
Redox Regulation in Age-Related Cataracts: Roles for Glutathione, Vitamin C, and the NRF2 Signaling Pathway

 ,
,  , and
, and
Abstract
1. Introduction
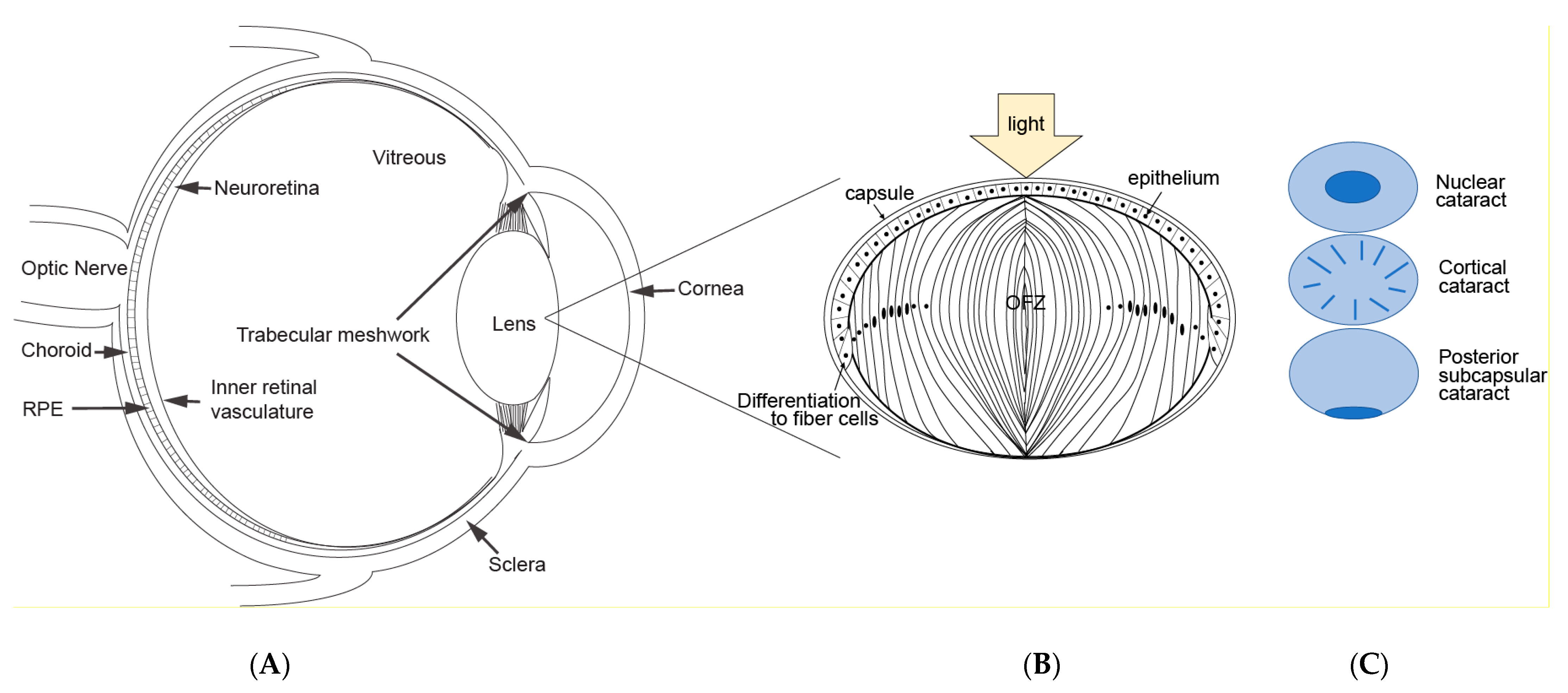
2. Lens Structure and Cataract
3. Glutathione and the Lens
3.1. Glutathione Regulation in the Lens
3.2. Mechanisms Involved in the Maintenance of GSH Homeostasis in the Lens
3.3. GSH Levels in Aging and Cataract
3.4. In Vivo Analysis of GSH Deficiency and Cataracts
3.5. Glutathione and Protein Homeostasis
4. Vitamin C and the Lens
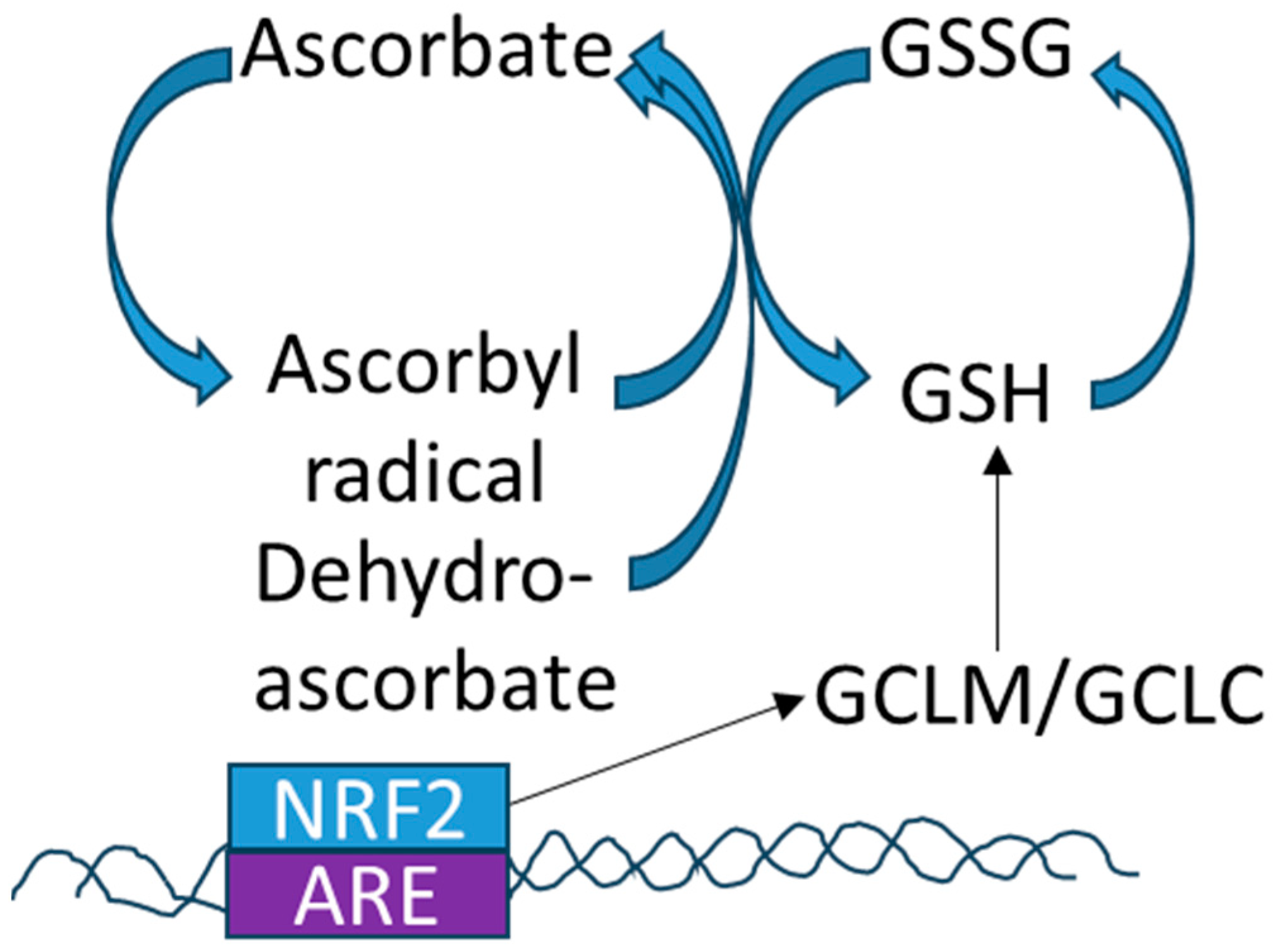
4.1. Redox and Vitamin C
4.2. Vitamin C Levels in Lens and Cataract
4.3. Too Much of a Good Thing?

{kind=link}
{kind=link}
{kind=link}
| Association | Type of Study | Findings | Ref. |
|---|---|---|---|
| INDEYE | Cross-sectional | Decreased risk of all catraract types in highest vs. lowest tertiles of plasma vitamin C | [82,83] |
| Nutrition and Vision Project | Retrospective | Decreased risk of cataract with ≥363 mg/day Vitamin C | [83] |
| Intervention | Length of Intervention | Findings | Ref. |
| REACT | 3 years | Small decrease in opacification upon supplementation | [85] |
| APC | 5 years | No difference in cataract upon supplememtation | [86] |
| PHS | 11.2 years | 9% lower risk in multivitamin supplemented group | [87] |
| Swedish mammography cohort | 8.2 years | 25% increased risk with high dose supplementation of vitamin C | [92] |
| Cohort of Swedish men | 8.4 years | 21% increased risk with high dose supplementation of vitamin C alone | [93] |
5. NRF2 and Cataract
5.1. NRF2 Antioxidant Response
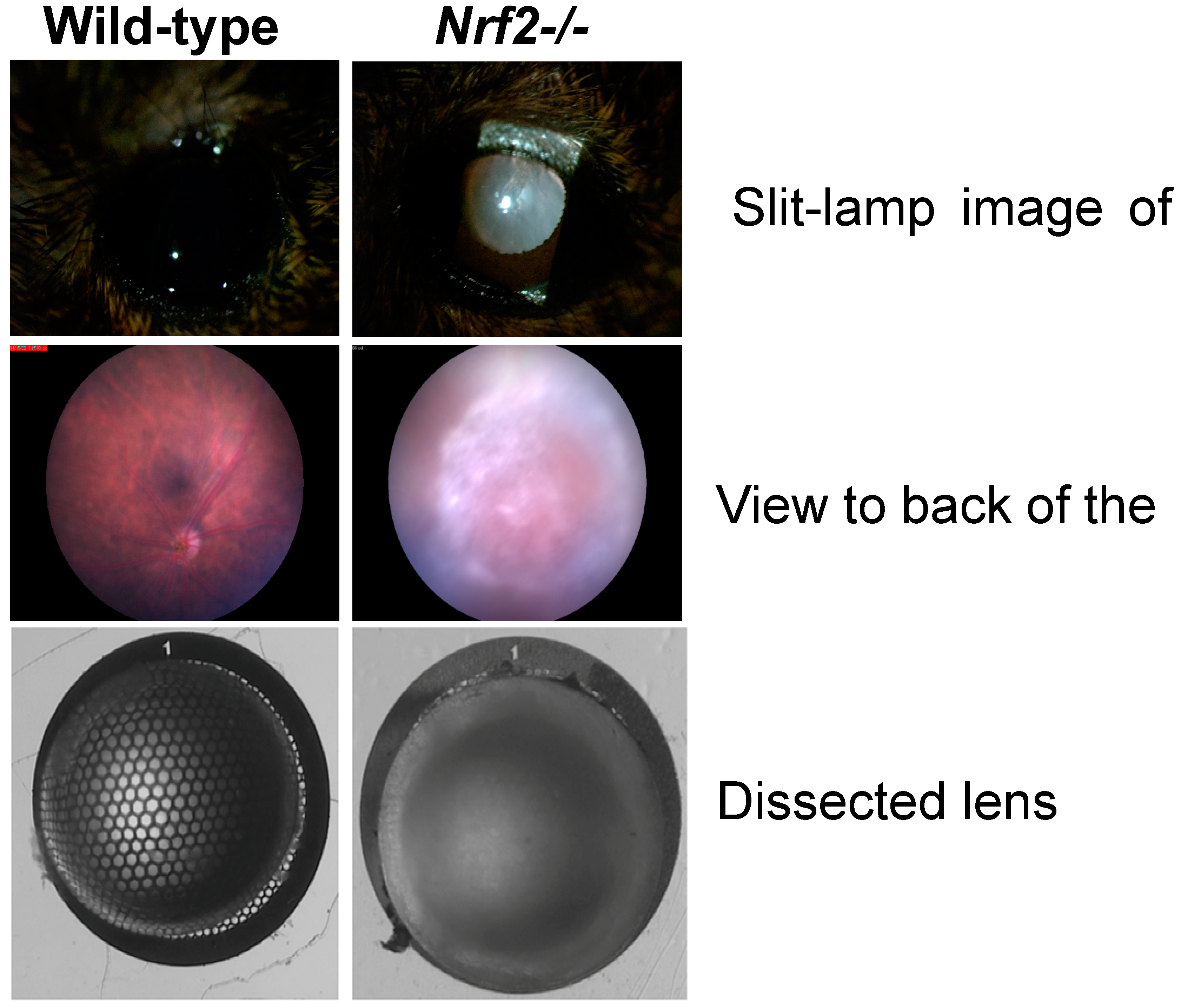
5.2. NRF2 in the Lens
6. Concluding Remarks and Future Perspectives
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pascolini, D.; Mariotti, S.P. Global estimates of visual impairment: 2010. Br. J. Ophthalmol. 2011, 96, 614–618. [Google Scholar] [CrossRef] [PubMed]
- Serebryany, E.; Thorn, D.C.; Quintanar, L. Redox chemistry of lens crystallins: A system of cysteines. Exp. Eye Res. 2021, 211, 108707. [Google Scholar] [CrossRef]
- Lim, J.C.; Grey, A.; Zahraei, A.; Donaldson, P.J. Age-dependent changes in glutathione metabolism pathways in the lens: New insights into therapeutic strategies to prevent cataract formation—A review. Clin. Exp. Ophthalmol. 2020, 48, 1031–1042. [Google Scholar] [CrossRef]
- Weikel, K.A.; Garber, C.; Baburins, A.; Taylor, A. Nutritional modulation of cataract. Nutr. Rev. 2014, 72, 30–47. [Google Scholar] [CrossRef]
- Vrensen, G.F. Early cortical lens opacities: A short overview. Acta Ophthalmol. 2009, 87, 602–610. [Google Scholar] [CrossRef]
- Der Perng, M.; Sandilands, A.; Kuszak, J.; Dahm, R.; Wegener, A.; Prescott, A.R.; Quinlan, R.A. The Intermediate Filament Systems in the Eye Lens. Methods Cell Biol. 2004, 78, 597–624. [Google Scholar] [CrossRef]
- Srivastava, O.P. Age-related increase in concentration and aggregation of degraded polypeptides in human lenses. Exp. Eye Res. 1988, 47, 525–543. [Google Scholar] [CrossRef] [PubMed]
- Shiels, A.; Hejtmancik, J.F. Molecular Genetics of Cataract. Prog. Mol. Biol. Transl. Sci. 2015, 134, 203–218. [Google Scholar] [CrossRef] [PubMed]
- Hammond, C.J.; Snieder, H.; Spector, T.D.; Gilbert, C.E. Genetic and Environmental Factors in Age-Related Nuclear Cataracts in Monozygotic and Dizygotic Twins. N. Engl. J. Med. 2000, 342, 1786–1790. [Google Scholar] [CrossRef]
- Framingham Offspring Eye Study Group. Familial aggregation of lens opacities: The Framingham Eye Study and the Framingham Offspring Eye Study. Am. J. Epidemiol. 1994, 140, 555–564. [Google Scholar]
- Heiba, I.M.; Elston, R.C.; Klein, B.E.; Klein, R. Evidence for a major gene for cortical cataract. Investig. Opthalmol. Vis. Sci. 1995, 36, 227–235. [Google Scholar]
- Jun, G.; Guo, H.; Klein, B.E.K.; Klein, R.; Wang, J.J.; Mitchell, P.; Miao, H.; Lee, K.E.; Joshi, T.; Buck, M.; et al. EPHA2 Is Associated with Age-Related Cortical Cataract in Mice and Humans. PLoS Genet. 2009, 5, e1000584. [Google Scholar] [CrossRef]
- Bhagyalaxmi, S.; Srinivas, P.; Barton, K.A.; Kumar, K.R.; Vidyavathi, M.; Petrash, J.M.; Reddy, G.B.; Padma, T. A novel mutation (F71L) in αA-Crystallin with defective chaperone-like function associated with age-related cataract. Biochim. Biophys. Acta 2009, 1792, 974–981. [Google Scholar] [CrossRef]
- Bennett, T.M.; Zhou, Y.; Meyer, K.J.; Anderson, M.G.; Shiels, A. Whole-exome sequencing prioritizes candidate genes for hereditary cataract in the Emory mouse mutant. G3 2023, 13, jkad055. [Google Scholar] [CrossRef] [PubMed]
- Mura, C.V.; Gong, X.; Taylor, A.; Villalobos-Molina, R.; Scrofano, M.M. Effects of calorie restriction and aging on the expression of antioxidant enzymes and ubiquitin in the liver of Emory mice. Mech. Ageing Dev. 1996, 91, 115–129. [Google Scholar] [CrossRef] [PubMed]
- Rowan, S.; Jiang, S.; Francisco, S.G.; Pomatto, L.C.D.; Ma, Z.; Jiao, X.; Campos, M.M.; Aryal, S.; Patel, S.D.; Mahaling, B.; et al. Aged Nrf2-Null Mice Develop All Major Types of Age-Related Cataracts. Investig. Opthalmol. Vis. Sci. 2021, 62, 10. [Google Scholar] [CrossRef]
- Bennett, M.H.; Cooper, J.S. Hyperbaric Cataracts; StatPearls: Treasure Island, FL, USA, 2023. [Google Scholar]
- Borchman, D.; Giblin, F.J.; Leverenz, V.R.; Reddy, V.N.; Lin, L.R.; Yappert, M.C.; Tang, D.; Li, L. Impact of aging and hyperbaric oxygen in vivo on guinea pig lens lipids and nuclear light scatter. Investig. Opthalmol. Vis. Sci. 2000, 41, 3061–3073. [Google Scholar]
- Richter, G.M.; Torres, M.; Choudhury, F.; Azen, S.P.; Varma, R. Risk Factors for Cortical, Nuclear, Posterior Subcapsular, and Mixed Lens Opacities: The Los Angeles Latino Eye Study. Ophthalmology 2012, 119, 547–554. [Google Scholar] [CrossRef]
- Kelly, S.P.; Thornton, J.; Edwards, R.; Sahu, A.; Harrison, R. Smoking and cataract: Review of causal association. J. Cataract. Refract. Surg. 2005, 31, 2395–2404. [Google Scholar] [CrossRef]
- Kisic, B.; Miric, D.; Zoric, L.; Ilic, A.; Dragojevic, I. Antioxidant Capacity of Lenses with Age-Related Cataract. Oxidative Med. Cell. Longev. 2012, 2012, 467130. [Google Scholar] [CrossRef]
- Tsao, Y.-T.; Wu, W.-C.; Chen, K.-J.; Liu, C.-F.; Hsueh, Y.-J.; Cheng, C.-M.; Chen, H.-C. An Assessment of Cataract Severity Based on Antioxidant Status and Ascorbic Acid Levels in Aqueous Humor. Antioxidants 2022, 11, 397. [Google Scholar] [CrossRef] [PubMed]
- Nye-Wood, M.G.; Spraggins, J.M.; Caprioli, R.M.; Schey, K.L.; Donaldson, P.J.; Grey, A.C. Spatial distributions of glutathione and its endogenous conjugates in normal bovine lens and a model of lens aging. Exp. Eye Res. 2017, 154, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Tsentalovich, Y.P.; Verkhovod, T.D.; Yanshole, V.V.; Kiryutin, A.S.; Yanshole, L.V.; Fursova, A.; Stepakov, D.A.; Novoselov, V.P.; Sagdeev, R.Z. Metabolomic composition of normal aged and cataractous human lenses. Exp. Eye Res. 2015, 134, 15–23. [Google Scholar] [CrossRef]
- Grey, A.C.; Demarais, N.J.; West, B.J.; Donaldson, P.J. A quantitative map of glutathione in the aging human lens. Int. J. Mass Spectrom. 2019, 437, 58–68. [Google Scholar] [CrossRef]
- Fan, X.; Monnier, V.M.; Whitson, J. Lens glutathione homeostasis: Discrepancies and gaps in knowledge standing in the way of novel therapeutic approaches. Exp. Eye Res. 2017, 156, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Forman, H.J.; Zhang, H.; Rinna, A. Glutathione: Overview of its protective roles, measurement, and biosynthesis. Mol. Asp. Med. 2009, 30, 1–12. [Google Scholar] [CrossRef]
- Giblin, F.J.; Zhou, Y.; Bennett, T.M.; Shiels, A.; Kyei, S.; Koffuor, G.A.; Ramkissoon, P.; Abu, E.K.; Sarpong, J.F.; Zhou, Y.-F.; et al. Glutathione: A Vital Lens Antioxidant. J. Ocul. Pharmacol. Ther. 2000, 16, 121–135. [Google Scholar] [CrossRef]
- Sasaki, H.; Giblin, F.J.; Winkler, B.S.; Chakrapani, B.; Leverenz, V.; Shu, C.C. A protective role for glutathione-dependent reduction of dehydroascorbic acid in lens epithelium. Investig. Opthalmol. Vis. Sci. 1995, 36, 1805. [Google Scholar]
- Harding, J.J. Free and protein-bound glutathione in normal and cataractous human lenses. Biochem. J. 1970, 117, 957–960. [Google Scholar] [CrossRef]
- Katakura, K.; Kishida, K.; Hirano, H. Changes in rat lens proteins and glutathione reductase activity with advancing age. Int. J. Vitam. Nutr. Res. 2004, 74, 329–333. [Google Scholar] [CrossRef]
- Wei, M.; Xing, K.-Y.; Fan, Y.-C.; Libondi, T.; Lou, M.F. Loss of Thiol Repair Systems in Human Cataractous Lenses. Investig. Opthalmol. Vis. Sci. 2014, 56, 598–605. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.; Lam, Y.C.; Kistler, J.; Donaldson, P.J. Molecular Characterization of the Cystine/Glutamate Exchanger and the Excitatory Amino Acid Transporters in the Rat Lens. Investig. Opthalmol. Vis. Sci. 2005, 46, 2869–2877. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.; Lorentzen, K.A.; Kistler, J.; Donaldson, P.J. Molecular identification and characterisation of the glycine transporter (GLYT1) and the glutamine/glutamate transporter (ASCT2) in the rat lens. Exp. Eye Res. 2006, 83, 447–455. [Google Scholar] [CrossRef] [PubMed]
- Martis, R.M.; Donaldson, P.J.; Li, B.; Middleditch, M.; Kallingappa, P.K.; Lim, J.C. Mapping of the cystine–glutamate exchanger in the mouse eye: A role for xCT in controlling extracellular redox balance. Histochem. Cell Biol. 2019, 152, 293–310. [Google Scholar] [CrossRef]
- Li, B.; Li, L.; Donaldson, P.J.; Lim, J.C. Dynamic regulation of GSH synthesis and uptake pathways in the rat lens epithelium. Exp. Eye Res. 2010, 90, 300–307. [Google Scholar] [CrossRef]
- Mackic, J.B.; Jinagouda, S.; McCOMB, G.J.; Weiss, M.H.; Kannan, R.; Kaplowitz, N.; Zlokovic, B.V. Transport of Circulating Reduced Glutathione at the Basolateral Side of the Anterior Lens Epithelium: Physiologic Importance and Manipulations. Exp. Eye Res. 1996, 62, 29–38. [Google Scholar] [CrossRef]
- Stewart-DeHaan, P.J.; Dzialoszynski, T.; Trevithick, J.R. Modelling cortical cataractogenesis XXIV: Uptake by the lens of glutathione injected into the rat. Mol. Vis. 1999, 5, 37. [Google Scholar]
- Whitson, J.A.; Sell, D.R.; Goodman, M.C.; Monnier, V.M.; Fan, X. Evidence of Dual Mechanisms of Glutathione Uptake in the Rodent Lens: A Novel Role for Vitreous Humor in Lens Glutathione Homeostasis. Investig. Opthalmol. Vis. Sci. 2016, 57, 3914–3925. [Google Scholar] [CrossRef]
- Zlokovic, B.V.; Mackic, J.B.; McComb, J.; Kaplowitz, N.; Weiss, M.H.; Kannan, R. Blood-to-lens Transport of Reduced Glutathione in an In Situ Perfused Guinea-pig Eye. Exp. Eye Res. 1994, 59, 487–496. [Google Scholar] [CrossRef]
- Kinoshita, J.H.; Masurat, T. Studies on the Glutathione in Bovine Lens. Arch. Ophthalmol. 1957, 57, 266–274. [Google Scholar] [CrossRef]
- Umapathy, A.; Li, B.; Donaldson, P.J.; Lim, J.C. Functional characterisation of glutathione export from the rat lens. Exp. Eye Res. 2018, 166, 151–159. [Google Scholar] [CrossRef]
- Miller, S.P.; Arya, D.V.; Srivastava, S.K. Studies of gamma-glutamyl transpeptidase in human ocular tissues. Exp. Eye Res. 1976, 22, 329–334. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.; Jacques, P.F.; Nadler, D.; Morrow, F.; Sulsky, S.I.; Shepard, D. Relationship in humans between ascorbic acid consumption and levels of total and reduced ascorbic acid in lens, aqueous humor, and plasma. Curr. Eye Res. 1991, 10, 751–759. [Google Scholar] [CrossRef]
- Tessier, F.; Moreaux, V.; Birlouez-Aragon, I.; Junes, P.; Mondon, H. Decrease in vitamin C concentration in human lenses during cataract progression. Int. J. Vitam. Nutr. Res. 1998, 68, 309–315. [Google Scholar]
- Truscott, R.J. Age-related nuclear cataract—Oxidation is the key. Exp. Eye Res. 2005, 80, 709–725. [Google Scholar] [CrossRef] [PubMed]
- Yeum, K.J.; Taylor, A.; Tang, G.; Russell, R.M. Measurement of carotenoids, retinoids, and tocopherols in human lenses. Investig. Opthalmol. Vis. Sci. 1995, 36, 2756–2761. [Google Scholar]
- Sethna, S.S.; Holleschau, A.M.; Rathbun, W.B. Activity of Glutathione Synthesis Enzymes in Human Lens Related to Age. Curr. Eye Res. 1982, 2, 735–742. [Google Scholar] [CrossRef]
- Yan, H.; Harding, J.J.; Xing, K.; Lou, M.F. Revival of Glutathione Reductase in Human Cataractous and Clear Lens Extracts by Thioredoxin and Thioredoxin Reductase, in Conjunction with α-Crystallin or Thioltransferase. Curr. Eye Res. 2007, 32, 455–463. [Google Scholar] [CrossRef]
- Chevez-Barrios, P.; Wiseman, A.L.; Rojas, E.; Ou, C.N.; Lieberman, M.W. Cataract development in gamma-glutamyl transpeptidase-deficient mice. Exp. Eye Res. 2000, 71, 575–582. [Google Scholar] [CrossRef] [PubMed]
- Slavi, N.; Rubinos, C.; Li, L.; Sellitto, C.; White, T.W.; Mathias, R.; Srinivas, M. Connexin 46 (Cx46) Gap Junctions Provide a Pathway for the Delivery of Glutathione to the Lens Nucleus. J. Biol. Chem. 2014, 289, 32694–32702. [Google Scholar] [CrossRef]
- Vaghefi, E.; Donaldson, P.J. The lens internal microcirculation system delivers solutes to the lens core faster than would be predicted by passive diffusion. Am. J. Physiol. Integr. Comp. Physiol. 2018, 315, R994–R1002. [Google Scholar] [CrossRef]
- Wells, P.G.; Wilson, B.; Lubek, B.M. In vivo murine studies on the biochemical mechanism of naphthalene cataractogenesis. Toxicol. Appl. Pharmacol. 1989, 99, 466–473. [Google Scholar] [CrossRef]
- Calvin, H.I.; Wu, J.-X.; Viswanadhan, K.; Fu, J.S.-C. Modifications in Lens Protein Biosynthesis Signal the Initiation of Cataracts Induced by Buthionine Sulfoximine in Mice. Exp. Eye Res. 1996, 63, 357–368. [Google Scholar] [CrossRef] [PubMed]
- Calvin, H.I.; Medvedovsky, C.; David, J.C.; Broglio, T.M.; Hess, J.L.; Fu, S.C.; Worgul, B.V. Rapid deterioration of lens fibers in GSH-depleted mouse pups. Investig. Opthalmol. Vis. Sci. 1991, 32, 1916–1924. [Google Scholar]
- Calvin, H.I.; Medvedovsky, C.; Worgul, B.V. Near-Total Glutathione Depletion and Age-Specific Cataracts Induced by Buthionine Sulfoximine in Mice. Science 1986, 233, 553–555. [Google Scholar] [CrossRef]
- Mårtensson, J.; Steinherz, R.; Jain, A.; Meister, A. Glutathione ester prevents buthionine sulfoximine-induced cataracts and lens epithelial cell damage. Proc. Natl. Acad. Sci. USA 1989, 86, 8727–8731. [Google Scholar] [CrossRef]
- Carey, J.W.; Pinarci, E.Y.; Penugonda, S.; Karacal, H.; Ercal, N. In vivo inhibition of l-buthionine-(S,R)-sulfoximine-induced cataracts by a novel antioxidant, N-acetylcysteine amide. Free. Radic. Biol. Med. 2011, 50, 722–729. [Google Scholar] [CrossRef]
- Elanchezhian, R.; Sakthivel, M.; Isai, M.; Geraldine, P.; Thomas, P. Evaluation of lenticular antioxidant and redox system components in the lenses of acetyl-L-carnitine treatment in BSO-induced glutathione deprivation. Mol. Vis. 2009, 15, 1485–1491. [Google Scholar]
- Sevin, G.; Kerry, Z.; Sozer, N.; Ozsarlak-Sozer, G. Taurine supplementation protects lens against glutathione depletion. Eur. Rev. Med. Pharmacol. Sci. 2021, 25, 4520–4526. [Google Scholar] [CrossRef] [PubMed]
- Whitson, J.A.; Wilmarth, P.A.; Klimek, J.; Monnier, V.M.; David, L.; Fan, X. Proteomic analysis of the glutathione-deficient LEGSKO mouse lens reveals activation of EMT signaling, loss of lens specific markers, and changes in stress response proteins. Free. Radic. Biol. Med. 2017, 113, 84–96. [Google Scholar] [CrossRef]
- Dalton, T.P.; Dieter, M.Z.; Yang, Y.; Shertzer, H.G.; Nebert, D.W. Knockout of the Mouse Glutamate Cysteine Ligase Catalytic Subunit (Gclc) Gene: Embryonic Lethal When Homozygous, and Proposed Model for Moderate Glutathione Deficiency When Heterozygous. Biochem. Biophys. Res. Commun. 2000, 279, 324–329. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.-Z.; Osei-Frimpong, J.; Kala, G.; Kala, S.V.; Barrios, R.J.; Habib, G.M.; Lukin, D.J.; Danney, C.M.; Matzuk, M.M.; Lieberman, M.W. Glutathione synthesis is essential for mouse development but not for cell growth in culture. Proc. Natl. Acad. Sci. USA 2000, 97, 5101–5106. [Google Scholar] [CrossRef]
- Chen, Y.; Yang, Y.; Miller, M.L.; Shen, D.; Shertzer, H.G.; Stringer, K.F.; Wang, B.; Schneider, S.N.; Nebert, D.W.; Dalton, T.P. Hepatocyte-specific Gclc deletion leads to rapid onset of steatosis with mitochondrial injury and liver failure. Hepatology 2007, 45, 1118–1128. [Google Scholar] [CrossRef]
- Fan, X.; Liu, X.; Hao, S.; Wang, B.; Robinson, M.L.; Monnier, V.M. The LEGSKO Mouse: A Mouse Model of Age-Related Nuclear Cataract Based on Genetic Suppression of Lens Glutathione Synthesis. PLoS ONE 2012, 7, e50832. [Google Scholar] [CrossRef] [PubMed]
- Whitson, J.A.; Zhang, X.; Medvedovic, M.; Chen, J.; Wei, Z.; Monnier, V.M.; Fan, X. Transcriptome of the GSH-Depleted Lens Reveals Changes in Detoxification and EMT Signaling Genes, Transport Systems, and Lipid Homeostasis. Investig. Opthalmol. Vis. Sci. 2017, 58, 2666–2684. [Google Scholar] [CrossRef] [PubMed]
- Ohtsu, A.; Kitahara, S.; Fujii, K. Anticataractogenic property of gamma-glutamylcysteine ethyl ester in an animal model of cataract. Ophthalmic Res. 1991, 23, 51–58. [Google Scholar] [CrossRef]
- Zarka, M.H.; Bridge, W.J. Oral administration of gamma-glutamylcysteine increases intracellular glutathione levels above homeostasis in a randomised human trial pilot study. Redox Biol. 2017, 11, 631–636. [Google Scholar] [CrossRef]
- Akopian, D.; Rape, M. Principles of Ubiquitin-Dependent Signaling. Annu. Rev. Cell Dev. Biol. 2018, 34, 137–162. [Google Scholar] [CrossRef]
- Jahngen-Hodge, J.; Obin, M.S.; Gong, X.; Shang, F.; Nowell, T.R.; Gong, J.; Abasi, H.; Blumberg, J.; Taylor, A. Regulation of Ubiquitin-conjugating Enzymes by Glutathione Following Oxidative Stress. J. Biol. Chem. 1997, 272, 28218–28226. [Google Scholar] [CrossRef]
- Shang, F.; Taylor, A. Oxidative stress and recovery from oxidative stress are associated with altered ubiquitin conjugating and proteolytic activities in bovine lens epithelial cells. Biochem. J. 1995, 307 Pt 1, 297–303. [Google Scholar] [CrossRef]
- Zetterberg, M.; Zhang, X.; Taylor, A.; Liu, B.; Liang, J.J.; Shang, F. Glutathiolation enhances the degradation of gammaC-crystallin in lens and reticulocyte lysates, partially via the ubiquitin-proteasome pathway. Investig. Ophthalmol. Vis. Sci. 2006, 47, 3467–3473. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.C.; Arredondo, M.C.; Braakhuis, A.J.; Donaldson, P.J. Vitamin C and the Lens: New Insights into Delaying the Onset of Cataract. Nutrients 2020, 12, 3142. [Google Scholar] [CrossRef] [PubMed]
- Timoshnikov, V.A.; Kobzeva, T.V.; Polyakov, N.E.; Kontoghiorghes, G.J. Redox Interactions of Vitamin C and Iron: Inhibition of the Pro-Oxidant Activity by Deferiprone. Int. J. Mol. Sci. 2020, 21, 3967. [Google Scholar] [CrossRef] [PubMed]
- Gęgotek, A.; Skrzydlewska, E. Antioxidative and Anti-Inflammatory Activity of Ascorbic Acid. Antioxidants 2022, 11, 1993. [Google Scholar] [CrossRef]
- Waly, M.I.; Al-Attabi, Z.; Guizani, N. Low Nourishment of Vitamin C Induces Glutathione Depletion and Oxidative Stress in Healthy Young Adults. Prev. Nutr. Food Sci. 2015, 20, 198–203. [Google Scholar] [CrossRef]
- Lenton, K.J.; Sané, A.T.; Therriault, H.; Cantin, A.M.; Payette, H.; Wagner, J.R. Vitamin C augments lymphocyte glutathione in subjects with ascorbate deficiency. Am. J. Clin. Nutr. 2003, 77, 189–195. [Google Scholar] [CrossRef]
- Johnston, C.S.; Meyer, C.G.; Srilakshmi, J.C. Vitamin C elevates red blood cell glutathione in healthy adults. Am. J. Clin. Nutr. 1993, 58, 103–105. [Google Scholar] [CrossRef]
- Tram, N.K.; McLean, R.M.; Swindle-Reilly, K.E. Glutathione Improves the Antioxidant Activity of Vitamin C in Human Lens and Retinal Epithelial Cells: Implications for Vitreous Substitutes. Curr. Eye Res. 2021, 46, 470–481. [Google Scholar]
- Shang, F.; Lu, M.; Dudek, E.; Reddan, J.; Taylor, A. Vitamin C and vitamin E restore the resistance of GSH-depleted lens cells to H2O2. Free. Radic. Biol. Med. 2003, 34, 521–530. [Google Scholar] [CrossRef]
- Chiu, C.-J.; Taylor, A. Nutritional antioxidants and age-related cataract and maculopathy. Exp. Eye Res. 2007, 84, 229–245. [Google Scholar] [CrossRef]
- Dherani, M.; Murthy, G.V.S.; Gupta, S.K.; Young, I.S.; Maraini, G.; Camparini, M.; Price, G.M.; John, N.; Chakravarthy, U.; Fletcher, A.E. Blood Levels of Vitamin C, Carotenoids and Retinol Are Inversely Associated with Cataract in a North Indian Population. Investig. Opthalmol. Vis. Sci. 2008, 49, 3328–3335. [Google Scholar] [CrossRef]
- Ravindran, R.D.; Vashist, P.; Gupta, S.K.; Young, I.S.; Maraini, G.; Camparini, M.; Jayanthi, R.; John, N.; Fitzpatrick, K.E.; Chakravarthy, U.; et al. Inverse Association of Vitamin C with Cataract in Older People in India. Ophthalmology 2011, 118, 1958–1965.e2. [Google Scholar] [CrossRef]
- Taylor, A.; Jacques, P.F.; Chylack, L.T.; Hankinson, S.E.; Khu, P.M.; Rogers, G.; Friend, J.; Tung, W.; Wolfe, J.K.; Padhye, N.; et al. Long-term intake of vitamins and carotenoids and odds of early age-related cortical and posterior subcapsular lens opacities. Am. J. Clin. Nutr. 2002, 75, 540–549. [Google Scholar] [CrossRef]
- Chylack, L.T., Jr.; Brown, N.P.; Bron, A.; Hurst, M.; Köpcke, W.; Thien, U.; Schalch, W. The Roche European American Cataract Trial (REACT): A randomized clinical trial to investigate the efficacy of an oral antioxidant micronutrient mixture to slow progression of age-related cataract. Ophthalmic Epidemiol. 2002, 9, 49–80. [Google Scholar]
- Gritz, D.C.; Srinivasan, M.; Smith, S.D.; Kim, U.; Lietman, T.M.; Wilkins, J.H.; Priyadharshini, B.; John, R.K.; Aravind, S.; Prajna, N.V.; et al. The Antioxidants in Prevention of Cataracts Study: Effects of antioxidant supplements on cataract progression in South India. Br. J. Ophthalmol. 2006, 90, 847–851. [Google Scholar]
- Christen, W.G.; Glynn, R.J.; Manson, J.E.; MacFadyen, J.; Bubes, V.; Schvartz, M.; Buring, J.E.; Sesso, H.D.; Gaziano, J.M. Effects of Multivitamin Supplement on Cataract and Age-Related Macular Degeneration in a Randomized Trial of Male Physicians. Ophthalmology 2013, 121, 525–534. [Google Scholar] [CrossRef]
- Jiang, H.; Yin, Y.; Wu, C.-R.; Liu, Y.; Guo, F.; Li, M.; Ma, L. Dietary vitamin and carotenoid intake and risk of age-related cataract. Am. J. Clin. Nutr. 2019, 109, 43–54. [Google Scholar] [CrossRef]
- Fan, X.; Sell, D.R.; Hao, C.; Liu, S.; Wang, B.; Wesson, D.W.; Siedlak, S.; Zhu, X.; Kavanagh, T.J.; Harrison, F.E.; et al. Vitamin C is a source of oxoaldehyde and glycative stress in age-related cataract and neurodegenerative diseases. Aging Cell 2020, 19, e13176. [Google Scholar] [CrossRef]
- Fan, X.; Reneker, L.W.; Obrenovich, M.E.; Strauch, C.; Cheng, R.; Jarvis, S.M.; Ortwerth, B.J.; Monnier, V.M. Vitamin C mediates chemical aging of lens crystallins by the Maillard reaction in a humanized mouse model. Proc. Natl. Acad. Sci. USA 2006, 103, 16912–16917. [Google Scholar] [CrossRef]
- Smuda, M.; Henning, C.; Raghavan, C.T.; Johar, K.; Vasavada, A.R.; Nagaraj, R.H.; Glomb, M.A. Comprehensive Analysis of Maillard Protein Modifications in Human Lenses: Effect of Age and Cataract. Biochemistry 2015, 54, 2500–2507. [Google Scholar] [CrossRef]
- Rautiainen, S.; Lindblad, B.E.; Morgenstern, R.; Wolk, A. Vitamin C supplements and the risk of age-related cataract: A population-based prospective cohort study in women. Am. J. Clin. Nutr. 2010, 91, 487–493. [Google Scholar] [CrossRef] [PubMed]
- Selin, J.Z.; Rautiainen, S.; Lindblad, B.E.; Morgenstern, R.; Wolk, A. High-Dose Supplements of Vitamins C and E, Low-Dose Multivitamins, and the Risk of Age-related Cataract: A Population-based Prospective Cohort Study of Men. Am. J. Epidemiol. 2013, 177, 548–555. [Google Scholar] [CrossRef]
- Yamamoto, M.; Kensler, T.W.; Motohashi, H. The KEAP1-NRF2 System: A Thiol-Based Sensor-Effector Apparatus for Maintaining Redox Homeostasis. Physiol. Rev. 2018, 98, 1169–1203. [Google Scholar] [CrossRef] [PubMed]
- Kasai, S.; Shimizu, S.; Tatara, Y.; Mimura, J.; Itoh, K. Regulation of Nrf2 by Mitochondrial Reactive Oxygen Species in Physiology and Pathology. Biomolecules 2020, 10, 320. [Google Scholar] [CrossRef]
- Liu, X.-F.; Hao, J.-L.; Xie, T.; Malik, T.H.; Lu, C.-B.; Liu, C.; Shu, C.; Zhou, D.-D. Nrf2 as a target for prevention of age-related and diabetic cataracts by against oxidative stress. Aging Cell 2017, 16, 934–942. [Google Scholar] [CrossRef]
- Palsamy, P.; Ayaki, M.; Elanchezhian, R.; Shinohara, T. Promoter demethylation of Keap1 gene in human diabetic cataractous lenses. Biochem. Biophys. Res. Commun. 2012, 423, 542–548. [Google Scholar] [CrossRef]
- Tonelli, C.; Chio, I.I.C.; Tuveson, D.A. Transcriptional Regulation by Nrf2. Antioxid. Redox Signal. 2018, 29, 1727–1745. [Google Scholar] [CrossRef]
- Bukke, V.N.; Moola, A.; Serviddio, G.; Vendemiale, G.; Bellanti, F. Nuclear factor erythroid 2-related factor 2-mediated signaling and metabolic associated fatty liver disease. World J. Gastroenterol. 2022, 28, 6909–6921. [Google Scholar] [CrossRef]
- Tossetta, G.; Fantone, S.; Piani, F.; Crescimanno, C.; Ciavattini, A.; Giannubilo, S.R.; Marzioni, D. Modulation of NRF2/KEAP1 Signaling in Preeclampsia. Cells 2023, 12, 1545. [Google Scholar] [CrossRef]
- Audousset, C.; McGovern, T.; Martin, J.G. Role of Nrf2 in Disease: Novel Molecular Mechanisms and Therapeutic Approaches—Pulmonary Disease/Asthma. Front. Physiol. 2021, 12, 727806. [Google Scholar] [CrossRef]
- Cuadrado, A.; Rojo, A.I.; Wells, G.; Hayes, J.D.; Cousin, S.P.; Rumsey, W.L.; Attucks, O.C.; Franklin, S.; Levonen, A.-L.; Kensler, T.W.; et al. Therapeutic targeting of the NRF2 and KEAP1 partnership in chronic diseases. Nat. Rev. Drug Discov. 2019, 18, 295–317. [Google Scholar] [CrossRef]
- Ghareghomi, S.; Habibi-Rezaei, M.; Arese, M.; Saso, L.; Moosavi-Movahedi, A.A. Nrf2 Modulation in Breast Cancer. Biomedicines 2022, 10, 2668. [Google Scholar] [CrossRef]
- Elanchezhian, R.; Palsamy, P.; Madson, C.J.; Mulhern, M.L.; Lynch, D.W.; Troia, A.M.; Usukura, J.; Shinohara, T. Low glucose under hypoxic conditions induces unfolded protein response and produces reactive oxygen species in lens epithelial cells. Cell Death Dis. 2012, 3, e301. [Google Scholar] [CrossRef]
- Gong, W.; Zhu, G.; Li, J.; Yang, X. LncRNA MALAT1 promotes the apoptosis and oxidative stress of human lens epithelial cells via p38MAPK pathway in diabetic cataract. Diabetes Res. Clin. Pract. 2018, 144, 314–321. [Google Scholar] [CrossRef]
- Palsamy, P.; Bidasee, K.R.; Shinohara, T. Valproic acid suppresses Nrf2/Keap1 dependent antioxidant protection through induction of endoplasmic reticulum stress and Keap1 promoter DNA demethylation in human lens epithelial cells. Exp. Eye Res. 2014, 121, 26–34. [Google Scholar] [CrossRef]
- Wu, Z.M.; Yin, X.X.; Ji, L.; Gao, Y.Y.; Pan, Y.M.; Lu, Q.; Wang, J. Ginkgo biloba extract prevents against apoptosis induced by high glucose in human lens epithelial cells. Acta Pharmacol. Sin. 2008, 29, 1042–1050. [Google Scholar]
- Yang, S.; Yang, X.; Cao, G. Acetyl-l-carnitine prevents homocysteine-induced suppression of Nrf2/Keap1 mediated antioxidation in human lens epithelial cells. Mol. Med. Rep. 2015, 12, 1145–1150. [Google Scholar] [CrossRef]
- Zheng, X.Y.; Xu, J.; Chen, X.I.; Li, W.; Wang, T.Y. Attenuation of oxygen fluctuation-induced endoplasmic reticulum stress in human lens epithelial cells. Exp. Ther. Med. 2015, 10, 1883–1887. [Google Scholar] [CrossRef]
- Fang, W.; Ye, Q.; Yao, Y.; Xiu, Y.; Gu, F.; Zhu, Y. Protective Effects of Trimetazidine in Retarding Selenite-Induced Lens Opacification. Curr. Eye Res. 2019, 44, 1325–1336. [Google Scholar] [CrossRef]
- Palsamy, P.; Bidasee, K.R.; Shinohara, T. Selenite cataracts: Activation of endoplasmic reticulum stress and loss of Nrf2/Keap1-dependent stress protection. Biochim. Biophys. Acta. 2014, 1842, 1794–1805. [Google Scholar]
- Elanchezhian, R.; Palsamy, P.; Madson, C.J.; Lynch, D.W.; Shinohara, T. Age-related cataracts: Homocysteine coupled endoplasmic reticulum stress and suppression of Nrf2-dependent antioxidant protection. Chem. Biol. Interact. 2012, 200, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Luo, W.; Luo, X.; Yong, Z.; Zhong, X. Effects of Rosa laevigata Michx. extract on reactive oxygen species production and mitochondrial membrane potential in lens epithelial cells cultured under high glucose. Int. J. Clin. Exp. Med. 2015, 8, 15759–15765. [Google Scholar]
- Park, J.Y.; Kang, K.A.; Kim, K.C.; Cha, J.W.; Kim, E.H.; Hyun, J.W. Morin Induces Heme Oxygenase-1 via ERK-Nrf2 Signaling Pathway. J. Cancer Prev. 2013, 18, 249–256. [Google Scholar] [CrossRef]
- Gao, Y.; Yan, Y.; Huang, T. Human age-related cataracts: Epigenetic suppression of the nuclear factor erythroid 2-related factor 2-mediated antioxidant system. Mol. Med. Rep. 2014, 11, 1442–1447. [Google Scholar] [CrossRef] [PubMed]
- Lou, M.F.; Dickerson, J.E. Protein-thiol mixed disulfides in human lens. Exp. Eye Res. 1992, 55, 889–896. [Google Scholar] [CrossRef] [PubMed]
- Michael, R.; Bron, A.J. The ageing lens and cataract: A model of normal and pathological ageing. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 1278–1292. [Google Scholar] [CrossRef]
- Vrensen, G.F. UV-B and early cortical and nuclear changes in the human lens. Doc. Ophthalmol. 1995, 88, 255–261. [Google Scholar] [CrossRef]
- Rowan, S.; Jiang, S.; Chang, M.-L.; Volkin, J.; Cassalman, C.; Smith, K.M.; Streeter, M.D.; Spiegel, D.A.; Moreira-Neto, C.; Rabbani, N.; et al. A low glycemic diet protects disease-prone Nrf2-deficient mice against age-related macular degeneration. Free. Radic. Biol. Med. 2020, 150, 75–86. [Google Scholar] [CrossRef]
- Rowan, S.; Jiang, S.; Korem, T.; Szymanski, J.; Chang, M.-L.; Szelog, J.; Cassalman, C.; Dasuri, K.; McGuire, C.; Nagai, R.; et al. Involvement of a gut–retina axis in protection against dietary glycemia-induced age-related macular degeneration. Proc. Natl. Acad. Sci. USA 2017, 114, E4472–E4481. [Google Scholar] [CrossRef]
- Uchiki, T.; Weikel, K.A.; Jiao, W.; Shang, F.; Caceres, A.; Pawlak, D.; Handa, J.T.; Brownlee, M.; Nagaraj, R.; Taylor, A. Glycation-altered proteolysis as a pathobiologic mechanism that links dietary glycemic index, aging, and age-related disease (in nondiabetics). Aging Cell 2012, 11, 1–13. [Google Scholar] [CrossRef]
- Weikel, K.A.; FitzGerald, P.; Shang, F.; Caceres, M.A.; Bian, Q.; Handa, J.T.; Stitt, A.W.; Taylor, A. Natural History of Age-Related Retinal Lesions That Precede AMD in Mice Fed High or Low Glycemic Index Diets. Investig. Opthalmol. Vis. Sci. 2012, 53, 622–632. [Google Scholar] [CrossRef]
- Mura, C.V.; Roh, S.; Smith, D.; Palmer, V.; Padhye, N.; Taylor, A. Cataract incidence and analysis of lens crystallins in the water-, urea- and SDS-soluble fractions of Emory mice fed a diet restricted by 40% in calories. Curr. Eye Res. 1993, 12, 1081–1091. [Google Scholar] [CrossRef]
- Taylor, A.; Jahngen-Hodge, J.; Smith, D.E.; Palmer, V.J.; Dallal, G.E.; Lipman, R.D.; Padhye, N.; Frei, B. Dietary restriction delays cataract and reduces ascorbate levels in emory mice. Exp. Eye Res. 1995, 61, 55–62. [Google Scholar] [CrossRef]
- Taylor, A.; Lipman, R.D.; Jahngen-Hodge, J.; Palmer, V.; Smith, D.; Padhye, N.; Dallal, G.E.; Cyr, D.E.; Laxman, E.; Shepard, D.; et al. Dietary calorie restriction in the Emory mouse: Effects on lifespan, eye lens cataract prevalence and progression, levels of ascorbate, glutathione, glucose, and glycohemoglobin, tail collagen breaktime, DNA and RNA oxidation, skin integrity, fecundity, and cancer. Mech. Ageing Dev. 1995, 79, 33–57. [Google Scholar] [CrossRef]
- Taylor, A.; Zuliani, A.M.; Hopkins, R.E.; Dallal, G.E.; Treglia, P.; Kuck, J.F.R.; Kuck, K. Moderate caloric restriction delays cataract formation in the Emory mouse. FASEB J. 1989, 3, 1741–1746. [Google Scholar] [CrossRef]
- Mune, M.; Meydani, M.; Jahngen-Hodge, J.; Martin, A.; Smith, D.; Palmer, V.; Blumberg, J.B.; Taylor, A. Effect of calorie restriction on liver and kidney glutathione in aging emory mice. Age 1995, 18, 43–49. [Google Scholar] [CrossRef]
- Pomatto, L.C.D.; Dill, T.; Carboneau, B.; Levan, S.; Kato, J.; Mercken, E.M.; Pearson, K.J.; Bernier, M.; de Cabo, R. Deletion of Nrf2 shortens lifespan in C57BL6/J male mice but does not alter the health and survival benefits of caloric restriction. Free Radic. Biol. Med. 2020, 152, 650–658. [Google Scholar]
- Zhao, F.; Ci, X.; Man, X.; Li, J.; Wei, Z.; Zhang, S. Food-Derived Pharmacological Modulators of the Nrf2/ARE Pathway: Their Role in the Treatment of Diseases. Molecules 2021, 26, 1016. [Google Scholar]
- Szczesny-Malysiak, E.; Stojak, M.; Campagna, R.; Grosicki, M.; Jamrozik, M.; Kaczara, P.; Chlopicki, S. Bardoxolone Methyl Displays Detrimental Effects on Endothelial Bioenergetics, Suppresses Endothelial ET-1 Release, and Increases Endothelial Permeability in Human Microvascular Endothelium. Oxidative Med. Cell. Longev. 2020, 2020, 4678252. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bejarano, E.; Weinberg, J.; Clark, M.; Taylor, A.; Rowan, S.; Whitcomb, E.A. Redox Regulation in Age-Related Cataracts: Roles for Glutathione, Vitamin C, and the NRF2 Signaling Pathway. Nutrients 2023, 15, 3375. https://doi.org/10.3390/nu15153375
Bejarano E, Weinberg J, Clark M, Taylor A, Rowan S, Whitcomb EA. Redox Regulation in Age-Related Cataracts: Roles for Glutathione, Vitamin C, and the NRF2 Signaling Pathway. Nutrients. 2023; 15(15):3375. https://doi.org/10.3390/nu15153375
Chicago/Turabian StyleBejarano, Eloy, Jasper Weinberg, Madison Clark, Allen Taylor, Sheldon Rowan, and Elizabeth A. Whitcomb. 2023. "Redox Regulation in Age-Related Cataracts: Roles for Glutathione, Vitamin C, and the NRF2 Signaling Pathway" Nutrients 15, no. 15: 3375. https://doi.org/10.3390/nu15153375
APA StyleBejarano, E., Weinberg, J., Clark, M., Taylor, A., Rowan, S., & Whitcomb, E. A. (2023). Redox Regulation in Age-Related Cataracts: Roles for Glutathione, Vitamin C, and the NRF2 Signaling Pathway. Nutrients, 15(15), 3375. https://doi.org/10.3390/nu15153375