 **, 3 ** | 5.89 (5.65–9.64) | 1, 2, 3 **, 4 ** |
| 6 | C and SF oil (n = 1) | 13.4 | 1 | 2.69 | - | 4.31 | - | 4.31 | - |
| Category | Salt Content | Protein Content |
|---|---|---|
| Soy and wheat protein, n = 8 | 1.4 (1.1–1.8) | 13.0 (11.4–5.4) |
| Whole bean, n = 4 | 1.3 (1.2–1.4) | 8.7 (7.0–11.4) |
| Cheese, n = 2 | 1.1 (1.1–1.1) | 13.3 (11.3–15.3) |
| Soy protein, n = 9 | 1.1 (1.0–1.2) | 12.5 (11.1–14.3) |
| Pea protein, n = 12 | 1.5 (1.2–2.1) | 9.5 (6.9–12.9) |
| Mycoprotein, n = 5 | 0.7 (0.3–1.0) | 13.3 (10.4–14.4) |
| Other, n = 3 | 1.5 (1.4–1.8) | 14.0 (11.9–23.8) |
| Tempeh 1, n = 1 | 1.03 ± 25% | 6.7 ± 10% |
| Soy and Wheat Protein | Soy Protein | Pea Protein | Mycoprotein | Whole Bean | Cheese | Other | Tempeh | mg/kg BW | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Essential | (n = 8) | (n = 9) | (n = 12) | (n = 5) | (n = 4) | (n = 2) | (n = 3) | (n = 1) | per day 1 | ||||||||
| Histidine | 3.8 | (3.5–4.5) | 3.6 | (3.1–3.9) | 2.8 | (2.5–3.4) | 3.0 | (2.9–3.1) | 2.4 | (2.3–2.8) | 2.6 | (2.5–2.7) | 3.7 | (3.0–3.8) | 1.7 | ± 0.1 | 10 |
| Isoleucine | 6.9 | (6.5–7.1) | 6.3 | (6.0–6.4) | 4.9 | (4.2–3.4) | 6.1 | (5.6–6.2) | 3.9 | (3.7–4.7) | 4.7 | (4.7–4.8) | 6.3 | (5.2–6.6) | 2.9 | ± 0.02 | 20 |
| Leucine | 11.9 | (11.0–12.0) | 10.3 | (9.5–10.6) | 8.0 | (6.8–9.3) | 10.2 | (8.7–10.7) | 6.2 | (5.8–8.0) | 8.4 | (8.1–8.7) | 11.8 | (9.4–12.1) | 3.9 | ± 0.01 | 39 |
| Lysine | 8.4 | (7.6–8.8) | 8.1 | (7.9–9.1) | 7.8 | (6.3–8.8) | 9.2 | (9.0–9.8) | 5.2 | (84.9–6.6) | 6.9 | (6.8–7.0) | 5.3 | (4.0–5.5) | 4.3 | ± 0.05 | 30 |
| Methionine | 1.1 | (0.9–1.8) | 1.1 | (1.0–1.6) | 1.0 | (0.8–1.3) | 2.3 | (2.0–2.4) | 1.1 | (1.0–1.3) | 1.9 | (1.7–2.1) | 1.5 | (1.3–1.5) | 1.2 | ± 0.02 | |
| Phenylalanine | 8.3 | (7.8–8.4) | 7.0 | (6.9–7.3) | 5.6 | (4.6–6.3) | 6.1 | (5.2–6.2) | 4.4 | (4.2–5.3) | 5.2 | (5.1–5.4) | 8.7 | (6.6–8.8) | 2.9 | ± 0.04 | 25 2 |
| Threonine | 6.2 | (6.0–6.4) | 5.8 | (5.2–5.9) | 4.3 | (3.7–4.9) | 6.1 | (5.9–6.3) | 3.5 | (3.2–4.5) | 4.0 | (3.9–4.1) | 4.9 | (4.3–5.3) | 2.8 | ± 0.00 | 15 |
| Valine | 7.1 | (6.6–7.4) | 6.3 | (6.0–6.3) | 5.0 | (4.5–5.8) | 6.7 | (6.6–7.0) | 4.2 | (3.9–5.2) | 5.7 | (5.6–5.9) | 6.7 | (5.4–7.0) | 3.1 | ± 0.00 | 26 |
| Non essential | |||||||||||||||||
| Alanine | 6.4 | (5.9–6.6) | 5.7 | (5.3–5.8) | 4.5 | (4.0–5.2) | 6.7 | (6.4–6.9) | 3.9 | (3.6–4.7) | 3.8 | (3.7–3.9) | 4.7 | (4.2–5.1) | 3.0 | ± 0.1 | - |
| Arginine | 8.5 | (6.8–8.9) | 8.6 | (6.4–9.4) | 6.9 | (6.1–8.3) | 6.2 | (5.7–6.8) | 5.3 | (4.9–7.0) | 3.6 | (3.4–3.7) | 6.2 | (5.5–6.7) | 2.7 | ± 0.1 | |
| Aspartic acid | 14.3 | (12.7–14.8) | 13.8 | (13.6–15.4) | 11.5 | (9.6–13.1) | 10.9 | (10.8–11.3) | 8.9 | (8.2–11.0) | 7.5 | (7.4–7.6) | 9.8 | (8.3–9.8) | 6.0 | ± 0.02 | - |
| Cysteine | 0.5 | (0.1–1.0) | 0.2 | (0.1–0.5) | 0.4 | (0.3–0.5) | 0.4 | (0.4–0.5) | 0.05 | (0.05–0.05) | 0.4 | (0.3–0.5) | - | ||||
| Glutamic acid | 34.1 | (32.2–36.2) | 25.7 | (22.3–27.1) | 16.6 | (13.9–20.0) | 15.8 | (13.8–20.5) | 12.5 | (12.4–15.7) | 18.8 | (18.0–19.6) | 48.3 | (31.9–54.1) | 7.9 | ± 0.1 | - |
| Glycine | 5.9 | (5.5–6.3) | 5.3 | (4.6–5.5) | 4.1 | (3.5–4.7) | 4.7 | (4.6–4.8) | 3.4 | (3.3–4.1) | 2.6 | (2.6–2.7) | 5.5 | (4.5–5.7) | 2.4 | ± 0.2 | - |
| Proline | 10.7 | (10.0–11.3) | 7.2 | (6.7–8.0) | 4.6 | (3.9–5.4) | 5.7 | (4.9–8.3) | 3.7 | (3.7–4.5) | 8.3 | (7.9–8.7) | 16.8 | (10.5–18.8) | 2.5 | ± 0.04 | - |
| Serine | 8.2 | (7.7–8.7) | 7.1 | (6.5–7.4) | 5.4 | (4.8–6.2) | 6.8 | (6.0–6.8) | 4.6 | (4.4–5.8) | 5.7 | (5.6–5.8) | 8.5 | (6.6–8.7) | 3.1 | ± 0.06 | - |
| Tyrosine | 6.0 | (5.8–6.3) | 5.4 | (5.3–5.7) | 4.4 | (4.0–4.9) | 5.0 | (4.4–5.1) | 3.7 | (3.6–4.3) | 4.8 | (4.7–4.9) | 6.1 | (4.7–6.2) | 2.4 | ± 0.00 | - |
| Meat Substitutes Based on: | Total DF | Soluble DF | Insoluble DF |
|---|---|---|---|
| Soy and wheat protein extract, n = 8 | 10.9 (8.0–13.9) | 3.3 (4.5–4.9) | 7.7 (5.9–10.0) |
| Whole bean, n = 4 | 11.4 (9.2–13.2) | 3.6 (2.1–5.1) | 6.8 (6.6–10.0) |
| Soy protein extract, n = 9 | 10.5 (9.6–13.3) | 4.3 (3.3–4.9) | 6.7 (6.0–8.2) |
| Pea protein extract, n = 12 | 8.0 (7.1–9.7) | 2.7 (2.1–3.3) | 5.3 (4.3–6.6) |
| Mycoprotein, n = 5 | 9.9 (6.8–17.8) | 1.3 (1.0–3.4) | 7.4 (5.8–16) |
| Tempeh, n = 1 | 21.5 | 6.1 | 15.4 |
| Cheese, n = 2 | 6.7 (6.2–7.3) | 2.2 (1.8–2.5) | 4.6 (4.4–4.8) |
| Other, n = 3 | 5.6 (4.4–8.8) | 1.4 (0.5–4.3) | 4.1 (3.9–4.5) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mayer Labba, I.-C.; Steinhausen, H.; Almius, L.; Bach Knudsen, K.E.; Sandberg, A.-S. Nutritional Composition and Estimated Iron and Zinc Bioavailability of Meat Substitutes Available on the Swedish Market. Nutrients 2022, 14, 3903. https://doi.org/10.3390/nu14193903
Mayer Labba I-C, Steinhausen H, Almius L, Bach Knudsen KE, Sandberg A-S. Nutritional Composition and Estimated Iron and Zinc Bioavailability of Meat Substitutes Available on the Swedish Market. Nutrients. 2022; 14(19):3903. https://doi.org/10.3390/nu14193903
Chicago/Turabian StyleMayer Labba, Inger-Cecilia, Hannah Steinhausen, Linnéa Almius, Knud Erik Bach Knudsen, and Ann-Sofie Sandberg. 2022. "Nutritional Composition and Estimated Iron and Zinc Bioavailability of Meat Substitutes Available on the Swedish Market" Nutrients 14, no. 19: 3903. https://doi.org/10.3390/nu14193903
APA StyleMayer Labba, I.-C., Steinhausen, H., Almius, L., Bach Knudsen, K. E., & Sandberg, A.-S. (2022). Nutritional Composition and Estimated Iron and Zinc Bioavailability of Meat Substitutes Available on the Swedish Market. Nutrients, 14(19), 3903. https://doi.org/10.3390/nu14193903