
Probiotic Fermented Goat’s and Sheep’s Milk: Effect of Type and Dose of Collagen on Survival of Four Strains of Probiotic Bacteria during Simulated In Vitro Digestion Conditions



Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Fermented Milk Manufacture
- Sheep’s milk.
- Sheep’s milk + 1.5% bovine collagen.
- Sheep’s milk + 3.0% bovine collagen.
- Sheep’s milk + 1.5% collagen protein hydrolysate.
- Sheep’s milk + 3.0% collagen protein hydrolysate.
- Goat’s milk.
- Goat’s milk + 1.5% bovine collagen.
- Goat’s milk + 3.0% bovine collagen.
- Goat’s milk + 1.5% collagen protein hydrolysate.
- Goat’s milk + 3.0% collagen protein hydrolysate.
2.3. In Vitro Digestion Process
2.4. Microbiological Analysis
2.5. Statistical Analysis
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ranadheera, C.S.; Vidanarachchi, J.K.; Rocha, R.S.; Cruz, A.G.; Ajlouni, S. Probiotic Delivery through Fermentation: Dairy vs. Non-Dairy Beverages. Fermentation 2017, 3, 67. [Google Scholar] [CrossRef] [Green Version]
- Melchior, S.; Marino, M.; D’Este, F.; Innocente, N.; Nicoli, M.C.; Calligaris, S. Effect of the formulation and structure of monoglyceride-based gels on the viability of probiotic Lactobacillus rhamnosus upon in vitro digestion. Food Funct. 2021, 12, 351. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, M.; Azcarate-Peril, M.A.; Barnard, A.; Benoit, V.; Grimaldi, R.; Guyonnet, D.; Holscher, H.D.; Hunter, K.; Manurung, S.; Obis, D.; et al. Shaping the Future of Probiotics and Prebiotics. Trends Microbiol. 2021, 29, 667–685. [Google Scholar] [CrossRef] [PubMed]
- Terpou, A.; Papadaki, A.; Lappa, I.K.; Kachrimanidou, V.; Bosnea, L.A.; Kopsahelis, N. Probiotics in Food Systems: Significance and Emerging Strategies towards Improved Viability and Delivery of Enhanced Beneficial Value. Nutrients 2019, 11, 1591. [Google Scholar] [CrossRef] [Green Version]
- European Pharmacopoeia Commission. 3053E General monograph on live biotherapeutic products. Eur. Pharmacop. 2019, 9, 7. [Google Scholar]
- Dale, H.F.; Rasmussen, S.H.; Asiller, Ö.Ö.; Lied, G.A. Probiotics in irritable bowel syndrome: An up-to-date systematic review. Nutrients 2019, 11, 2048. [Google Scholar] [CrossRef] [Green Version]
- Barichella, M.; Pacchetti, C.; Bolliri, C.; Cassani, E.; Lorio, L.; Pusani, C.; Pinelli, G.; Privitera, G.; Cesari, I.; Faierman, S.A.; et al. Probiotics and prebiotic fiber for constipation associated with Parkinson disease: An RCT. Neurology 2016, 87, 1274–1280. [Google Scholar] [CrossRef]
- Toejing, P.; Khampithum, N.; Sirilun, S.; Chaiyasut, C.; Lailerd, N. Influence of Lactobacillus paracasei HII01 Supplementation on Glycemia and Inflammatory biomarkers in Type 2 Diabetes: A randomized clinical trial. Foods 2021, 10, 1455. [Google Scholar] [CrossRef]
- Lv, T.; Ye, M.; Luo, F.; Hu, B.; Wang, A.; Chen, J.; Yan, J.; He, Z.; Chen, F.; Qian, C.; et al. Probiotics treatment improves cognitive impairment in patients and animals: A systematic review and meta-analysis. Neurosci. Biobehav. Rev. 2021, 120, 159–172. [Google Scholar] [CrossRef]
- Lopez-Santamarina, A.; Lamas, A.; del Carmen Mondragón, A.; Cardelle-Cobas, A.; Regal, P.; Rodriguez-Avila, J.A.; Miranda, J.M.; Franco, C.M.; Cepeda, A. Probiotic Effects against Virus Infections: New Weapons for an Old War. Foods 2021, 10, 130. [Google Scholar] [CrossRef]
- Alvarez-Vieites, E.; López-Santamarina, A.; Miranda, J.M.; Mondragón, A.D.C.; Lamas, A.; Cardelle-Cobas, A.; Nebot, C.; Franco, C.M.; Cepeda, A. Influence of the intestinal microbiota on diabetes management. Curr. Pharm. Biotechnol. 2020, 21, 1603–1615. [Google Scholar] [CrossRef]
- Roca-Saavedra, P.; Mendez-Vilabrille, V.; Miranda, J.M.; Nebot, C.; Cardelle-Cobas, A.; Franco, C.M.; Cepeda, A. Food additives, contaminants and other minor components: Effects on human gut microbiota—A review. J. Physiol. Biochem. 2018, 74, 69–83. [Google Scholar] [CrossRef]
- Quinto, E.; Jiménez, P.; Caro, I.; Tejero, J.; Mateo, J.; Girbés, T. Probiotic Lactic Acid Bacteria: A Review. Food Nutr. Sci. 2014, 5, 1765–1775. [Google Scholar] [CrossRef] [Green Version]
- Umu, Ö.C.O.; Rudi, K.; Diep, D.B. Modulation of the gut microbiota by prebiotic fibres and bacteriocins. Microb. Ecol. Health Dis. 2017, 28, 1348886. [Google Scholar] [CrossRef] [Green Version]
- Ou, Y.C.; Fu, H.C.; Tseng, C.W. The influence of probiotics on genital high-risk human papilloma virus clearance and quality of cervical smear: A randomized placebo-controlled trial. BMC Women Health 2019, 19, 103. [Google Scholar] [CrossRef] [Green Version]
- Kowalczyk, M.; Znamirowska-Piotrowska, A.; Buniowska-Olejnik, M.; Pawlos, M. Sheep’s Milk Symbiotic Ice Cream: Effect of Inulin and Apple Fiber on the Survival of Five Probiotic Bacterial Strains during Simulated In Vitro Digestion Conditions. Nutrients 2022, 14, 4454. [Google Scholar] [CrossRef]
- Paliwoda, A.; Nowak, A. Factors determining the adhesive capacity of Lactobacillus bacteria. Post. Mikrobiol.-Adv. Microbiol. 2017, 56, 196–204. [Google Scholar]
- Jungersen, M.; Wind, A.; Johansen, E.; Christensen, J.E.; Stuer-Lauridsen, B.; Eskesen, D. The Science behind the Probiotic Strain Bifidobacterium animalis subsp. lactis BB-12®. Microorganisms 2014, 2, 92–110. [Google Scholar] [CrossRef] [Green Version]
- Kumar, M.; Verma, V.; Nagpal, R.; Kumar, A.; Gautam, S.K.; Behare, P.V.; Grover, C.R.; Aggarwal, P.K. Effect of probiotic fermented milk and chlorophyllin on gene expressions and genotoxicity during AFB1-induced hepatocellular carcinoma. Gene 2011, 490, 54–59. [Google Scholar] [CrossRef]
- Balthazar, C.F.; Santillo, A.; Guimarães, J.T.; Capozzi, V.; Russo, P.; Caroprese, M.; Marino, R.; Esmerino, E.A.; Raices, R.S.L.; Silva, M.C.; et al. Novel milk–juice beverage with fermented sheep’s milk and strawberry (Fragaria × ananassa): Nutritional and functional characterization. J. Dairy Sci. 2019, 102, 10724–10736. [Google Scholar] [CrossRef] [Green Version]
- Espitia, P.J.P.; Batista, R.A.; Azeredo, H.M.C.; Otoni, C.G. Probiotics and their potential applications in active edible films and coatings. Food Res. Int. 2016, 90, 42–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ospanov, A.; Velyamov, S.; Tlevlessova, D.; Schetinina, E.; Kairbayeva, A.; Makeeva, R.; Tastanova, R. Survival of lactic acid bacteria when using the developed yogurt from the milk of small cattle under in-vitro conditions. Food Sci. Technol. 2023, 43, e117722. [Google Scholar] [CrossRef]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimera, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dantas, D.L.S.; Viera, V.B.; Soares, J.K.B.; Santos, K.M.O.; Egito, A.S.; Figueirêdo, R.M.F.; Lima, M.S.; Machado, N.A.F.; Souza, M.F.V.; Conceição, M.L.; et al. Pilosocereus gounellei (xique-xique) flour: Improving the nutritional, bioactive, and technological properties of probiotic goat-milk yogurt. LWT 2022, 158, 113165. [Google Scholar] [CrossRef]
- Verruck, S.; Barretta, C.; Miotto, M.; Canella, M.H.M.; Liz, G.R.; Maran, B.M.; Garcia, S.G.; Silveira, S.M.; Vieira, C.R.W.; Cruz, A.G.; et al. Evaluation of the interaction between microencapsulated Bifidobacterium BB-12 added in goat’s milk Frozen Yogurt and Escherichia coli in the large intestine. Food Res. Int. 2020, 127, 108690. [Google Scholar] [CrossRef]
- Wang, X.; Gao, S.; Yun, S.; Zhang, M.; Peng, L.; Li, Y.; Zhou, Y. Microencapsulating alginate-based polymers for probiotics delivery systems and their application. Pharmaceuticals 2022, 15, 644. [Google Scholar] [CrossRef]
- Melchior, S.; Marino, M.; Innocente, N.; Calligaris, S.; Nicoli, M.C. Effect of Different Biopolymer-Based Structured Systems on the Survival of Probiotic Strains during Storage and in Vitro Digestion. J. Sci. Food Agric. 2020, 100, 3902–3909. [Google Scholar] [CrossRef]
- Okpara, M.O. Microbial enzymes and their applications in food industry: A mini-review. Adv. Enzym. Res. 2022, 10, 23–47. [Google Scholar] [CrossRef]
- Yao, M.; Xie, J.; Du, H.; McClements, D.J.; Xiao, H.; Li, L. Progress in Microencapsulation of Probiotics: A Review. Compr. Rev. Food Sci. Food Saf. 2020, 19, 857–874. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Cui, S.W.; Chen, M.; Li, Y.; Liang, R.; Xu, F.; Zhong, F. Protective Approaches and Mechanisms of Microencapsulation to the Survival of Probiotic Bacteria during Processing, Storage and Gastrointestinal Digestion: A Review. Crit. Rev. Food Sci. Nutr. 2019, 59, 2863–2878. [Google Scholar] [CrossRef]
- Ranadheera, C.S.; Naumovski, N.; Ajlouni, S. Non-bovine milk products as emerging probiotic carriers: Recent developments and innovations. Curr. Opin. Food Sci. 2018, 22, 109–114. [Google Scholar] [CrossRef]
- Gustaw, W.; Kozioł, J.; Waśko, A.; Skrzypczak, K.; Michalak-Majewska, M.; Nastaj, M. Physicochemical properties and survival of Lactobacillus casei in fermented milk beverages produced with addition of selected milk protein preparations. Zywnosc-Nauka Technol. Jakosc 2015, 6, 129–139. [Google Scholar] [CrossRef]
- Rasika, D.M.D.; Munasinghe, M.A.D.D.; Vidanarachchi, J.K.; Cruz, A.G.; Ajlouni, S.; Ranadheera, C.S. Probiotics and prebiotics in non-bovine milk. Adv. Food Nutr. Res. 2020, 94, 339–384. [Google Scholar] [CrossRef]
- Varga, L.; Süle, J.; Nagy, P. Short communication: Survival of the characteristic microbiota in probiotic fermented camel, cow, goat, and sheep’s milks during refrigerated storage. J. Dairy Sci. 2014, 97, 2039–2044. [Google Scholar] [CrossRef] [Green Version]
- Costa, M.P.; Frasao, B.S.; Silva, A.C.O.; Freitas, M.Q.; Franco, R.M.; Conte-Junior, C.A. Cupuassu (Theobroma grandiflorum) pulp, probiotic, and prebiotic: Influence on color, apparent viscosity, and texture of goat milk yogurts. J. Dairy Sci. 2015, 98, 5995–6003. [Google Scholar] [CrossRef]
- Martinez, G.M.; Suarez, V.H. Lechería Caprina: Producción, Manejo, Sanidad, Calidad de Leche; INTA Ediciones; Instituto Nacional de Tecnologías de los Alimentos: Buenos Aires, Argentina, 2018. [Google Scholar]
- Fonseca, C.R.; Bordin, K.; Fernandes, A.M.; Rodrigues, C.E.; Corassin, C.H.; Cruz, A.G.; Oliveira, C.A. Storage of refrigerated raw goat milk affecting the quality of whole milk powder. J. Dairy Sci. 2013, 96, 4716–4724. [Google Scholar] [CrossRef] [Green Version]
- Kapila, R.; Kavadi, P.K.; Kapila, S. Comparative evaluation of allergic sensitization to milk proteins of cow, buffalo and goat. Small Rumin. Res. 2013, 112, 191–198. [Google Scholar] [CrossRef]
- Teng, F.; Reis, M.G.; Yang, L.; Ma, Y.; Day, L. Structural characteristics of triacylglycerols contribute to the distinct in vitro gastric digestibility of sheep’s and cow milk fat prior to and after homogenization. Food Res. Int. 2020, 130, 108911. [Google Scholar] [CrossRef]
- Mohapatraa, A.; Shindeb, A.K.; Singh, R. Sheep’s milk: A pertinent functional food. Small Rumin. Res. 2019, 181, 6–11. [Google Scholar] [CrossRef]
- Czyżak-Runowska, G.; Wójtowski, J.A.; Łęska, B.; Bielińska-Nowak, S.; Pytlewski, J.; Antkowiak, I.; Stanisławski, D. Lactose Content and Selected Quality Parameters of Sheep’s Milk Fermented Beverages during Storage. Animals 2022, 12, 3105. [Google Scholar] [CrossRef]
- Günter, E.A.; Martynov, V.V.; Ananchenko, B.A.; Martinson, E.A.; Litvinets, S.G. The gel strength and swelling in the gastrointestinal environment of pectin/κ-carrageenan gel particles based on pectins with different degrees of methylesterification. Mater. Today Commun. 2022, 33, 104986. [Google Scholar] [CrossRef]
- Jin, H.S.; Fei, Y.S.; Yan, C.K.; Kuan, C.H.; Wei, S.W.Y. Effect of gums coating materials on the survival of microencapsulated probiotics under simulated gastrointestinal conditions. Mater. Today Proc. 2020, 29, 16–19. [Google Scholar] [CrossRef]
- Caillard, R.; Lapointe, N. In vitro gastric survival of commercially available probiotic strains and oral dosage forms. Int. J. Pharm. 2017, 519, 125–127. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Medronho, B.; dos Santos, T.; Nunes-Correia, I.; Granja, P.; Miguel, M.G.; Lindman, B. On the Viability, Cytotoxicity and Stability of Probiotic Bacteria Entrapped in Cellulose-Based Particles. Food Hydrocoll. 2018, 82, 457–465. [Google Scholar] [CrossRef]
- Obradovic, N.; Pajic-Lijakovic, I.; Krunic, T.; Belovic, M.; Rakin, M.; Bugarski, B. Effect of Encapsulated Probiotic Starter Culture on Rheological and Structural Properties of Natural Hydrogel Carriers Affected by Fermentation and Gastrointestinal Conditions. Food Biophys. 2020, 15, 18–31. [Google Scholar] [CrossRef]
- Aryana, K.J.; Olson, D.W. A 100-Year Review: Yogurt and other cultured dairy products. J. Dairy Sci. 2017, 100, 9987–10013. [Google Scholar] [CrossRef] [Green Version]
- Szopa, K.; Znamirowska-Piotrowska, A.; Szajnar, K.; Pawlos, M. Effect of Collagen Types, Bacterial Strains and Storage Duration on the Quality of Probiotic Fermented Sheep’s Milk. Molecules 2022, 27, 3028. [Google Scholar] [CrossRef]
- Szopa, K.; Pawlos, M.; Znamirowska-Piotrowska, A. Effect of Storage Time and Bacterial Strain on the Quality of Probiotic Goat’s Milk Using Different Types and Doses of Collagens. Molecules 2023, 28, 657. [Google Scholar] [CrossRef]
- Ramasubramanian, L.; Webb, R.; Arcy, R.B.; Deeth, H. Characteristic of calcium-milk coagulum. J. Food Eng. 2013, 114, 147–152. [Google Scholar] [CrossRef]
- Commission Regulation (EC). No 1662/2006 of 6 November 2006 Amending Regulation (EC) No 853/2004 of the European Parliament and of the Council Laying down Specific Hygiene Rules for Food of Animal Origin (Text with EEA Relevance). Off. J. Eur. Union 2006, L320, 1–10. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/HTML/?uri=CELEX:32006R1662&from=EN (accessed on 20 June 2022).
- Szajnar, K.; Pawlos, M.; Znamirowska, A. The Effect of the Addition of Chokeberry Fiber on the Quality of Sheep’s Milk Fermented by Lactobacillus rhamnosus and Lact. acidophilus. Int. J. Food Sci. 2021, 2021, 7928745. [Google Scholar] [CrossRef]
- Buniowska, M.; Carbonell-Capella, J.M.; Frigola, A.; Esteve, M.J. Bioaccessibility of bioactive compounds after non-thermal processing of an exotic fruit juice blend sweetened with Stevia rebaudiana. Food Chem. 2017, 221, 1834–1842. [Google Scholar] [CrossRef]
- Silva, C.C.; da Silva Barros, E.L.; Verruck, S.; Maran, B.M.; Canella, M.H.M.; Esmerino, E.A.; Ramon Silva, R.; Prudencio, E.S. How ice cream manufactured with concentrated milk serves as a protective probiotic carrier? An in vitro gastrointestinal assay. Food Sci. Technol. 2022, 42, 28621. [Google Scholar] [CrossRef]
- Znamirowska, A.; Szajnar, K.; Pawlos, M. Probiotic Fermented Milk with Collagen. Dairy 2020, 1, 126–134. [Google Scholar] [CrossRef]
- Lima, K.G.D.; Kruger, M.F.; Behrens, J.; Destro, M.T.; Landgraf, M.; Franco, B.D.G.M. Evaluation of culture media for enumeration of Lactobacillus acidophilus, Lactobacillus casei and Bifidobacterium animalis in the presence of Lactobacillus delbrueckii subsp. bulgaricus and Streptococcus thermophilus. LWT Food Sci. Technol. 2009, 42, 491–495. [Google Scholar] [CrossRef]
- Sun, J.; Chen, H.; Qiao, Y.; Liu, G.; Leng, C.; Zhang, Y.; Lv, X.; Feng, Z. The nutrient requirements of Lactobacillus rhamnosus GG and their application to fermented milk. J. Dairy. Sci. 2019, 102, 5971–5978. [Google Scholar] [CrossRef]
- Turkmen, N. The Nutritional Value and Health Benefits of Goat Milk Components. Nutr. Dairy Their Implic. Health Dis. 2017, 35, 441–449. [Google Scholar] [CrossRef]
- Lima, M.J.R.; Teixeira-Lemos, E.; Oliveira, J.; Teixeira-Lemos, L.P.; Monteiro, A.M.C.; Costa, J.M. Nutritional and Health Profile of Goat Products: Focus on Health Benefits of Goat Milk. In Goat Science; IntechOpen: London, UK, 2018; pp. 198–231. [Google Scholar] [CrossRef] [Green Version]
- Flis, Z.; Molik, E. Importance of Bioactive Substances in Sheep’s Milk in Human Health. Int. J. Mol. Sci. 2021, 22, 4364. [Google Scholar] [CrossRef]
- Hu, X.; Liu, C.; Zhang, H.; Hossen Md, A.; Sameen, D.E.; Dai, J.; Qin, W.; Liu, Y.; Li, S. In vitro digestion of sodium alginate/pectin co-encapsulated Lactobacillus bulgaricus and its application in yogurt bilayer beads. Int. J. Biol. Macromol. 2021, 193, 1050–1058. [Google Scholar] [CrossRef]
- Prestes, A.A.; Verruck, S.; Vargas, M.O.; Canella, M.H.M.; Silva, C.C.; Barros, E.L.S.; Dantas, A.; Oliveira, L.V.A.; Maran, B.M.; Matos, M.; et al. Influence of guabiroba pulp (campomanesia xanthocarpa o. berg) added to fermented milk on probiotic survival under in vitro simulated gastrointestinal conditions. Food Res. Int. 2021, 141, 110135. [Google Scholar] [CrossRef]
- Haji, F.; Cheon, J.; Baek, J.; Wang, Q.; Tam, K.C. Application of Pickering emulsions in probiotic encapsulation—A review. Curr. Res. Food Sci. 2022, 5, 1603–1615. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.T.H.; Gathercole, J.L.; Day, L.; Dalziel, J.E. Differences in peptide generation following in vitro gastrointestinal digestion of yogurt and milk from cow, sheep’s and goat. Food Chem. 2020, 317, 126419. [Google Scholar] [CrossRef] [PubMed]
- Dalziel, J.E.; Dunstan, K.E.; Dewhurst, H.; Van Gendt, M.; Young, W.; Carpenter, E. Goat milk increases gastric emptying and alters caecal short chain fatty acid profile compared with cow milk in healthy rats. Food Funct. 2020, 11, 8573–8582. [Google Scholar] [CrossRef] [PubMed]
- Naissinger da Silva, M.; Tagliapietra, B.L.; Flores, V.A.; Richards, N.S.P.S. In vitro test to evaluate survival in the gastrointestinal tract of commercial probiotics. Curr. Res. Food Sci. 2021, 4, 320–325. [Google Scholar] [CrossRef] [PubMed]
- Ziarno, M.; Zaręba, D. Effects of milk components and food additives on survival of three bifidobacteria strains in fermented milk under simulated gastrointestinal tract conditions. Microb. Ecol. Health Dis. 2015, 26, 1. [Google Scholar] [CrossRef]
- De Oliveira Galdino, I.K.C.P.; Oliveira, M.M.; Oliveira, A.T.; da Silva, G.M.; de Oliveira, T.A.; dos Santos, K.M.O.; do Egito, A.S.; Buriti, F.C.A. Fermentative behavior of native lactobacilli in goat milk and their survival under in vitro simulated gastrointestinal conditions. LWT 2020, 135, 109905. [Google Scholar] [CrossRef]
- Wu, S.C.; Wang, F.J.; Pan, C.L. Growth and survival of lactic acid bacteria during the fermentation and storage of seaweed oligosaccharides solution. J. Mar. Sci. Technol. 2007, 15, 104–114. [Google Scholar] [CrossRef]
- Śliżewska, K.; Chlebicz-Wójcik, A. Growth Kinetics of Probiotic Lactobacillus Strains in the Alternative, Cost-Efficient Semi-Solid Fermentation Medium. Biology 2020, 9, 423. [Google Scholar] [CrossRef]
- König, H.; Berkelmann-Löhnertz, B. Maintenance of wine-associated microorganisms. In Biology of Microorganisms on Grapes, in Must and in Wine; König, H., Gottfried, U., Fröhlich, J., Eds.; Springer: Cham, Switzerland, 2017; pp. 549–571. [Google Scholar] [CrossRef]
- Chr. Hansen: Our Probiotic Strains. Available online: https://www.chr-hansen.com/en/human-health-and-probiotics/our-probiotic-strains/l-casei (accessed on 4 January 2022).
- Amund, O.D. Exploring the Relationship between Exposure to Technological and Gastrointestinal Stress and Probiotic Functional Properties of lactobacilli and bifidobacteria. Can. J. Microbiol. 2016, 62, 715–725. [Google Scholar] [CrossRef]
- Afzaal, M.; Khan, A.U.; Saeed, F.; Arshad, M.S.; Khan, M.A.; Saeed, M.; Maan, A.A.; Khan, M.K.; Ismail, Z.; Ahmed, A. Survival and stability of free and encapsulated probiotic bacteria under simulated gastrointestinal conditions and in ice cream. Food Sci. Nutr. 2020, 8, 1649–1656. [Google Scholar] [CrossRef] [Green Version]
- Doherty, S.B.; Gee, V.L.; Ross, R.P.; Stanton, C.; Fitzgerald, G.F.; Brodkorb, A. Development and characterisation of whey protein micro-beads as potential matrices for probiotic protection. Food Hydrocoll. 2011, 25, 1604–1617. [Google Scholar] [CrossRef]
- Ranadheera, C.S.; Evans, C.A.; Adams, M.C.; Baines, S.K. Effect of dairy probiotic combinations on in vitro gastrointestinal tolerance, intestinal epithelial cell adhesion and cytokine secretion. J. Funct. Foods 2014, 8, 18–25. [Google Scholar] [CrossRef]
- Ayyash, M.; Abdalla, A.; Alhammadi, A.; Ranadheera, C.S.; Baig, M.A.; Al-Ramadi, B.; Chen, G.; Kamal-Eldin, A.; Huppertz, T. Probiotic survival, biological functionality and untargeted metabolomics of the bioaccessible compounds in fermented camel and bovine milk after in vitro digestion. Food Chem. 2021, 363, 130243. [Google Scholar] [CrossRef]
- Moumita, S.; Goderska, K.; Johnson, E.M.; Das, B.; Indira, D.; Yadav, R.; Kumari, S.; Jayabalan, R. Evaluation of the viability of free and encapsulated lactic acid bacteria using in-vitro gastro intestinal model and survivability studies of synbiotic microcapsules in dry food matrix during storage. LWT—Food Sci. Technol. 2017, 77, 460–467. [Google Scholar] [CrossRef]
- Radicioni, M.; Koirala, R.; Fiore, W.; Leuratti, C.; Guglielmetti, S.; Arioli, S. Survival of L. casei DG® (Lactobacillus paracasei CNCMI1572) in the gastrointestinal tract of a healthy paediatric population. Eur. J. Nutr. 2019, 58, 3161–3170. [Google Scholar] [CrossRef] [Green Version]
- Ferrario, C.; Taverniti, V.; Milani, C.; Fiore, W.; Laureati, M.; De Noni, I.; Stuknyte, M.; Chouaia, B.; Riso, P.; Guglielmetti, S. Modulation of fecal Clostridiales bacteria and butyrate by probiotic intervention with Lactobacillus paracasei DG varies among healthy adults. J. Nutr. 2014, 144, 1787–1796. [Google Scholar] [CrossRef] [Green Version]
- Castro-Bravo, N.; Wells, J.M.; Margolles, A.; Ruas-Madiedo, P. Interactions of surface exopolysaccharides from Bifidobacterium and Lactobacillus within the intestinal environment. Front. Microbiol. 2018, 9, 2426. [Google Scholar] [CrossRef] [Green Version]
- Balzaretti, S.; Taverniti, V.; Guglielmetti, S.; Fiore, W.; Minuzzo, M.; Ngo, N.H.; Ngere, B.J.; Sadiq, S.; Humphreys, P.N.; Laws, A.P. A novel rhamnose-rich hetero-exopolysaccharide isolated from Lactobacillus paracasei DG activates THP-1 human monocytic cells. Appl. Environ. Microbiol. 2017, 83, 702–716. [Google Scholar] [CrossRef] [Green Version]
- Leeuwendaal, N.; Hayes, J.J.; Stanton, C.; O’Toole, P.W.; Beresford, T.P. Protection of Candidate Probiotic Lactobacilli by Cheddar Cheese Matrix during Simulated Gastrointestinal Digestion. J. Funct. Foods 2022, 92, 105042. [Google Scholar] [CrossRef]
- Florence, A.C.R.; de Oliveira, M.N.; Delile, A.; Béal, C. Survival of Bifidobacterium strains in organic fermented milk is improved as a result of membrane fatty acid composition. Int. Dairy J. 2016, 61, 1–9. [Google Scholar] [CrossRef]
- Verruck, S.; Prudêncio, E.S.; Vieira, C.R.W.; Amante, E.R.; Amboni, R.D.M.C. The buffalo Minas Frescal cheese as a protective matrix of Bifidobacterium BB-12 under in vitro simulated gastrointestinal conditions. LWT—Food Sci. Technol. 2015, 63, 1179–1183. [Google Scholar] [CrossRef]





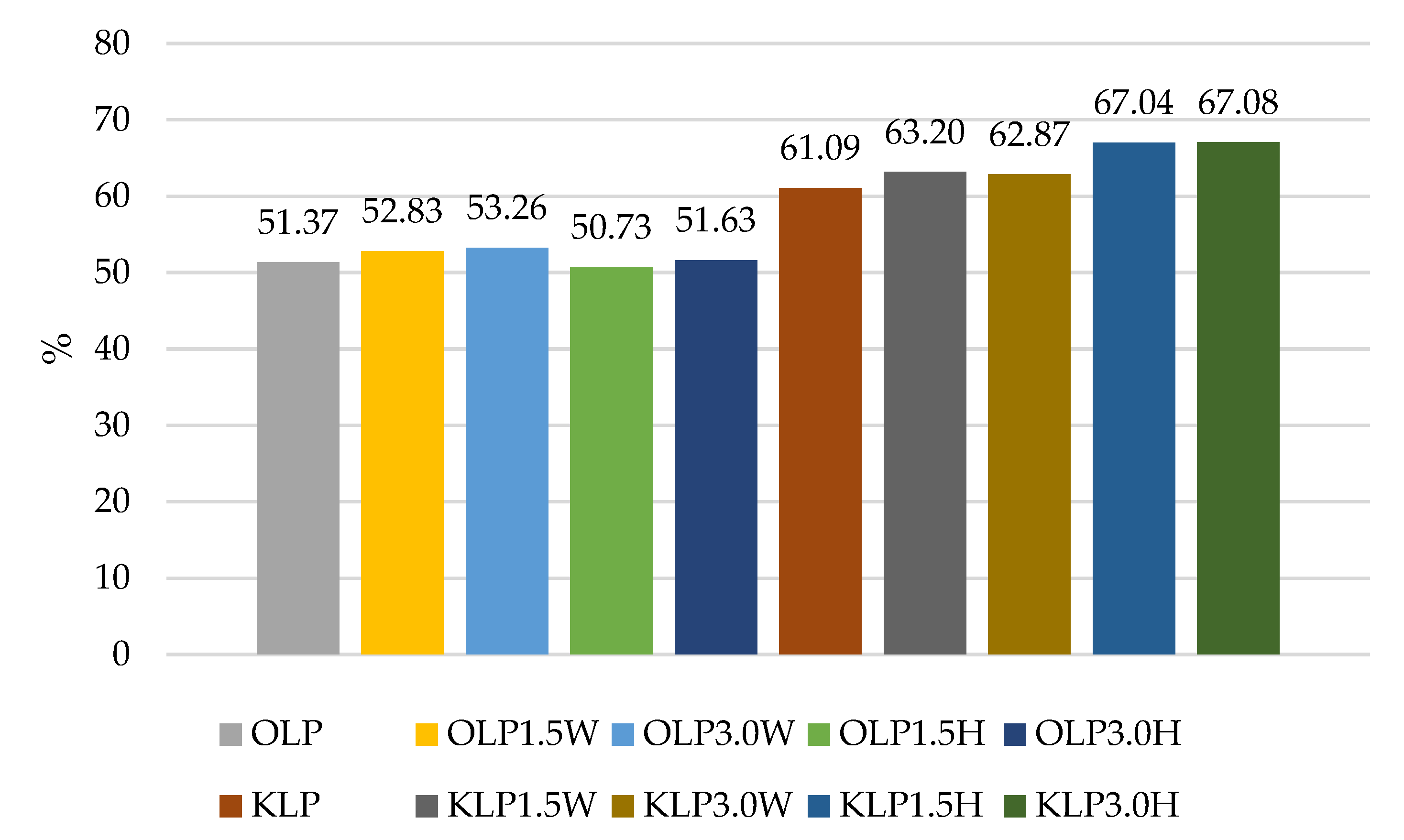{kind=link}
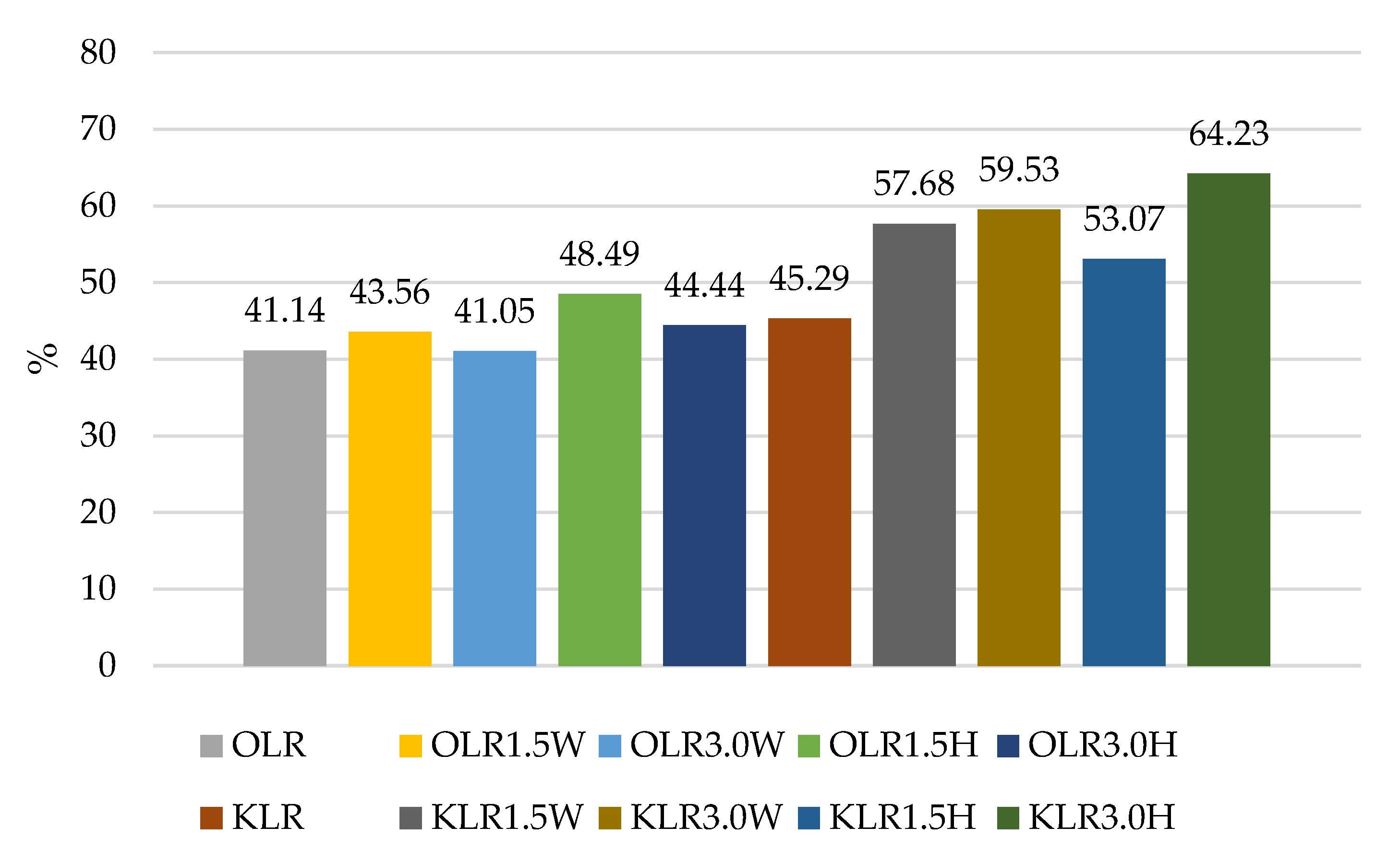{kind=link}
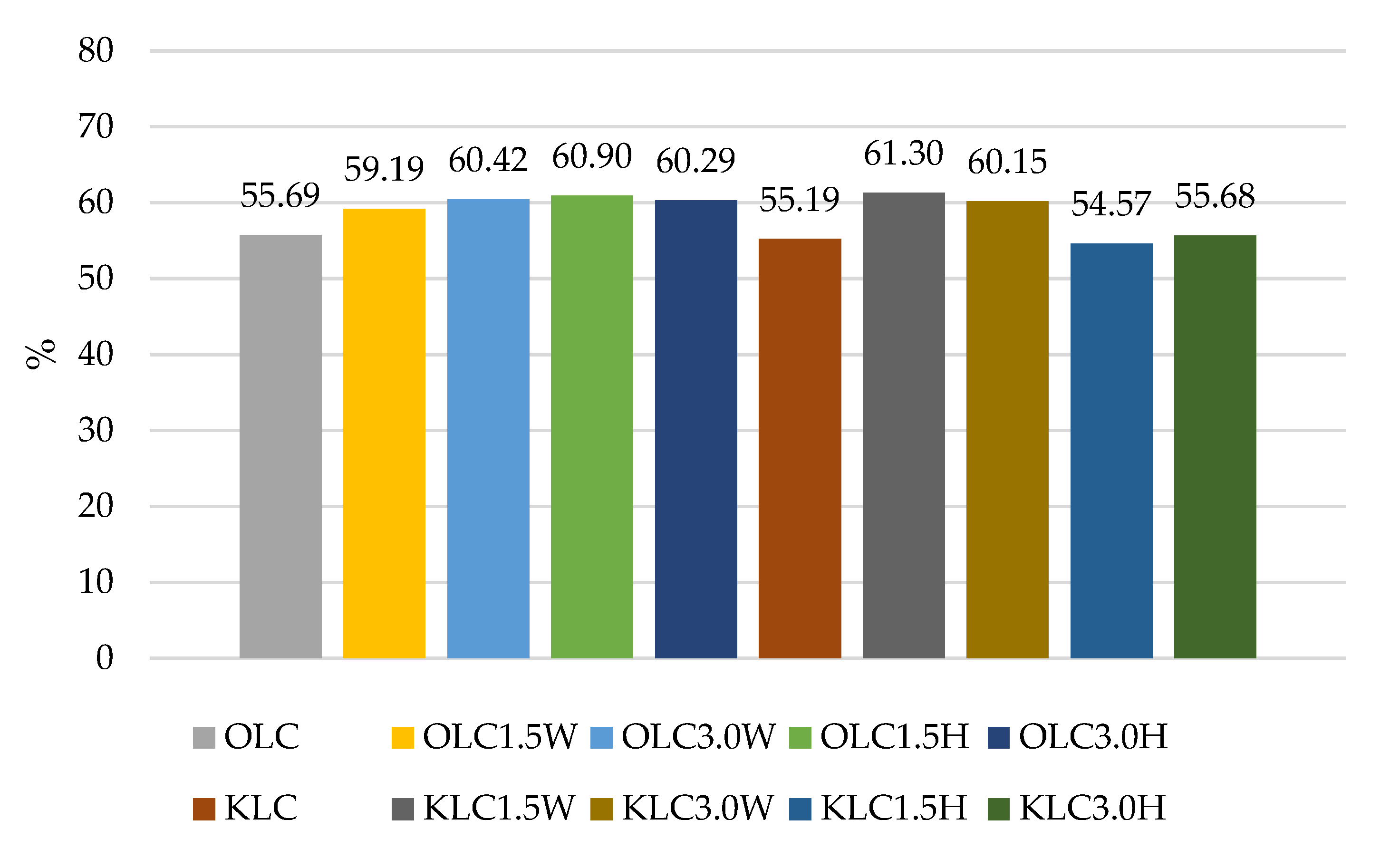{kind=link}
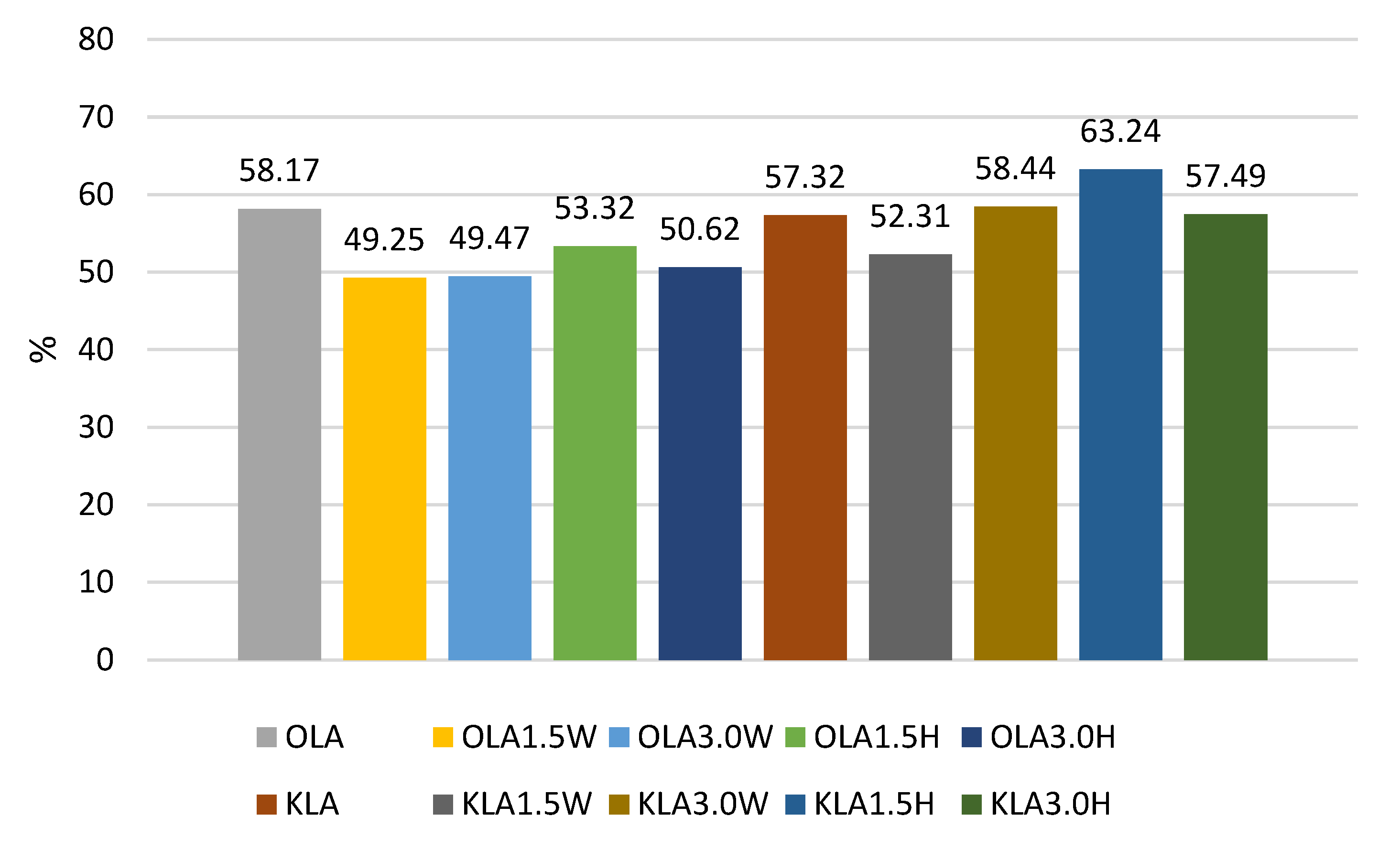{kind=link}
| Probiotic Strain | Batch | ||||
|---|---|---|---|---|---|
| Control | 1.5% Bovine Collagen | 3.0% Bovine Collagen | 1.5% Collagen Protein Hydrolysate | 3.0% Collagen Protein Hydrolysate | |
| Lacticaseibacillus casei 431 | OLC | OLC1.5W | OLC3.0W | OLC1.5H | OLC3.0H |
| Lacticaseibacillus paracasei L-26 | OLP | OLP1.5W | OLP3.0W | OLP1.5H | OLP3.0H |
| Lactobacillus acidophilus LA-5 | OLA | OLA1.5W | OLA3.0W | OLA1.5H | OLA3.0H |
| Lacticaseibacillus rhamnosus Lr-32 | OLR | OLR1.5W | OLR3.0W | OLR1.5H | OLR3.0H |
| Probiotic Strain | Batch | ||||
|---|---|---|---|---|---|
| Control | 1.5% Bovine Collagen | 3.0% Bovine Collagen | 1.5% Collagen Protein Hydrolysate | 3.0% Collagen Protein Hydrolysate | |
| Lacticaseibacillus casei 431 | KLC | KLC1.5W | KLC3.0W | KLC1.5H | KLC3.0H |
| Lacticaseibacillus paracasei L-26 | KLP | KLP1.5W | KLP3.0W | KLP1.5H | KLP3.0H |
| Lactobacillus acidophilus LA-5 | KLA | KLA1.5W | KLA3.0W | KLA1.5H | KLA3.0H |
| Lacticaseibacillus rhamnosus Lr-32 | KLR | KLR1.5W | KLR3.0W | KLR1.5H | KLR3.0H |
| Experimental Batch | Viable Counts of Probiotic Bacteria, log cfu g−1 | |||
|---|---|---|---|---|
| Digestion Phase | ||||
| Before Digestion | Oral Cavity | Stomach | Small Intensine | |
| OLP | 10.92 Cb ± 0.14 | 10.50 Bb ± 0.17 | 5.40 Ba ± 0.13 | 5.61 Aa ± 0.21 |
| OLP1.5W | 10.77 Cb ± 0.15 | 10.84 Cb ± 0.12 | 5.46 Ba ± 0.14 | 5.69 Aa ± 0.11 |
| OLP3.0W | 10.89 Cb ± 0.04 | 10.85 Cb ± 0.12 | 5.52 Ba ± 0.16 | 5.80 Aa ± 0.11 |
| OLP1.5H | 10.90 Cb ± 0.25 | 10.92 Cb ± 0.03 | 5.55 Ba ± 0.05 | 5.53 Aa ± 0.11 |
| OLP3.0H | 10.46 Bb ± 0.23 | 10.53 BCb ± 0.37 | 5.35 Ba ± 0.14 | 5.40 Aa ± 0.12 |
| KLP | 9.02 Ab ± 0.22 | 9.09 Ab ± 0.11 | 5.06 Aa ± 0.27 | 5.51 Aa ± 0.16 |
| KLP1.5W | 9.24 Ab ± 0.18 | 9.17 Ab ± 0.39 | 5.84 Ca ± 0.05 | 5.84 Ba ± 0.09 |
| KLP3.0W | 9.40 Ab ± 0.23 | 9.45 Ab ± 0.68 | 5.92 Ca ± 0.20 | 5.91 Aa ± 0.45 |
| KLP1.5H | 8.92 Ac ± 0.16 | 8.77 Ac ± 0.18 | 5.45 Ba ± 0.21 | 5.98 Bb ± 0.17 |
| KLP3.0H | 8.90 Ac ± 0.22 | 8.70 Ac ± 0.30 | 5.47 Ba ± 0.19 | 5.97 Bb ± 0.07 |
| Experimental Batch | Viable Counts of Probiotic Bacteria, log cfu g−1 | |||
|---|---|---|---|---|
| Digestion Phase | ||||
| Before Digestion | Oral Cavity | Stomach | Small Intensine | |
| OLR | 10.21 Cb ± 0.23 | 9.90 Ab ± 0.24 | 4.06 Ba ± 0.08 | 4.20 Aa ± 0.40 |
| OLR1.5W | 10.17 Cb ± 0.21 | 9.88 ABb ± 0.15 | 3.58 Aa ± 0.68 | 4.43 Aa ± 0.34 |
| OLR3.0W | 10.50 Db ± 0.24 | 9.88 Bb ± 0.42 | 3.78 Aa ± 0.62 | 4.31 Aa ± 0.39 |
| OLR1.5H | 10.27 Cb ± 0.28 | 9.79 Abb ± 0.21 | 3.65 Aa ± 0.16 | 4.98 Aa ± 0.97 |
| OLR3.0H | 10.26 Cb ± 0.08 | 9.85 Ab ± 0.14 | 3.87 Aa ± 0.32 | 4.56 Aa ± 0.36 |
| KLR | 9.23 Bb ± 0.13 | 9.25 ABb ± 0.38 | 4.02 Ba ± 0.52 | 4.18 Aa ± 0.19 |
| KLR1.5W | 9.05 Ac ± 0.20 | 8.92 Ac ± 0.18 | 3.69 Aa ± 0.58 | 5.22 Bb ± 0.22 |
| KLR3.0W | 8.97 Ac ± 0.07 | 8.99 Ac ± 0.29 | 4.10 Ba ± 0.13 | 5.34 Bb ± 0.21 |
| KLR1.5H | 9.12 Ac ± 0.14 | 9.07 Ac ± 0.22 | 4.00 Ba ± 0.06 | 4.84 Ab ± 0.61 |
| KLR3.0H | 9.17 ABc ± 0.40 | 8.99 Ac ± 0.11 | 4.28 Ba ± 0.17 | 5.89 Bb ± 0.30 |
| Experimental Batch | Viable Counts of Probiotic Bacteria, log cfu g−1 | |||
|---|---|---|---|---|
| Digestion Phase | ||||
| Before Digestion | Oral Cavity | Stomach | Small Intensine | |
| OLC | 9.57 Bc ± 0.34 | 9.75 Cc ± 0.09 | 4.94 Ba ± 0.11 | 5.33 Bb ± 0.19 |
| OLC1.5W | 9.68 Bb ± 0.34 | 9.74 Cb ± 0.09 | 5.72 Ca ± 0.18 | 5.73 Ca ± 0.11 |
| OLC3.0W | 9.60 Bc ± 0.09 | 9.57 Bc ± 0.14 | 5.74 Cb ± 0.04 | 5.80 Ca ± 0.09 |
| OLC1.5H | 9.54 Bb ± 0.04 | 9.47 Bb ± 0.10 | 5.79 Ca ± 0.14 | 5.81 Ca ± 0.28 |
| OLC3.0H | 9.52 Bb ± 0.05 | 9.42 Bb ± 0.06 | 5.78 Ca ± 0.78 | 5.74 Ca ± 0.16 |
| KLC | 9.15 Ac ± 0.39 | 9.44 Bc ± 0.12 | 4.25 Aa ± 0.06 | 5.05 Ab ± 0.21 |
| KLC1.5W | 9.56 ABc ± 0.47 | 9.38 Ac ± 0.21 | 5.65 Ca ± 0.19 | 5.86 Ba ± 0.14 |
| KLC3.0W | 9.51 Bb ± 0.13 | 9.58 Bb ± 0.18 | 5.72 Ca ± 0.09 | 5.72 Ba ± 0.22 |
| KLC1.5H | 9.20 Ac ± 0.12 | 9.10 Ac ± 0.28 | 5.81 Cb ± 0.14 | 5.02 Aa ± 0.37 |
| KLC3.0H | 9.07 Ab ± 0.21 | 8.95 Ab ± 0.16 | 5.64 Ca ± 0.36 | 5.05 Aa ± 0.50 |
| Experimental Batches | Viable Counts of Probiotic Bacteria, log cfu g−1 | |||
|---|---|---|---|---|
| Digestion Phase | ||||
| Before Digestion | Oral Cavity | Stomach | Small Intensine | |
| OLA | 9.18 ABb ± 0.10 | 9.14 Bb ± 0.18 | 4.72 Aa ± 0.45 | 5.34 Ba ± 0.37 |
| OLA1.5W | 9.34 Bb ± 0.06 | 9.35 Cb ± 0.11 | 4.73 Aa ± 0.49 | 4.60 Aa ± 0.39 |
| OLA3.0W | 9.38 Bb ± 0.09 | 9.31 Cb ± 0.34 | 4.42 Aa ± 0.61 | 4.64 Aa ± 0.29 |
| OLA1.5H | 9.49 Bb ± 0.11 | 9.42 Cb ± 0.18 | 4.46 Aa ± 0.68 | 5.06 Ba ± 0.28 |
| OLA3.0H | 9.76 Bb ± 0.11 | 9.70 Cb ± 0.18 | 4.72 Aa ± 0.28 | 4.94 Ba ± 0.20 |
| KLA | 8.95 Ab ± 0.38 | 8.82 Bb ± 0.24 | 5.45 Ba ± 0.12 | 5.13 Ba ± 0.19 |
| KLA1.5W | 8.87 Ab ± 0.11 | 8.60 Ab ± 0.25 | 5.27 Ba ± 0.22 | 4.64 Aa ± 0.60 |
| KLA3.0W | 8.71 Ab ± 0.62 | 8.36 Ab ± 0.54 | 4.85 Aa ± 0.59 | 5.09 Ba ± 0.34 |
| KLA1.5H | 8.27 Ab ± 0.19 | 8.44 Ab ± 0.39 | 5.37 Ba ± 0.14 | 5.23 Ba ± 0.71 |
| KLA3.0H | 8.28 Ac ± 0.11 | 8.22 Ac ± 0.27 | 5.10 Bb ± 0.05 | 4.76 Aa ± 0.77 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szopa, K.; Szajnar, K.; Pawlos, M.; Znamirowska-Piotrowska, A. Probiotic Fermented Goat’s and Sheep’s Milk: Effect of Type and Dose of Collagen on Survival of Four Strains of Probiotic Bacteria during Simulated In Vitro Digestion Conditions. Nutrients 2023, 15, 3241. https://doi.org/10.3390/nu15143241
Szopa K, Szajnar K, Pawlos M, Znamirowska-Piotrowska A. Probiotic Fermented Goat’s and Sheep’s Milk: Effect of Type and Dose of Collagen on Survival of Four Strains of Probiotic Bacteria during Simulated In Vitro Digestion Conditions. Nutrients. 2023; 15(14):3241. https://doi.org/10.3390/nu15143241
Chicago/Turabian StyleSzopa, Kamil, Katarzyna Szajnar, Małgorzata Pawlos, and Agata Znamirowska-Piotrowska. 2023. "Probiotic Fermented Goat’s and Sheep’s Milk: Effect of Type and Dose of Collagen on Survival of Four Strains of Probiotic Bacteria during Simulated In Vitro Digestion Conditions" Nutrients 15, no. 14: 3241. https://doi.org/10.3390/nu15143241
APA StyleSzopa, K., Szajnar, K., Pawlos, M., & Znamirowska-Piotrowska, A. (2023). Probiotic Fermented Goat’s and Sheep’s Milk: Effect of Type and Dose of Collagen on Survival of Four Strains of Probiotic Bacteria during Simulated In Vitro Digestion Conditions. Nutrients, 15(14), 3241. https://doi.org/10.3390/nu15143241