
Neurite Outgrowth-Promoting Compounds from Cockscomb Hydrolysate

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Purification of Fractions Showing Neurite Outgrowth-Promoting Activity from Cockscomb Hydrolysate
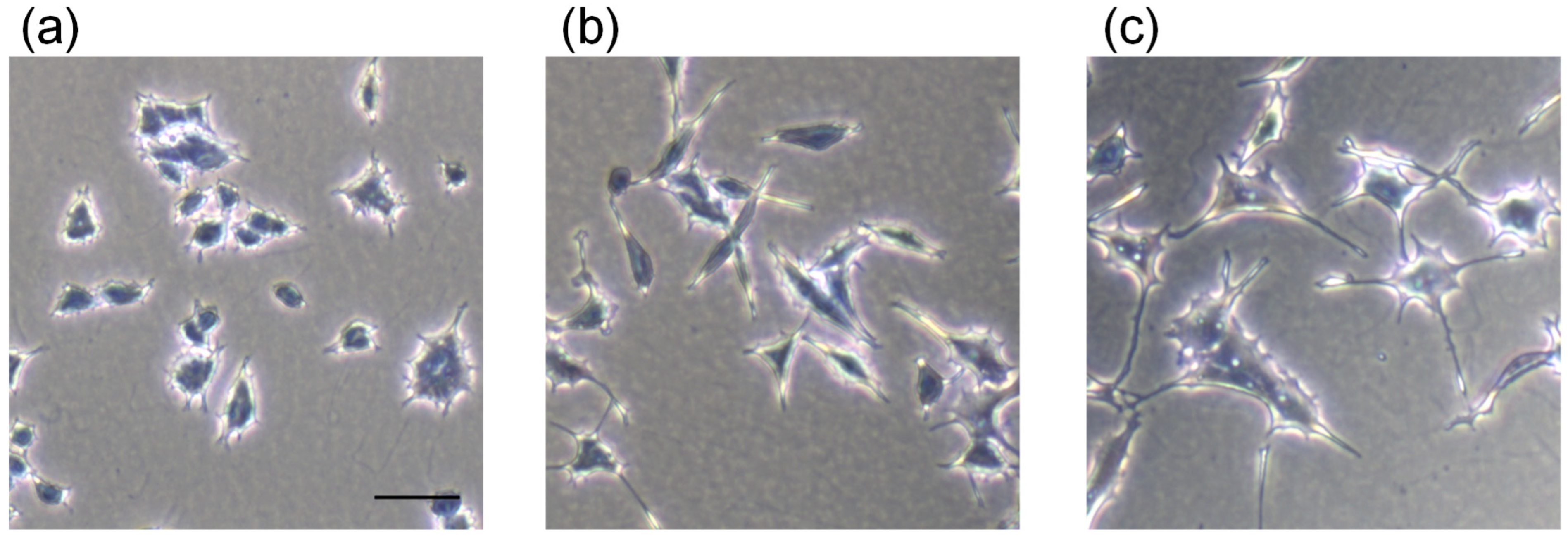
2.3. Neurite Outgrowth-Promoting Activity
3. Results and Discussion
3.1. Purification of Neurite Outgrowth-Promoting Compounds from Cockscomb Hydrolysate
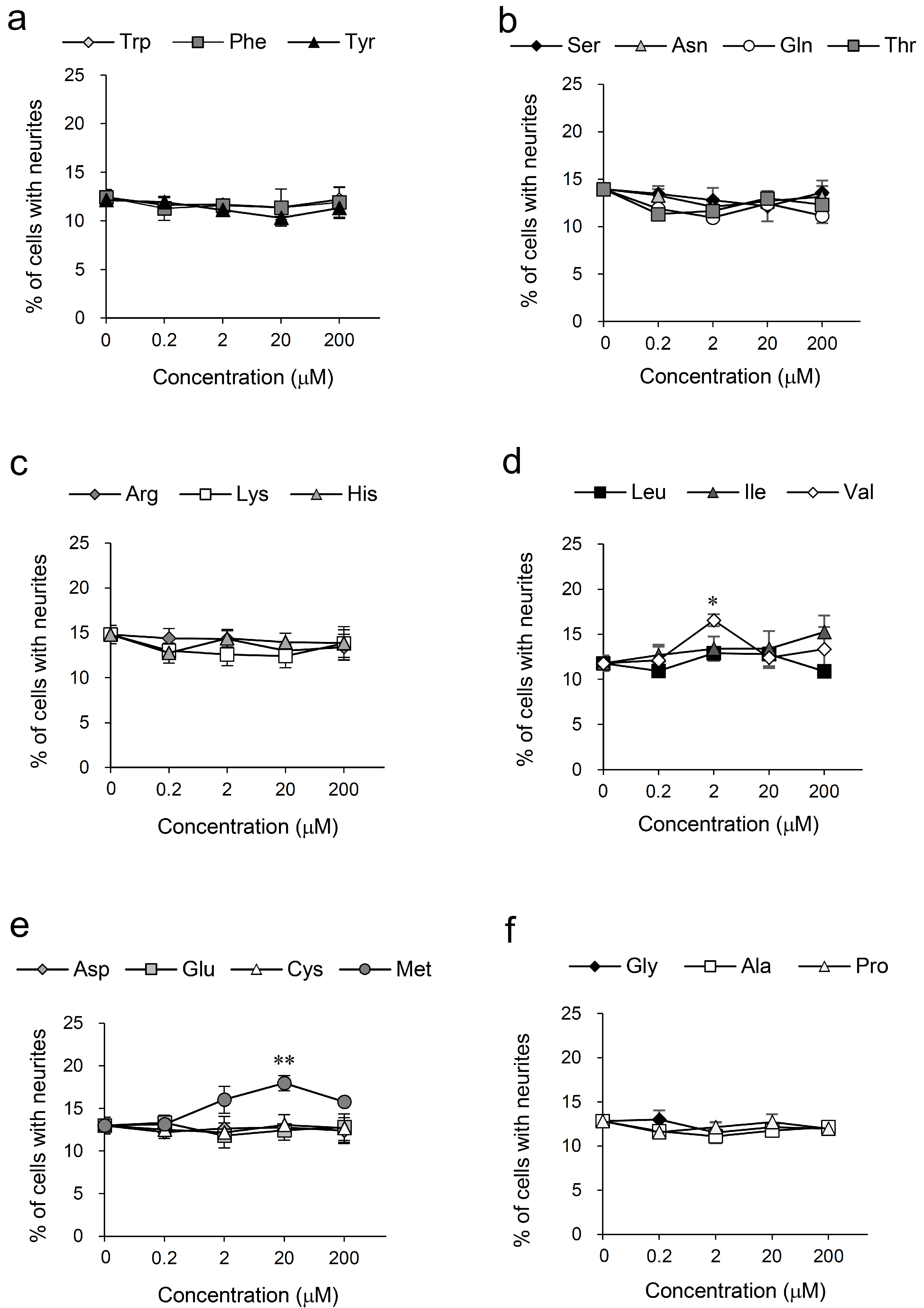
3.2. Screening for Amino Acids Having Neurite Outgrowth-Promoting Activity
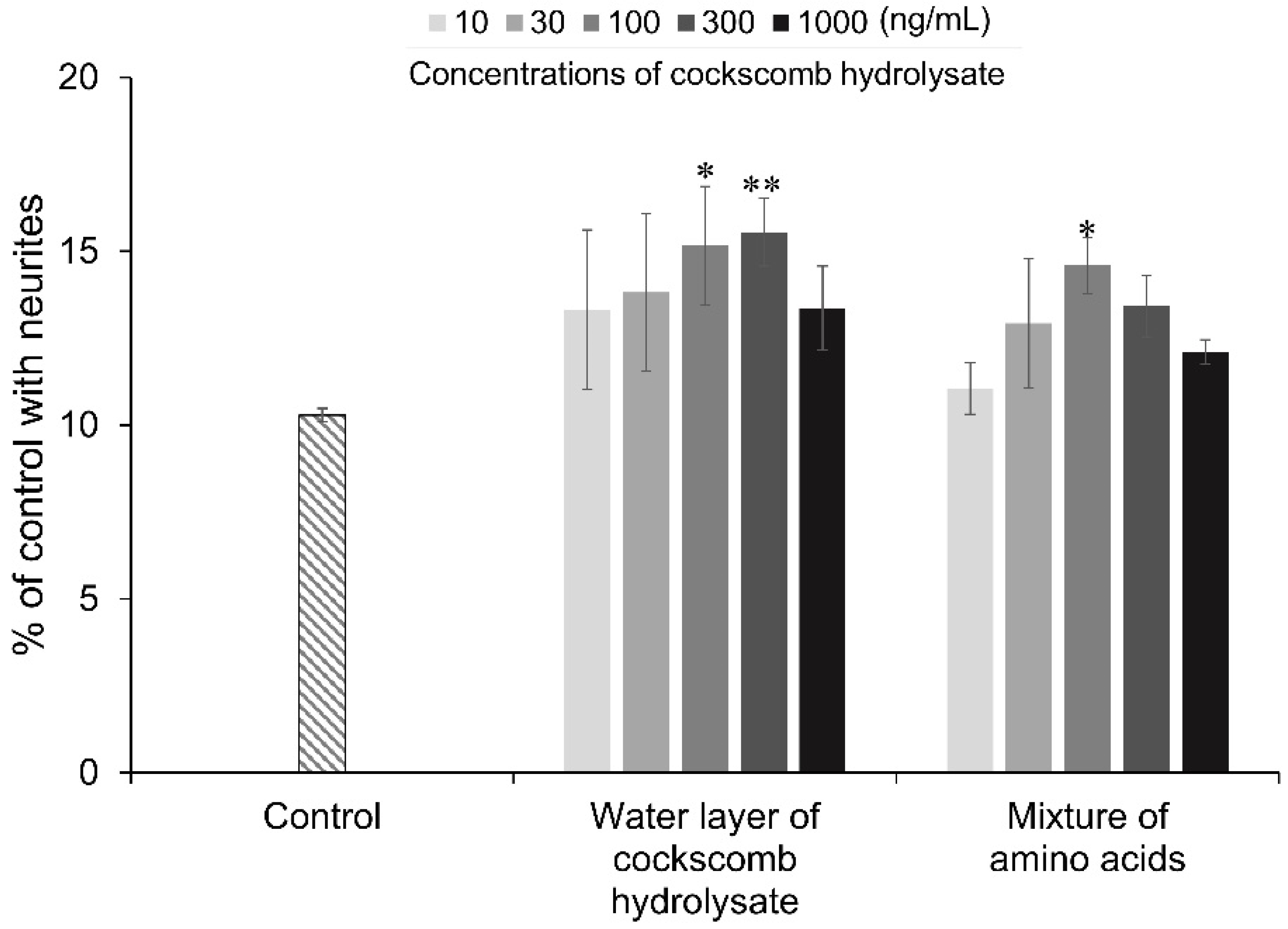
3.3. Neurite Outgrowth-Promoting Activities of Mixtures of Amino Acids
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kanazawa, K. Research and Development for bioavailable functional foods in food science. Food Sci. Technol. Res. 2018, 24, 183–191. [Google Scholar] [CrossRef]
- Lee, H.W.; Park, Y.S.; Jung, J.S.; Shin, W.S. Chitosan oligosaccharides, dp 2-8, have prebiotic effect on the Bifidobacterium bifidium and Lactobacillus sp. Anaerobe 2002, 8, 319–324. [Google Scholar] [CrossRef]
- Hirano, S.; Iwata, M.; Yamanaka, K.; Tanaka, H.; Toda, T.; Inui, H. Enhancement of serum lysozyme activity by injecting a mixture of chitosan oligosaccharides intravenously in rabbits. Agric. Biol. Chem. 1991, 55, 2623–2625. [Google Scholar]
- Sakai, S.; Akiyama, H.; Sato, Y.; Yoshioka, Y.; Linhardt, R.J.; Goda, Y.; Maitani, T.; Toida, T. Chondroitin sulfate intake inhibits the IgE-mediated allergic response by down-regulating Th2 responses in mice. J. Biol. Chem. 2006, 281, 19872–19880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsufuji, H.; Matsui, T.; Seki, E.; Osajima, K.; Nakashima, M.; Osajima, Y. Angiotensin I-converting enzyme inhibitory peptides in an alkaline protease hydrolyzate derived from sardine muscle. Biosci. Biotechnol. Biochem. 1994, 58, 2244–2245. [Google Scholar] [CrossRef] [PubMed]
- Matsufuji, H.; Matsui, T.; Ohshige, S.; Kawasaki, T.; Osajima, K.; Osajima, Y. Antihypertensive effects of angiotensin fragments in SHR. Biosci. Biotechnol. Biochem. 1995, 59, 1398–1401. [Google Scholar] [CrossRef] [Green Version]
- Manso, M.A.; Miguel, M.; Even, J.; Hernández, R.; Aleixandre, A.; López-Fandiño, R. Effect of the long-term intake of an egg white hydrolysate on the oxidative status and blood lipid profile of spontaneously hypertensive rats. Food Chem. 2008, 109, 361–367. [Google Scholar] [CrossRef]
- Morimatsu, F.; Ito, M.; Budijanto, S.; Watanabe, I.; Furukawa, Y.; Kimura, S. Plasma cholesterol-suppressing effect of papain-hydrolyzed pork meat in rats fed hypercholesterolemic diet. J. Nutr. Sci. Vitaminol. 1996, 43, 145–153. [Google Scholar] [CrossRef] [Green Version]
- Neuman, M.G.; Nanau, R.M.; Oruña-Sanchez, L.; Coto, G. Hyaluronic acid and wound healing. J. Pharm. Pharm. Sci. 2015, 18, 53–60. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.; Zhang, H.; Wu, H.; Wang, C.; Shi, X.; Xie, J.; He, J.; Yang, J. Cytoprotective effect of hyaluronic acid and hydroxypropyl methylcellulose against DNA damage induced by thimerosal in Chang conjunctival cells. Graefes Arch. Clin. Exp. Ophthalmol. 2012, 250, 1459–1466. [Google Scholar] [CrossRef]
- Torigoe, K.; Tanaka, H.F.; Ohkochi, H.; Miyasaka, M.; Yamanokuchi, H.; Yoshida, K.; Yoshida, T. Hyaluronan tetrasaccharide promotes regeneration of peripheral nerve: In vivo analysis by film model method. Brain Res. 2011, 1385, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Koyama, Y.; Nomura, Y. Effects of collagen peptide ingestion on UV-B-induced skin damage. Biosci. Biotechnol. Biochem. 2009, 73, 930–932. [Google Scholar] [CrossRef] [PubMed]
- Ogura, M.; Takabe, W.; Yagi, M.; Wakayama, S.; Yonei, Y. Study for investigation of symptomatic improvement and safety of the ingestion of rooster comb degradation product containing low-molecular hyaluronic acid (INJUV) in individuals with knee and lower back pain; open-label trial with no control group. Glycative Stress Res. 2018, 5, 55–67. [Google Scholar]
- Zhou, X.; Tai, A.; Yamamoto, I. Enhancement of neurite outgrowth in PC12 cells stimulated with cyclic AMP and NGF by 6-acylated ascorbic acid 2-O-α-glucosides (6-Acyl-AA-2G), novel lipophilic ascorbate derivatives. Biol. Pharm. Bull. 2003, 26, 341–346. [Google Scholar] [CrossRef] [Green Version]
- Kamata, Y.; Shiraga, H.; Tai, A.; Kawamoto, Y.; Gohda, E. Induction of neurite outgrowth in PC12 cells by the medium-chain fatty acid octanoic acid. Neuroscience 2007, 146, 1073–1081. [Google Scholar] [CrossRef]
- Duan, L.; Hope, J.M.; Guo, S.; Ong, Q.; François, A.; Kaplan, L.; Scherrer, G.; Cui, B. Optical activation of trkA signaling. ACS Synth. Biol. 2018, 7, 1685–1693. [Google Scholar] [CrossRef] [Green Version]
- Mitra, S.; Behbahani, H.; Eriksdotter, M. Innovative therapy for Alzheimer’s disease-with focus on biodelivery of NGF. Flont. Neurosci. 2019, 13, 38. [Google Scholar] [CrossRef] [Green Version]
- Greene, L.A.; Tischler, A.S. Establishment of a noradrenergic clonal line of rat adrenal pheochromocytoma cells which respond to nerve growth factor. Proc. Natl. Acad. Sci. USA 1976, 73, 2424–2428. [Google Scholar] [CrossRef] [Green Version]
- Vaudry, D.; Stork, P.J.; Lazarovici, P.; Eiden, L.E. Signaling pathways for PC12 cell differentiation: Making the right connections. Science 2002, 296, 1648–1649. [Google Scholar] [CrossRef]
- Stessin, A.M.; Zippin, J.H.; Kamenetsky, M.; Hess, K.C.; Hess, K.C.; Levin, L.R. Soluble adenylyl cyclase mediates nerve growth factor-induced activation of Rap1. J. Biol. Chem. 2006, 281, 17253–17258. [Google Scholar] [CrossRef] [Green Version]
- Blomstrand, E.; Saltin, B. BCAA intake affects protein metabolism in muscle after but not during exercise in humans. Am. J. Physiol. Endocrinol. Metab. 2001, 281, E365–E374. [Google Scholar] [CrossRef] [PubMed]
- Hiroshige, K.; Sonta, T.; Suda, T.; Kanegae, K.; Ohtani, A. Oral supplementation of branched-chain amino acid improves nutritional status in elderly patients on chronic haemodialysis. Nephrol. Dial. Transplant. 2001, 16, 1856–1862. [Google Scholar] [CrossRef] [PubMed]
- Gluud, L.L.; Dam, G.; Les, I.; Marchesini, G.; Borre, M.; Aagaard, N.K.; Vilstrup, H. Branched-chain amino acids for people with hepatic encephalopathy. Cochrane Database Syst. Rev. 2017, 5, CD001939. [Google Scholar] [CrossRef] [PubMed]
- González-Domínguez, R.; García-Barrera, T.; Gómez-Ariza, J.L. Metabolite profiling for the identification of altered metabolic pathways in Alzheimer’s disease. J. Pharm. Biomed. Anal. 2015, 107, 75–81. [Google Scholar] [CrossRef]
- Parlesak, A.; Bode, C.; Bode, J.C. Free methionine supplementation limits alcohol-induced liver damage in rats. Alcohol. Clin. Exp. Res. 1998, 22, 352–358. [Google Scholar] [CrossRef]
- Van Brummelen, R.; du Toit, D. L-methionine as immune supportive supplement: A clinical evaluation. Amino Acids. 2007, 33, 157–163. [Google Scholar] [CrossRef]
- Li, Q.; Cui, J.; Fang, C.; Zhang, X.; Li, L. S-adenosylmethionine administration attenuates low brain-derived neurotrophic factor expression induced by chronic cerebrovascular hypoperfusion or beta amyloid treatment. Neurosci. Bull. 2016, 32, 153–161. [Google Scholar] [CrossRef] [Green Version]
- Wortmann, M. Dementia: A global health priority—Highlights from an ADI and World Health Organization report. Alzheimer’s Res. Ther. 2012, 4, 40. [Google Scholar] [CrossRef] [Green Version]
- Zheng, C.; Geetha, T.; Gearing, M.; Babu, J.R. Amyloid β-abrogated TrkA ubiquitination in PC12 cells analogous to Alzheimer’s disease. J. Neurochem. 2015, 133, 919–925. [Google Scholar] [CrossRef]
- Kristiansen, M.; Ham, J. Programmed cell death during neuronal development: The sympathetic neuron model. Cell Death Differ. 2014, 21, 1025–1035. [Google Scholar] [CrossRef] [Green Version]
- Fischer, W.; Wictorin, K.; Björklund, A.; Williams, L.R.; Varon, S.; Gage, F.H. Amelioration of cholinergic neuron atrophy and spatial memory impairment in aged rats by nerve growth factor. Nature 1987, 329, 65–68. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, R.A.; O’Kane, R.L.; Simpson, I.A.; Viña, J.R. Structure of the blood-brain barrier and its role in the transport of amino acids. J. Nutr. 2006, 136, 218S–226S. [Google Scholar] [CrossRef] [PubMed]
- Poduslo, J.F.; Curran, G.L. Permeability at the blood-brain and blood-nerve barriers of the neurotrophic factors: NGF, CNTF, NT-3, BDNF. Mol. Brain Res. 1996, 36, 280–286. [Google Scholar] [CrossRef]
- Connor, B.; Dragunow, M. The role of neuronal growth factors in neurodegenerative disorders of the human brain. Brain Res. Rev. 1998, 27, 1–39. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koga, T.; Tai, A. Neurite Outgrowth-Promoting Compounds from Cockscomb Hydrolysate. Nutrients 2022, 14, 1422. https://doi.org/10.3390/nu14071422
Koga T, Tai A. Neurite Outgrowth-Promoting Compounds from Cockscomb Hydrolysate. Nutrients. 2022; 14(7):1422. https://doi.org/10.3390/nu14071422
Chicago/Turabian StyleKoga, Takeru, and Akihiro Tai. 2022. "Neurite Outgrowth-Promoting Compounds from Cockscomb Hydrolysate" Nutrients 14, no. 7: 1422. https://doi.org/10.3390/nu14071422
APA StyleKoga, T., & Tai, A. (2022). Neurite Outgrowth-Promoting Compounds from Cockscomb Hydrolysate. Nutrients, 14(7), 1422. https://doi.org/10.3390/nu14071422