
Prenatal Choline Supplementation Alters One Carbon Metabolites in a Rat Model of Periconceptional Alcohol Exposure

 , ,
, , 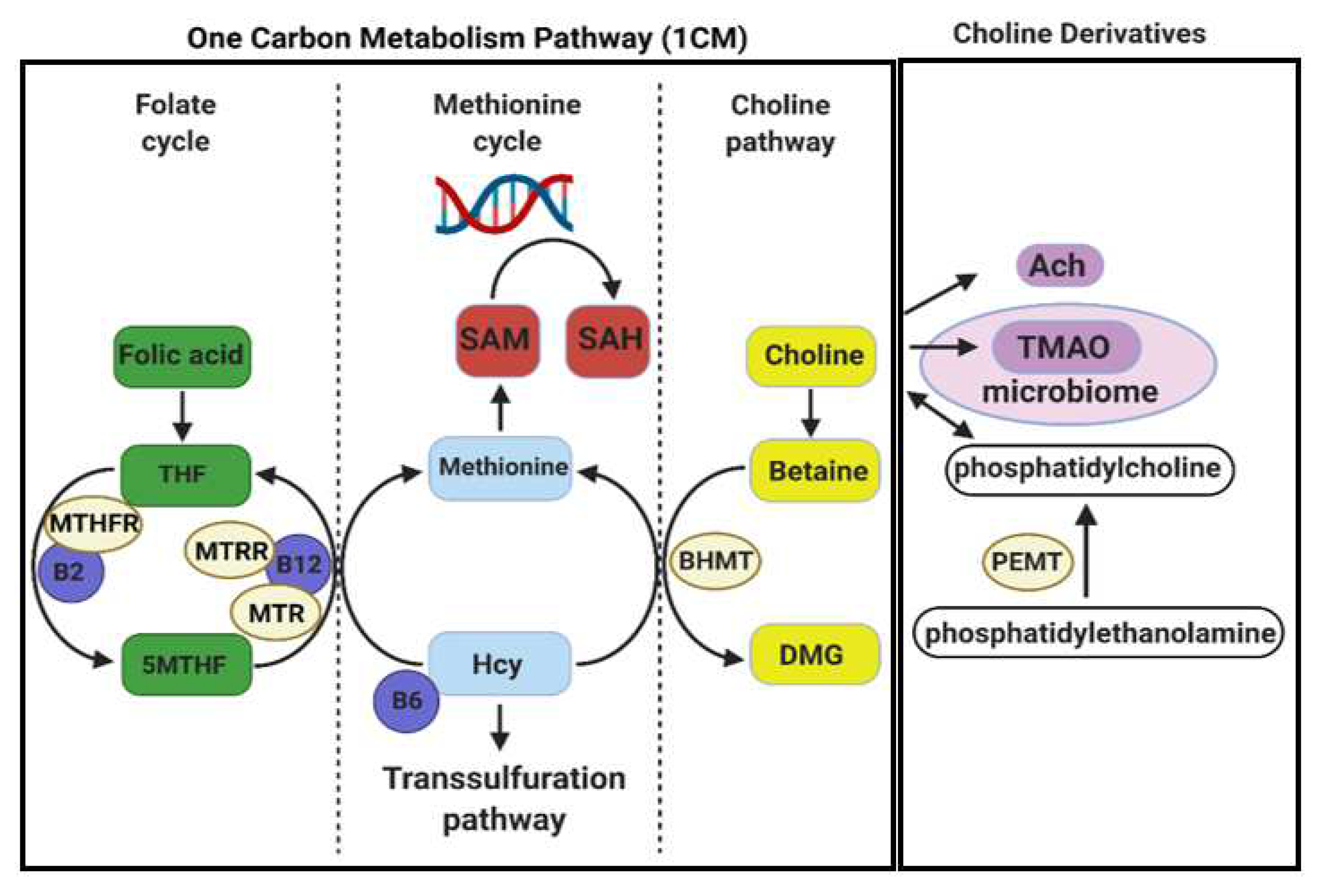{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Treatment and Diets
2.2. Blood and Tissue Collection
2.3. Liquid Chromatography-Tandem Mass Spectrometry (LC-MS/MS)
2.4. Quantitative PCR (qPCR) Analysis
2.5. Statistical Analyses
3. Results
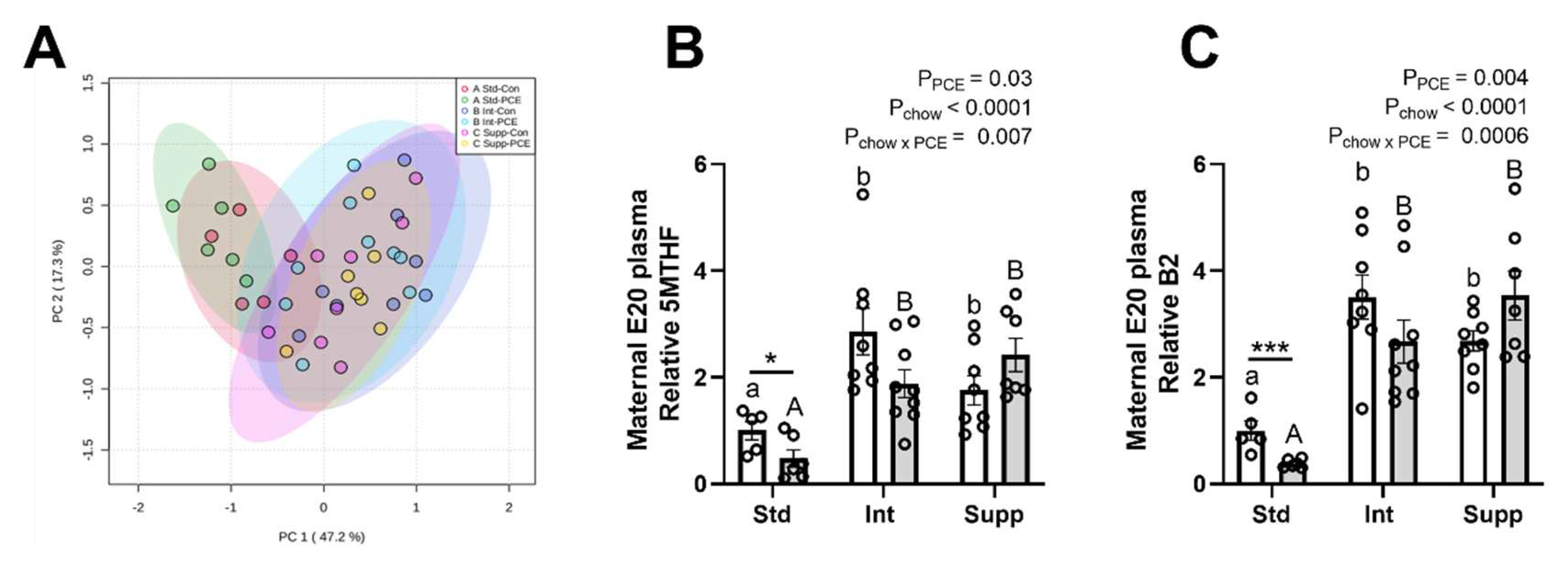
3.1. E20 Maternal Plasma: One-Carbon Metabolism Molecules
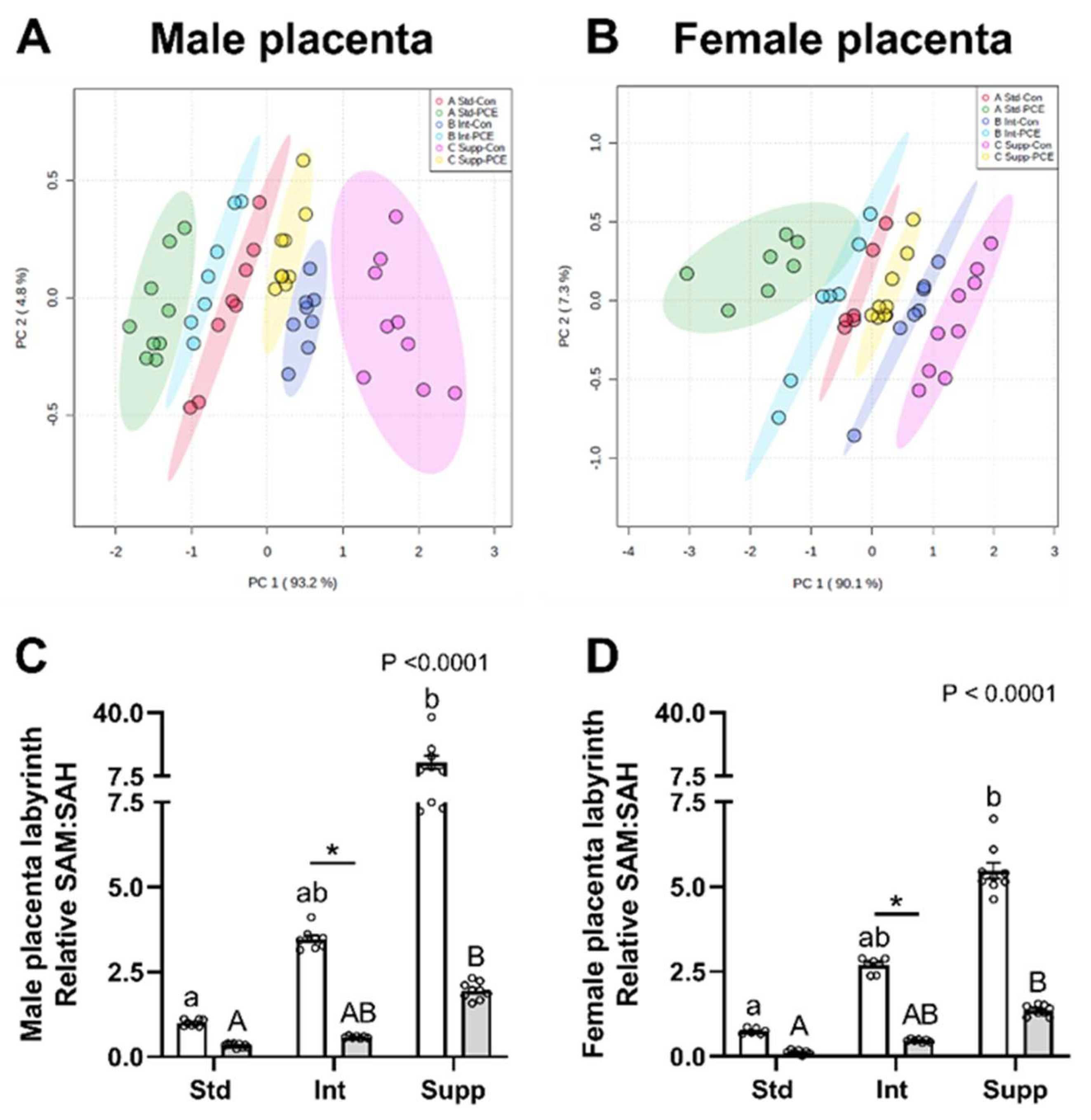
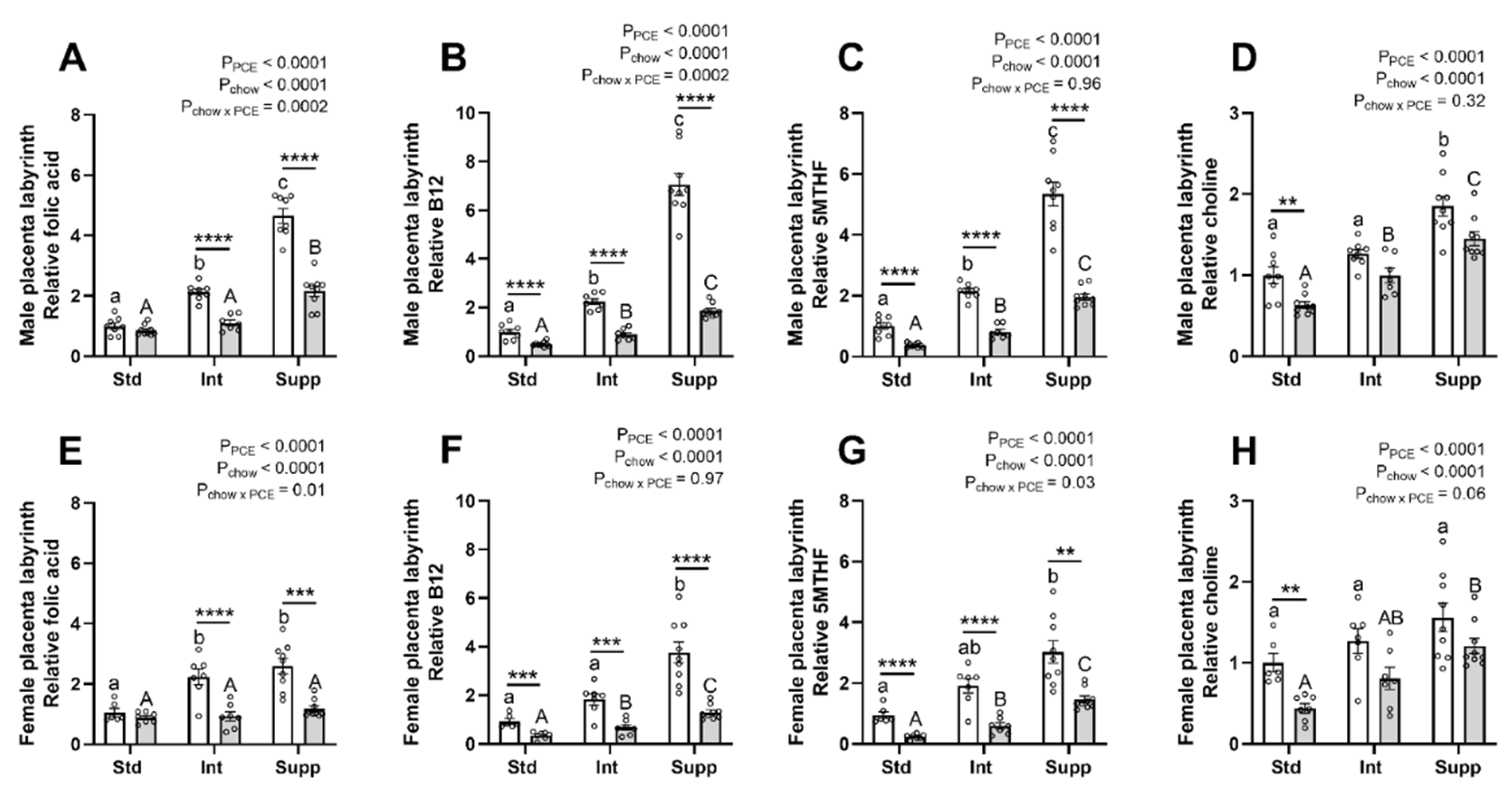
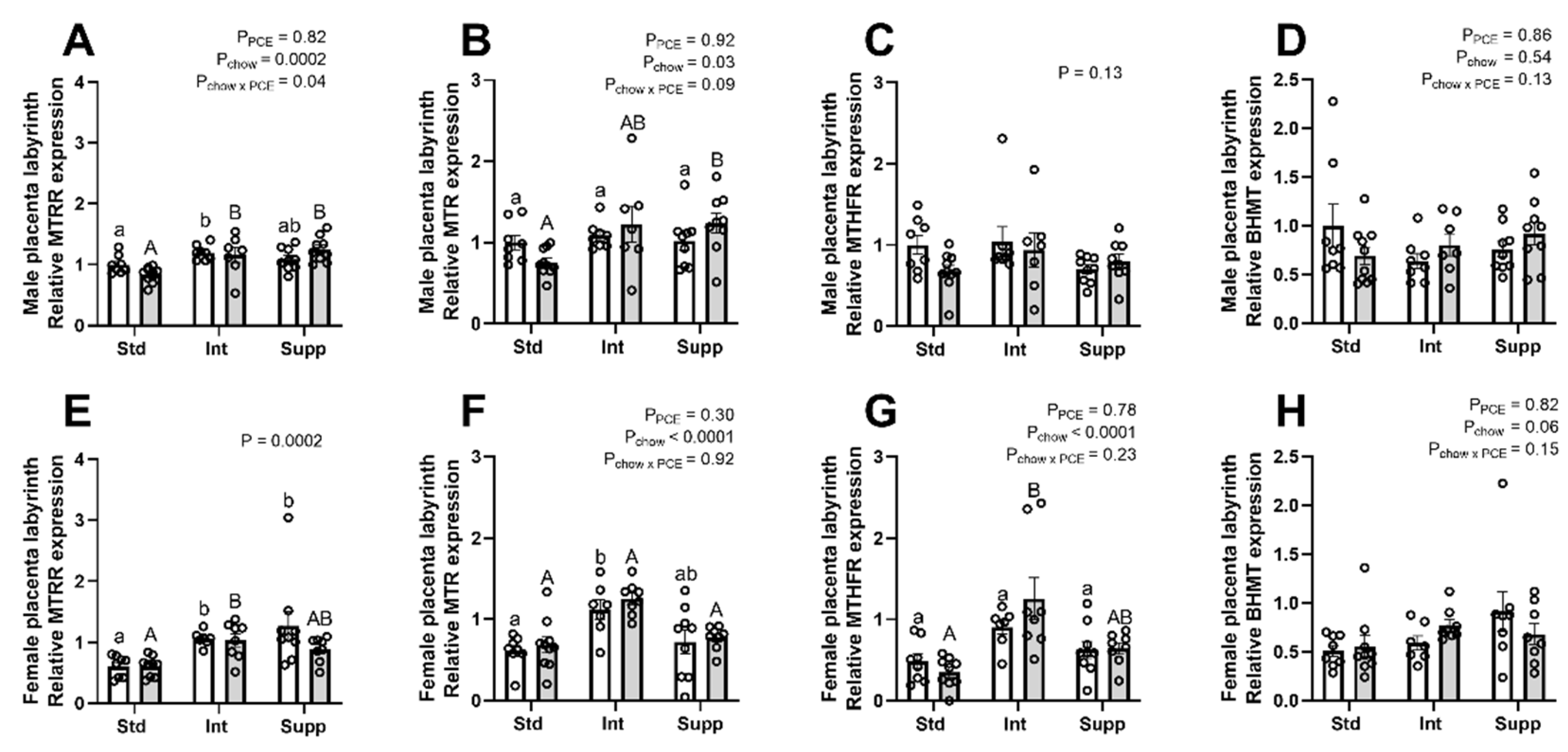
3.2. Placental Labyrinth: One-Carbon Metabolism Molecules
3.3. Placental Labyrinth: Expression of Genes Encoding One-Carbon Metabolism Pathway Enzymes
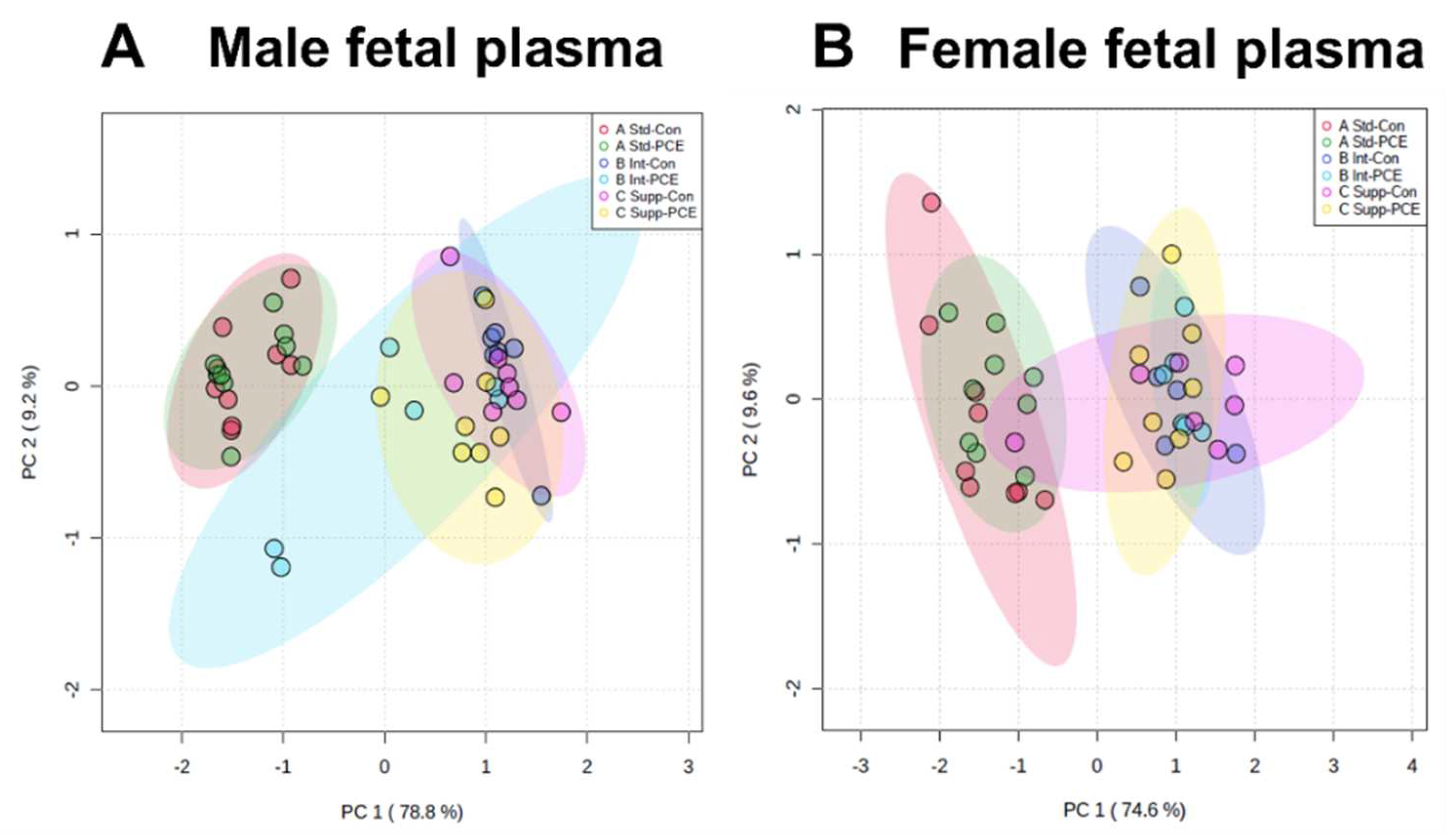
3.4. Fetal Plasma: One-Carbon Metabolism Molecules
3.5. Fetal Liver and Brain: One-Carbon Metabolism Molecules
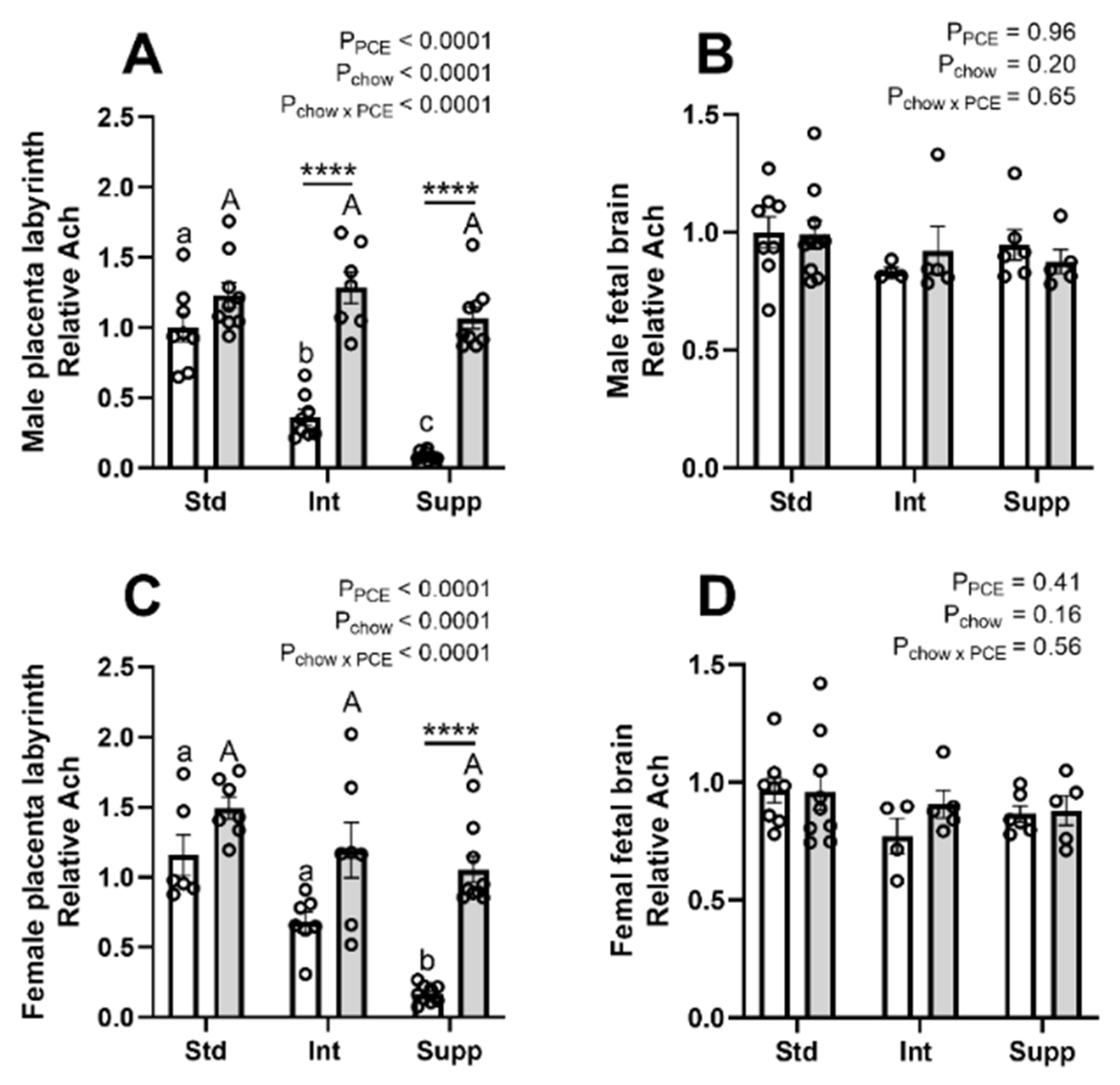
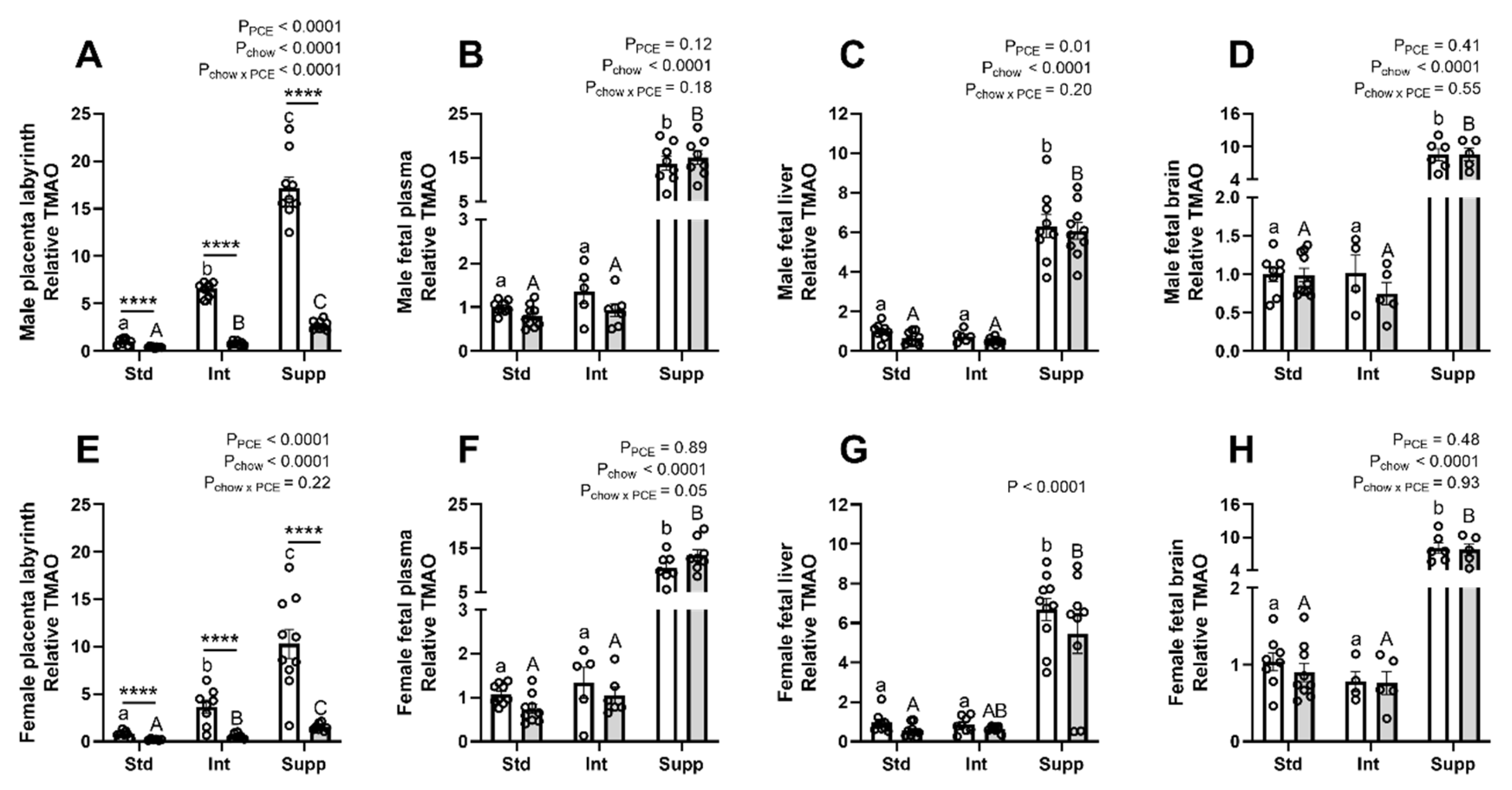
3.6. Choline Derivatives in Maternal, Placental, and Fetal Compartments: Acetylcholine and Trimethylamine-Oxide
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aliyu, M.H.; Wilson, R.E.; Zoorob, R.; Chakrabarty, S.; Alio, A.P.; Kirby, R.S.; Salihu, H.M. Alcohol consumption during pregnancy and the risk of early stillbirth among singletons. Alcohol 2008, 42, 369–374. [Google Scholar] [CrossRef] [PubMed]
- Patra, J.; Bakker, R.; Irving, H.; Jaddoe, V.W.; Malini, S.; Rehm, J. Dose-response relationship between alcohol consumption before and during pregnancy and the risks of low birthweight, preterm birth and small for gestational age (SGA)-a systematic review and meta-analyses. BJOG 2011, 118, 1411–1421. [Google Scholar] [CrossRef] [PubMed]
- Kesmodel, U.; Wisborg, K.; Olsen, S.F.; Henriksen, T.B.; Secher, N.J. Moderate alcohol intake in pregnancy and the risk of spontaneous abortion. Alcohol Alcohol. 2002, 37, 87–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- May, P.A.; Chambers, C.D.; Kalberg, W.O.; Zellner, J.; Feldman, H.; Buckley, D.; Kopald, D.; Hasken, J.M.; Xu, R.; Honerkamp-Smith, G.; et al. Prevalence of Fetal Alcohol Spectrum Disorders in 4 US Communities. JAMA 2018, 319, 474–482. [Google Scholar] [CrossRef] [PubMed]
- May, P.A.; Blankenship, J.; Marais, A.S.; Gossage, J.P.; Kalberg, W.O.; Barnard, R.; De Vries, M.; Robinson, L.K.; Adnams, C.M.; Buckley, D.; et al. Approaching the prevalence of the full spectrum of fetal alcohol spectrum disorders in a South African population-based study. Alcohol. Clin. Exp. Res. 2013, 37, 818–830. [Google Scholar] [CrossRef] [Green Version]
- Fitzpatrick, J.P.; Latimer, J.; Olson, H.C.; Carter, M.; Oscar, J.; Lucas, B.R.; Doney, R.; Salter, C.; Try, J.; Hawkes, G.; et al. Prevalence and profile of Neurodevelopment and Fetal Alcohol Spectrum Disorder (FASD) amongst Australian Aboriginal children living in remote communities. Res. Dev. Disabil. 2017, 65, 114–126. [Google Scholar] [CrossRef]
- Popova, S.; Lange, S.; Probst, C.; Gmel, G.; Rehm, J. Estimation of national, regional, and global prevalence of alcohol use during pregnancy and fetal alcohol syndrome: A systematic review and meta-analysis. Lancet Glob. Health 2017, 5, e290–e299. [Google Scholar] [CrossRef] [Green Version]
- O’Keeffe, L.M.; Kearney, P.M.; McCarthy, F.P.; Khashan, A.S.; Greene, R.A.; North, R.A.; Poston, L.; McCowan, L.M.; Baker, P.N.; Dekker, G.A.; et al. Prevalence and predictors of alcohol use during pregnancy: Findings from international multicentre cohort studies. BMJ Open 2015, 5, e006323. [Google Scholar] [CrossRef] [Green Version]
- Fleming, T.P.; Kwong, W.Y.; Porter, R.; Ursell, E.; Fesenko, I.; Wilkins, A.; Miller, D.J.; Watkins, A.J.; Eckert, J.J. The embryo and its future. Biol. Reprod. 2004, 71, 1046–1054. [Google Scholar] [CrossRef] [Green Version]
- Kaminen-Ahola, N. Fetal alcohol spectrum disorders: Genetic and epigenetic mechanisms. Prenat. Diagn. 2020, 40, 1189–1192. [Google Scholar] [CrossRef]
- Gutherz, O.R.; Deyssenroth, M.; Li, Q.; Hao, K.; Jacobson, J.L.; Chen, J.; Jacobson, S.W.; Carter, R.C. Potential roles of imprinted genes in the teratogenic effects of alcohol on the placenta, somatic growth, and the developing brain. Exp. Neurol. 2022, 347, 113919. [Google Scholar] [CrossRef]
- Zeisel, S.H. Nutrition in pregnancy: The argument for including a source of choline. Int. J. Womens Health 2013, 5, 193–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bekdash, R.A.; Zhang, C.; Sarkar, D.K. Gestational choline supplementation normalized fetal alcohol-induced alterations in histone modifications, DNA methylation, and proopiomelanocortin (POMC) gene expression in β-endorphin-producing POMC neurons of the hypothalamus. Alcohol. Clin. Exp. Res. 2013, 37, 1133–1142. [Google Scholar] [CrossRef] [Green Version]
- Idrus, N.M.; Breit, K.R.; Thomas, J.D. Dietary choline levels modify the effects of prenatal alcohol exposure in rats. Neurotoxicol. Teratol. 2017, 59, 43–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobson, S.W.; Carter, R.C.; Molteno, C.D.; Stanton, M.E.; Herbert, J.S.; Lindinger, N.M.; Lewis, C.E.; Dodge, N.C.; Hoyme, H.E.; Zeisel, S.H.; et al. Efficacy of Maternal Choline Supplementation during Pregnancy in Mitigating Adverse Effects of Prenatal Alcohol Exposure on Growth and Cognitive Function: A Randomized, Double-Blind, Placebo-Controlled Clinical Trial. Alcohol. Clin. Exp. Res. 2018, 42, 1327–1341. [Google Scholar] [CrossRef]
- Kable, J.A.; Coles, C.D.; Keen, C.L.; Uriu-Adams, J.Y.; Jones, K.L.; Yevtushok, L.; Kulikovsky, Y.; Zymak-Zakutnya, N.; Dubchak, I.; Akhmedzhanova, D.; et al. The Impact of Micronutrient Supplementation in Alcohol-Exposed Pregnancies on Reaction Time Responses of Preschoolers in Ukraine. Alcohol 2022, 99, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Akison, L.K.; Nitert, M.D.; Clifton, V.L.; Moritz, K.M.; Simmons, D.G. Review: Alterations in placental glycogen deposition in complicated pregnancies: Current preclinical and clinical evidence. Placenta 2017, 54, 52–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ernst, A.M.; Gimbel, B.A.; de Water, E.; Eckerle, J.K.; Radke, J.P.; Georgieff, M.K.; Wozniak, J.R. Prenatal and Postnatal Choline Supplementation in Fetal Alcohol Spectrum Disorder. Nutrients 2022, 14, 688. [Google Scholar] [CrossRef]
- Taesuwan, S.; McDougall, M.Q.; Malysheva, O.V.; Bender, E.; Nevins, J.E.H.; Devapatla, S.; Vidavalur, R.; Caudill, M.A.; Klatt, K.C. Choline metabolome response to prenatal choline supplementation across pregnancy: A randomized controlled trial. FASEB J. 2021, 35, e22063. [Google Scholar] [CrossRef]
- Gatarek, P.; Kaluzna-Czaplinska, J. Trimethylamine N-oxide (TMAO) in human health. EXCLI J. 2021, 20, 301–319. [Google Scholar] [CrossRef]
- Wen, Y.; Peng, L.; Xu, R.; Zang, N.; Huang, Q.; Zhong, M. Maternal serum trimethylamine-N-oxide is significantly increased in cases with established preeclampsia. Pregnancy Hypertens. 2019, 15, 114–117. [Google Scholar] [CrossRef] [PubMed]
- Gårdebjer, E.M.; Cuffe, J.S.; Pantaleon, M.; Wlodek, M.E.; Moritz, K.M. Periconceptional alcohol consumption causes fetal growth restriction and increases glycogen accumulation in the late gestation rat placenta. Placenta 2014, 35, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Kalisch-Smith, J.I.; Steane, S.E.; Simmons, D.G.; Pantaleon, M.; Anderson, S.T.; Akison, L.K.; Wlodek, M.E.; Moritz, K.M. Periconceptional alcohol exposure causes female-specific perturbations to trophoblast differentiation and placental formation in the rat. Development 2019, 146, dev172205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steane, S.E.; Fielding, A.M.; Kent, N.L.; Andersen, I.; Browne, D.J.; Tejo, E.N.; Gårdebjer, E.M.; Kalisch-Smith, J.I.; Sullivan, M.A.; Moritz, K.M.; et al. Maternal choline supplementation in a rat model of periconceptional alcohol exposure: Impacts on the fetus and placenta. Alcohol. Clin. Exp. Res. 2021, 45, 2130–2146. [Google Scholar] [CrossRef]
- Cuffe, J.S.; O’Sullivan, L.; Simmons, D.G.; Anderson, S.T.; Moritz, K.M. Maternal corticosterone exposure in the mouse has sex-specific effects on placental growth and mRNA expression. Endocrinology 2012, 153, 5500–5511. [Google Scholar] [CrossRef] [Green Version]
- Pang, Z.; Chong, J.; Zhou, G.; de Lima Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the gap between raw spectra and functional insights. Nucleic Acids Res. 2021, 49, W388–W396. [Google Scholar] [CrossRef]
- Lionaki, E.; Ploumi, C.; Tavernarakis, N. One-Carbon Metabolism: Pulling the Strings behind Aging and Neurodegeneration. Cells 2022, 11, 214. [Google Scholar] [CrossRef]
- Rosenzweig, A.; Blenis, J.; Gomes, A.P. Beyond the Warburg Effect: How Do Cancer Cells Regulate One-Carbon Metabolism? Front. Cell Dev. Biol. 2018, 6, 90. [Google Scholar] [CrossRef]
- Baumgartner, H.K.; Trinder, K.M.; Galimanis, C.E.; Post, A.; Phang, T.; Ross, R.G.; Winn, V.D. Characterization of choline transporters in the human placenta over gestation. Placenta 2015, 36, 1362–1369. [Google Scholar] [CrossRef] [Green Version]
- Obeid, R.; Munz, W.; Jäger, M.; Schmidt, W.; Herrmann, W. Biochemical indexes of the B vitamins in cord serum are predicted by maternal B vitamin status. Am. J. Clin. Nutr. 2005, 82, 133–139. [Google Scholar] [CrossRef]
- Kim, Y.I.; Miller, J.W.; da Costa, K.A.; Nadeau, M.; Smith, D.; Selhub, J.; Zeisel, S.H.; Mason, J.B. Severe folate deficiency causes secondary depletion of choline and phosphocholine in rat liver. J. Nutr. 1994, 124, 2197–2203. [Google Scholar] [CrossRef] [PubMed]
- Selhub, J.; Seyoum, E.; Pomfret, E.A.; Zeisel, S.H. Effects of choline deficiency and methotrexate treatment upon liver folate content and distribution. Cancer Res. 1991, 51, 16–21. [Google Scholar] [PubMed]
- Yan, J.; Jiang, X.; West, A.A.; Perry, C.A.; Malysheva, O.V.; Devapatla, S.; Pressman, E.; Vermeylen, F.; Stabler, S.P.; Allen, R.H.; et al. Maternal choline intake modulates maternal and fetal biomarkers of choline metabolism in humans. Am. J. Clin. Nutr. 2012, 95, 1060–1071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, J.; Jiang, X.; West, A.A.; Perry, C.A.; Malysheva, O.V.; Brenna, J.T.; Stabler, S.P.; Allen, R.H.; Gregory, J.F., 3rd; Caudill, M.A. Pregnancy alters choline dynamics: Results of a randomized trial using stable isotope methodology in pregnant and nonpregnant women. Am. J. Clin. Nutr. 2013, 98, 1459–1467. [Google Scholar] [CrossRef] [PubMed]
- Halsted, C.H.; Villanueva, J.A.; Devlin, A.M.; Chandler, C.J. Metabolic interactions of alcohol and folate. J. Nutr. 2002, 132, 2367s–2372s. [Google Scholar] [CrossRef] [Green Version]
- Subramanian, V.S.; Subramanya, S.B.; Ghosal, A.; Said, H.M. Chronic alcohol feeding inhibits physiological and molecular parameters of intestinal and renal riboflavin transport. Am. J. Physiol. Cell Physiol. 2013, 305, C539–C546. [Google Scholar] [CrossRef] [Green Version]
- Barak, A.J.; Beckenhauer, H.C.; Tuma, D.J. Methionine synthase. a possible prime site of the ethanolic lesion in liver. Alcohol 2002, 26, 65–67. [Google Scholar] [CrossRef]
- Cherukad, J.; Wainwright, V.; Watson, E.D. Spatial and temporal expression of folate-related transporters and metabolic enzymes during mouse placental development. Placenta 2012, 33, 440–448. [Google Scholar] [CrossRef]
- Pickell, L.; Li, D.; Brown, K.; Mikael, L.G.; Wang, X.L.; Wu, Q.; Luo, L.; Jerome-Majewska, L.; Rozen, R. Methylenetetrahydrofolate reductase deficiency and low dietary folate increase embryonic delay and placental abnormalities in mice. Birth Defects Res. A Clin. Mol. Teratol. 2009, 85, 531–541. [Google Scholar] [CrossRef]
- Lo, J.O.; Schabel, M.C.; Roberts, V.H.J.; Morgan, T.K.; Fei, S.S.; Gao, L.; Ray, K.G.; Lewandowski, K.S.; Newman, N.P.; Bohn, J.A.; et al. Effects of early daily alcohol exposure on placental function and fetal growth in a rhesus macaque model. Am. J. Obstet. Gynecol. 2022, 226, 130.e1–130.e11. [Google Scholar] [CrossRef]
- Kalisch-Smith, J.I.; Simmons, D.G.; Dickinson, H.; Moritz, K.M. Review: Sexual dimorphism in the formation, function and adaptation of the placenta. Placenta 2017, 54, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Meakin, A.S.; Cuffe, J.S.M.; Darby, J.R.T.; Morrison, J.L.; Clifton, V.L. Let’s Talk about Placental Sex, Baby: Understanding Mechanisms That Drive Female- and Male-Specific Fetal Growth and Developmental Outcomes. Int. J. Mol. Sci. 2021, 22, 6386. [Google Scholar] [CrossRef] [PubMed]
- Padmanabhan, N.; Rakoczy, J.; Kondratowicz, M.; Menelaou, K.; Blake, G.E.T.; Watson, E.D. Multigenerational analysis of sex-specific phenotypic differences at midgestation caused by abnormal folate metabolism. Environ. Epigenet. 2017, 3, dvx014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solanky, N.; Requena Jimenez, A.; D’Souza, S.W.; Sibley, C.P.; Glazier, J.D. Expression of folate transporters in human placenta and implications for homocysteine metabolism. Placenta 2010, 31, 134–143. [Google Scholar] [CrossRef]
- Batistel, F.; Alharthi, A.S.; Yambao, R.R.C.; Elolimy, A.A.; Pan, Y.X.; Parys, C.; Loor, J.J. Methionine Supply during Late-Gestation Triggers Offspring Sex-Specific Divergent Changes in Metabolic and Epigenetic Signatures in Bovine Placenta. J. Nutr. 2019, 149, 6–17. [Google Scholar] [CrossRef]
- Zeisel, S. Choline, other methyl-donors and epigenetics. Nutrients 2017, 9, 445. [Google Scholar] [CrossRef]
- Loke, Y.J.; Muggli, E.; Nguyen, L.; Ryan, J.; Saffery, R.; Elliott, E.J.; Halliday, J.; Craig, J.M. Time- and sex-dependent associations between prenatal alcohol exposure and placental global DNA methylation. Epigenomics 2018, 10, 981–991. [Google Scholar] [CrossRef]
- Martin, E.; Smeester, L.; Bommarito, P.A.; Grace, M.R.; Boggess, K.; Kuban, K.; Karagas, M.R.; Marsit, C.J.; O’Shea, T.M.; Fry, R.C. Sexual epigenetic dimorphism in the human placenta: Implications for susceptibility during the prenatal period. Epigenomics 2017, 9, 267–278. [Google Scholar] [CrossRef]
- Gårdebjer, E.M.; Anderson, S.T.; Pantaleon, M.; Wlodek, M.E.; Moritz, K.M. Maternal alcohol intake around the time of conception causes glucose intolerance and insulin insensitivity in rat offspring, which is exacerbated by a postnatal high-fat diet. FASEB J. 2015, 29, 2690–2701. [Google Scholar] [CrossRef] [Green Version]
- Burgess, D.J.; Dorey, E.S.; Gardebjer, E.M.; Bielefeldt-Ohmann, H.; Moritz, K.M.; Cuffe, J.S.M. Periconceptional ethanol exposure alters the stress axis in adult female but not male rat offspring. Stress 2019, 22, 347–357. [Google Scholar] [CrossRef]
- Burgess, D.J.; Lucia, D.; Cuffe, J.S.M.; Moritz, K.M. Periconceptional ethanol exposure alters hypothalamic-pituitary-adrenal axis function, signalling elements and associated behaviours in a rodent model. Psychoneuroendocrinology 2020, 122, 104901. [Google Scholar] [CrossRef] [PubMed]
- Derbyshire, E.; Obeid, R. Choline, Neurological Development and Brain Function: A Systematic Review Focusing on the First 1000 Days. Nutrients 2020, 12, 1731. [Google Scholar] [CrossRef] [PubMed]
- Sastry, B.V. Human placental cholinergic system. Biochem. Pharmacol. 1997, 53, 1577–1586. [Google Scholar] [CrossRef]
- Virdee, M.S.; Saini, N.; Kay, C.D.; Neilson, A.P.; Kwan, S.T.C.; Helfrich, K.K.; Mooney, S.M.; Smith, S.M. An enriched biosignature of gut microbiota-dependent metabolites characterizes maternal plasma in a mouse model of fetal alcohol spectrum disorder. Sci. Rep. 2021, 11, 248. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Steane, S.E.; Kumar, V.; Cuffe, J.S.M.; Moritz, K.M.; Akison, L.K. Prenatal Choline Supplementation Alters One Carbon Metabolites in a Rat Model of Periconceptional Alcohol Exposure. Nutrients 2022, 14, 1874. https://doi.org/10.3390/nu14091874
Steane SE, Kumar V, Cuffe JSM, Moritz KM, Akison LK. Prenatal Choline Supplementation Alters One Carbon Metabolites in a Rat Model of Periconceptional Alcohol Exposure. Nutrients. 2022; 14(9):1874. https://doi.org/10.3390/nu14091874
Chicago/Turabian StyleSteane, Sarah E., Vinod Kumar, James S. M. Cuffe, Karen M. Moritz, and Lisa K. Akison. 2022. "Prenatal Choline Supplementation Alters One Carbon Metabolites in a Rat Model of Periconceptional Alcohol Exposure" Nutrients 14, no. 9: 1874. https://doi.org/10.3390/nu14091874
APA StyleSteane, S. E., Kumar, V., Cuffe, J. S. M., Moritz, K. M., & Akison, L. K. (2022). Prenatal Choline Supplementation Alters One Carbon Metabolites in a Rat Model of Periconceptional Alcohol Exposure. Nutrients, 14(9), 1874. https://doi.org/10.3390/nu14091874