


Effects of Vitamin D3 Supplementation and Aerobic Training on Autophagy Signaling Proteins in a Rat Model Type 2 Diabetes Induced by High-Fat Diet and Streptozotocin


 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
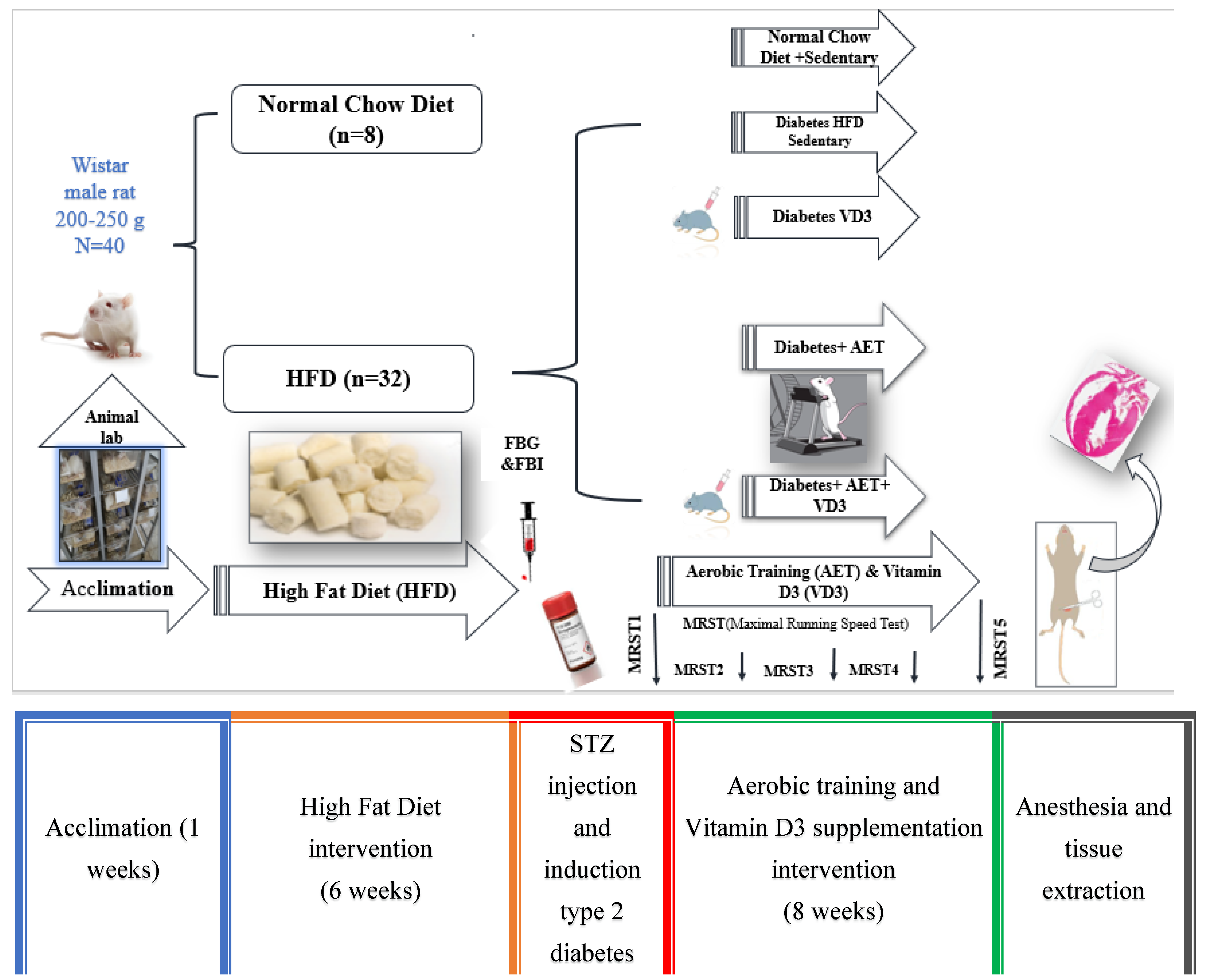
2.1. Preparation of Laboratory Animals and Their Maintenance
2.2. Induction of Diabetes Cardiomyopathy in Rat Model and Grouping Based on This
2.3. Measuring Maximum Running Speed and the Aerobic Training Protocol
2.4. Vitamin D3 Supplementation Protocol
2.5. Western Blot
2.6. Statistical Analysis
3. Results
3.1. Effects of Aerobic Training in Combination with Vitamin D3 Supplementation on Time-to Exhaustion (TTE)
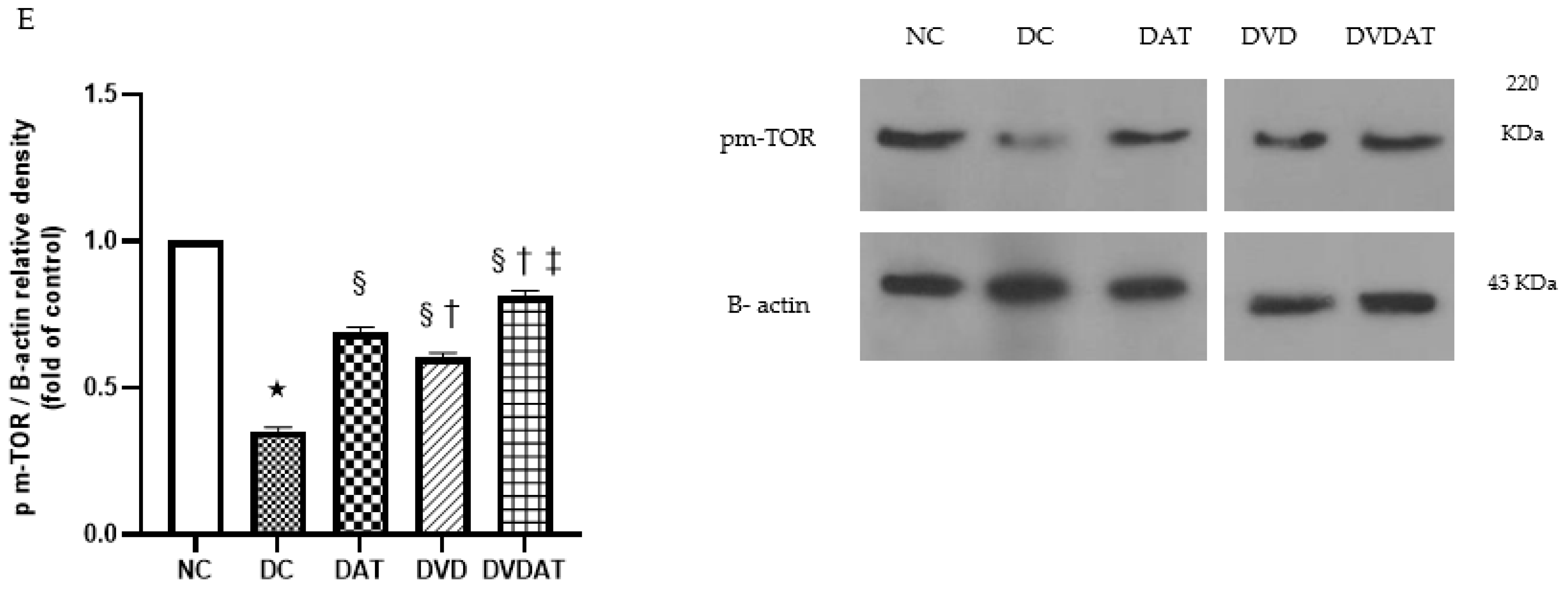
3.2. Effects of Aerobic Training along with Vitamin D3 Supplementation on p-AMPK, p-AMPK/AMPK, mTOR, and pm-TOR Proteins
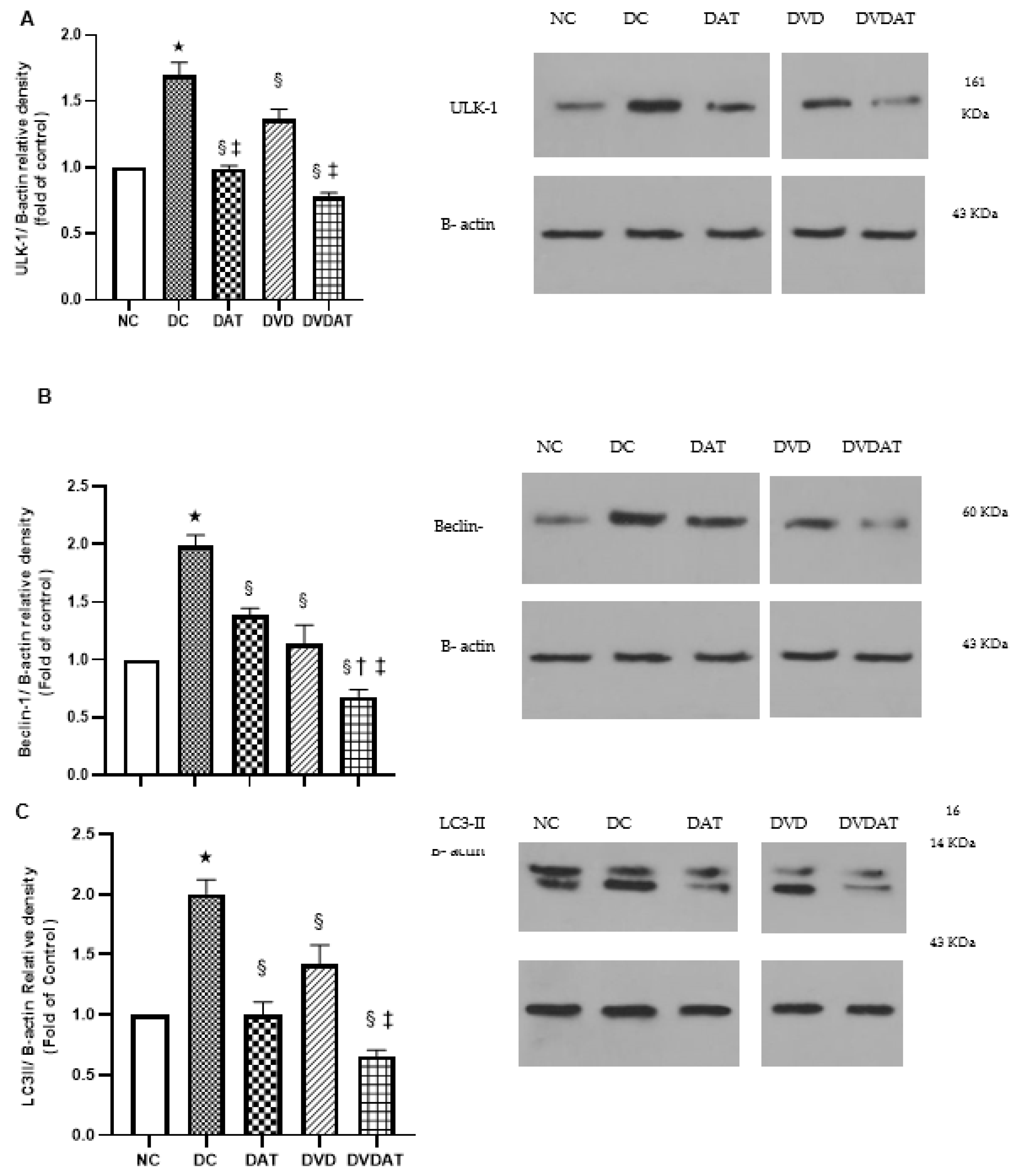
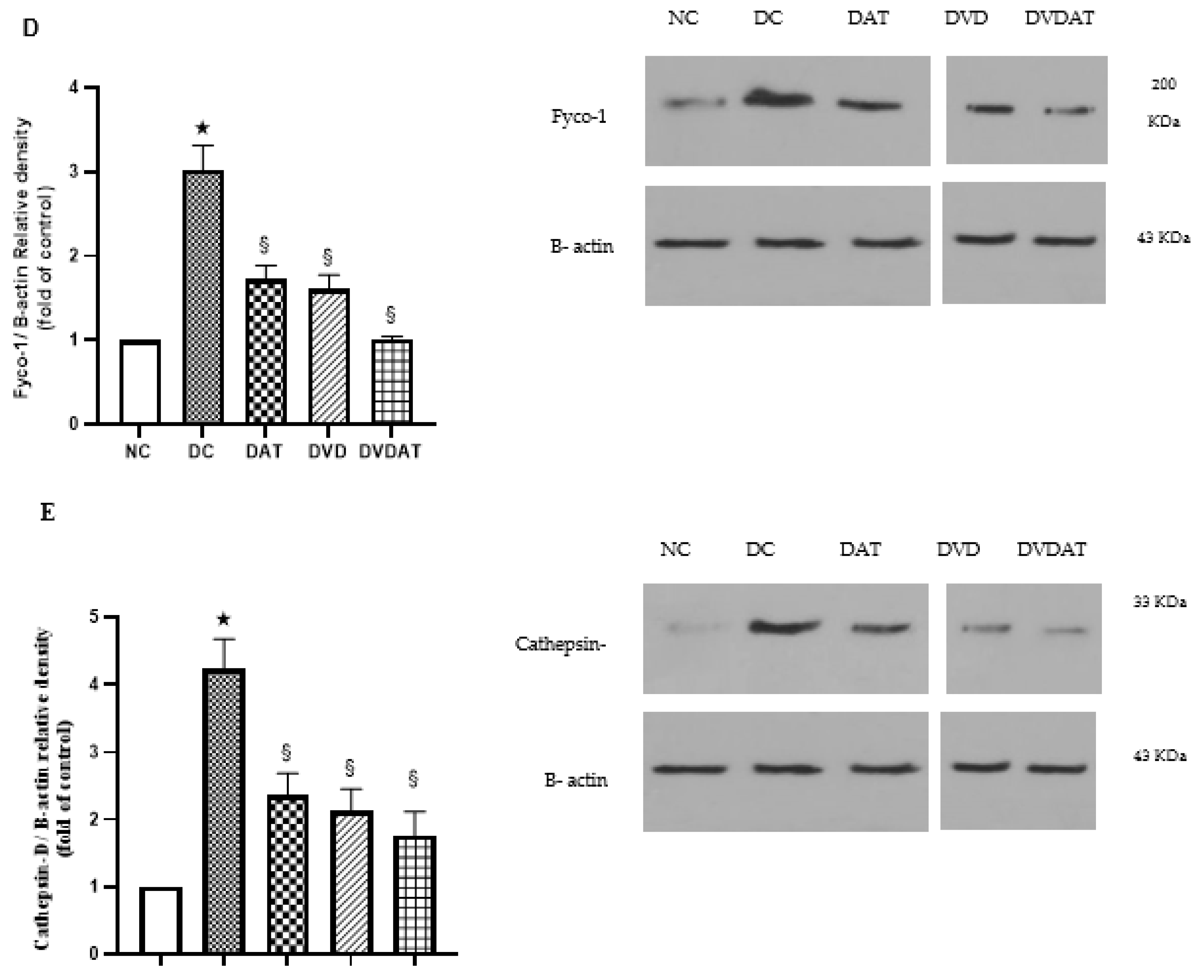
3.3. Effects of Aerobic Training along with Vitamin D3 Supplementation on ULK-1, Beclin-1, LC3II, Fyco-1 and Cathepsin D protein Levels
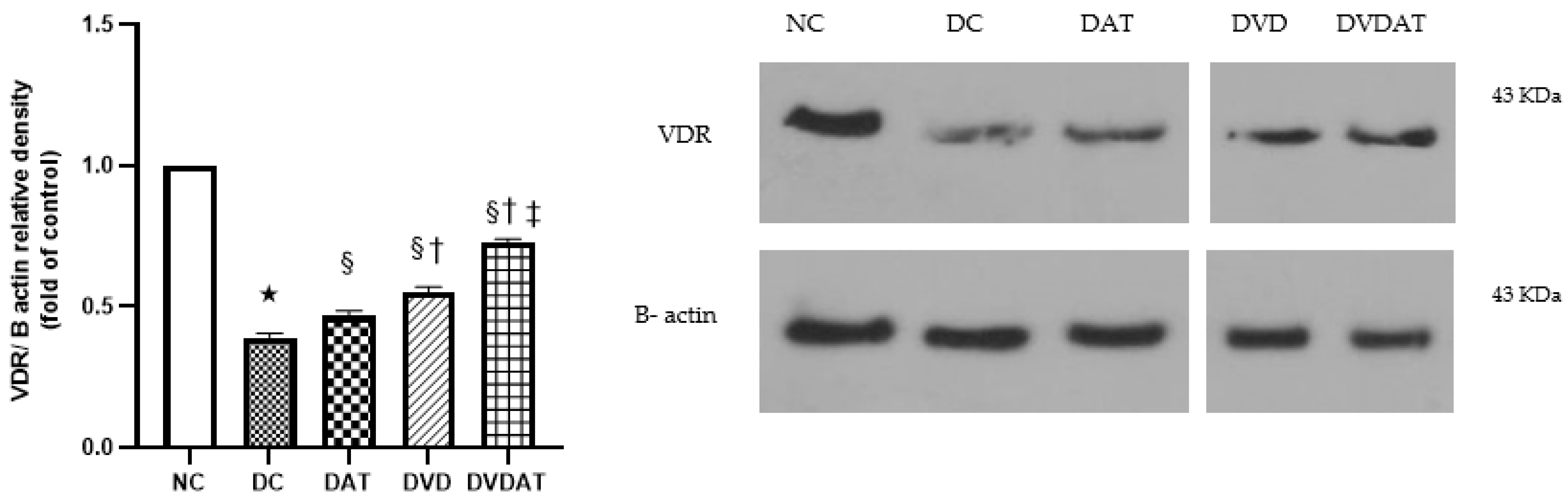
3.4. Effects of Aerobic Training along with Vitamin D3 Supplementation on VDR Protein
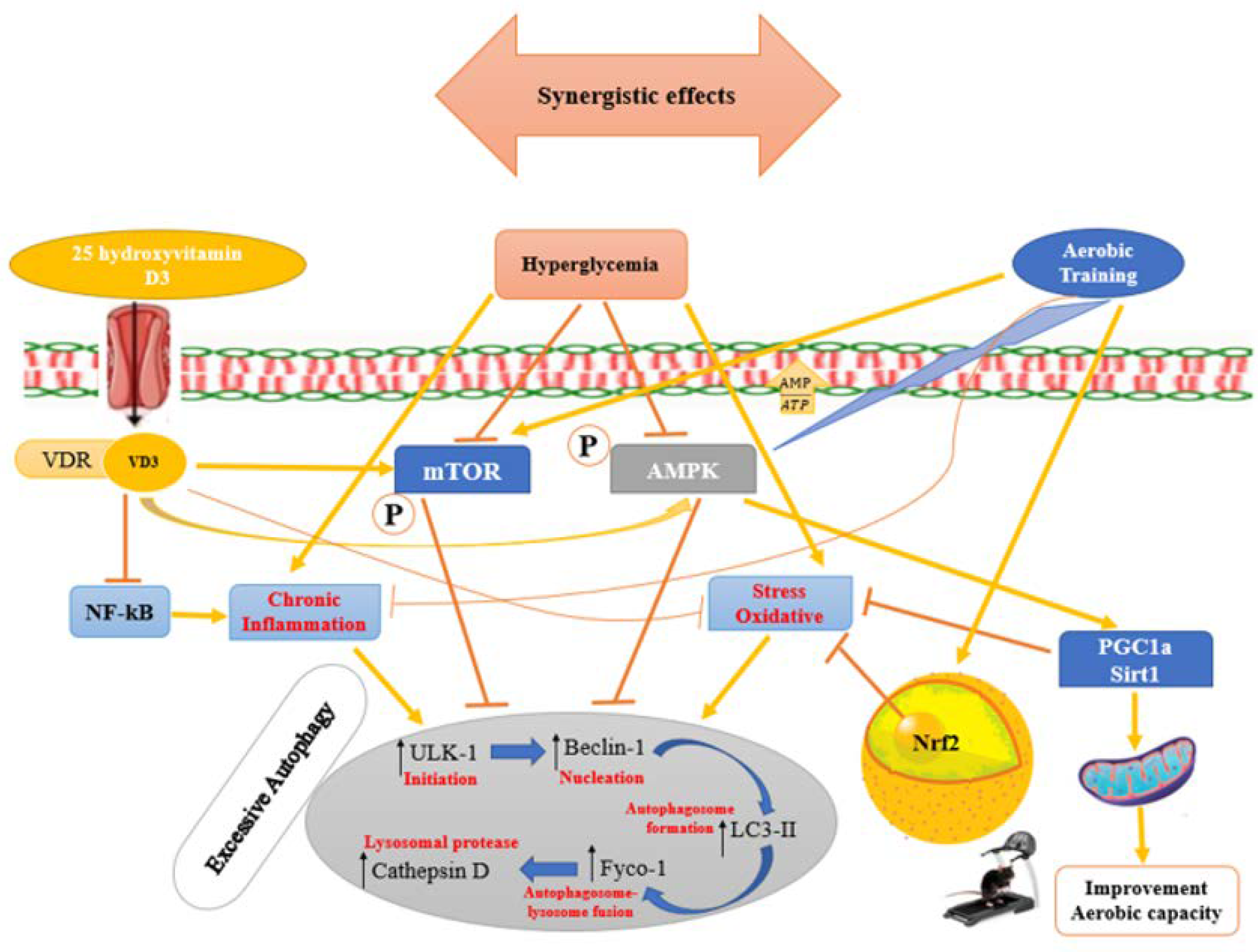
4. Discussion
4.1. Impact of Type 2 Diabetes Mellitus on Left Ventricular Heart Tissue
4.2. Effects of Aerobic Training on the Levels of Autophagy-Related Proteins in Rats with Diabetic Cardiomyopathy
4.3. Effects of Vitamin D3 on Autophagy-Related Protein Levels in Rats with Diabetic Cardiomyopathy
4.4. Effects of Vitamin D3, Aerobic Training, and Their Interaction on Cardiac Functional Capacity in T2DM Rats
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Katare, R.G.; Caporali, A.; Oikawa, A.; Meloni, M.; Emanueli, C.; Madeddu, P. Vitamin B1 Analog Benfotiamine Prevents Diabetes-Induced Diastolic Dysfunction and Heart Failure Through Akt/Pim-1–Mediated Survival Pathway. Circ. Heart Fail. 2010, 3, 294–305. [Google Scholar] [CrossRef] [PubMed]
- Plutzky, J. Macrovascular Effects and Safety Issues of Therapies for Type 2 Diabetes. Am. J. Cardiol. 2011, 108, 25B–32B. [Google Scholar] [CrossRef] [PubMed]
- Rubler, S.; Dlugash, J.; Yuceoglu, Y.Z.; Kumral, T.; Branwood, A.W.; Grishman, A. New type of cardiomyopathy associated with diabetic glomerulosclerosis. Am. J. Cardiol. 1972, 30, 595–602. [Google Scholar] [CrossRef]
- Kannel, W.B.; Hjortland, M.; Castelli, W.P. Role of diabetes in congestive heart failure: The Framingham study. Am. J. Cardiol. 1974, 34, 29–34. [Google Scholar] [CrossRef]
- Shaid, S.; Brandts, C.H.; Serve, H.; Dikic, I. Ubiquitination and selective autophagy. Cell Death Differ. 2013, 20, 21–30. [Google Scholar] [CrossRef]
- Dewanjee, S.; Vallamkondu, J.; Kalra, R.S.; John, A.; Reddy, P.H.; Kandimalla, R. Autophagy in the diabetic heart: A potential pharmacotherapeutic target in diabetic cardiomyopathy. Ageing Res. Rev. 2021, 68, 101338. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, S.; Zhou, S.; Yan, X.; Wang, Y.; Chen, J.; Mellen, N.; Kong, M.; Gu, J.; Tan, Y.; et al. Sulforaphane prevents the development of cardiomyopathy in type 2 diabetic mice probably by reversing oxidative stress-induced inhibition of LKB1/AMPK pathway. J. Mol. Cell. Cardiol. 2014, 77, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Kitada, M.; Koya, D. Autophagy in metabolic disease and ageing. Nat. Rev. Endocrinol. 2021, 17, 647–661. [Google Scholar] [CrossRef] [PubMed]
- Russell, R.C.; Tian, Y.; Yuan, H.; Park, H.W.; Chang, Y.-Y.; Kim, J.; Kim, H.; Neufeld, T.P.; Dillin, A.; Guan, K.-L. ULK1 induces autophagy by phosphorylating Beclin-1 and activating VPS34 lipid kinase. Nat. Cell Biol. 2013, 15, 741–750. [Google Scholar] [CrossRef]
- Shpilka, T.; Weidberg, H.; Pietrokovski, S.; Elazar, Z. Atg8: An autophagy-related ubiquitin-like protein family. Genome Biol. 2011, 12, 226. [Google Scholar] [CrossRef]
- Kanamori, H.; Takemura, G.; Goto, K.; Tsujimoto, A.; Mikami, A.; Ogino, A.; Watanabe, T.; Morishita, K.; Okada, H.; Kawasaki, M.; et al. Autophagic adaptations in diabetic cardiomyopathy differ between type 1 and type 2 diabetes. Autophagy 2015, 11, 1146–1160. [Google Scholar] [CrossRef]
- Marsh, B.J.; Soden, C.; Alarcón, C.; Wicksteed, B.L.; Yaekura, K.; Costin, A.J.; Morgan, G.P.; Rhodes, C.J. Regulated Autophagy Controls Hormone Content in Secretory-Deficient Pancreatic Endocrine β-cells. Mol. Endocrinol. 2007, 21, 2255–2269. [Google Scholar] [CrossRef]
- Mellor, K.; Ritchie, R.; Delbridge, L. Autophagy is Upregulated in Hearts of Insulin Resistant Mice. Heart Lung Circ. 2010, 19, S39–S40. [Google Scholar] [CrossRef]
- Munasinghe, P.E.; Riu, F.; Dixit, P.; Edamatsu, M.; Saxena, P.; Hamer, N.S.; Galvin, I.F.; Bunton, R.W.; Lequeux, S.; Jones, G.; et al. Type-2 diabetes increases autophagy in the human heart through promotion of Beclin-1 mediated pathway. Int. J. Cardiol. 2016, 202, 13–20. [Google Scholar] [CrossRef]
- Tarawan, V.M.; Gunadi, J.W.; Lesmana, R.; Goenawan, H.; Meilina, D.E.; Sipayung, J.A.; Wargasetia, T.L.; Widowati, W.; Limyati, Y.; Supratman, U. Alteration of Autophagy Gene Expression by Different Intensity of Exercise in Gastrocnemius and Soleus Muscles of Wistar Rats. J. Sports Sci. Med. 2019, 18, 146–154. [Google Scholar] [PubMed]
- He, C.; Bassik, M.C.; Moresi, V.; Sun, K.; Wei, Y.; Zou, Z.; An, Z.; Loh, J.; Fisher, J.; Sun, Q.; et al. Exercise-induced BCL2-regulated autophagy is required for muscle glucose homeostasis. Nature 2012, 481, 511–515. [Google Scholar] [CrossRef] [PubMed]
- Martinet, W.; De Meyer, G.R. Autophagy in atherosclerosis: A cell survival and death phenomenon with therapeutic potential. Circ. Res. 2009, 104, 304–317. [Google Scholar] [CrossRef]
- Jafari, A.; Zarghami Khameneh, A.; Nikookheslat, S.; Karimi, P.; Pashaei, Z. Effects of Two Months of High Intensity Interval Training and Caffeine Supplementation on the Expression of Beclin-1 and Bcl-2 Proteins in the Myocardium of Type 2 Male Diabetic Rats. J. Appl. Health Stud. Sport Physiol. 2021, 8, 83–91. [Google Scholar]
- Daneshyar, S.; Tavoosidana, G.; Bahmani, M.; Basir, S.S.; Delfan, M.; Laher, I.; Saeidi, A.; Granacher, U.; Zouhal, H. Combined effects of high fat diet and exercise on autophagy in white adipose tissue of mice. Life Sci. 2023, 314, 121335. [Google Scholar] [CrossRef]
- Dong, J.-Y.; Zhang, W.; Chen, J.J.; Zhang, Z.-L.; Han, S.-F.; Qin, L.-Q. Vitamin D Intake and Risk of Type 1 Diabetes: A Meta-Analysis of Observational Studies. Nutrients 2013, 5, 3551–3562. [Google Scholar] [CrossRef]
- Assalin, H.B.; Rafacho, B.P.; dos Santos, P.P.; Ardisson, L.P.; Roscani, M.G.; Chiuso-Minicucci, F.; Barbisan, L.F.; Fernandes, A.A.H.; Azevedo, P.S.; Minicucci, M.F.; et al. Impact of the Length of Vitamin D Deficiency on Cardiac Remodeling. Circ. Heart Fail. 2013, 6, 809–816. [Google Scholar] [CrossRef]
- Hu, F.; Yan, L.; Lu, S.; Ma, W.; Wang, Y.; Wei, Y.; Yan, X.; Zhao, X.; Chen, Z.; Wang, Z.; et al. Effects of 1, 25-Dihydroxyvitamin D3 on Experimental Autoimmune Myocarditis in Mice. Cell. Physiol. Biochem. 2016, 38, 2219–2229. [Google Scholar] [CrossRef]
- Yao, T.; Ying, X.; Zhao, Y.; Yuan, A.; He, Q.; Tong, H.; Ding, S.; Liu, J.; Peng, X.; Gao, E.; et al. Vitamin D Receptor Activation Protects Against Myocardial Reperfusion Injury Through Inhibition of Apoptosis and Modulation of Autophagy. Antioxidants Redox Signal. 2015, 22, 633–650. [Google Scholar] [CrossRef]
- Smith, A.C.; Fosse, R.T.; Ramp, W.K.; Swindle, M.M. Ethics and regulations for the care and use of laboratory animals. In Animal Models in Orthopaedic Research; CRC Press: Boca Raton, FL, USA, 2020; pp. 3–14. Available online: https://www.taylorfrancis.com/chapters/edit/10.1201/9780429173479-2/ethics-regulations-care-use-laboratory-animals-alison-smith-richard-fosse-warren-ramp-michael-swindle (accessed on 30 April 2020).
- Ali, T.M.; Abo-Salem, O.M.; El Esawy, B.H.; El Askary, A. The Potential Protective Effects of Diosmin on Streptozotocin-Induced Diabetic Cardiomyopathy in Rats. Am. J. Med. Sci. 2020, 359, 32–41. [Google Scholar] [CrossRef]
- Mansor, L.S.; Gonzalez, E.R.; Cole, M.A.; Tyler, D.J.; Beeson, J.H.; Clarke, K.; Carr, C.A.; Heather, L.C. Cardiac metabolism in a new rat model of type 2 diabetes using high-fat diet with low dose streptozotocin. Cardiovasc. Diabetol. 2013, 12, 136. [Google Scholar] [CrossRef]
- Ansari, M.; Gopalakrishnan, S.; Kurian, G.A. Streptozotocin-induced type II diabetic rat administered with nonobesogenic high-fat diet is highly susceptible to myocardial ischemia–reperfusion injury: An insight into the function of mitochondria. J. Cell. Physiol. 2019, 234, 4104–4114. [Google Scholar] [CrossRef]
- Kanter, M.; Aksu, F.; Takir, M.; Kostek, O.; Kanter, B.; Oymagil, A. Effects of Low Intensity Exercise Against Apoptosis and Oxidative Stress in Streptozotocin-induced Diabetic Rat Heart. Exp. Clin. Endocrinol. Diabetes 2016, 125, 583–591. [Google Scholar] [CrossRef]
- Bedford, T.G.; Tipton, C.M.; Wilson, N.C.; Oppliger, R.A.; Gisolfi, C.V.; Rabelo, P.C.R.; Horta, N.A.C.; Cordeiro, L.M.S.; Poletini, M.O.; Coimbra, C.C.; et al. Maximum oxygen consumption of rats and its changes with various experimental procedures. J. Appl. Physiol. 1979, 47, 1278–1283. [Google Scholar] [CrossRef] [PubMed]
- Machado, M.V.; Martins, R.L.; Borges, J.; Antunes, B.R.; Estato, V.; Vieira, A.B.; Tibiriçá, E. Exercise Training Reverses Structural Microvascular Rarefaction and Improves Endothelium-Dependent Microvascular Reactivity in Rats with Diabetes. Metab. Syndr. Relat. Disord. 2016, 14, 298–304. [Google Scholar] [CrossRef] [PubMed]
- Mehdipoor, M.; Damirchi, A.; Tousi, S.M.T.R.; Babaei, P. Concurrent vitamin D supplementation and exercise training improve cardiac fibrosis via TGF-β/Smad signaling in myocardial infarction model of rats. J. Physiol. Biochem. 2021, 77, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Qin, Y.; Wang, Y.; Meng, S.; Xian, H.; Che, H.; Lv, J.; Li, Y.; Yu, Y.; Bai, Y.; et al. Metformin Inhibits the NLRP3 Inflammasome via AMPK/mTOR-dependent Effects in Diabetic Cardiomyopathy. Int. J. Biol. Sci. 2019, 15, 1010–1019. [Google Scholar] [CrossRef]
- Yang, F.; Zhang, L.; Gao, Z.; Sun, X.; Yu, M.; Dong, S.; Wu, J.; Zhao, Y.; Xu, C.; Zhang, W.; et al. Exogenous H2S Protects Against Diabetic Cardiomyopathy by Activating Autophagy via the AMPK/mTOR Pathway. Cell. Physiol. Biochem. 2017, 43, 1168–1187. [Google Scholar] [CrossRef]
- Wang, D.; Yin, Y.; Wang, S.; Zhao, T.; Gong, F.; Zhao, Y.; Wang, B.; Huang, Y.; Cheng, Z.; Zhu, G.; et al. FGF1ΔHBS prevents diabetic cardiomyopathy by maintaining mitochondrial homeostasis and reducing oxidative stress via AMPK/Nur77 suppression. Signal Transduct. Target. Ther. 2021, 6, 133. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhang, L.; Hu, J.; Lin, J.; Wang, T.; Duan, Y.; Man, W.; Feng, J.; Sun, L.; Jia, H.; et al. MST1 coordinately regulates autophagy and apoptosis in diabetic cardiomyopathy in mice. Diabetologia 2016, 59, 2435–2447. [Google Scholar] [CrossRef]
- Kim, Y.A.; Kim, Y.S.; Oh, S.L.; Kim, H.-J.; Song, W. Autophagic response to exercise training in skeletal muscle with age. J. Physiol. Biochem. 2013, 69, 697–705. [Google Scholar] [CrossRef] [PubMed]
- Jokar, M.; Sherafati-Moghadam, M. The effect of 4 weeks high intensity interval training (HIIT) on the content of FOXO3a Beclin-1 proteins in the left ventricular heart tissue with type 2 diabetic rats. Feyz J. Kashan Univ. Med. Sci. 2020, 24, 160–169. [Google Scholar]
- Menon, M.B.; Dhamija, S. Beclin 1 Phosphorylation–at the Center of Autophagy Regulation. Front. Cell Dev. Biol. 2018, 6, 137. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Kwon, I.; Jang, Y.; Song, W.; Cosio-Lima, L.M.; Roltsch, M.H. Potential signaling pathways of acute endurance exercise-induced cardiac autophagy and mitophagy and its possible role in cardioprotection. J. Physiol. Sci. 2017, 67, 639–654. [Google Scholar] [CrossRef]
- Nilsson, P.; Loganathan, K.; Sekiguchi, M.; Matsuba, Y.; Hui, K.; Tsubuki, S.; Tanaka, M.; Iwata, N.; Saito, T.; Saido, T.C. Aβ Secretion and Plaque Formation Depend on Autophagy. Cell Rep. 2013, 5, 61–69. [Google Scholar] [CrossRef]
- Luo, J.; Yan, D.; Li, S.; Liu, S.; Zeng, F.; Cheung, C.W.; Liu, H.; Irwin, M.G.; Huang, H.; Xia, Z. Allopurinol reduces oxidative stress and activates Nrf2/p62 to attenuate diabetic cardiomyopathy in rats. J. Cell. Mol. Med. 2020, 24, 1760–1773. [Google Scholar] [CrossRef]
- Mei, Y.; Thompson, M.D.; Cohen, R.A.; Tong, X. Autophagy and oxidative stress in cardiovascular diseases. Biochim. et Biophys. Acta (BBA) Mol. Basis Dis. 2014, 1852, 243–251. [Google Scholar] [CrossRef]
- Mizushima, N.; Komatsu, M. Autophagy: Renovation of Cells and Tissues. Cell 2011, 147, 728–741. [Google Scholar] [CrossRef]
- McKnight, N.C.; Yue, Z. Beclin 1, an Essential Component and Master Regulator of PI3K-III in Health and Disease. Curr. Pathobiol. Rep. 2013, 1, 231–238. [Google Scholar] [CrossRef]
- Ohri, S.S.; Vashishta, A.; Proctor, M.; Fusek, M.; Vetvicka, V. The propeptide of cathepsin D increases proliferation, invasion and metastasis of breast cancer cells. Int. J. Oncol. 2008, 32, 491–498. [Google Scholar] [CrossRef]
- Pankiv, S.; Alemu, E.A.; Brech, A.; Bruun, J.-A.; Lamark, T.; Øvervatn, A.; Bjørkøy, G.; Johansen, T. FYCO1 is a Rab7 effector that binds to LC3 and PI3P to mediate microtubule plus end–directed vesicle transport. J. Cell Biol. 2010, 188, 253–269. [Google Scholar] [CrossRef]
- Kuhn, C.; Menke, M.; Senger, F.; Mack, C.; Dierck, F.; Hille, S.; Schmidt, I.; Brunke, G.; Bünger, P.; Schmiedel, N.; et al. FYCO1 Regulates Cardiomyocyte Autophagy and Prevents Heart Failure Due to Pressure Overload In Vivo. JACC: Basic Transl. Sci. 2021, 6, 365–380. [Google Scholar] [CrossRef]
- Diniz, T.A.; Junior, E.A.d.L.; Teixeira, A.A.; Biondo, L.A.; da Rocha, L.A.F.; Valadão, I.C.; Silveira, L.S.; Cabral-Santos, C.; de Souza, C.O.; Neto, J.C.R. Aerobic training improves NAFLD markers and insulin resistance through AMPK-PPAR-α signaling in obese mice. Life Sci. 2021, 266, 118868. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Geng, L.; Ying, L.; Shu, L.; Ye, K.; Yang, R.; Liu, Y.; Wang, Y.; Cai, Y.; Jiang, X.; et al. FGF21–Sirtuin 3 Axis Confers the Protective Effects of Exercise Against Diabetic Cardiomyopathy by Governing Mitochondrial Integrity. Circulation 2022, 146, 1537–1557. [Google Scholar] [CrossRef]
- Asokan, S.M.; Wang, T.; Wang, M.-F.; Lin, W.-T. A novel dipeptide from potato protein hydrolysate augments the effects of exercise training against high-fat diet-induced damages in senescence-accelerated mouse-prone 8 by boosting pAMPK/SIRT1/PGC-1α/pFOXO3 pathway. Aging 2020, 12, 7334–7349. [Google Scholar] [CrossRef] [PubMed]
- de Bem, G.F.; Costa, C.A.; Santos, I.B.; Cordeiro, V.d.S.C.; de Carvalho, L.C.R.M.; de Souza, M.A.V.; Soares, R.d.A.; Sousa, P.J.d.C.; Ognibene, D.T.; Resende, A.C.; et al. Antidiabetic effect of Euterpe oleracea Mart. (açaí) extract and exercise training on high-fat diet and streptozotocin-induced diabetic rats: A positive interaction. PLoS ONE 2018, 13, e0199207. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.; Park, H.; Cho, S.; Lee, M. Vitamin D3 supplementation modulates inflammatory responses from the muscle damage induced by high-intensity exercise in SD rats. Cytokine 2013, 63, 27–35. [Google Scholar] [CrossRef]
- Li, F.-H.; Li, T.; Su, Y.-M.; Ai, J.-Y.; Duan, R.; Liu, T.C.-Y. Cardiac basal autophagic activity and increased exercise capacity. J. Physiol. Sci. 2018, 68, 729–742. [Google Scholar] [CrossRef]
- Zou, M.-H.; Xie, Z. Regulation of interplay between autophagy and apoptosis in the diabetic heart. Autophagy 2013, 9, 624–625. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chen, C.; Yao, F.; Su, Q.; Liu, D.; Xue, R.; Dai, G.; Fang, R.; Zeng, J.; Chen, Y.; et al. AMPK inhibits cardiac hypertrophy by promoting autophagy via mTORC1. Arch. Biochem. Biophys. 2014, 558, 79–86. [Google Scholar] [CrossRef]
- Byrne, N.J.; Rajasekaran, N.S.; Abel, E.D.; Bugger, H. Therapeutic potential of targeting oxidative stress in diabetic cardiomyopathy. Free. Radic. Biol. Med. 2021, 169, 317–342. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wang, C.; Yan, F.; Wang, T.; He, Y.; Li, H.; Xia, Z.; Zhang, Z. N-Acetylcysteine Attenuates Diabetic Myocardial Ischemia Reperfusion Injury through Inhibiting Excessive Autophagy. Mediat. Inflamm. 2017, 2017, 9257291. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, J.; Cretoiu, D.; Li, G.; Xiao, J. Exercise-mediated regulation of autophagy in the cardiovascular system. J. Sport Health Sci. 2020, 9, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Haye, A.; Ansari, M.A.; Rahman, S.O.; Shamsi, Y.; Ahmed, D.; Sharma, M. Role of AMP-activated protein kinase on cardio-metabolic abnormalities in the development of diabetic cardiomyopathy: A molecular landscape. Eur. J. Pharmacol. 2020, 888, 173376. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, Y.; Liu, X.; Yuan, X.; Xiang, M.; Liu, J.; Zhang, L.; Zhu, S.; Lu, J.; Tang, Q.; et al. Exercise and Metformin Intervention Prevents Lipotoxicity-Induced Hepatocyte Apoptosis by Alleviating Oxidative and ER Stress and Activating the AMPK/Nrf2/HO-1 Signaling Pathway in db/db Mice. Oxidative Med. Cell. Longev. 2022, 2022, 2297268. [Google Scholar] [CrossRef]
- Abbaszadeh, S.; Yadegari, P.; Imani, A.; Taghdir, M. Vitamin D3 protects against lead-induced testicular toxicity by modulating Nrf2 and NF-κB genes expression in rat. Reprod. Toxicol. 2021, 103, 36–45. [Google Scholar] [CrossRef] [PubMed]
- McMillan, E.M.; Paré, M.-F.; Baechler, B.L.; Graham, D.A.; Rush, J.W.E.; Quadrilatero, J. Autophagic Signaling and Proteolytic Enzyme Activity in Cardiac and Skeletal Muscle of Spontaneously Hypertensive Rats following Chronic Aerobic Exercise. PLoS ONE 2015, 10, e011938. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, R.-Y. Effects of high-load exercise induced skeletal muscle injury on autophagy ultrastructure and Beclin1 and LC3-II / I in rats. Chin. J. Appl. Physiol. 2020, 36, 296–300. [Google Scholar]
- Chang, Y.; Ma, X. Effects of high intensity training on rat cardiomyocyte autophagy. J. Sci. Med. Sport 2017, 20, e118. [Google Scholar] [CrossRef]
- Mejías-Peña, Y.; Estébanez, B.; Rodriguez-Miguelez, P.; Fernandez-Gonzalo, R.; Almar, M.; de Paz, J.A.; González-Gallego, J.; Cuevas, M.J. Impact of resistance training on the autophagy-inflammation-apoptosis crosstalk in elderly subjects. Aging 2017, 9, 408–418. [Google Scholar] [CrossRef] [PubMed]
- Jung, C.H.; Jun, C.B.; Ro, S.-H.; Kim, Y.-M.; Otto, N.M.; Cao, J.; Kundu, M.; Kim, D.-H. ULK-Atg13-FIP200 Complexes Mediate mTOR Signaling to the Autophagy Machinery. Mol. Biol. Cell 2009, 20, 1992–2003. [Google Scholar] [CrossRef] [PubMed]
- Mirsepasi, M.; Banaeifar, A.; Azarbayjani, M.-A.; Arshadi, S. The Effect of 12 Weeks Aerobic Training on Expression of AKT1 and mTORc1 genes in the Left Ventricle of Type 2 Diabetic Rats. Iran. J. Diabetes Obes. 2018, 10, 137–143. [Google Scholar]
- Kwon, I.; Song, W.; Jang, Y.; Choi, M.D.; Vinci, D.M.; Lee, Y. Elevation of hepatic autophagy and antioxidative capacity by endurance exercise is associated with suppression of apoptosis in mice. Ann. Hepatol. 2020, 19, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Ghareghani, P.; Shanaki, M.; Ahmadi, S.; Khoshdel, A.R.; Rezvan, N.; Meshkani, R.; Delfan, M.; Gorgani-Firuzjaee, S. Aerobic endurance training improves nonalcoholic fatty liver disease (NAFLD) features via miR-33 dependent autophagy induction in high fat diet fed mice. Obes. Res. Clin. Pract. 2018, 12, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Zhang, H.; Zeng, Z.; Wu, L.; Zhang, Y.; Guo, Y.; Lv, J.; Wang, C.; Fan, J.; Chen, N. Lifelong Aerobic Exercise Alleviates Sarcopenia by Activating Autophagy and Inhibiting Protein Degradation via the AMPK/PGC-1α Signaling Pathway. Metabolites 2021, 11, 323. [Google Scholar] [CrossRef] [PubMed]
- Kang, E.-B.; Cho, J.-Y. Effect of treadmill exercise on PI3K/AKT/mTOR, autophagy, and Tau hyperphosphorylation in the cerebral cortex of NSE/htau23 transgenic mice. J. Exerc. Nutr. Biochem. 2015, 19, 199–209. [Google Scholar] [CrossRef]
- Gunadi, J.W.; Tarawan, V.M.; Setiawan, I.; Lesmana, R.; Wahyudianingsih, R.; Supratman, U. Cardiac hypertrophy is stimulated by altered training intensity and correlates with autophagy modulation in male Wistar rats. BMC Sports Sci. Med. Rehabilitation 2019, 11, 9. [Google Scholar] [CrossRef]
- Wu, Y.; Li, X.; Zhu, J.X.; Xie, W.; Le, W.; Fan, Z.; Jankovic, J.; Pan, T. Resveratrol-Activated AMPK/SIRT1/Autophagy in Cellular Models of Parkinson’s Disease. Neurosignals 2011, 19, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Manna, P.; Achari, A.E.; Jain, S.K. Vitamin D supplementation inhibits oxidative stress and upregulate SIRT1/AMPK/GLUT4 cascade in high glucose-treated 3T3L1 adipocytes and in adipose tissue of high fat diet-fed diabetic mice. Arch. Biochem. Biophys. 2017, 615, 22–34. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Lee, H.; Lim, Y. Vitamin D3 improves lipophagy-associated renal lipid metabolism and tissue damage in diabetic mice. Nutr. Res. 2020, 80, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Li, H.-X.; Gao, J.-M.; Liang, J.-Q.; Xi, J.-M.; Fu, M.; Wu, Y.-J. Vitamin D3potentiates the growth inhibitory effects of metformin in DU145 human prostate cancer cells mediated by AMPK/mTOR signalling pathway. Clin. Exp. Pharmacol. Physiol. 2015, 42, 711–717. [Google Scholar] [CrossRef]
- Aragón-Herrera, A.; Feijóo-Bandín, S.; Santiago, M.O.; Barral, L.; Campos-Toimil, M.; Gil-Longo, J.; Pereira, T.M.C.; García-Caballero, T.; Rodríguez-Segade, S.; Rodríguez, J.; et al. Empagliflozin reduces the levels of CD36 and cardiotoxic lipids while improving autophagy in the hearts of Zucker diabetic fatty rats. Biochem. Pharmacol. 2019, 170, 113677. [Google Scholar] [CrossRef]
- Lim, H.; Lee, H.; Lim, Y. Effect of vitamin D3 supplementation on hepatic lipid dysregulation associated with autophagy regulatory AMPK/Akt-mTOR signaling in type 2 diabetic mice. Exp. Biol. Med. 2021, 246, 1139–1147. [Google Scholar] [CrossRef]
- Lee, T.-W.; Chang, C.-J.; Lien, G.-S.; Kao, Y.-H.; Chao, T.-F.; Chen, Y.-J. Potential of vitamin D in treating diabetic cardiomyopathy. Nutr. Res. 2015, 35, 269–279. [Google Scholar] [CrossRef]
- Dai, J.; Liang, Y.; Li, H.; Zhou, W.; Wang, B.; Gong, A.; Zhang, R. Vitamin D enhances resistance to aspergillus fumigatus in mice via inhibition of excessive autophagy. Am. J. Transl. Res. 2018, 10, 381–391. [Google Scholar]
- Guo, X.; Lin, H.; Liu, J.; Wang, D.; Li, D.; Jiang, C.; Tang, Y.; Wang, J.; Zhang, T.; Li, Y.; et al. 1,25-Dihydroxyvitamin D attenuates diabetic cardiac autophagy and damage by vitamin D receptor-mediated suppression of FoxO1 translocation. J. Nutr. Biochem. 2020, 80, 108380. [Google Scholar] [CrossRef]
- Li, A.; Yi, B.; Han, H.; Yang, S.; Hu, Z.; Zheng, L.; Wang, J.; Liao, Q.; Zhang, H. Vitamin D-VDR (vitamin D receptor) regulates defective autophagy in renal tubular epithelial cell in streptozotocin-induced diabetic mice via the AMPK pathway. Autophagy 2021, 18, 877–890. [Google Scholar] [CrossRef] [PubMed]
- Tavera-Mendoza, L.E.; Westerling, T.; Libby, E.; Marusyk, A.; Cato, L.; Cassani, R.; Cameron, L.A.; Ficarro, S.B.; Marto, J.A.; Klawitter, J.; et al. Vitamin D receptor regulates autophagy in the normal mammary gland and in luminal breast cancer cells. Proc. Natl. Acad. Sci. USA 2017, 114, E2186–E2194. [Google Scholar] [CrossRef]
- Huang, D.; Guo, Y.; Li, X.; Pan, M.; Liu, J.; Zhang, W.; Mai, K. Vitamin D3/VDR inhibits inflammation through NF-κB pathway accompanied by resisting apoptosis and inducing autophagy in abalone Haliotis discus hannai. Cell Biol. Toxicol. 2021, 39, 885–906. [Google Scholar] [CrossRef]
- Wu, S.; Sun, J. Vitamin D, vitamin D receptor, and macroautophagy in inflammation and infection. Discov. Med. 2011, 11, 325–335. [Google Scholar] [PubMed]
- Bhutia, S.K. Vitamin D in autophagy signaling for health and diseases: Insights on potential mechanisms and future perspectives. J. Nutr. Biochem. 2021, 99, 108841. [Google Scholar] [CrossRef] [PubMed]
- Chung, C.; Silwal, P.; Kim, I.; Modlin, R.L.; Jo, E.-K. Vitamin D-Cathelicidin Axis: At the Crossroads between Protective Immunity and Pathological Inflammation during Infection. Immune Netw. 2020, 20, e12. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Wang, K.; Zhang, J.; Cao, Z.-B. Aerobic Exercise Ameliorates Myocardial Fibrosis via Affecting Vitamin D Receptor and Transforming Growth Factor-β1 Signaling in Vitamin D-Deficient Mice. Nutrients 2023, 15, 741. [Google Scholar] [CrossRef]
- Kobayashi, S.; Xu, X.; Chen, K.; Liang, Q. Suppression of autophagy is protective in high glucose-induced cardiomyocyte injury. Autophagy 2012, 8, 577–592. [Google Scholar] [CrossRef]
- Khodir, S.A.; Samaka, R.M.; Ameen, O. Autophagy and mTOR Pathways Mediate the Potential Renoprotective Effects of Vitamin D on Diabetic Nephropathy. Int. J. Nephrol. 2020, 2020, 7941861. [Google Scholar] [CrossRef]
- Li, Q.; Wang, W.; Ma, Q.; Xia, R.; Gao, B.; Zhu, G.; Wang, J. Moxibustion Improves Chronic Heart Failure by Inhibiting Autophagy and Inflammation via Upregulation of mTOR Expression. Evidence-Based Complement. Altern. Med. 2021, 2021, 6635876. [Google Scholar] [CrossRef]
- Song, B.; Yan, X.; Li, R.; Zhang, H. Ghrelin ameliorates chronic obstructive pulmonary disease–associated infllammation and autophagy. Biotechnol. Appl. Biochem. 2020, 68, 356–365. [Google Scholar] [CrossRef]
- Zha, L.; Wu, G.; Xiao, H.; Xiao, Y. Vitamin D Attenuates Airway Inflammation in Asthmatic Guinea Pigs Using Mammalian Target of Rapamycin-Mediated Autophagy. J. Interf. Cytokine Res. 2022, 42, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, N. Autophagy Is a Promoter for Aerobic Exercise Performance during High Altitude Training. Oxidative Med. Cell. Longev. 2018, 2018, 3617508. [Google Scholar] [CrossRef] [PubMed]
- Hooper, P.L.; Balogh, G.; Rivas, E.; Kavanagh, K.; Vigh, L. The importance of the cellular stress response in the pathogenesis and treatment of type 2 diabetes. Cell Stress Chaperon 2014, 19, 447–464. [Google Scholar] [CrossRef] [PubMed]
- Ruderman, N.B.; Xu, X.J.; Nelson, L.; Cacicedo, J.M.; Saha, A.K.; Lan, F.; Ido, Y. AMPK and SIRT1: A long-standing partnership? Am. J. Physiol. Endocrinol. Metab. 2010, 298, E751–E760. [Google Scholar] [CrossRef] [PubMed]
- Karbasforooshan, H.; Karimi, G. The role of SIRT1 in diabetic cardiomyopathy. BioMedicine 2017, 90, 386–392. [Google Scholar] [CrossRef] [PubMed]
- Valle, M.S.; Russo, C.; Malaguarnera, L. Protective role of vitamin D against oxidative stress in diabetic retinopathy. Diabetes/Metabolism Res. Rev. 2021, 37, e3447. [Google Scholar] [CrossRef]
- Li, W.; Yang, Q.; Mao, Z. Chaperone-mediated autophagy: Machinery, regulation and biological consequences. Cell. Mol. Life Sci. 2010, 68, 749–763. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Num. Week | MRS T1 | First Week | Secound Week | MRS T2 | Third Week | Fourth Week | MRS T3 | Fifth Week | Sixth Week | MRS T4 | Seventh Week | Eight Week | MRS T5 |
| Intensity (% MRS T) | 55% | 55% | 55% | 60% | 60% | 60% | 65% | 65% | |||||
| Duration (min) | 30 | 35 | 40 | 45 | 50 | 55 | 60 | 60 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Golpasandi, H.; Rahimi, M.R.; Ahmadi, S.; Łubkowska, B.; Cięszczyk, P. Effects of Vitamin D3 Supplementation and Aerobic Training on Autophagy Signaling Proteins in a Rat Model Type 2 Diabetes Induced by High-Fat Diet and Streptozotocin. Nutrients 2023, 15, 4024. https://doi.org/10.3390/nu15184024
Golpasandi H, Rahimi MR, Ahmadi S, Łubkowska B, Cięszczyk P. Effects of Vitamin D3 Supplementation and Aerobic Training on Autophagy Signaling Proteins in a Rat Model Type 2 Diabetes Induced by High-Fat Diet and Streptozotocin. Nutrients. 2023; 15(18):4024. https://doi.org/10.3390/nu15184024
Chicago/Turabian StyleGolpasandi, Hadi, Mohammad Rahman Rahimi, Slahadin Ahmadi, Beata Łubkowska, and Paweł Cięszczyk. 2023. "Effects of Vitamin D3 Supplementation and Aerobic Training on Autophagy Signaling Proteins in a Rat Model Type 2 Diabetes Induced by High-Fat Diet and Streptozotocin" Nutrients 15, no. 18: 4024. https://doi.org/10.3390/nu15184024
APA StyleGolpasandi, H., Rahimi, M. R., Ahmadi, S., Łubkowska, B., & Cięszczyk, P. (2023). Effects of Vitamin D3 Supplementation and Aerobic Training on Autophagy Signaling Proteins in a Rat Model Type 2 Diabetes Induced by High-Fat Diet and Streptozotocin. Nutrients, 15(18), 4024. https://doi.org/10.3390/nu15184024