
Salvia plebeia R. Br. Water Extract Ameliorates Hepatic Steatosis in a Non-Alcoholic Fatty Liver Disease Model by Regulating the AMPK Pathway

Abstract
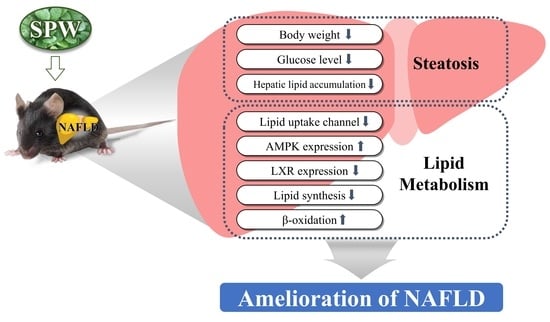
:
1. Introduction
2. Materials and Methods
2.1. Preparation of SPW
2.2. Measurement of Total Phenolic Compounds and Flavonoid Content
2.3. Cell Culture and Cell Viability Assay
2.4. Oil Red O Staining
2.5. AdipoRed Assay
2.6. Animals and Experimental Design
2.7. Oral Glucose Tolerance Test (OGTT)
2.8. Histological Analysis of the Liver
2.9. Biochemical Assays
2.10. Total RNA Isolation and Real-Time Polymerase Chain Reaction
2.11. Statistical Analysis
3. Results
3.1. Total Phenolic Compounds and Flavonoid Content of SPW was Determinated
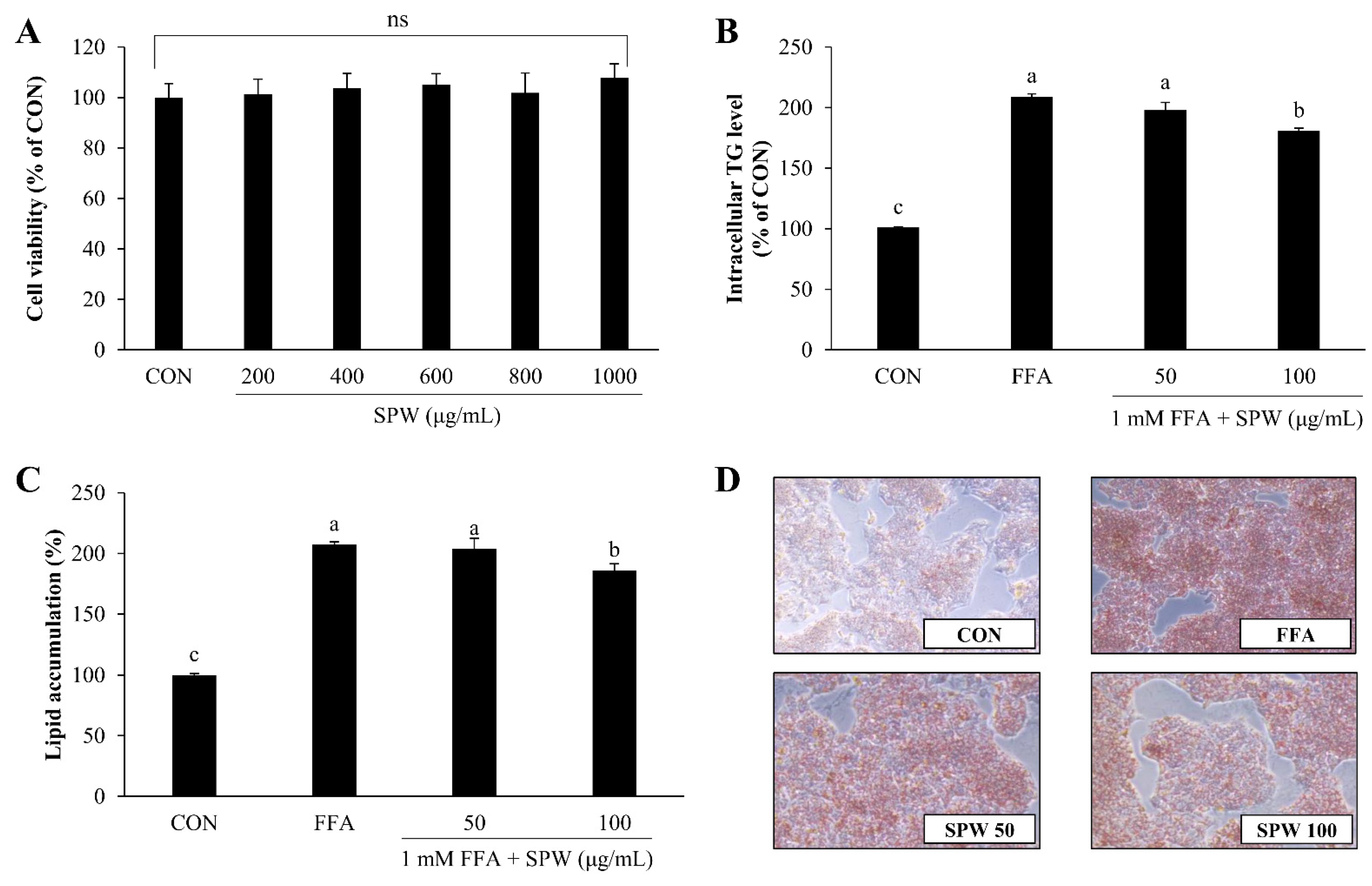
3.2. SPW Ameliorates FFA-Induced Lipid Accumulation
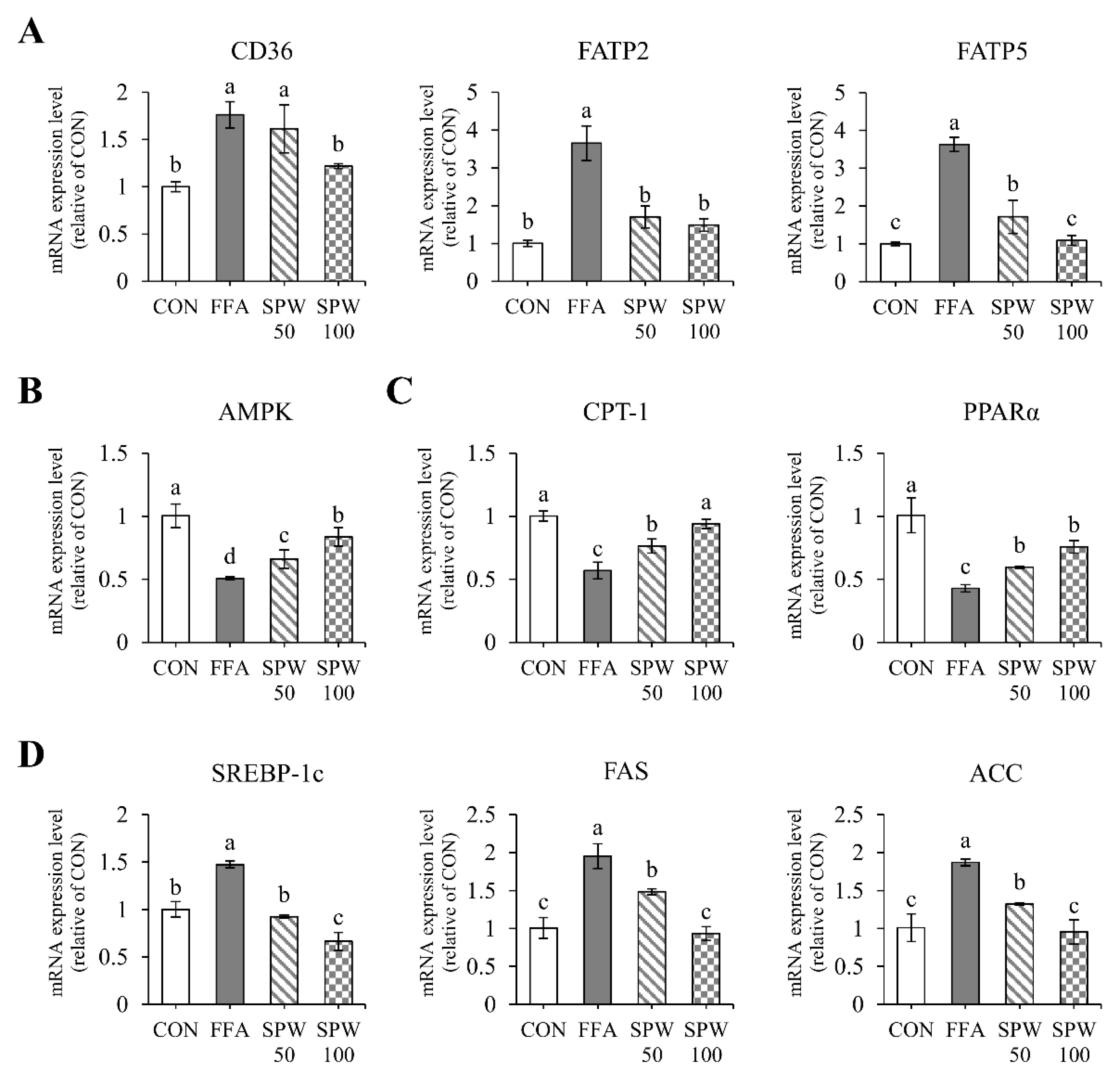
3.3. SPW Regulates Lipid Metabolism via the AMPK Pathway in FFA-Induced Hepatic Steatosis
3.4. SPW Improves HFD-Induced Body Weight Gain
3.5. SPW Improves HFD-Induced Glucose Level
3.6. SPW Ameliorates HFD-Induced Hepatic Steatosis and Liver Injury
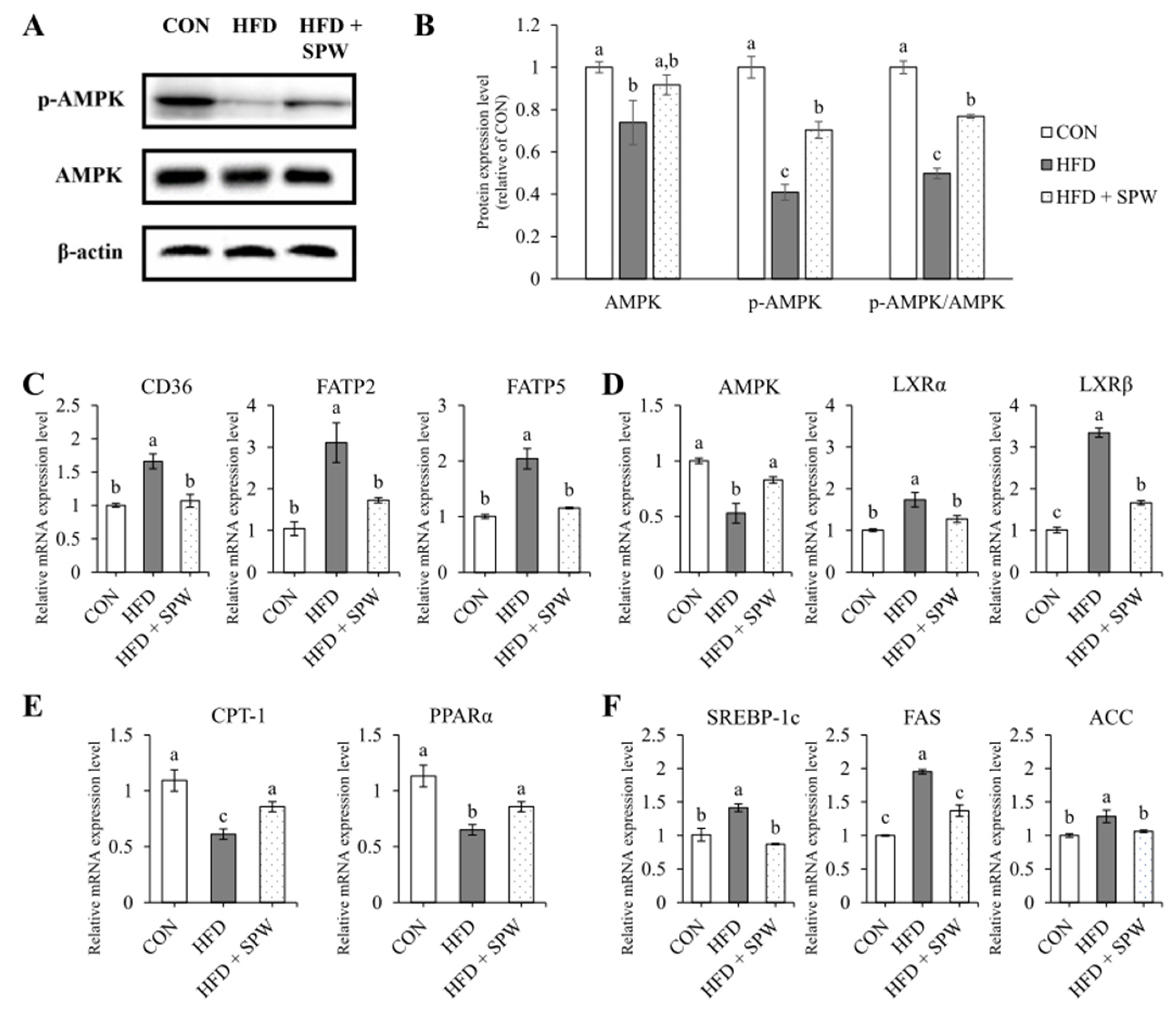
3.7. SPW Regulates Lipid Metabolism via AMPK Pathway in HFD-Induced Hepatic Steatosis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Im, H.J.; Ahn, Y.C.; Wang, J.-H.; Lee, M.M.; Son, C.G. Systematic review on the prevalence of nonalcoholic fatty liver disease in South Korea. Clin. Res. Hepatol. Gastroenterol. 2021, 45, 101526. [Google Scholar] [CrossRef] [PubMed]
- Mitra, S.; De, A.; Chowdhury, A. Epidemiology of non-alcoholic and alcoholic fatty liver diseases. Transl. Gastroenterol. Hepatol. 2020, 5, 16. [Google Scholar] [CrossRef] [PubMed]
- Buzzetti, E.; Pinzani, M.; Tsochatzis, E.A. The multiple-hit pathogenesis of non-alcoholic fatty liver disease (NAFLD). Metabolism 2016, 65, 1038–1048. [Google Scholar] [CrossRef] [PubMed]
- Karim, M.; Al-Mahtab, M.; Rahman, S.; Debnath, C. Non-alcoholic Fatty Liver Disease (NAFLD)—A Review. Mymensingh Med. J. MMJ 2015, 24, 873–880. [Google Scholar]
- He, J.; Lee, J.H.; Febbraio, M.; Xie, W. The emerging roles of fatty acid translocase/CD36 and the aryl hydrocarbon receptor in fatty liver disease. Exp. Biol. Med. 2011, 236, 1116–1121. [Google Scholar] [CrossRef]
- Enooku, K.; Tsutsumi, T.; Kondo, M.; Fujiwara, N.; Sasako, T.; Shibahara, J.; Kado, A.; Okushin, K.; Fujinaga, H.; Nakagomi, R. Hepatic FATP5 expression is associated with histological progression and loss of hepatic fat in NAFLD patients. J. Gastroenterol. 2020, 55, 227–243. [Google Scholar] [CrossRef]
- Mun, J.; Kim, S.; Yoon, H.G.; You, Y.; Kim, O.K.; Choi, K.C.; Lee, Y.H.; Lee, J.; Park, J.; Jun, W. Water Extract of Curcuma longa L. Ameliorates Non-Alcoholic Fatty Liver Disease. Nutrients 2019, 11, 2536. [Google Scholar] [CrossRef] [Green Version]
- Oliveras-Ferraros, C.; Vazquez-Martin, A.; Fernández-Real, J.M.; Menendez, J.A. AMPK-sensed cellular energy state regulates the release of extracellular Fatty Acid Synthase. Biochem. Biophys. Res. Commun. 2009, 378, 488–493. [Google Scholar] [CrossRef]
- Hu, Y.; Yin, F.; Liu, Z.; Xie, H.; Xu, Y.; Zhou, D.; Zhu, B. Acerola polysaccharides ameliorate high-fat diet-induced non-alcoholic fatty liver disease through reduction of lipogenesis and improvement of mitochondrial functions in mice. Food Funct. 2020, 11, 1037–1048. [Google Scholar] [CrossRef]
- Lund, E.G.; Peterson, L.B.; Adams, A.D.; Lam, M.H.; Burton, C.A.; Chin, J.; Guo, Q.; Huang, S.; Latham, M.; Lopez, J.C.; et al. Different roles of liver X receptor alpha and beta in lipid metabolism: Effects of an alpha-selective and a dual agonist in mice deficient in each subtype. Biochem. Pharmacol. 2006, 71, 453–463. [Google Scholar] [CrossRef]
- Liang, Y.-Y.; Wan, X.-H.; Niu, F.-J.; Xie, S.-M.; Guo, H.; Yang, Y.-Y.; Guo, L.-Y.; Zhou, C.-Z. Salvia plebeia R. Br.: An overview about its traditional uses, chemical constituents, pharmacology and modern applications. Biomed. Pharmacother. 2020, 121, 109589. [Google Scholar] [CrossRef] [PubMed]
- Seo, H.W.; Suh, J.H.; Kyung, J.-S.; Jang, K.H.; So, S.-H. Subacute oral toxicity and bacterial mutagenicity study of a mixture of korean red ginseng (Panax ginseng CA Meyer) and Salvia plebeia R. Br. Extracts. Toxicol. Res. 2019, 35, 215–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bang, S.; Ha, T.K.Q.; Lee, C.; Li, W.; Oh, W.-K.; Shim, S.H. Antiviral activities of compounds from aerial parts of Salvia plebeia R. Br. J. Ethnopharmacol. 2016, 192, 398–405. [Google Scholar] [CrossRef] [PubMed]
- Qu, X.J.; Xia, X.; Wang, Y.S.; Song, M.J.; Liu, L.L.; Xie, Y.Y.; Cheng, Y.N.; Liu, X.J.; Qiu, L.L.; Xiang, L.; et al. Protective effects of Salvia plebeia compound homoplantaginin on hepatocyte injury. Food Chem. Toxicol. 2009, 47, 1710–1715. [Google Scholar] [CrossRef]
- Lee, G.T.; Duan, C.H.; Lee, J.-N.; Lee, K.-S.; Hong, J.-T.; Lee, K.-K. Phytochemical constituents from Salvia plebeia. Nat. Prod. Sci. 2010, 16, 207–210. [Google Scholar]
- Weng, X.; Wang, W. Antioxidant activity of compounds isolated from Salvia plebeia. Food Chem. 2000, 71, 489–493. [Google Scholar] [CrossRef]
- Choi, S.I.; Cho, I.H.; Han, S.H.; Jeon, Y.J.; Choi, J.G.; Kim, J.S.; Lee, J.H. Antiobesity Effects of Salvia plebeia R. Br. Extract in High-Fat Diet-Induced Obese Mice. J. Med. Food 2016, 19, 1048–1056. [Google Scholar] [CrossRef]
- Won, H.R. Effect of Salvia plebeia water extract on antioxidant activity and lipid composition of rats fed a high fat-high cholesterol diet. Korean J. Community Living Sci. 2016, 27, 233–243. [Google Scholar] [CrossRef] [Green Version]
- Meda, A.; Lamien, C.E.; Romito, M.; Millogo, J.; Nacoulma, O.G. Determination of the total phenolic, flavonoid and proline contents in Burkina Fasan honey, as well as their radical scavenging activity. Food Chem. 2005, 91, 571–577. [Google Scholar] [CrossRef]
- Atanassova, M.; Georgieva, S.; Ivancheva, K. Total phenolic and total flavonoid contents, antioxidant capacity and biological contaminants in medicinal herbs. J. Univ. Chem. Technol. Metall. 2011, 46, 81–88. [Google Scholar]
- Roehm, N.W.; Rodgers, G.H.; Hatfield, S.M.; Glasebrook, A.L. An improved colorimetric assay for cell proliferation and viability utilizing the tetrazolium salt XTT. J. Immunol. Methods 1991, 142, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Iverson, S.J.; Lang, S.L.; Cooper, M.H. Comparison of the Bligh and Dyer and Folch methods for total lipid determination in a broad range of marine tissue. Lipids 2001, 36, 1283–1287. [Google Scholar] [CrossRef] [PubMed]
- Mu, H.-N.; Zhou, Q.; Yang, R.-Y.; Tang, W.-Q.; Li, H.-X.; Wang, S.-M.; Li, J.; Chen, W.-X.; Dong, J. Caffeic acid prevents non-alcoholic fatty liver disease induced by a high-fat diet through gut microbiota modulation in mice. Food Res. Int. 2021, 143, 110240. [Google Scholar] [CrossRef]
- Balachander, G.J.; Subramanian, S.; Ilango, K. Rosmarinic acid attenuates hepatic steatosis by modulating ER stress and autophagy in oleic acid-induced HepG2 cells. RSC Adv. 2018, 8, 26656–26663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Liu, R.; Shen, Z.; Cai, G. Combination of luteolin and lycopene effectively protect against the “two-hit” in NAFLD through Sirt1/AMPK signal pathway. Life Sci. 2020, 256, 117990. [Google Scholar] [CrossRef] [PubMed]
- Han, L.P.; Sun, B.; Li, C.J.; Xie, Y.; Chen, L.M. Effect of celastrol on toll-like receptor 4-mediated inflammatory response in free fatty acid-induced HepG2 cells. Int. J. Mol. Med. 2018, 42, 2053–2061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.-C.; Chen, H.-J.; Huang, B.-M.; Chen, Y.-C.; Chang, C.-F. Polyphenol-Rich Extracts from Toona sinensis Bark and Fruit Ameliorate Free Fatty Acid-Induced Lipogenesis through AMPK and LC3 Pathways. J. Clin. Med. 2019, 8, 1664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Im, A.-R.; Kim, Y.H.; Lee, H.W.; Song, K.H. Water extract of Dolichos lablab attenuates hepatic lipid accumulation in a cellular nonalcoholic fatty liver disease model. J. Med. Food 2016, 19, 495–503. [Google Scholar] [CrossRef]
- Mitmesser, S.H.; Carr, T.P. Trans fatty acids alter the lipid composition and size of apoB-100-containing lipoproteins secreted by HepG2 cells. J. Nutr. Biochem. 2005, 16, 178–183. [Google Scholar] [CrossRef]
- Gaggini, M.; Morelli, M.; Buzzigoli, E.; DeFronzo, R.A.; Bugianesi, E.; Gastaldelli, A. Non-alcoholic fatty liver disease (NAFLD) and its connection with insulin resistance, dyslipidemia, atherosclerosis and coronary heart disease. Nutrients 2013, 5, 1544–1560. [Google Scholar] [CrossRef]
- Goorden, S.; Buffart, T.E.; Bakker, A.; Buijs, M.M. Liver disorders in adults: ALT and AST. Ned. Tijdschr. Voor Geneeskd. 2013, 157, A6443. [Google Scholar]
- Chiang, J.Y.; Ferrell, J.M. Bile acid metabolism in liver pathobiology. Gene Expr. 2018, 18, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rui, L. Energy metabolism in the liver. Compr. Physiol. 2014, 4, 177–179. [Google Scholar] [PubMed] [Green Version]
- Chen, L.; Chen, X.-W.; Huang, X.; Song, B.-L.; Wang, Y.; Wang, Y. Regulation of glucose and lipid metabolism in health and disease. Sci. China Life Sci. 2019, 62, 1420–1458. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Shen, L.; Tan, Z.; Zhang, P.; Zhao, X.; Xu, Y.; Gan, M.; Yang, Q.; Ma, J.; Jiang, A.A. Betaine supplementation enhances lipid metabolism and improves insulin resistance in mice fed a high-fat diet. Nutrients 2018, 10, 131. [Google Scholar] [CrossRef] [Green Version]
- Sekizkardes, H.; Chung, S.T.; Chacko, S.; Haymond, M.W.; Startzell, M.; Walter, M.; Walter, P.J.; Lightbourne, M.; Brown, R.J. Free fatty acid processing diverges in human pathologic insulin resistance conditions. J. Clin. Investig. 2020, 130, 3592–3602. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Yoo, W.; Park, H.M.; Lim, S.Y.; Shin, D.H.; Kim, S.; Park, H.Y.; Jeong, T.S. Arazyme Suppresses Hepatic Steatosis and Steatohepatitis in Diet-Induced Non-Alcoholic Fatty Liver Disease-Like Mouse Model. Int. J. Mol. Sci. 2019, 20, 2325. [Google Scholar] [CrossRef] [Green Version]
- Oligschlaeger, Y.; Shiri-Sverdlov, R. NAFLD Preclinical Models: More than a Handful, Less of a Concern? Biomedicines 2020, 8, 28. [Google Scholar] [CrossRef] [Green Version]
- Castro, R.E.; Diehl, A.M. Towards a definite mouse model of NAFLD. J. Hepatol. 2018, 69, 272–274. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, A.; Terauchi, Y. Lessons from mouse models of high-fat diet-induced NAFLD. Int. J. Mol. Sci. 2013, 14, 21240–21257. [Google Scholar] [CrossRef] [Green Version]
- Prill, S.; Caddeo, A.; Baselli, G.; Jamialahmadi, O.; Dongiovanni, P.; Rametta, R.; Kanebratt, K.P.; Pujia, A.; Pingitore, P.; Mancina, R.M. The TM6SF2 E167K genetic variant induces lipid biosynthesis and reduces apolipoprotein B secretion in human hepatic 3D spheroids. Sci. Rep. 2019, 9, 11585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mundi, M.S.; Velapati, S.; Patel, J.; Kellogg, T.A.; Abu Dayyeh, B.K.; Hurt, R.T. Evolution of NAFLD and its management. Nutr. Clin. Pract. 2020, 35, 72–84. [Google Scholar] [CrossRef] [PubMed]
- Utzschneider, K.M.; Kahn, S.E.; Polidori, D.C. Hepatic insulin extraction in NAFLD is related to insulin resistance rather than liver fat content. J. Clin. Endocrinol. Metab. 2019, 104, 1855–1865. [Google Scholar] [CrossRef] [Green Version]
- Wilson, C.G.; Tran, J.L.; Erion, D.M.; Vera, N.B.; Febbraio, M.; Weiss, E.J. Hepatocyte-Specific Disruption of CD36 Attenuates Fatty Liver and Improves Insulin Sensitivity in HFD-Fed Mice. Endocrinology 2016, 157, 570–585. [Google Scholar] [CrossRef] [PubMed]
- Qiu, P.; Wang, H.; Zhang, M.; Zhang, M.; Peng, R.; Zhao, Q.; Liu, J. FATP2-targeted therapies—A role beyond fatty liver disease. Pharmacol. Res. 2020, 161, 105228. [Google Scholar] [CrossRef] [PubMed]
- Kumari, A.; Pathak, D.P.; Asthana, S. Bile acids mediated potential functional interaction between FXR and FATP5 in the regulation of Lipid Metabolism. Int. J. Biol. Sci. 2020, 16, 2308. [Google Scholar] [CrossRef] [PubMed]
- Hardwick, J.P.; Eckman, K.; Lee, Y.K.; Abdelmegeed, M.A.; Esterle, A.; Chilian, W.M.; Chiang, J.Y.; Song, B.-J. Chapter Five—Eicosanoids in Metabolic Syndrome. In Advances in Pharmacology; Webb, D.R., Ed.; Academic Press: Cambridge, MA, USA, 2013; Volume 66, pp. 157–266. [Google Scholar]
- Jeon, S.-M. Regulation and function of AMPK in physiology and diseases. Exp. Mol. Med. 2016, 48, e245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Xu, S.; Mihaylova, M.M.; Zheng, B.; Hou, X.; Jiang, B.; Park, O.; Luo, Z.; Lefai, E.; Shyy, J.Y.J.; et al. AMPK phosphorylates and inhibits SREBP activity to attenuate hepatic steatosis and atherosclerosis in diet-induced insulin-resistant mice. Cell Metab. 2011, 13, 376–388. [Google Scholar] [CrossRef] [Green Version]
- Foretz, M.; Viollet, B. Regulation of hepatic metabolism by AMPK. J. Hepatol. 2011, 54, 827–829. [Google Scholar] [CrossRef]
- Calkin, A.C.; Tontonoz, P. Transcriptional integration of metabolism by the nuclear sterol-activated receptors LXR and FXR. Nat. Rev. Mol. Cell Biol. 2012, 13, 213–224. [Google Scholar] [CrossRef] [Green Version]
- Komati, R.; Spadoni, D.; Zheng, S.; Sridhar, J.; Riley, K.E.; Wang, G. Ligands of Therapeutic Utility for the Liver X Receptors. Molecules 2017, 22, 88. [Google Scholar] [CrossRef] [PubMed]
- Kratzer, A.; Buchebner, M.; Pfeifer, T.; Becker, T.M.; Uray, G.; Miyazaki, M.; Miyazaki-Anzai, S.; Ebner, B.; Chandak, P.G.; Kadam, R.S. Synthetic LXR agonist attenuates plaque formation in apoE-/-mice without inducing liver steatosis and hypertriglyceridemias. J. Lipid Res. 2009, 50, 312–326. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′ to 3′) | |
|---|---|---|
| hCD36 | Forward Reverse | 5′-TGGAACAGAGGCTGACAACT-3′ 5′-TTGATTTTTGATAGATATGGG-3′ |
| hFATP2 | Forward Reverse | 5′-CTTTCAGCACATTGCTGATTACCT-3′ 5′-CAGTGATCTCAATGGTGTCCTGTAT-3′ |
| hFATP5 | Forward Reverse | 5′-AGCTCCTGCGGTACTTGTGT-3′ 5′-AAGGTCTCCCACACATCAGC-3′ |
| hAMPK | Forward Reverse | 5′-GGCACCCTCCCATTTGATG-3′ 5′-ACACCCCCTCGGATCTTCTT-3′ |
| hAMPK | Forward Reverse | 5′-GGCACCCTCCCATTTGATG-3′ 5′-ACACCCCCTCGGATCTTCTT-3′ |
| hCPT-1 | Forward Reverse | 5′-TGTTGGGTATGCTGTTCATGACA-3′ 5′-GCGGCCTGGGTAGGAAGA-3′ |
| hPPARα | Forward Reverse | 5′-AACATCCAAGAGATTTCGCAATC-3′ 5′-CCGTAAAGCCAAAGCTTCCA-3′ |
| hSREBP-1c | Forward Reverse | 5′-CGGAACCATCTTGGCAACA-3′ 5′-GCCGGTTGATAGGCAGCTT-3′ |
| hFAS | Forward Reverse | 5′-CGCTCGGCATGGCTATCT-3′ 5′-CTCGTTGAAGAACGCATCCA-3′ |
| hACC | Forward Reverse | 5′-TGCAGATCTTAGCGGACCAA-3′ 5′-GCCTGCGTTGTACAGAGCAA-3′ |
| hβ-Actin | Forward Reverse | 5′-ACGGCCAGGTCATCACTATTG-3′ 5′-CAAGAAGGAAGGCTGGAAAAGA-3′ |
| mCD36 | Forward Reverse | 5′-TTGAAGGCATTCCCACGTATC-3′ 5′-CGGACCCGTTGGCAAA-3′ |
| mFATP2 | Forward Reverse | 5′-GGAACCACAGGTCTTCCAAA-3′ 5′-TAAAGTAGCCCCAACCACGA-3′ |
| mFATP5 | Forward Reverse | 5′-GGAACCACAGGTCTTCCAAA-3′ 5′-TAAAGTAGCCCCAACCACGA-3′ |
| mAMPK | Forward Reverse | 5′-TTCGTGCCGCCCCTTT-3′ 5′-GGTCAGCATGCCCACAAAA-3′ |
| mLXRα | Forward Reverse | 5′-CCTCTGGCTTCCATTACAAC-3′ 5′-CTTCTGACAGCACACACTC-3′ |
| mLXRβ | Forward Reverse | 5′-CACCATTGAGATCATGTTGC-3′ 5′-TTGATCCTCGTGTAGGAGAG-3′ |
| mCPT-1 | Forward Reverse | 5′-GTGACTGGTGGGAGGAATAC-3′ 5′-GAGCATCTCCATGGCGTAG-3′ |
| mPPARα | Forward Reverse | 5′-TGGCAAAAGGCAAGGAGAAG-3′ 5′-CCCTCTACATAGAACTGCAA-3′ |
| mSREBP-1c | Forward Reverse | 5′-TGGCTTGGTGATGCTATGTTG-3′ 5′-GACCATCAAGGCCCCTCAA-3′ |
| mFAS | Forward Reverse | 5′-GAAGTGTCTGGACTGTGTCATTTTTAC-3′ 5′-TTAATTGTGGGATCAGGAGAGCAT-3′ |
| mACC | Forward Reverse | 5′-TCCCCAAGTTCTTCACGTTCA-3′ 5′-CAGGCTCCAAGTGGCGATAA-3′ |
| mβ-Actin | Forward Reverse | 5′-AGCCATGTACGTAGCCATCC-3′ 5′-CTCTCAGCTGTGGTGGTGAA-3′ |
| Total Phenolic Compounds Content (mg GAE/100 g SPW) | Flavonoid Content (mg CE/100 g SPW) | |
|---|---|---|
| SPW | 6984.69 ± 138.25 | 5334.44 ± 172.78 |
| Groups | CON | HFD | HFD + SPW |
|---|---|---|---|
| Body mass (g) | |||
| Initial (A) | 22.78 ± 0.37 ns | 22.79 ± 0.33 | 22.76 ± 0.28 |
| Final (B) | 35.45 ± 0.66 c | 45.81 ± 0.67 a | 43.36 ± 0.61 b |
| Weight gain (B–A) | 12.67 ± 0.72 c | 23.10 ± 0.55 a | 20.61 ± 0.58 b |
| Food intake (g/day) | 3.23 ± 0.03 a | 2.52 ± 0.04 b | 2.66 ± 0.05 b |
| Energy intake (kcal/day) | 12.60 ± 0.13 ns | 13.19 ± 0.19 | 13.93 ± 0.25 |
| FER (%) | 4.62 ± 0.27 c | 10.61 ± 0.26 a | 9.19 ± 0.31 b |
| Liver weight (g) | 1.44 ± 0.02 b | 1.94 ± 0.05 a | 1.55 ± 0.12 b |
| Liver index (%) | 3.68 ± 0.04 b | 4.19 ± 0.10 a | 3.09 ± 0.21 c |
| Relative fat weight (g) | |||
| Perirenal fat | 0.75 ± 0.03 c | 1.39 ± 0.07 a | 1.08 ± 0.06 b |
| Epididymal fat | 1.49 ± 0.03 b | 2.12 ± 0.12 a | 1.62 ± 0.05 b |
| Groups | CON | HFD | HFD + SPW |
|---|---|---|---|
| Serum | |||
| TG (mg/dL) | 101.79 ± 1.05 c | 134.23 ± 3.51 a | 118.16 ± 1.91 b |
| TC (mg/dL) | 239.44 ± 1.27 b | 278.38 ± 5.56 a | 237.44 ± 3.22 b |
| HDL (mg/dL) | 122.12 ± 1.72 b | 95.75 ± 0.72 a | 119.55 ± 2.40 b |
| LDL (mg/dL) | 96.96 ± 2.06 b | 155.78 ± 5.13 a | 96.25 ± 4.79 b |
| NEFA (mEq/L) | 80.33 ± 2.66 b | 120.33 ± 5.61 a | 81.00 ± 3.22 b |
| Liver tissue | |||
| TG (mg/g liver) | 5.77 ± 0.35 c | 10.73 ± 0.07 a | 8.93 ± 0.05 b |
| TC (mg/g liver) | 2.80 ± 0.06 b | 4.16 ± 0.12 a | 2.75 ± 0.09 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bae, S.; Lee, Y.-H.; Lee, J.; Park, J.; Jun, W. Salvia plebeia R. Br. Water Extract Ameliorates Hepatic Steatosis in a Non-Alcoholic Fatty Liver Disease Model by Regulating the AMPK Pathway. Nutrients 2022, 14, 5379. https://doi.org/10.3390/nu14245379
Bae S, Lee Y-H, Lee J, Park J, Jun W. Salvia plebeia R. Br. Water Extract Ameliorates Hepatic Steatosis in a Non-Alcoholic Fatty Liver Disease Model by Regulating the AMPK Pathway. Nutrients. 2022; 14(24):5379. https://doi.org/10.3390/nu14245379
Chicago/Turabian StyleBae, Subin, Yoo-Hyun Lee, Jeongmin Lee, Jeongjin Park, and Woojin Jun. 2022. "Salvia plebeia R. Br. Water Extract Ameliorates Hepatic Steatosis in a Non-Alcoholic Fatty Liver Disease Model by Regulating the AMPK Pathway" Nutrients 14, no. 24: 5379. https://doi.org/10.3390/nu14245379