
Oudemansiella raphanipies Polysaccharides Improve Lipid Metabolism Disorders in Murine High-Fat Diet-Induced Non-Alcoholic Fatty Liver Disease

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Purification of Polysaccharides
2.2. Characterization of Monosaccharides
2.3. Fourier Transform-Infrared (FT-IR) Spectroscopy
2.4. Determination of Molecular Weight (Mw)
2.5. Development of NAFLD Mouse Model
2.6. Biochemical Assays
2.7. Liver Histopathological Examination
2.8. Metabolite Extraction
2.9. Statistical Analysis
3. Results and Discussion
3.1. Primary Structure of ORPS-1
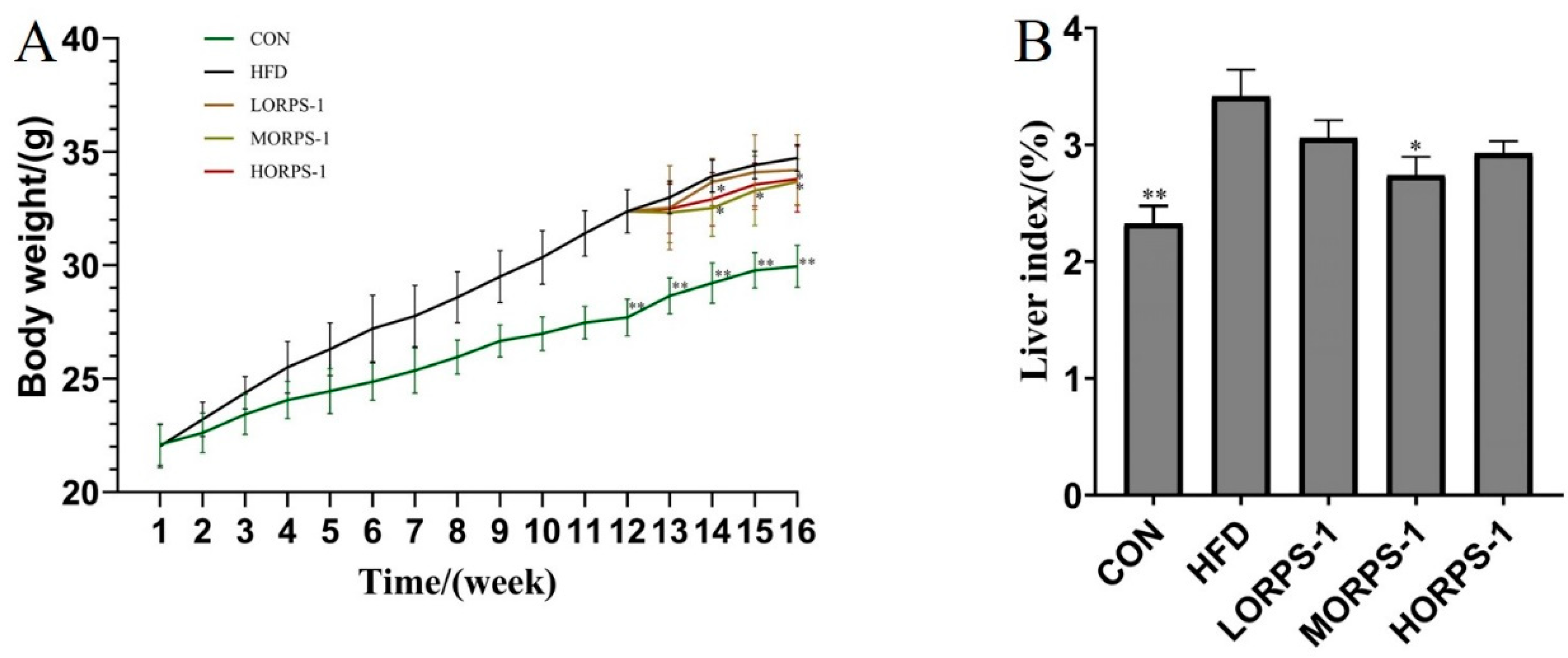
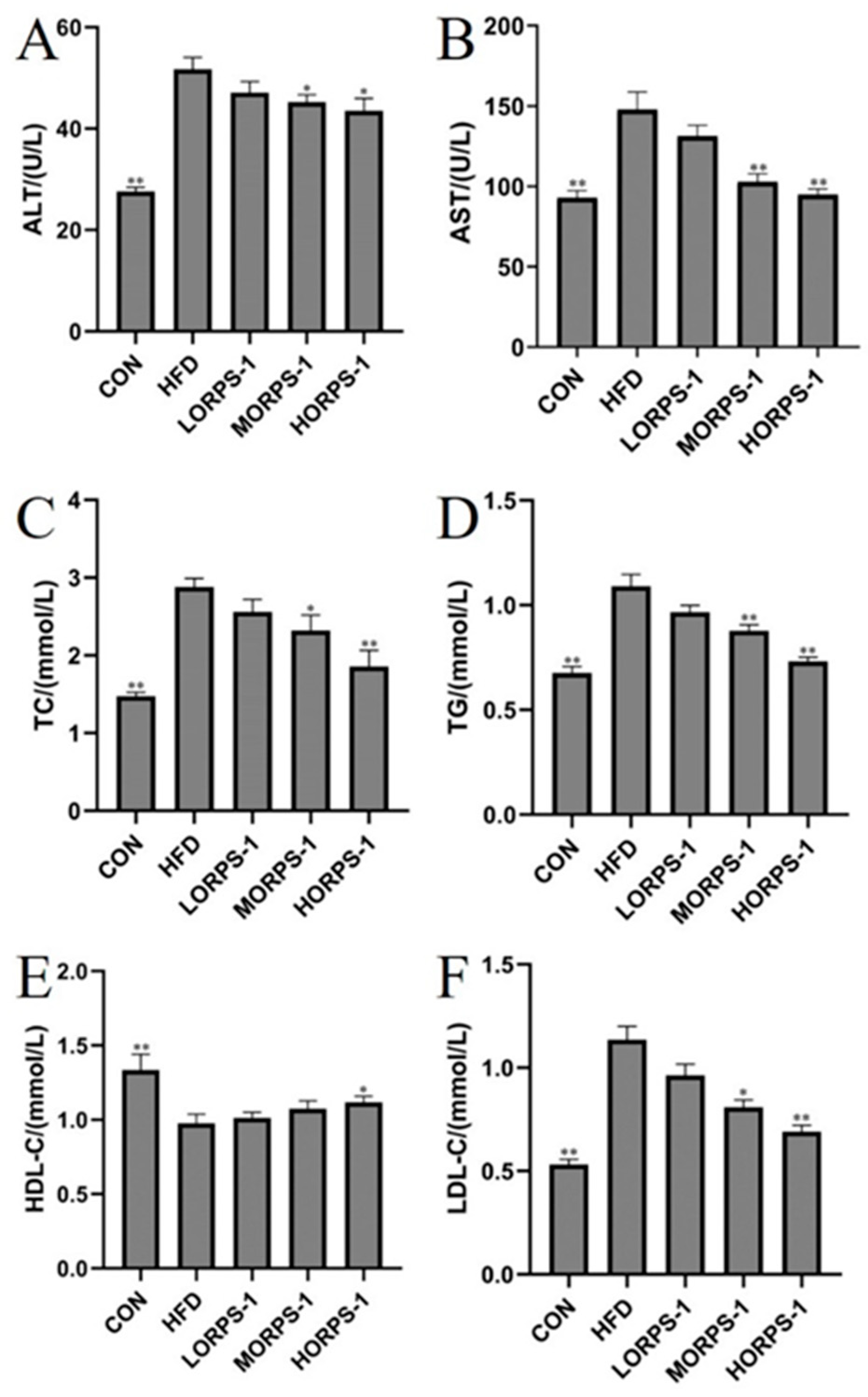
3.2. ORPS-1 Improved Liver Function in HFD Mice
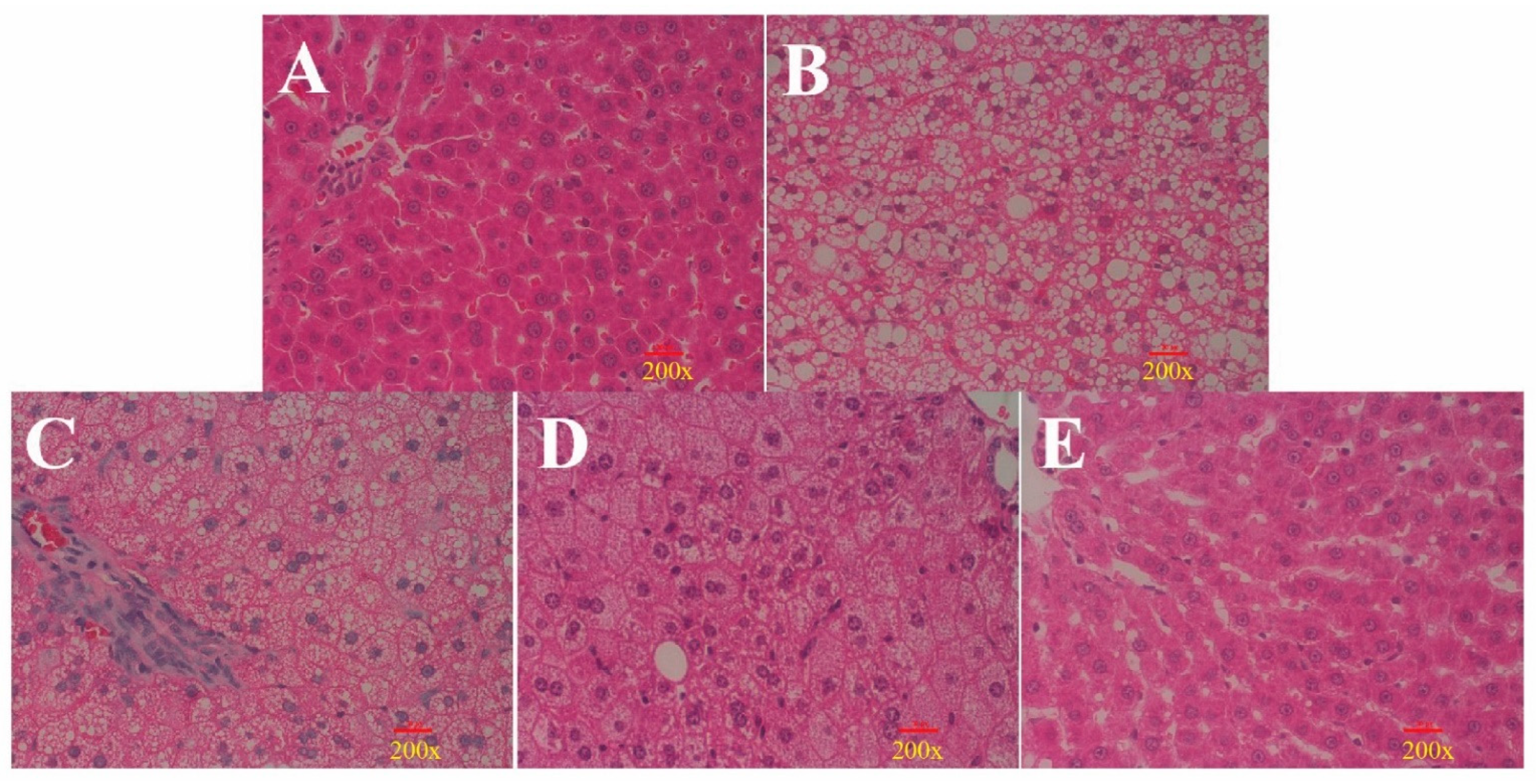
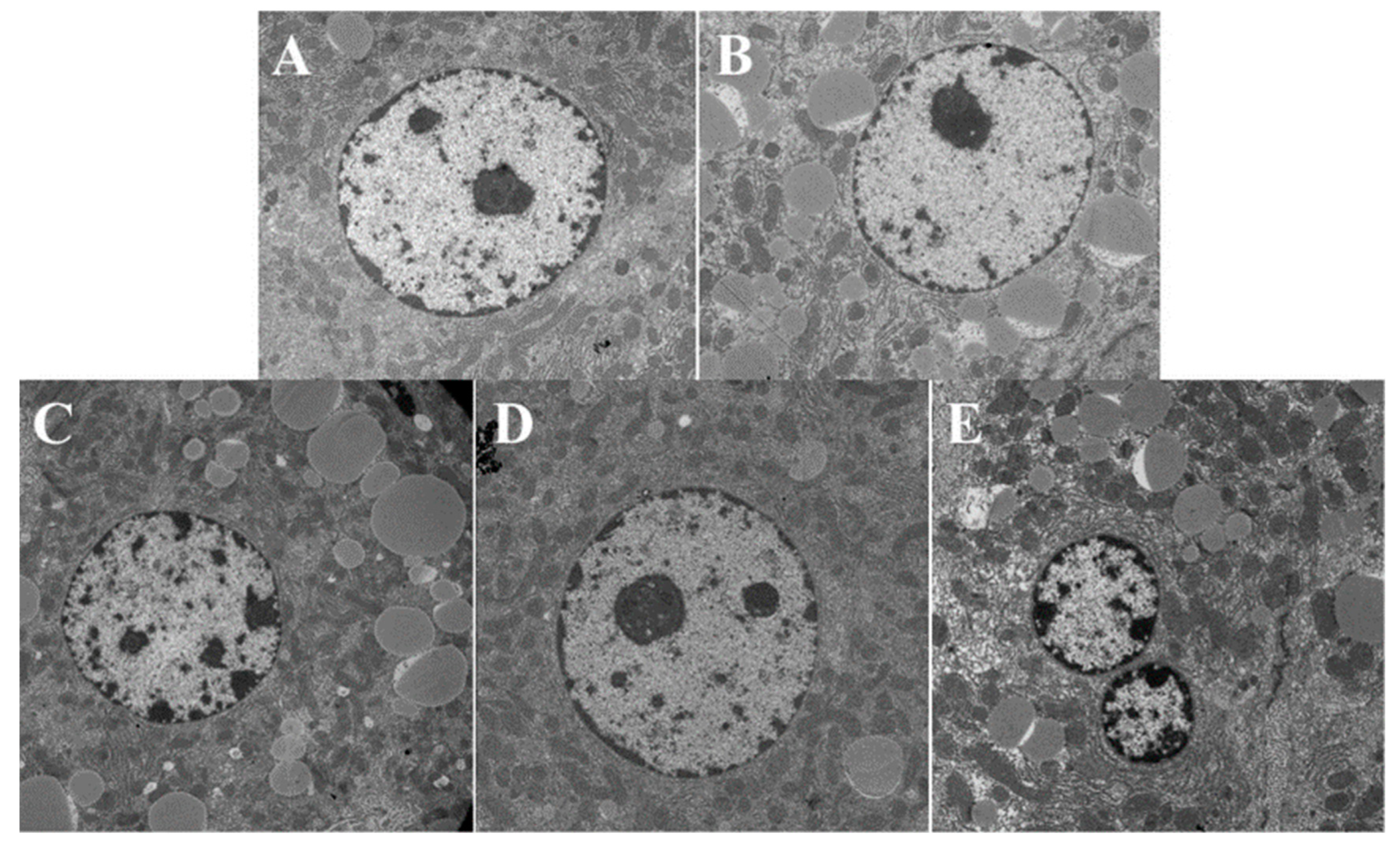
3.3. Histological Analysis Shows That ORPS-1 Treatment Ameliorates Liver Injury
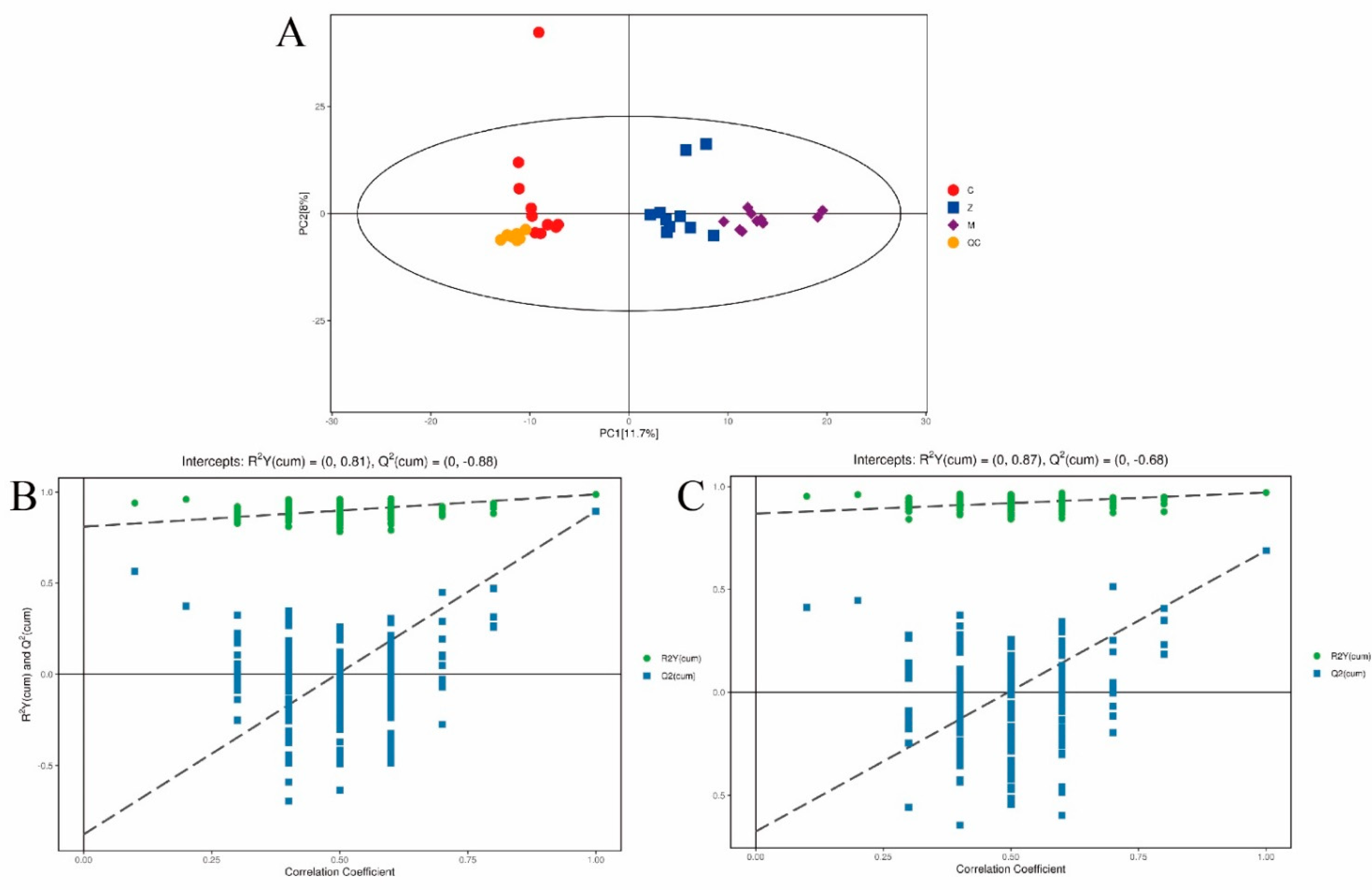
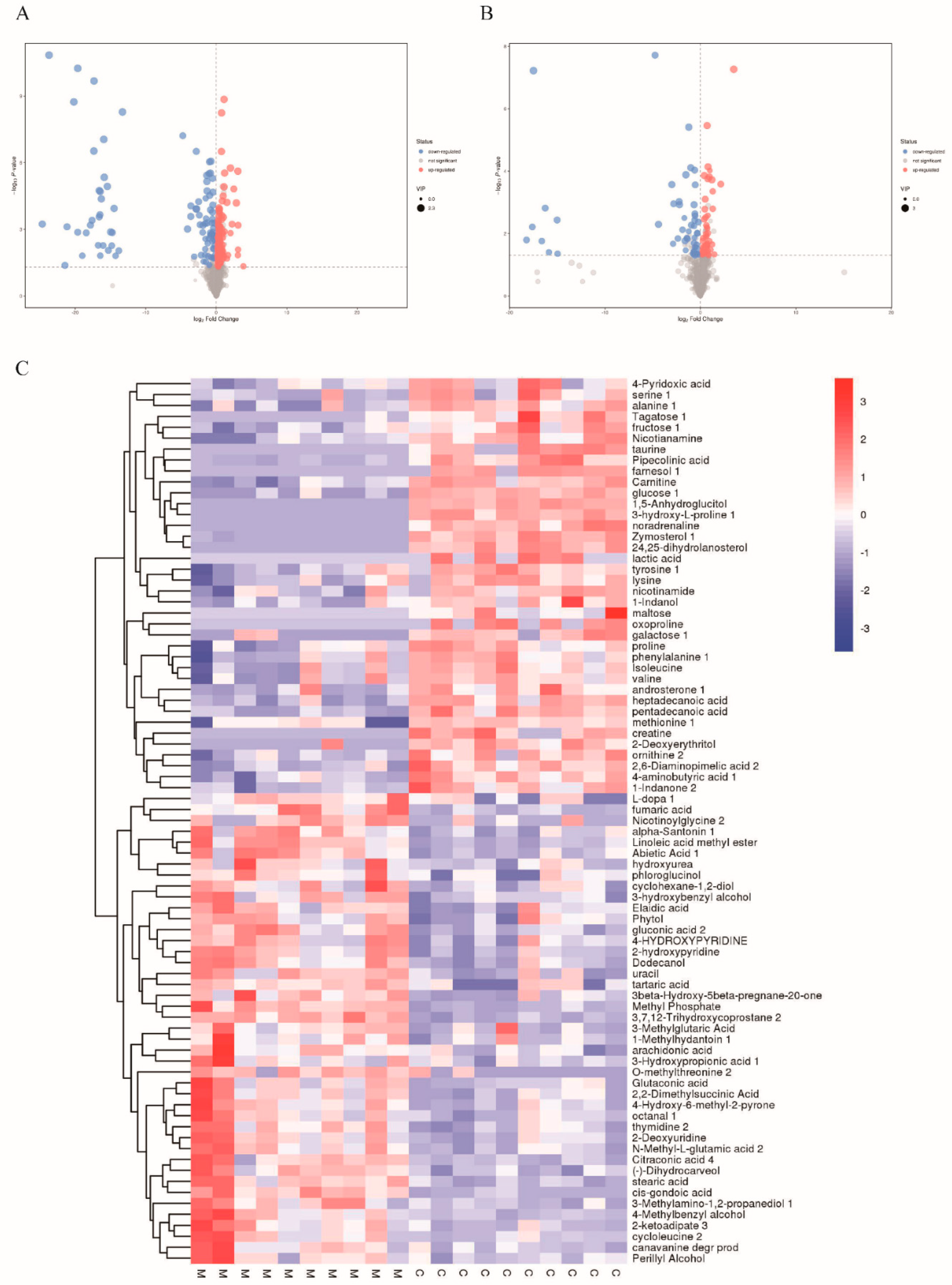
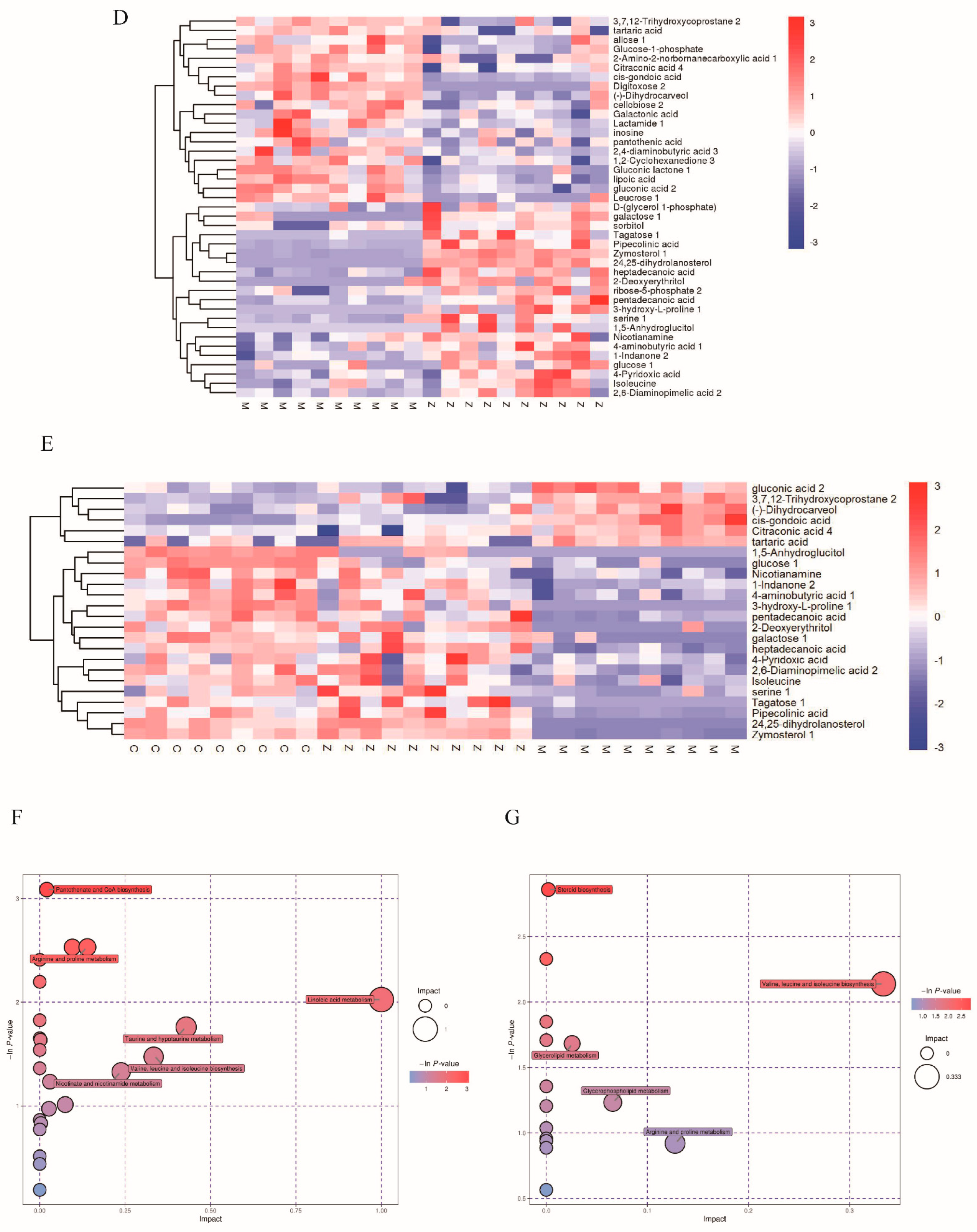
3.4. Metabolic Profiling of Mice Liver Tissue
3.5. Analysis of Total Metabolites
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, X.; Wu, M.; Liu, Z.; Yuan, H.; Wu, X.; Shi, T.; Chen, X.; Zhang, T. Increasing prevalence of NAFLD/NASH among children, adolescents and young adults from 1990 to 2017: A population-based observational study. BMJ 2021, 11, e042843. [Google Scholar] [CrossRef] [PubMed]
- Abdul, G.R.; Muhammad, S.; Mobeen, Z.M.; Shagufta, J.; Muhammad, S.N.S.; Kaukab, R.G. Identification of metabolic risk phenotypes predisposing to Non-Alcoholic Fatty Liver Disease in a Pakistani Cohort. Pak. J. Med. Sci. 2017, 33, 121–126. [Google Scholar] [CrossRef]
- Temple, J.L.; Cordero, P.; Li, J.; Nguyen, V.; Oben, J. A guide to non-alcoholic fatty liver disease in childhood and adolescence. Int. J. Mol. Sci. 2016, 17, 947. [Google Scholar] [CrossRef] [PubMed]
- Radaelli, M.G.; Martucci, F.; Perra, S.; Accornero, S.; Castoldi, G.; Lattuada, G.; Manzoni, G.; Perseghin, G. NAFLD/NASH in patients with type 2 diabetes and related treatment options. J. Endocrinol. Invest. 2018, 41, 509–521. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Liu, J.; Yan, C.; Zhang, C.; Pan, W.; Zhang, W.; Lu, Y.; Chen, L.; Chen, Y. Sarcodon aspratus polysaccharides ameliorated obesity-induced metabolic disorders and modulated gut microbiota dysbiosis in mice fed a high-fat diet. Food Funct. 2020, 11, 2588–2602. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.; Yang, M.; Yang, Y.; Yu, C.; Wang, C. Antrodia cinnamomea prevents obesity, dyslipidemia, and the derived fatty liver via regulating AMPK and SREBP signaling. Am. J. Chin. Med. 2017, 45, 67–83. [Google Scholar] [CrossRef]
- Li, Y.; Lu, X.; Li, X.; Guo, X.; Sheng, Y.; Li, Y.; Xu, G.; Han, X.; An, L.; Du, P. Effects of Agaricus blazei Murrill polysaccharides on hyperlipidemic rats by regulation of intestinal microflora. Food. Sci. Nutr. 2020, 8, 2758–2772. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Duan, X.; Duan, S.; Li, C.; Hu, B.; Liu, A.; Wu, Y.; Wu, H.; Chen, H.; Wu, W. Effects of in vitro digestion and fecal fermentation on the stability and metabolic behavior of polysaccharides from Craterellus cornucopioides. Food Funct. 2020, 11, 6899–6910. [Google Scholar] [CrossRef]
- Liu, Y.; Li, Y.; Zhang, H.; Li, C.; Zhang, Z.; Liu, A.; Chen, H.; Hu, B.; Luo, Q.; Lin, B.; et al. Polysaccharides from Cordyceps miltaris cultured at different pH: Sugar composition and antioxidant activity. Int. J. Biol. Macromol. 2020, 162, 349–358. [Google Scholar] [CrossRef]
- Xu, X.; Yang, J.; Ning, Z.; Zhang, X. Lentinula edodes-derived polysaccharide rejuvenates mice in terms of immune responses and gut microbiota. Food Funct. 2015, 6, 2653–2663. [Google Scholar] [CrossRef]
- Kerezoudi, E.N.; Mitsou, E.K.; Gioti, K.; Terzi, E.; Avgousti, I.; Panagiotou, A.; Koutrotsios, G.; Zervakis, G.I.; Mountzouris, K.C.; Tenta, R.; et al. Fermentation of Pleurotus ostreatus and Ganoderma lucidum mushrooms and their extracts by the gut microbiota of healthy and osteopenic women: Potential prebiotic effect and impact of mushroom fermentation products on human osteoblasts. Food Funct. 2021, 12, 1529–1546. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Qin, C.; Shu, X.; Chen, R.; Song, H.; Li, Q.; Xu, H. Effects of copper on induction of thiol-compounds and antioxidant enzymes by the fruiting body of Oudemansiella radicata. Ecotox. Environ. Safe 2015, 111, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yi, C.; Han, J.; Ming, T.; Zhou, J.; Lu, C.; Li, Y.; Su, X. Novel high-docosahexaenoic-acid tuna oil supplementation modulates gut microbiota and alleviates obesity in high-fat diet mice. Food Sci. Nutr. 2020, 8, 6513–6527. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Juan, L.; Song, X.; Zhang, J.; Wang, X.; Jing, H.; Ren, Z.; Li, S.; Zhang, C.; Jia, L. Antioxidative, anti-inflammation and lung-protective effects of mycelia selenium polysaccharides from Oudemansiella radicata. Inter. J Bio. Macromol. 2017, 104, 1158–1164. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, Y.; Ke, Y.; Li, C.; Zhang, Z.; Liu, A.; Luo, Q.; Lin, B.; He, J.; Wu, W. Processing of four different cooking methods of Oudemansiella radicata: Effects on in vitro bioaccessibility of nutrients and antioxidant activity. Food Chem. 2021, 337, 128007. [Google Scholar] [CrossRef]
- Liu, Q.; Zhu, M.; Geng, X.; Wang, H.; Tzi, B.N. Characterization of polysaccharides with antioxidant and hepatoprotective activities from the edible mushroom Oudemansiella radicata. Molecules 2017, 22, 234. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Jing, S.; Jiang, R.; Wang, C.; Zhang, C.; Chen, J.; Li, H. Metabolomics study of the therapeutic mechanism of Schisandra chinensis lignans on aging rats induced by D-galactose. Clin. Interv. in Aging. 2018, 13, 829–841. [Google Scholar] [CrossRef] [Green Version]
- Zheng, G.; Chen, Z.; Chengye, T.; Zhenzhen, R.; Xinling, S.; Xiuxiu, W.; Nuo, X.; Huijuan, J.; Shangshang, L.; Weiru, L. Characterization, antioxidation, anti-inflammation and renoprotection effects of selenized mycelia polysaccharides from Oudemansiella radicata. Carbohyd. Poly. 2018, 181, 1224–1234. [Google Scholar]
- Jahn, D.; Kircher, S.; Hermanns, H.M.; Geier, A. Animal models of NAFLD from a hepatologist’s point of view. BBA.-Mol. Basis. Dis. 2019, 1865, 943–953. [Google Scholar] [CrossRef]
- Yu, K.; Huang, K.; Jiang, S.; Tang, X.; Huang, X.; Sun, L.; Pang, L.; Mo, C. Protective function on liver and proteomic analysis of the improvement mechanism of Sedum sarmentosum Bunge extract on nonalcoholic fatty liver disease in Nile tilapia. Aquaculture 2021, 531, 735977. [Google Scholar] [CrossRef]
- Dunn, W.B.; Broadhurst, D.; Begley, P.; Zelena, E.; Mcintyre, S.F.; Anderson, N.; Brown, M.; D, K.J.; Halsall, A.; N, H.J. Procedures for large-scale metabolic profiling of serum and plasma using gas chromatography and liquid chromatography coupled to mass spectrometry. Nat. Protoc. 2018, 6, 1060–1083. [Google Scholar] [CrossRef] [PubMed]
- Maity, P.; Sen, I.K.; Chakraborty, I.; Mondal, S.; Bar, H.; Bhanja, S.K.; Mandal, S.; Maity, G.N. Biologically active polysaccharide from edible mushrooms: A review. Int. J. Biol. Macromol. 2021, 172, 408–417. [Google Scholar] [CrossRef] [PubMed]
- Maryama, S.; Akrama, A.; Alia, M.S.; Sarab, S.p.-N.-T. Physicochemical properties and surface activity characterization of water-soluble polysaccharide isolated from Balangu seed (Lallemantia royleana) gum. J. Food. Meas. Charact. 2020, 14, 3625–3632. [Google Scholar]
- Zhang, C.; Yu, Y.; Liang, Y.; Chen, X. Purification, partial characterization and antioxidant activity of polysaccharides from Glycyrrhiza uralensis. Int. J. Bio. Macromol. 2015, 79, 681–686. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Shen, Q.; Liu, M.; Chen, Z.; Zhang, L.; Ren, Z.; Wang, W.; Dong, Y.; Wang, X.; Zhang, J. Antioxidant and hepatoprotective effects of intracellular mycelium polysaccharides from Pleurotus geesteranus against alcoholic liver diseases. Inter. J. Bio. Macromol. 2018, 114, 979–988. [Google Scholar] [CrossRef]
- Zhao, H.; Li, H.; Lai, Q.; Yang, Q.; Dong, Y.; Liu, X.; Wang, W.; Zhang, J.; Jia, L. Antioxidant and hepatoprotective activities of modified polysaccharides from Coprinus comatus in mice with alcohol-induced liver injury. Int. J. Biolog. Macromol. 2019, 127, 476–485. [Google Scholar] [CrossRef]
- Zhang, L.; Ren, X.; He, J.; Li, W.-N.; Hao, J.-W.; Jing, Y.-R.; Chai, J.-R.; Sun, X.-W.; Wan, C.-L. Protective effect of Phaeoporus obliquus polysaccharide against acute liver injury induced by carbon tetrachloride and alcohol in mice. Pak. J. Pharm. Sci. 2021, 34, 649–656. [Google Scholar]
- GiaBuu, T.; SaoMai, D.; Tram, N.T. Amelioration of single clove black garlic aqueous extract on dyslipidemia and hepatitis in chronic carbon tetrachloride intoxicated swiss albino mice. Inter. J. Hepatol. 2018, 2018, 1–9. [Google Scholar]
- Tan, G.; Lou, Z.; Liao, W.; Zhu, Z.; Dong, X.; Zhang, W.; Li, W.; Chai, Y. Potential biomarkers in mouse myocardium of doxorubicin-induced cardiomyopathy: A metabonomic method and its application. PLoS ONE 2011, 6, e27683. [Google Scholar] [CrossRef] [Green Version]
- Ajaz, S.; McPhail, M.J.; Gnudi, L.; SalmaMujib, F.M.T.; Napoli, S.; Carey, I.; Agarwal, K. Mitochondrial dysfunction as a mechanistic biomarker in patients with Non-Alcoholic fatty liver disease (NAFLD). Mitochondrion 2020, 57, 119–130. [Google Scholar] [CrossRef]
- ElenaRomero-Ibarguengoitia, M.; Vadillo-Ortega, F.; Caballero, A.; Ibarra-Gonzalez, I.; Herrera-Rosas, A.; Maria, F.-C.; Mireya, L.-H.; Antonio, G.-C.; Mummidi, S.; Duggirala, R.; et al. Family history and obesity in youth, their effect on acylcarnitine/aminoacids metabolomics and non-alcoholic fatty liver disease (NAFLD). Structural equation modeling approach. PLoS ONE 2018, 13, e0193138. [Google Scholar]
- Gaggini, M.; Carli, F.; Rosso, C.; Buzzigoli, E.; Marietti, M.; DellaLatta, V.; Ciociaro, D.; Abate, M.; Gambino, R.; Cassader, M.; et al. Altered amino acid concentrations in NAFLD: Impact of obesity and insulin resistance. Hepatology 2018, 67, 145–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- K, H.M. Mitigating factors and metabolic mechanisms in fructose-induced nonalcoholic fatty liver disease: The next challenge. Am. J. Clin. Nutr. 2012, 96, 951–952. [Google Scholar]
- Caviglia, G.P.; Rosso, C.; Armandi, A.; Gaggini, M.; Bugianesi, E. Interplay between oxidative stress and metabolic derangements in Non-Alcoholic Fatty Liver Disease: The role of Selenoprotein P. Int. J. Mol. Sci. 2020, 21, 8838. [Google Scholar]
- Cui, H.; Li, Y.; Cao, M.; Liao, J.; Liu, X.; Miao, J.; Fu, H.; Song, R.; Wen, W.; Zhang, Z.; et al. Untargeted metabolomic analysis of the effects and mechanism of nuciferine treatment on rats with nonalcoholic fatty liver disease. Front. Pharmacol. 2020, 11, 8538. [Google Scholar] [CrossRef]
- Isabel, A.; Su, Y.; Roger, G.-J. Suppression of endogenous glucose production by isoleucine and valine and impact of diet composition. Nutrients 2016, 8, 79–86. [Google Scholar]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, H.; Zhu, H.; Huo, G.; Li, S.; Wu, Y.; Zhou, F.; Hua, C.; Hu, Q. Oudemansiella raphanipies Polysaccharides Improve Lipid Metabolism Disorders in Murine High-Fat Diet-Induced Non-Alcoholic Fatty Liver Disease. Nutrients 2022, 14, 4092. https://doi.org/10.3390/nu14194092
Jiang H, Zhu H, Huo G, Li S, Wu Y, Zhou F, Hua C, Hu Q. Oudemansiella raphanipies Polysaccharides Improve Lipid Metabolism Disorders in Murine High-Fat Diet-Induced Non-Alcoholic Fatty Liver Disease. Nutrients. 2022; 14(19):4092. https://doi.org/10.3390/nu14194092
Chicago/Turabian StyleJiang, Haitao, Hua Zhu, Guangming Huo, Shengjie Li, Yulong Wu, Feng Zhou, Chun Hua, and Qiuhui Hu. 2022. "Oudemansiella raphanipies Polysaccharides Improve Lipid Metabolism Disorders in Murine High-Fat Diet-Induced Non-Alcoholic Fatty Liver Disease" Nutrients 14, no. 19: 4092. https://doi.org/10.3390/nu14194092