

Phenolic-Compound-Rich Opuntia littoralis Ethyl Acetate Extract Relaxes Arthritic Symptoms in Collagen-Induced Mice Model via Bone Morphogenic Markers

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Chemicals and Reagents
2.3. Preparation of OL Cladode Extract
2.4. Phytochemical Screening
2.4.1. Determination of Total Phenolic Acids
2.4.2. Determinations of Total Flavonoid Content (TFC)
2.4.3. Investigation of Polyphenolic Compounds Using HPLC
2.5. Antioxidant Assays of OL Extract
2.5.1. DPPH Assay
2.5.2. FRAP Assay
2.6. Anti-Inflammatory Activities of OL Extracts
2.7. Acute Oral Toxicity Research
2.8. Preparation of Arthritis Induction Solution
2.9. Induction of Arthritis in Mouse Model
2.10. Calculation of the Arthritic Score
2.11. Histopathological Assessment
2.12. Immunohistochemistry
2.13. Malondialdehyde (MDA) and Nitric Oxide (NO) Quantification
2.14. Cytokine Estimation
2.15. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
2.16. Western Blot
2.17. Statistical Analysis
3. Results
3.1. Phytochemical Analysis
3.2. Estimation of Phytochemicals of Opuntia littoralis Cladode Extracts
3.3. Analysis of Polyphenolic Compounds in EAE and HAE of OL Cladodes Using HPLC
3.4. Effect of OL Extracts on Antioxidant Activity
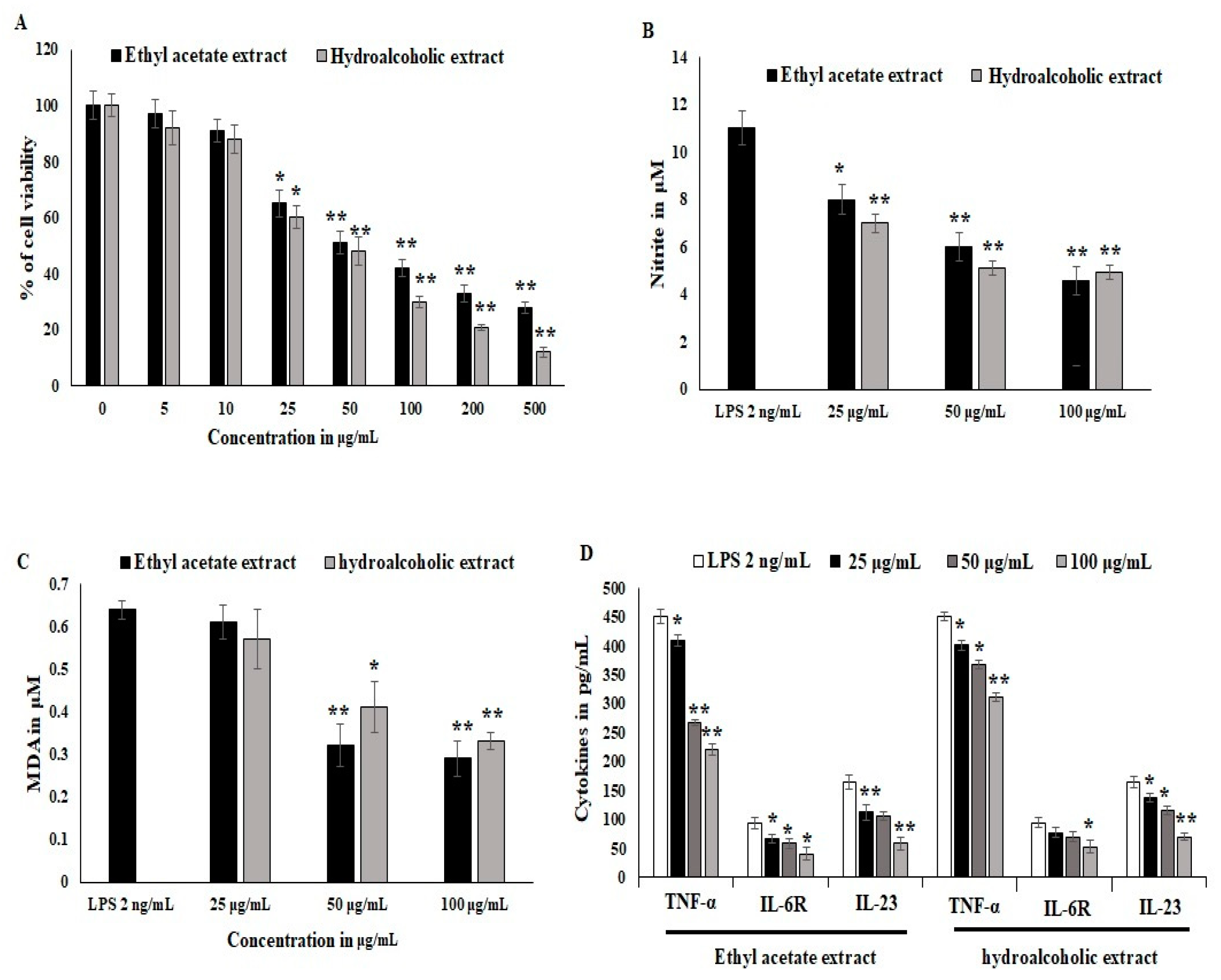
3.5. Effect of Ethyl Acetate and Hydro Alcoholic Extract on Inflammatory Cells
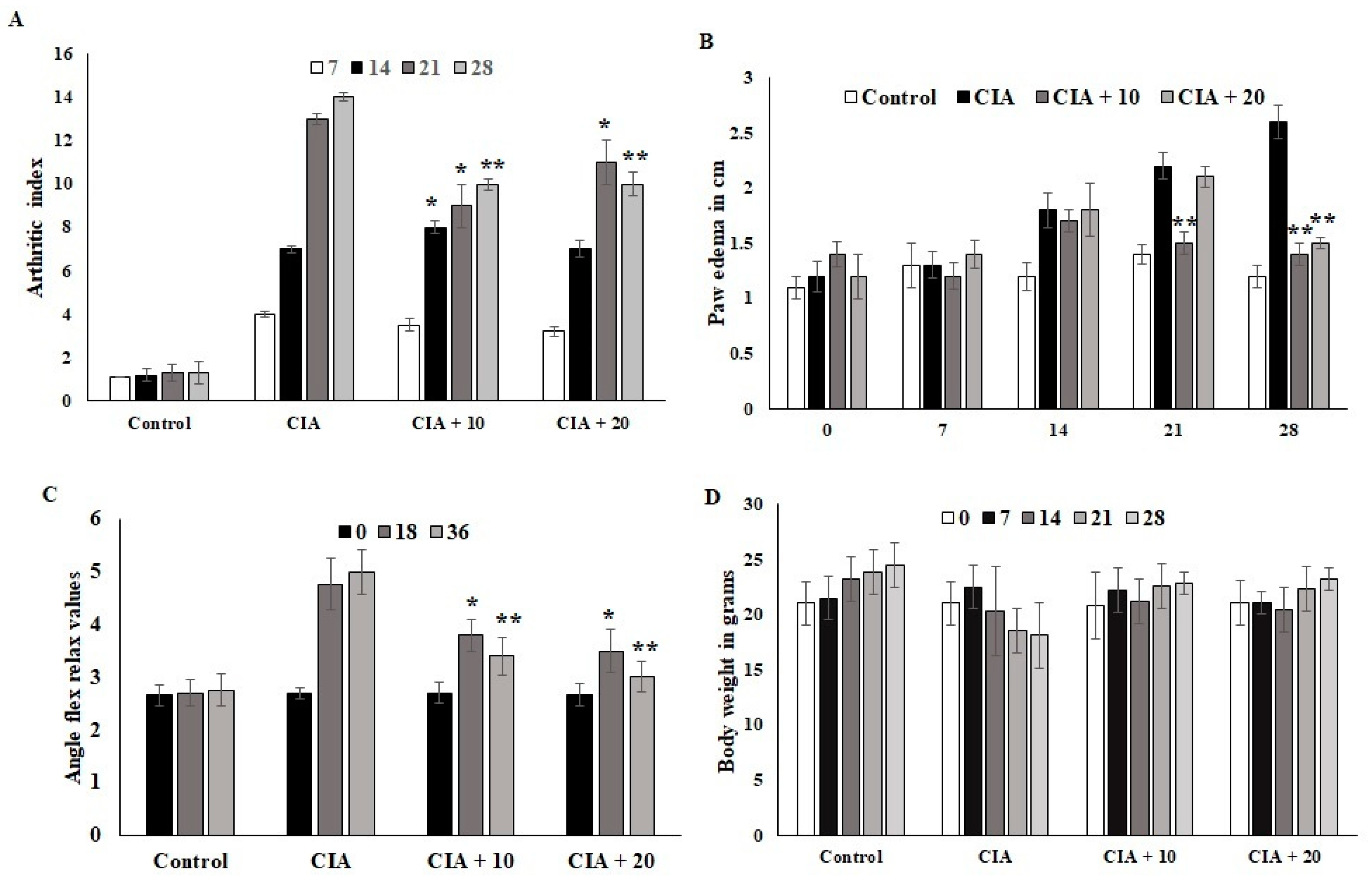
3.6. Effect of OL-EAE on Collagen-Induced Arthritic (CIA) Mice
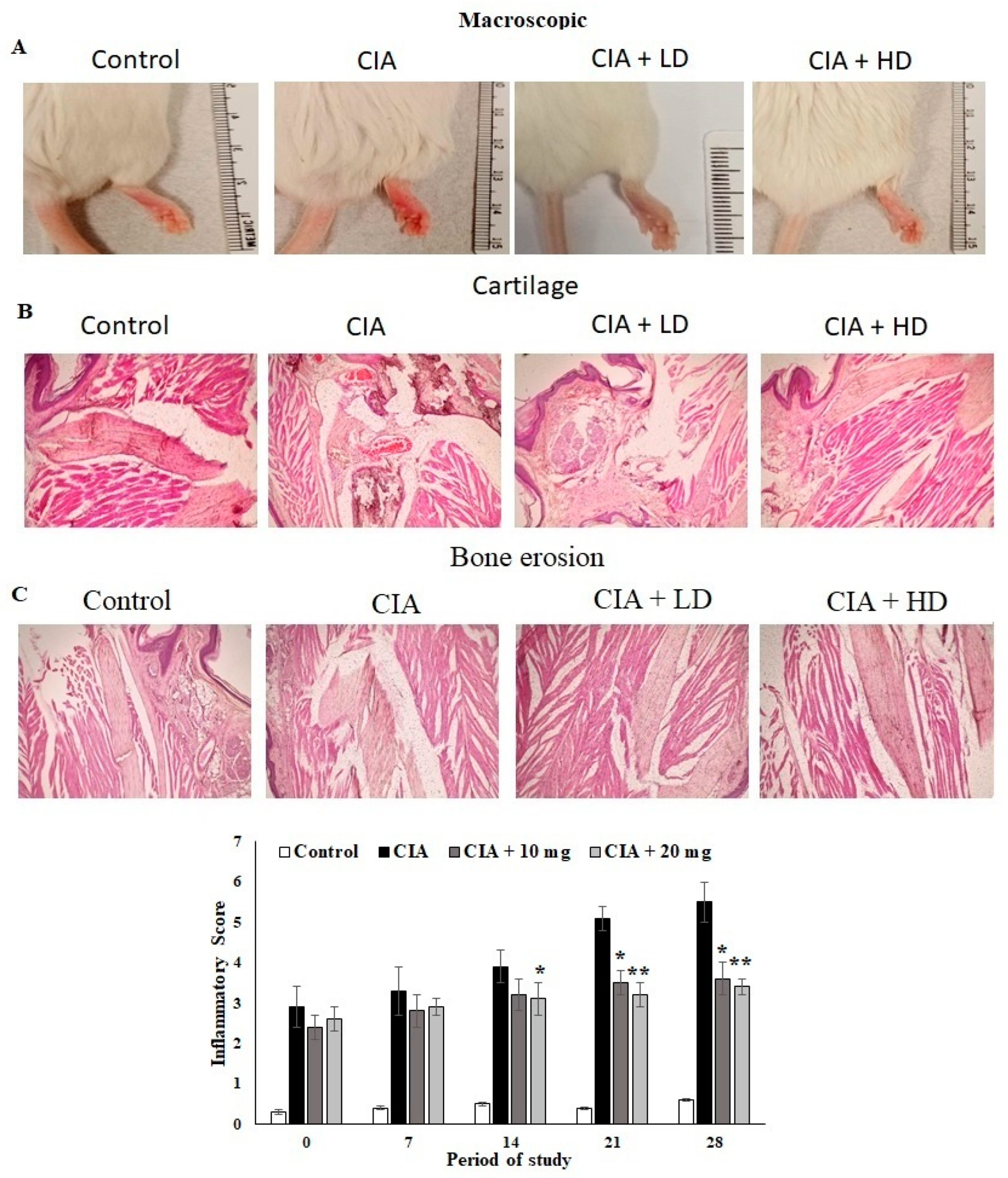
3.7. Effect of OL-EAE on Macroscopic and Microscopic Assessment of CIA Mice
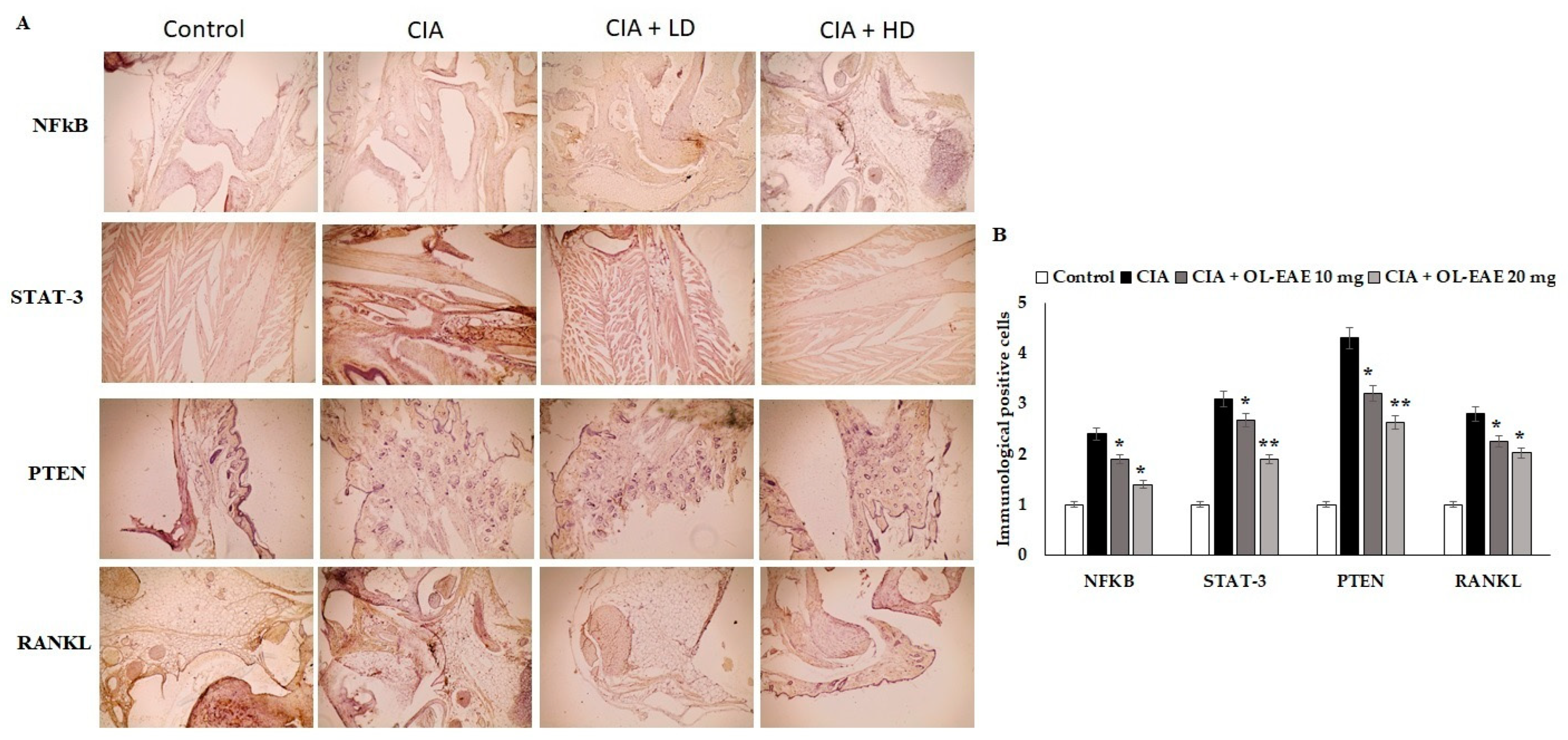
3.8. Effect of OL-EAE on Marker Localization in Arthritic Mice
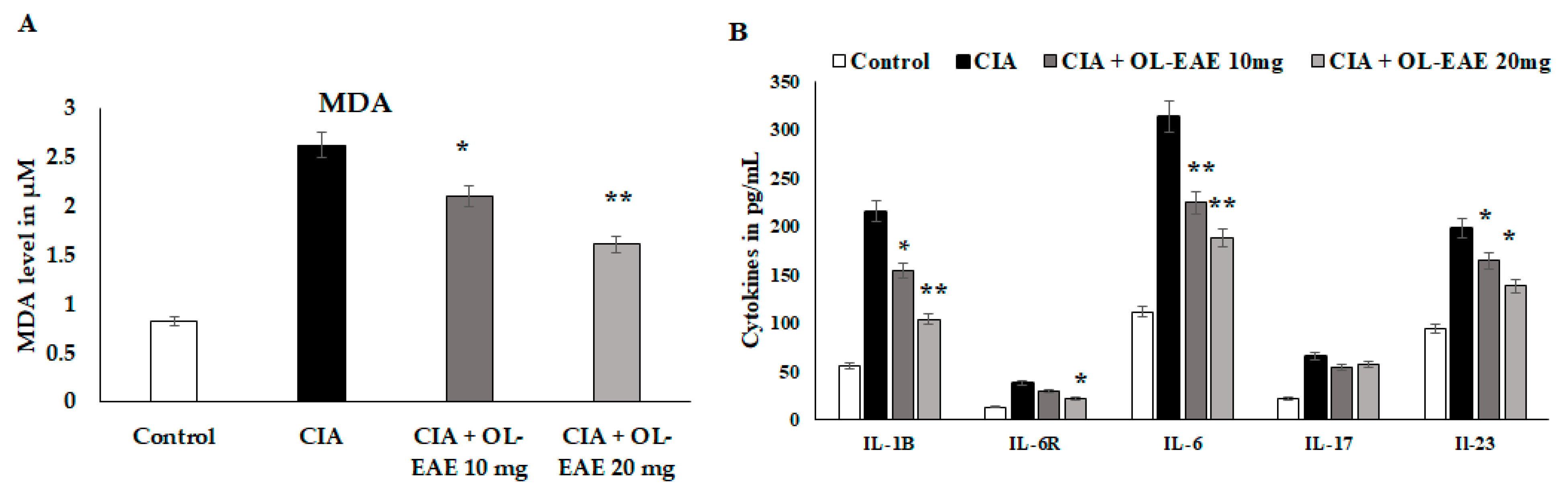
3.9. Regulation of Secondary Inflammatory Mediators in OL-EAE-Treated CIA Mice
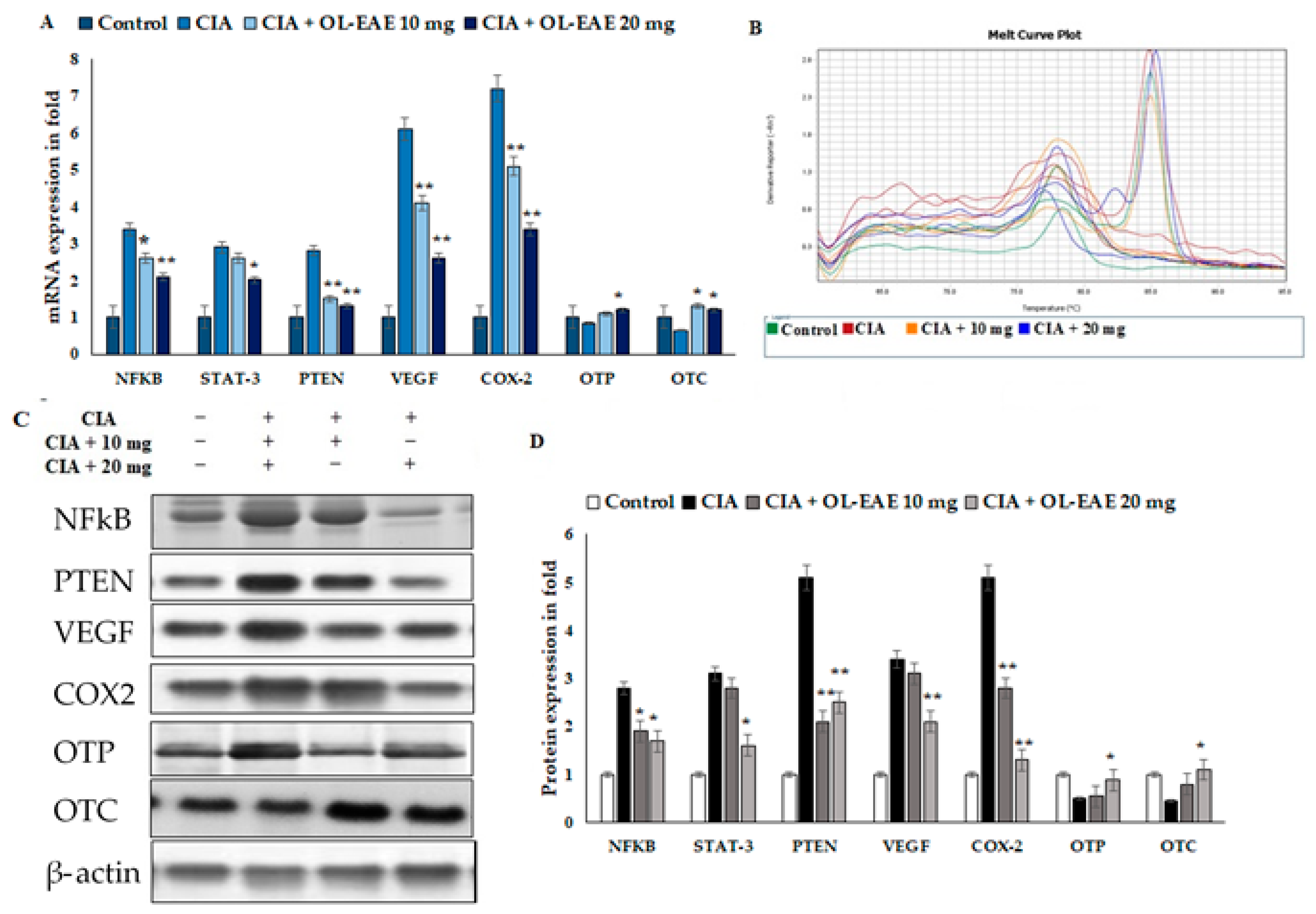
3.10. Effect of OL-EAE on Bone Formation and Inflammatory Markers in CIA Mice
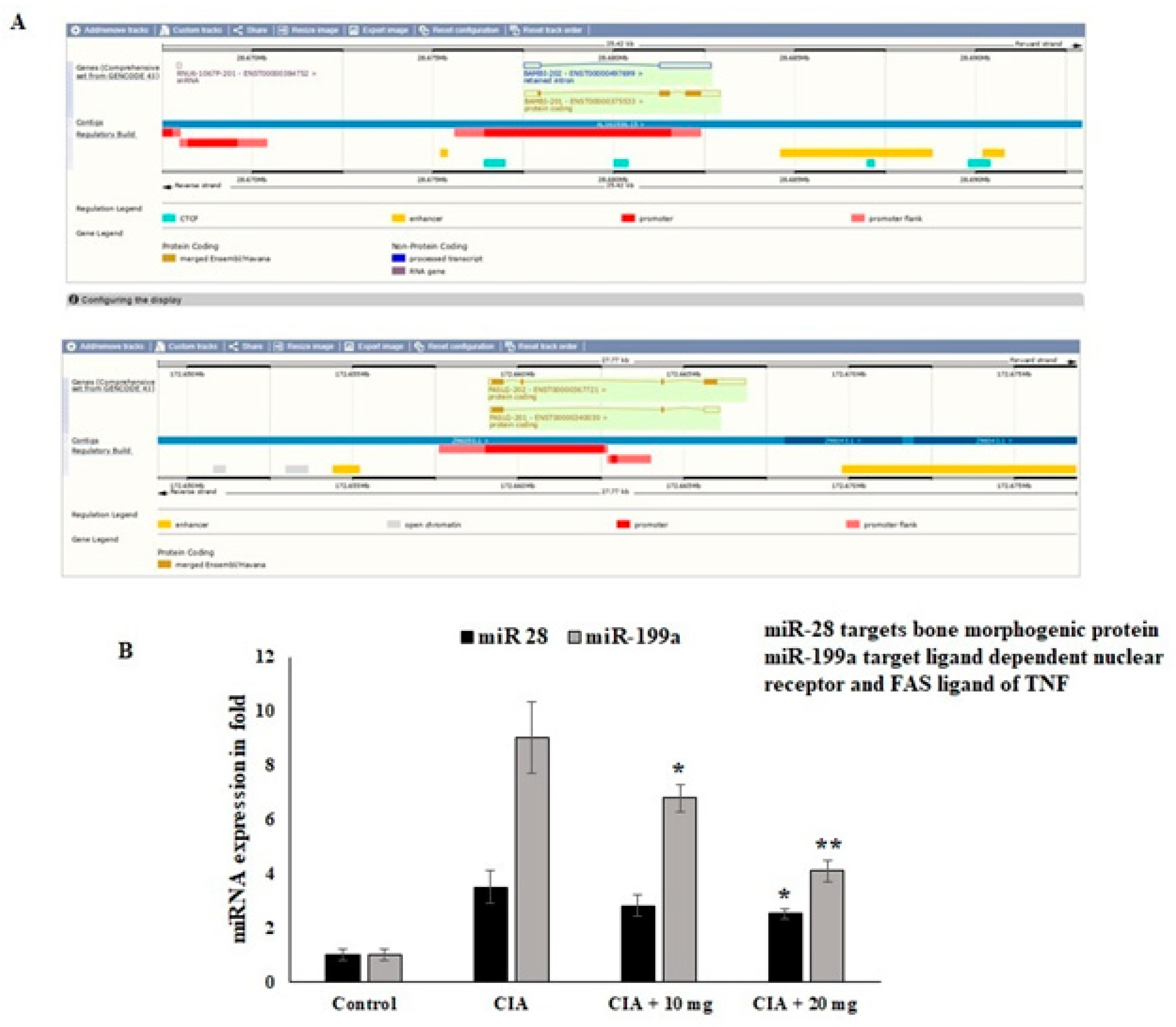
3.11. Effect of OL-EAE Extracts on Epigenetic Regulators (miR28 and miR-199a) of mRNA Targets
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Anjali, P.; Vimalavathini, R.; Kavimani, S. In Vitro Anti-Inflammatory and Anti-Arthritic Activity of Ethanolic Extract Of Leaves Of Pyrenacantha Volubilis (Eepv). In Vitro 2020, 13, 156–158. [Google Scholar] [CrossRef]
- Barrow, M. An Overview of the NF-kB mechanism of pathophysiology in rheumatoid arthritis, investigation of the NF-kB ligand RANKL and related nutritional interventions. Autoimmun. Rev. 2021, 20, 102741. [Google Scholar] [CrossRef]
- Alunno, A.; Carubbi, F.; Giacomelli, R.; Gerli, R. Cytokines in the pathogenesis of rheumatoid arthritis: New players and therapeutic targets. BMC Rheumatol. 2017, 1, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.; Jiang, S.; Dong, Q.; Dang, J.; Liu, Z.; Han, H.; Tao, Y.; Yue, H. Anti-rheumatoid arthritis effects of iridoid glucosides from Lamiophlomis rotata (Benth.) kudo on adjuvant-induced arthritis in rats by OPG/RANKL/NF-κB signaling pathways. J. Ethnopharmacol. 2021, 266, 113402. [Google Scholar] [CrossRef] [PubMed]
- Min, H.K.; Kim, S.M.; Baek, S.Y.; Woo, J.W.; Park, J.S.; Cho, M.L.; Lee, J.; Kwok, S.K.; Kim, S.W.; Park, S.H. Anthocyanin extracted from black soybean seed coats prevents autoimmune arthritis by suppressing the development of Th17 cells, and synthesis of proinflammatory cytokines by such cells via inhibition of NF-κB. PLoS ONE 2015, 10, e0138201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruscitti, P.; Cipriani, P.; Carubbi, F.; Liakouli, V.; Zazzeroni, F.; Di Benedetto, P.; Berardicurti, O.; Alesse, E.; Giacomelli, R. The role of IL-1β in the bone loss during rheumatic diseases. Mediat. Inflamm. 2015, 2015, 782382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volkov, M.; van Schie, K.A.; van der Woude, D. Autoantibodies and B Cells: The ABC of rheumatoid arthritis pathophysiology. Immunol. Rev. 2020, 294, 148–163. [Google Scholar] [CrossRef] [Green Version]
- Feng, J.H.; Jung, J.S.; Hwang, S.H.; Lee, S.K.; Lee, S.Y.; Kwak, Y.G.; Kim, D.H.; Song, C.Y.; Kim, M.J.; Lim, S.S.; et al. The mixture of Agrimonia pilosa Ledeb. and Salvia miltiorrhiza Bunge. extract produces analgesic and anti-inflammatory effects in a collagen-induced arthritis mouse model. Anim. Cells Syst. 2022, 26, 166–173. [Google Scholar] [CrossRef]
- Yang, X.; Chang, Y.; Wei, W. Emerging role of targeting macrophages in rheumatoid arthritis: Focus on polarization, metabolism and apoptosis. Cell Prolif. 2020, 53, e12854. [Google Scholar] [CrossRef]
- Chen, Z.; Bozec, A.; Ramming, A.; Schett, G. Anti-inflammatory and immune-regulatory cytokines in rheumatoid arthritis. Nat. Rev. Rheumatol. 2019, 15, 9–17. [Google Scholar] [CrossRef]
- Luan, J.; Hu, Z.; Cheng, J.; Zhang, R.; Yang, P.; Guo, H.; Nan, G.; Guo, N.; Gou, X. Applicability and implementation of the collagen induced arthritis mouse model, including protocols. Exp. Ther. Med. 2021, 22, 939. [Google Scholar] [CrossRef]
- Evangelatos, G.; Fragoulis, G.E.; Koulouri, V.; Lambrou, G.I. MicroRNAs in rheumatoid arthritis: From pathogenesis to clinical impact. Autoimmun. Rev. 2019, 18, 102391. [Google Scholar] [CrossRef] [PubMed]
- Bardin, P.; Marchal-Duval, E.; Sonneville, F.; Blouquit-Laye, S.; Rousselet, N.; Le Rouzic, P.; Corvol, H.; Tabary, O. Small RNA and transcriptome sequencing reveal the role of miR-199a-3p in inflammatory processes in cystic fibrosis airways. J. Pathol. 2018, 245, 410–420. [Google Scholar] [CrossRef]
- Wangyang, Y.; Yi, L.; Wang, T.; Feng, Y.; Liu, G.; Li, D.; Zheng, X. MiR-199a-3p inhibits proliferation and induces apoptosis in rheumatoid arthritis fibroblast-like synoviocytes via suppressing retinoblastoma 1. Biosci. Rep. 2018, 38, BSR20180982. [Google Scholar] [CrossRef] [Green Version]
- Tatangelo, M.; Tomlinson, G.; Paterson, J.M.; Ahluwalia, V.; Kopp, A.; Gomes, T.; Bansback, N.; Bombardier, C. Association of patient, prescriber, and region with the initiation of first prescription of biologic disease-modifying antirheumatic drug among older patients with rheumatoid arthritis and identical health insurance coverage. JAMA Netw. Open 2019, 2, e1917053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.F.; Wu, S.; Yan, Q.; Wu, Y.Y.; Chen, H.; Yin, S.Q.; Chen, X.; Wang, H.; Li, J. PTEN Methylation Promotes Inflammation and Activation of Fibroblast-Like Synoviocytes in Rheumatoid Arthritis. Front. Pharmacol. 2021, 12, 1747. [Google Scholar] [CrossRef]
- Harirforoosh, S.; Asghar, W.; Jamali, F. Adverse effects of nonsteroidal antiinflammatory drugs: An update of gastrointestinal, cardiovascular and renal complications. J. Pharm. Pharm. Sci. 2013, 16, 821–847. [Google Scholar] [CrossRef] [Green Version]
- Iriti, M.; Faoro, F. Chemical diversity and defence metabolism: How plants cope with pathogens and ozone pollution. Int. J. Mol. Sci. 2009, 10, 3371–3399. [Google Scholar] [CrossRef] [Green Version]
- Jannat, A.; John, P.; Bhatti, A.; Hayat, M.Q. Tomorou attenuates progression of rheumatoid arthritis through alteration in ULK-1 independent autophagy pathway in collagen induced arthritis mice model. Cell Death Discov. 2019, 5, 142. [Google Scholar] [CrossRef] [Green Version]
- Kang, E.J.; Kim, H.J.; Choi, J.H.; Noh, J.R.; Kim, J.H.; Lee, I.B.; Choi, Y.K.; Choi, D.H.; An, J.; Lee, C.H.; et al. Humulus japonicus extract ameliorates collagen-induced arthritis in mice through regulation of overall articular inflammation. Int. J. Mol. Med. 2020, 45, 417–428. [Google Scholar] [CrossRef]
- Kim, I.; Kim, H.; Lee, E.H.; Jo, G.; Na, C.S.; Kang, K.; Lee, T.H. Anti-Inflammatory Effect of Cudrania tricuspidata Extract and Stewartia koreana Extract Mixture in a Collagen-Induced Arthritis Mouse Model. Appl. Sci. 2021, 11, 6660. [Google Scholar] [CrossRef]
- El-Mostafa, K.; El Kharrassi, Y.; Badreddine, A.; Andreoletti, P.; Vamecq, J.; El Kebbaj, M.S.; Latruffe, N.; Lizard, G.; Nasser, B.; Cherkaoui-Malki, M. Nopal cactus (Opuntia ficus-indica) as a source of bioactive compounds for nutrition, health and disease. Molecules 2014, 19, 14879–14901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bargougui, A.; Tag, H.M.; Bouaziz, M.; Triki, S. Antimicrobial, antioxidant, total phenols and flavonoids content of four cactus (Opuntia ficus-indica) cultivars. Biomed. Pharmacol. J. 2019, 12, 1353–1368. [Google Scholar] [CrossRef]
- Zeghbib, W.; Boudjouan, F.; Vasconcelos, V.; Lopes, G. Phenolic Compounds’ Occurrence in Opuntia Species and Their Role in the Inflammatory Process: A Review. Molecules 2022, 27, 4763. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Moaty, H.I.; Gouda, H.M.; Sorour, W.A.; Metwally, A.; Hassan, L.M.; Youssef, A.K.; Alsheddi, T.H.S.; Galal, T.M. Nutritional composition of the edible wild plant Opuntia littoralis Englem. in relation to different seasons and edaphic factors. Appl. Ecol. Environ. Res. 2022, 20, 3655–3669. [Google Scholar] [CrossRef]
- Abd El-Moaty, H.I.; Sorour, W.A.; Youssef, A.K.; Gouda, H.M. Structural elucidation of phenolic compounds isolated from Opuntia littoralis and their antidiabetic, antimicrobial and cytotoxic activity. S. Afr. J. Bot. 2020, 131, 320–327. [Google Scholar] [CrossRef]
- Müller, J.; Heindl, A. Drying of medicinal plants. Frontis 2006, 17, 237–252. [Google Scholar]
- Lala, P.K. Lab Manuals of Pharmacognosy; CSI Publishers and Distributors: Calcutta, India, 1993; Volume 5, pp. 38–48. [Google Scholar]
- Ezeonu, C.S.; Ejikeme, C.M. Qualitative and quantitative determination of phytochemical contents of indigenous Nigerian softwoods. New J. Sci. 2016, 2016, 5601327. [Google Scholar] [CrossRef] [Green Version]
- Annapandian, V.M.; Rajagopal, S.S. Phytochemical evaluation and in vitro antioxidant activity of various solvent extracts of Leucas aspera (Willd.) Link leaves. Free Radic. Antioxid. 2017, 7, 166–171. [Google Scholar] [CrossRef] [Green Version]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Zou, Y.; Lu, Y.; Wei, D. Antioxidant activity of a flavonoid-rich extract of Hypericum perforatum L. in vitro. J. Agric. Food Chem. 2004, 52, 5032–5039. [Google Scholar] [CrossRef] [PubMed]
- Ben-Hammouda, M.; Kremer, R.J.; Minor, H.C.; Sarwar, M. A chemical basis for differential allelopathic potential of sorghum hybrids on wheat. J. Chem. Ecol. 1995, 21, 775–786. [Google Scholar] [CrossRef] [PubMed]
- Loganayaki, N.; Siddhuraju, P.; Manian, S. Antioxidant activity and free radical scavenging capacity of phenolic extracts from Helicteres isora L. and Ceiba pentandra L. J. Food Sci. Technol. 2013, 50, 687–695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, F.; Ariga, T.; Yoshimura, Y.; Nakazawa, H. Antioxidative and anti-glycation activity of garcinol from Garcinia indica fruit rind. J. Agric. Food Chem. 2000, 48, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, E.A.; Ibrahim, H.M.; Khalil, H.E. Pinocembrin reduces arthritic symptoms in mouse model via targeting Sox4 signaling molecules. J. Med. Food 2021, 24, 282–291. [Google Scholar] [CrossRef]
- Huang, D.; Jiang, S.; Du, Z.; Chen, Y.; Xue, D.; Wang, X.; Li, M.; Zhang, F.; Chen, W.; Sun, L. Analgesic and Anti-Arthritic Activities of Polysaccharides in Chaenomeles speciosa. Front. Pharmacol. 2022, 13, 744915. [Google Scholar] [CrossRef]
- Ibrahim, H.I.M.; AlZahrani, A.; Hanieh, H.; Ahmed, E.A.; Thirugnanasambantham, K. MicroRNA-7188-5p and miR-7235 regulates Multiple sclerosis in an experimental mouse model. Mol. Immunol. 2021, 139, 157–167. [Google Scholar] [CrossRef]
- Khalifa, A.; Sheikh, A.; Ibrahim, H.I.M. Bacillus amyloliquefaciens Enriched Camel Milk Attenuated Colitis Symptoms in Mice Model. Nutrients 2022, 14, 1967. [Google Scholar] [CrossRef]
- Khalil, H.E.; Ibrahim, H.-I.M.; Ahmed, E.A.; Emeka, P.M.; Alhaider, I.A. Orientin, a Bio-Flavonoid from Trigonella hamosa L.; Regulates COX-2/PGE-2 in A549 Cell Lines via miR-26b and miR-146a. Pharmaceuticals 2022, 15, 154. [Google Scholar] [CrossRef]
- Sharma, B.R.; Park, C.M.; Choi, J.W.; Rhyu, D.Y. Anti-nociceptive and anti-inflammatory effects of the methanolic extract of Opuntia humifusa stem. Avicenna J. Phytomed. 2017, 7, 366. [Google Scholar]
- Cha, M.N.; Jun, H.I.; Lee, W.J.; Kim, M.J.; Kim, M.K.; Kim, Y.S. Chemical composition and antioxidant activity of Korean cactus (Opuntia humifusa) fruit. Food Sci. Biotechnol. 2013, 22, 523–529. [Google Scholar] [CrossRef]
- Mabry, T.; Markham, K.R.; Thomas, M.B. The Systematic Identification of Flavonoids; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Bagdas, D.; Gul, Z.; Meade, J.A.; Cam, B.; Cinkilic, N.; Gurun, M.S. Pharmacologic overview of chlorogenic acid and its metabolites in chronic pain and inflammation. Curr. Neuropharmacol. 2020, 18, 216–228. [Google Scholar] [CrossRef]
- Bai, J.; Zhang, Y.; Tang, C.; Hou, Y.; Ai, X.; Chen, X.; Zhang, Y.; Wang, X.; Meng, X. Gallic acid: Pharmacological activities and molecular mechanisms involved in inflammation-related diseases. Biomed. Pharmacother. 2021, 133, 110985. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.L.; Wei, J.; Bi, L.Q. Rutin attenuates oxidative stress and proinflammatory cytokine level in adjuvant induced rheumatoid arthritis via inhibition of NF-κB. Pharmacology 2017, 100, 40–49. [Google Scholar] [CrossRef]
- Choi, E.M. Oxidative status of DBA/1J mice with type II collagen-induced arthritis. J. Appl. Toxicol. Int. J. 2007, 27, 472–481. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.Y.; Tsai, T.C.; Li, Y.T.; Ding, Y.C.; Wang, C.T.; Hsieh, J.L.; Wu, C.L.; Wu, P.T.; Shiau, A.L. Interleukin-23 Mediates Osteoclastogenesis in Collagen-Induced Arthritis by Modulating MicroRNA-223. Int. J. Mol. Sci. 2022, 23, 9718. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Xie, D.; Gong, W.; Wu, H.; Qiang, Y. Pentahydroxy flavonoid isolated from Madhuca indica ameliorated adjuvant-induced arthritis via modulation of inflammatory pathways. Sci. Rep. 2021, 11, 17971. [Google Scholar] [CrossRef]
- Alabarse, P.V.; Silva, J.M.; Santo, R.C.; Oliveira, M.S.; Almeida, A.S.; de Oliveira, M.S.; Immig, M.L.; Freitas, E.C.; Teixeira, V.O.N.; Xavier, R.M.; et al. Metabolomic Biomarker Candidates for Skeletal Muscle Loss in the Collagen-Induced Arthritis (CIA) Model. J. Pers. Med. 2021, 11, 837. [Google Scholar] [CrossRef]
- Casanova-Vallve, N.; Constantin-Teodosiu, D.; Filer, A.; Hardy, R.S.; Greenhaff, P.L.; Chapman, V. Skeletal muscle dysregulation in rheumatoid arthritis: Metabolic and molecular markers in a rodent model and patients. PLoS ONE 2020, 15, e0235702. [Google Scholar] [CrossRef] [PubMed]
- Oike, T.; Sato, Y.; Kobayashi, T.; Miyamoto, K.; Nakamura, S.; Kaneko, Y.; Kobayashi, S.; Harato, K.; Saya, H.; Miyamoto, T.; et al. Stat3 as a potential therapeutic target for rheumatoid arthritis. Sci. Rep. 2017, 7, 10965. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Forward Sequence | Reverse Sequence | PCR Product Size |
|---|---|---|---|
| NF-KΒ | CATGAAGAGAAGACACTGACCATGGAAA | TGGATAGAGGCTAAGTGT AGACACG | 198 |
| COX-2 | TGTATGCTACCATCTGGCTTCGG | GTTTGGAACAGTCGCTCGTCATC | 147 |
| PTEN | TGTGGTCTGCCAGCTAAAGG | ACACACAGGTAACGGCTGAG | 192 |
| VEGF | TGCAGATTATGCGGATCAAACC | TGCATTCACATTTGTTGTGCTGTAG | 187 |
| OTP | ACATCCAGTACCCTGATGCTACAG | TGGCCTTGTATGCACCATTC | 204 |
| OTC | AGCAAAGGTGCAGCCTTTGT | GCGCCTGGGTCTCTTCACT | 176 |
| GAPDH | GCAAGGATACTGAGAGCAAGAG | GGATGGAATTGTGAGGGAGATG | 113 |
| Plant Extracts | EC50 Values (μg/mL) of Radical Scavenging | ||
|---|---|---|---|
| Hexane Extract | OL-EAE | OL-HAE | |
| DPPH | 2.22 ± 0.11 a | 1.8 ± 0.12 a | 3.2 ± 0.3 b |
| FRAP | 2.35 ± 0.08 a | 0.88 ± 0.03 bc | 1.35 ± 0.02 b |
| ABTS | 34.41 ± 2.47 a | 11.99 ± 1.66 c | 21.2 ± 1.02 bc |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Almansour, Z.H.; Ibrahim, H.-I.M.; Hamad, R.S.; Abd El-Moaty, H.I. Phenolic-Compound-Rich Opuntia littoralis Ethyl Acetate Extract Relaxes Arthritic Symptoms in Collagen-Induced Mice Model via Bone Morphogenic Markers. Nutrients 2022, 14, 5366. https://doi.org/10.3390/nu14245366
Almansour ZH, Ibrahim H-IM, Hamad RS, Abd El-Moaty HI. Phenolic-Compound-Rich Opuntia littoralis Ethyl Acetate Extract Relaxes Arthritic Symptoms in Collagen-Induced Mice Model via Bone Morphogenic Markers. Nutrients. 2022; 14(24):5366. https://doi.org/10.3390/nu14245366
Chicago/Turabian StyleAlmansour, Zainab H., Hairul-Islam Mohamed Ibrahim, Rabab S. Hamad, and Heba Ibrahim Abd El-Moaty. 2022. "Phenolic-Compound-Rich Opuntia littoralis Ethyl Acetate Extract Relaxes Arthritic Symptoms in Collagen-Induced Mice Model via Bone Morphogenic Markers" Nutrients 14, no. 24: 5366. https://doi.org/10.3390/nu14245366
APA StyleAlmansour, Z. H., Ibrahim, H.-I. M., Hamad, R. S., & Abd El-Moaty, H. I. (2022). Phenolic-Compound-Rich Opuntia littoralis Ethyl Acetate Extract Relaxes Arthritic Symptoms in Collagen-Induced Mice Model via Bone Morphogenic Markers. Nutrients, 14(24), 5366. https://doi.org/10.3390/nu14245366