
Dietary Conjugated Linoleic Acid Reduces Body Weight and Fat in Snord116m+/p− and Snord116m−/p− Mouse Models of Prader–Willi Syndrome

 , ,
, , 
Abstract
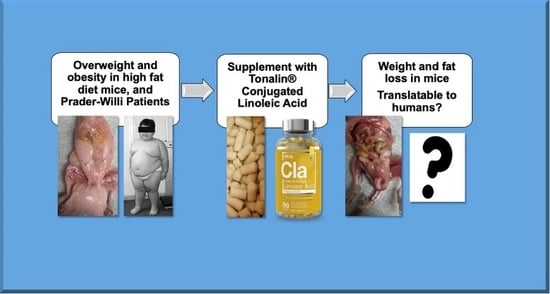
1. Introduction
2. Materials and Methods
2.1. Mouse Housing, Breeding, and Euthanasia
2.2. Study Design
2.3. Weekly Measurements
2.4. Study Pre- and Post-Measurements
2.4.1. Wheel Running
2.4.2. Elevated Plus
2.4.3. Glucose Tolerance Tests
2.5. End-of-Study Measurements
2.5.1. Rotarod Analysis
2.5.2. Indirect Calorimetry and Home-Cage Activity
2.5.3. In Vivo Muscle Function Testing
2.6. RNA Isolation
2.7. RNA-Seq
2.8. Reverse-Transcriptase Quantitative PCR
2.9. Microbiome Analysis
2.10. Histology
2.11. Statistical Analyses
3. Results
3.1. Body Weight and Fat Mass Are Reduced in PWS and PWS-KO Mice
3.2. Changes in Fasting Glucose and Glucose Tolerance with Genotype and Treatment
3.3. No Effect of CLA Treatment on Metabolism
3.4. CLA Modulates Spontaneous Wheel Running Levels without Effects on End-of-Study Rotarod, Home-Cage Activity, or Anxiety Measurements
3.5. In Vivo Muscle Function Modulation by CLA Treatment
3.6. CLA-Treatment Effects on Liver
3.7. RNA-Seq Analysis of Hypothalamic Gene Expression
3.8. Cecum Microbiome Analysis in CLA and Control Diet-Treated Animals
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, H.; Victor, A.K.; Klein, J.; Tacer, K.F.; Tai, D.J.; de Esch, C.; Nuttle, A.; Temirov, J.; Burnett, L.C.; Rosenbaum, M.; et al. Loss of MAGEL2 in Prader-Willi syndrome leads to decreased secretory granule and neuropeptide production. JCI Insight 2020, 5, e138576. [Google Scholar] [CrossRef] [PubMed]
- Tan, Q.; Potter, K.J.; Burnett, L.C.; Orsso, C.E.; Inman, M.; Ryman, D.C.; Haqq, A.M. Prader-Willi-Like Phenotype Caused by an Atypical 15q11.2 Microdeletion. Genes 2020, 11, 128. [Google Scholar] [CrossRef]
- Burnett, L.C.; LeDuc, C.A.; Sulsona, C.R.; Paull, D.; Rausch, R.; Eddiry, S.; Carli, J.F.; Morabito, M.V.; Skowronski, A.A.; Hubner, G.; et al. Deficiency in prohormone convertase PC1 impairs prohormone processing in Prader-Willi syndrome. J. Clin. Investig. 2017, 127, 293–305. [Google Scholar] [CrossRef] [PubMed]
- Bittel, D.C.; Kibiryeva, N.; Sell, S.M.; Strong, T.V.; Butler, M.G. Whole genome microarray analysis of gene expression in Prader-Willi syndrome. Am. J. Med. Genet. A 2007, 143, 430–442. [Google Scholar] [CrossRef] [PubMed]
- Kocher, M.A.; Huang, F.W.; Le, E.; Good, D.J. Snord116 Post-transcriptionally Increases Nhlh2 mRNA Stability: Implications for Human Prader-Willi Syndrome. Hum. Mol. Genet. 2021, 30, 1101–1110. [Google Scholar] [CrossRef] [PubMed]
- Good, D.J.; Porter, F.D.; Mahon, K.A.; Parlow, A.F.; Westphal, H.; Kirsch, I.R. Hypogonadism and obesity in mice with a targeted deletion of the Nhlh2 gene. Nat. Genet. 1997, 15, 397–401. [Google Scholar] [CrossRef]
- Ding, F.; Li, H.H.; Zhang, S.; Solomon, N.M.; Camper, S.A.; Cohen, P.; Francke, U. SnoRNA Snord116 (Pwcr1/MBII-85) deletion causes growth deficiency and hyperphagia in mice. PLoS ONE 2008, 3, e1709. [Google Scholar] [CrossRef]
- Polex-Wolf, J.; Lam, B.Y.; Larder, R.; Tadross, J.; Rimmington, D.; Bosch, F.; Cenzano, V.J.; Ayuso, E.; Ma, M.K.; Rainbow, K.; et al. Hypothalamic loss of Snord116 recapitulates the hyperphagia of Prader-Willi syndrome. J. Clin. Investig. 2018, 128, 960–969. [Google Scholar] [CrossRef]
- Kummerfeld, D.M.; Raabe, C.A.; Brosius, J.; Mo, D.; Skryabin, B.V.; Rozhdestvensky, T.S. A Comprehensive Review of Genetically Engineered Mouse Models for Prader-Willi Syndrome Research. Int. J. Mol. Sci. 2021, 22, 3613. [Google Scholar] [CrossRef]
- Eddiry, S.; Dienne, G.; Molinas, C.; Salles, J.; Auriol, F.C.; Gennero, I.; Bieth, E.; Skryabin, B.V.; Rozhdestvensky, T.S.; Burnett, L.C.; et al. SNORD116 and growth hormone therapy impact IGFBP7 in Prader-Willi syndrome. Genet. Med. 2021, 23, 1664–1672. [Google Scholar] [CrossRef]
- Qi, Y.; Purtell, L.; Fu, M.; Lee, N.J.; Aepler, J.; Zhang, L.; Loh, K.; Enriquez, R.F.; Baldock, P.A.; Zolotukhin, S.; et al. Snord116 is critical in the regulation of food intake and body weight. Sci. Rep. 2016, 6, 18614. [Google Scholar] [CrossRef] [PubMed]
- Pellikaan, K.; van Woerden, G.M.; Kleinendorst, L.; Rosenberg, A.G.W.; Horsthemke, B.; Grosser, C.; van Zutven, L.; van Rossum, E.F.C.; van der Lely, A.J.; Resnick, J.L.; et al. The Diagnostic Journey of a Patient with Prader-Willi-Like Syndrome and a Unique Homozygous SNURF-SNRPN Variant; Bio-Molecular Analysis and Review of the Literature. Genes 2021, 12, 875. [Google Scholar] [CrossRef]
- Hur, S.; Whitcomb, F.; Rhee, S.; Park, Y.; Good, D.J.; Park, Y. Effects of trans-10,cis-12 conjugated linoleic acid on body composition in genetically obese mice. J. Med. Food 2009, 12, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Gilliard, D.; Good, D.J.; Park, Y. Preventive effects of conjugated linoleic acid on obesity by improved physical activity in nescient basic helix-loop-helix 2 knockout mice during growth period. Food Funct. 2012, 3, 1280–1285. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Park, Y.; Kim, D.; Good, D.J.; Park, Y. Dietary conjugated nonadecadienoic acid prevents adult-onset obesity in nescient basic helix-loop-helix 2 knockout mice. J. Nutr. Biochem. 2013, 24, 556–566. [Google Scholar] [CrossRef]
- Kim, Y.; Kim, D.; Good, D.J.; Park, Y. Effects of postweaning administration of conjugated linoleic acid on development of obesity in nescient basic helix-loop-helix 2 knockout mice. J. Agric. Food Chem. 2015, 63, 5212–5223. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, Y.; Kim, Y.J.; Park, Y. Conjugated Linoleic Acid: Potential Health Benefits as a Functional Food Ingredient. Annu. Rev. Food Sci. Technol. 2016, 7, 221–244. [Google Scholar] [CrossRef]
- Yang, X. Growth hormone treatment for Prader-Willi syndrome: A review. Neuropeptides 2020, 83, 102084. [Google Scholar] [CrossRef]
- Hogg, S. A review of the validity and variability of the elevated plus-maze as an animal model of anxiety. Pharmacol. Biochem. Behav. 1996, 54, 21–30. [Google Scholar] [CrossRef]
- Coyle, C.A.; Jing, E.; Hosmer, T.; Powers, J.B.; Wade, G.; Good, D.J. Reduced voluntary activity precedes adult-onset obesity in Nhlh2 knockout mice. Physiol. Behav. 2002, 77, 387–402. [Google Scholar] [CrossRef]
- Weir, J.B. New methods for calculating metabolic rate with special reference to protein metabolism. J. Physiol. 1949, 109, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Hamm, S.E.; Fathalikhani, D.D.; Bukovec, K.E.; Addington, A.K.; Zhang, H.; Perry, J.B.; McMillan, R.P.; Lawlor, M.W.; Prom, M.J.; Vanden Avond, M.A.; et al. Voluntary wheel running complements microdystrophin gene therapy to improve muscle function in mdx mice. Mol. Ther. Methods Clin. Dev. 2021, 21, 144–160. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Ander, S. Differential Analysis of Count Data—The DESeq2 Package. 2016. Available online: https://citeseerx.ist.psu.edu/viewdoc/summary?doi=10.1.1.733.1832 (accessed on 29 September 2021).
- Love, M.; Anders, S.; Huber, W. Differential analysis of count data—The DESeq2 package—4 February 2014. Genome Biol. 2014, 15, 10–1186. [Google Scholar]
- Li, Y.; Andrade, J. DEApp: An interactive web interface for differential expression analysis of next generation sequence data. Source Code Biol. Med. 2017, 12, 2. [Google Scholar] [CrossRef]
- Motenko, H.; Neuhauser, S.B.; O’Keefe, M.; Richardson, J.E. MouseMine: A new data warehouse for MGI. Mamm. Genome 2015, 26, 325–330. [Google Scholar] [CrossRef]
- Kozich, J.J.; Westcott, S.L.; Baxter, N.T.; Highlander, S.K.; Schloss, P.D. Development of a dual-index sequencing strategy and curation pipeline for analyzing amplicon sequence data on the MiSeq Illumina sequencing platform. Appl. Environ. Microbiol. 2013, 79, 5112–5120. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Subramanian, S.; Faith, J.J.; Gevers, D.; Gordon, J.I.; Knight, R.; Mills, D.A.; Caporaso, J.G. Quality-filtering vastly improves diversity estimates from Illumina amplicon sequencing. Nat. Methods 2013, 10, 57–59. [Google Scholar] [CrossRef]
- Amir, A.; McDonald, D.; Navas-Molina, J.A.; Kopylova, E.; Morton, J.T.; Zech, X.Z.; Kightley, E.P.; Thompson, L.R.; Hyde, E.R.; Gonzalez, A.; et al. Deblur Rapidly Resolves Single-Nucleotide Community Sequence Patterns. mSystems 2017, 2, e00191-16. [Google Scholar] [CrossRef]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—Approximately maximum-likelihood trees for large alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef] [PubMed]
- McDonald, D.; Price, M.N.; Goodrich, J.; Nawrocki, E.P.; DeSantis, T.Z.; Probst, A.; Andersen, G.L.; Knight, R.; Hugenholtz, P. An improved Greengenes taxonomy with explicit ranks for ecological and evolutionary analyses of bacteria and archaea. ISME J. 2012, 6, 610–618. [Google Scholar] [CrossRef] [PubMed]
- Lozupone, C.; Lladser, M.E.; Knights, D.; Stombaugh, J.; Knight, R. UniFrac: An effective distance metric for microbial community comparison. ISME J. 2011, 5, 169–172. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Adhikari, A.; Copping, N.A.; Onaga, B.; Pride, M.C.; Coulson, R.L.; Yang, M.; Yasui, D.H.; LaSalle, J.M.; Silverman, J.L. Cognitive deficits in the Snord116 deletion mouse model for Prader-Willi syndrome. Neurobiol. Learn. Mem. 2019, 165, 106874. [Google Scholar] [CrossRef]
- Zieba, J.; Low, J.K.; Purtell, L.; Qi, Y.; Campbell, L.; Herzog, H.; Karl, T. Behavioural characteristics of the Prader-Willi syndrome related biallelic Snord116 mouse model. Neuropeptides 2015, 53, 71–77. [Google Scholar] [CrossRef]
- Pinkoski, C.; Chilibeck, P.D.; Candow, D.G.; Esliger, D.; Ewaschuk, J.B.; Facci, M.; Farthing, J.P.; Zello, G.A. The effects of conjugated linoleic acid supplementation during resistance training. Med. Sci. Sports Exerc. 2006, 38, 339–348. [Google Scholar] [CrossRef]
- Poirier, H.; Niot, I.; Clement, L.; Guerre-Millo, M.; Besnard, P. Development of conjugated linoleic acid (CLA)-mediated lipoatrophic syndrome in the mouse. Biochimie 2005, 87, 73–79. [Google Scholar] [CrossRef]
- Jing, E.; Nillni, E.A.; Sanchez, V.C.; Stuart, R.C.; Good, D.J. Deletion of the Nhlh2 transcription factor decreases the levels of the anorexigenic peptides alpha melanocyte-stimulating hormone and thyrotropin-releasing hormone and implicates prohormone convertases I and II in obesity. Endocrinology 2004, 145, 1503–1513. [Google Scholar] [CrossRef]
- Qi, Y.; Purtell, L.; Fu, M.; Zhang, L.; Zolotukhin, S.; Campbell, L.; Herzog, H. Hypothalamus Specific Re-Introduction of SNORD116 into Otherwise Snord116 Deficient Mice Increased Energy Expenditure. J. Neuroendocrinol. 2017, 29, e12457. [Google Scholar] [CrossRef]
- Kruger, M.; Ruschke, K.; Braun, T. NSCL-1 and NSCL-2 synergistically determine the fate of GnRH-1 neurons and control necdin gene expression. EMBO J. 2004, 23, 4353–4364. [Google Scholar] [CrossRef] [PubMed]
- Butler, M.G.; Miller, J.L.; Forster, J.L. Prader-Willi Syndrome—Clinical Genetics, Diagnosis and Treatment Approaches: An Update. Curr. Pediatr. Rev. 2019, 15, 207–244. [Google Scholar] [CrossRef] [PubMed]
- Zahova, S.; Isles, A.R. Animal models for Prader-Willi syndrome. Handb. Clin. Neurol. 2021, 181, 391–404. [Google Scholar] [CrossRef]
- Baldini, L.; Robert, A.; Charpentier, B.; Labialle, S. Phylogenetic and Molecular Analyses Identify SNORD116 Targets Involved in the Prader-Willi Syndrome. Mol. Biol. Evol. 2022, 39, msab348. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhuang, P.; Zhang, Y.; Shou, Q.; Lu, Y.; Wang, G.; Qiu, J.; Wang, J.; He, L.; Chen, J.; et al. Mixed conjugated linoleic acid sex-dependently reverses high-fat diet-induced insulin resistance via the gut-adipose axis. FASEB J. 2021, 35, e21466. [Google Scholar] [CrossRef]
- Fleck, A.K.; Hucke, S.; Teipel, F.; Eschborn, M.; Janoschka, C.; Liebmann, M.; Wami, H.; Korn, L.; Pickert, G.; Hartwig, M.; et al. Dietary conjugated linoleic acid links reduced intestinal inflammation to amelioration of CNS autoimmunity. Brain 2021, 144, 1152–1166. [Google Scholar] [CrossRef]
- Crovesy, L.; Masterson, D.; Rosado, E.L. Profile of the gut microbiota of adults with obesity: A systematic review. Eur. J. Clin. Nutr. 2020, 74, 1251–1262. [Google Scholar] [CrossRef]
- Ze, X.; Duncan, S.H.; Louis, P.; Flint, H.J. Ruminococcus bromii is a keystone species for the degradation of resistant starch in the human colon. ISME J. 2012, 6, 1535–1543. [Google Scholar] [CrossRef]
- Bosshard, P.P.; Zbinden, R.; Altwegg, M. Turicibacter sanguinis gen. nov., sp. nov., a novel anaerobic, Gram-positive bacterium. Int. J. Syst. Evol. Microbiol. 2002, 52, 1263–1266. [Google Scholar] [CrossRef]
- Coupaye, M.; Tauber, M.; Cuisset, L.; Laurier, V.; Bieth, E.; Lacorte, J.M.; Oppert, J.M.; Clement, K.; Poitou, C. Effect of Genotype and Previous GH Treatment on Adiposity in Adults With Prader-Willi Syndrome. J. Clin. Endocrinol. Metab. 2016, 101, 4895–4903. [Google Scholar] [CrossRef]
- Bekx, M.T.; Carrel, A.L.; Shriver, T.C.; Li, Z.; Allen, D.B. Decreased energy expenditure is caused by abnormal body composition in infants with Prader-Willi Syndrome. J. Pediatr. 2003, 143, 372–376. [Google Scholar] [CrossRef]
- Morales, J.S.; Valenzuela, P.L.; Pareja-Galeano, H.; Rincon-Castanedo, C.; Rubin, D.A.; Lucia, A. Physical exercise and Prader-Willi syndrome: A systematic review. Clin. Endocrinol. 2019, 90, 649–661. [Google Scholar] [CrossRef]
- DeLany, J.P.; Blohm, F.; Truett, A.A.; Scimeca, J.A.; West, D.B. Conjugated linoleic acid rapidly reduces body fat content in mice without affecting energy intake. Am. J. Physiol. 1999, 276, R1172–R1179. [Google Scholar] [CrossRef]
- Kamphuis, M.M.; Lejeune, M.P.; Saris, W.H.; Westerterp-Plantenga, M.S. The effect of conjugated linoleic acid supplementation after weight loss on body weight regain, body composition, and resting metabolic rate in overweight subjects. Int. J. Obes. Relat. Metab. Disord. 2003, 27, 840–847. [Google Scholar] [CrossRef]
- Pariza, M.W.; Park, Y.; Cook, M.E. Conjugated linoleic acid and the control of cancer and obesity. Toxicol. Sci. 1999, 52, 107–110. [Google Scholar] [CrossRef] [PubMed]
- Bellicha, A.; Coupaye, M.; Mosbah, H.; Tauber, M.; Oppert, J.M.; Poitou, C. Physical Activity in Patients with Prader-Willi Syndrome-A Systematic Review of Observational and Interventional Studies. J. Clin. Med. 2021, 10, 2528. [Google Scholar] [CrossRef] [PubMed]
- Reus, L.; Zwarts, M.; van Vlimmeren, L.A.; Willemsen, M.A.; Otten, B.J.; Nijhuis-van der Sanden, M.W. Motor problems in Prader-Willi syndrome: A systematic review on body composition and neuromuscular functioning. Neurosci. Biobehav. Rev. 2011, 35, 956–969. [Google Scholar] [CrossRef] [PubMed]
- Pamukoff, D.N.; Holmes, S.C.; Shumski, E.J.; Garcia, S.A.; Rubin, D.A. Plantar Flexor Function in Adults with and without Prader-Willi Syndrome. Med. Sci. Sports Exerc. 2020, 52, 2189–2197. [Google Scholar] [CrossRef]
- Capodaglio, P.; Vismara, L.; Menegoni, F.; Baccalaro, G.; Galli, M.; Grugni, G. Strength characterization of knee flexor and extensor muscles in Prader-Willi and obese patients. BMC Musculoskelet Disord. 2009, 10, 47. [Google Scholar] [CrossRef]
- Lee, S.R.; Khamoui, A.V.; Jo, E.; Zourdos, M.C.; Panton, L.B.; Ormsbee, M.J.; Kim, J.S. Effect of conjugated linoleic acids and omega-3 fatty acids with or without resistance training on muscle mass in high-fat diet-fed middle-aged mice. Exp. Physiol. 2017, 102, 1500–1512. [Google Scholar] [CrossRef]
- Shen, W.; Chuang, C.C.; Martinez, K.; Reid, T.; Brown, J.M.; Xi, L.; Hixson, L.; Hopkins, R.; Starnes, J.; McIntosh, M. Conjugated linoleic acid reduces adiposity and increases markers of browning and inflammation in white adipose tissue of mice. J. Lipid Res. 2013, 54, 909–922. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.; McIntosh, M.K. Nutrient Regulation: Conjugated Linoleic Acid’s Inflammatory and Browning Properties in Adipose Tissue. Annu. Rev. Nutr. 2016, 36, 183–210. [Google Scholar] [CrossRef] [PubMed]
- West, D.B.; Delany, J.P.; Camet, P.M.; Blohm, F.; Truett, A.A.; Scimeca, J. Effects of conjugated linoleic acid on body fat and energy metabolism in the mouse. Am. J. Physiol. 1998, 275, R667–R672. [Google Scholar] [CrossRef] [PubMed]
- Vyas, D.; Kadegowda, A.K.; Erdman, R.A. Dietary conjugated linoleic Acid and hepatic steatosis: Species-specific effects on liver and adipose lipid metabolism and gene expression. J. Nutr. Metab. 2012, 2012, 932928. [Google Scholar] [CrossRef] [PubMed]
- Della Casa, L.; Rossi, E.; Romanelli, C.; Gibellini, L.; Iannone, A. Effect of diets supplemented with different conjugated linoleic acid (CLA) isomers on protein expression in C57/BL6 mice. Genes Nutr. 2016, 11, 26. [Google Scholar] [CrossRef]
- Xiao, L.; Wang, Y.; Zhou, Y.; Sun, Y.; Sun, W.; Wang, L.; Zhou, C.; Zhou, J.; Zhang, J. Identification of a novel human cancer/testis gene MAEL that is regulated by DNA methylation. Mol. Biol. Rep. 2010, 37, 2355–2360. [Google Scholar] [CrossRef]
- Crespo-Yanez, X.; Aguilar-Gurrieri, C.; Jacomin, A.C.; Journet, A.; Mortier, M.; Taillebourg, E.; Soleilhac, E.; Weissenhorn, W.; Fauvarque, M.O. CHMP1B is a target of USP8/UBPY regulated by ubiquitin during endocytosis. PLoS Genet. 2018, 14, e1007456. [Google Scholar] [CrossRef]
- McCullough, J.; Clippinger, A.K.; Talledge, N.; Skowyra, M.L.; Saunders, M.G.; Naismith, T.V.; Colf, L.A.; Afonine, P.; Arthur, C.; Sundquist, W.I.; et al. Structure and membrane remodeling activity of ESCRT-III helical polymers. Science 2015, 350, 1548–1551. [Google Scholar] [CrossRef]
- Schmidt, O.; Teis, D. The ESCRT machinery. Curr. Biol. 2012, 22, R116–R120. [Google Scholar] [CrossRef]
- Chang, C.L.; Weigel, A.V.; Ioannou, M.S.; Pasolli, H.A.; Xu, C.S.; Peale, D.R.; Shtengel, G.; Freeman, M.; Hess, H.F.; Blackstone, C.; et al. Spastin tethers lipid droplets to peroxisomes and directs fatty acid trafficking through ESCRT-III. J. Cell Biol. 2019, 218, 2583–2599. [Google Scholar] [CrossRef]
- Liu, Y.; Hu, W.; Murakawa, Y.; Yin, J.; Wang, G.; Landthaler, M.; Yan, J. Cold-induced RNA-binding proteins regulate circadian gene expression by controlling alternative polyadenylation. Sci. Rep. 2013, 3, 2054. [Google Scholar] [CrossRef] [PubMed]
- Cataldi, M.; Arnaldi, D.; Tucci, V.; De Carli, F.; Patti, G.; Napoli, F.; Pace, M.; Maghnie, M.; Nobili, L. Sleep disorders in Prader-Willi syndrome, evidence from animal models and humans. Sleep Med. Rev. 2021, 57, 101432. [Google Scholar] [CrossRef]
- Turroni, F.; Marchesi, J.R.; Foroni, E.; Gueimonde, M.; Shanahan, F.; Margolles, A.; van Sinderen, D.; Ventura, M. Microbiomic analysis of the bifidobacterial population in the human distal gut. ISME J. 2009, 3, 745–751. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.R.; Xu, Y.; Cassidy, R.M.; Lu, Y.; Yang, Y.; Tian, J.; Li, D.P.; Van Drunen, R.; Ribas-Latre, A.; Cai, Z.L.; et al. Paraventricular hypothalamus mediates diurnal rhythm of metabolism. Nat. Commun. 2020, 11, 3794. [Google Scholar] [CrossRef]
- Walton, J.C.; McNeill, J.K.T.; Oliver, K.A.; Albers, H.E. Temporal Regulation of GABAA Receptor Subunit Expression: Role in Synaptic and Extrasynaptic Communication in the Suprachiasmatic Nucleus. eNeuro 2017, 4. [Google Scholar] [CrossRef]
- Rice, L.J.; Lagopoulos, J.; Brammer, M.; Einfeld, S.L. Reduced gamma-aminobutyric acid is associated with emotional and behavioral problems in Prader-Willi syndrome. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2016, 171, 1041–1048. [Google Scholar] [CrossRef] [PubMed]
- Egawa, K.; Saitoh, S.; Asahina, N.; Shiraishi, H. Variance in the pathophysiological impact of the hemizygosity of gamma-aminobutyric acid type A receptor subunit genes between Prader-Willi syndrome and Angelman syndrome. Brain Dev. 2021, 43, 521–527. [Google Scholar] [CrossRef] [PubMed]
- Ding, F.; Li, H.H.; Li, J.; Myers, R.M.; Francke, U. Neonatal maternal deprivation response and developmental changes in gene expression revealed by hypothalamic gene expression profiling in mice. PLoS ONE 2010, 5, e9402. [Google Scholar] [CrossRef]
- Khor, E.C.; Fanshawe, B.; Qi, Y.; Zolotukhin, S.; Kulkarni, R.N.; Enriquez, R.F.; Purtell, L.; Lee, N.J.; Wee, N.K.; Croucher, P.I.; et al. Prader-Willi Critical Region, a Non-Translated, Imprinted Central Regulator of Bone Mass: Possible Role in Skeletal Abnormalities in Prader-Willi Syndrome. PLoS ONE 2016, 11, e0148155. [Google Scholar] [CrossRef]
- Zahova, S.K.; Humby, T.; Davies, J.R.; Morgan, J.E.; Isles, A.R. Comparison of mouse models reveals a molecular distinction between psychotic illness in PWS and schizophrenia. Transl. Psychiatry 2021, 11, 433. [Google Scholar] [CrossRef]
- Peng, Y.; Tan, Q.; Afhami, S.; Deehan, E.C.; Liang, S.; Gantz, M.; Triador, L.; Madsen, K.L.; Walter, J.; Tun, H.M.; et al. The Gut Microbiota Profile in Children with Prader-Willi Syndrome. Genes 2020, 11, 904. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Yin, A.; Li, H.; Wang, R.; Wu, G.; Shen, J.; Zhang, M.; Wang, L.; Hou, Y.; Ouyang, H.; et al. Dietary Modulation of Gut Microbiota Contributes to Alleviation of Both Genetic and Simple Obesity in Children. EBioMedicine 2015, 2, 968–984. [Google Scholar] [CrossRef] [PubMed]
- Dahl, W.J.; Auger, J.; Alyousif, Z.; Miller, J.L.; Tompkins, T.A. Adults with Prader-Willi syndrome exhibit a unique microbiota profile. BMC Res. Notes 2021, 14, 51. [Google Scholar] [CrossRef] [PubMed]
- Biassoni, R.; Di Marco, E.; Squillario, M.; Barla, A.; Piccolo, G.; Ugolotti, E.; Gatti, C.; Minuto, N.; Patti, G.; Maghnie, M.; et al. Gut Microbiota in T1DM-Onset Pediatric Patients: Machine-Learning Algorithms to Classify Microorganisms as Disease Linked. J. Clin. Endocrinol. Metab. 2020, 105, e3114–e3126. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | Treatment 20% Fat Diet | Treatment 20% Fat Diet with CLA |
|---|---|---|
| WT | 8 | 8 |
| PWS (Snord116m+/p−) | 8 | 8 |
| PWS-KO (Snord116m−/p−) | 7 | 8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Knott, B.; Kocher, M.A.; Paz, H.A.; Hamm, S.E.; Fink, W.; Mason, J.; Grange, R.W.; Wankhade, U.D.; Good, D.J. Dietary Conjugated Linoleic Acid Reduces Body Weight and Fat in Snord116m+/p− and Snord116m−/p− Mouse Models of Prader–Willi Syndrome. Nutrients 2022, 14, 860. https://doi.org/10.3390/nu14040860
Knott B, Kocher MA, Paz HA, Hamm SE, Fink W, Mason J, Grange RW, Wankhade UD, Good DJ. Dietary Conjugated Linoleic Acid Reduces Body Weight and Fat in Snord116m+/p− and Snord116m−/p− Mouse Models of Prader–Willi Syndrome. Nutrients. 2022; 14(4):860. https://doi.org/10.3390/nu14040860
Chicago/Turabian StyleKnott, Brittney, Matthew A. Kocher, Henry A. Paz, Shelby E. Hamm, William Fink, Jordan Mason, Robert W. Grange, Umesh D. Wankhade, and Deborah J. Good. 2022. "Dietary Conjugated Linoleic Acid Reduces Body Weight and Fat in Snord116m+/p− and Snord116m−/p− Mouse Models of Prader–Willi Syndrome" Nutrients 14, no. 4: 860. https://doi.org/10.3390/nu14040860
APA StyleKnott, B., Kocher, M. A., Paz, H. A., Hamm, S. E., Fink, W., Mason, J., Grange, R. W., Wankhade, U. D., & Good, D. J. (2022). Dietary Conjugated Linoleic Acid Reduces Body Weight and Fat in Snord116m+/p− and Snord116m−/p− Mouse Models of Prader–Willi Syndrome. Nutrients, 14(4), 860. https://doi.org/10.3390/nu14040860