
Renoprotective Effects of Solid-State Cultivated Antrodia cinnamomea in Juvenile Rats with Chronic Kidney Disease

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Solid-State Cultures of Antrodia camphorata
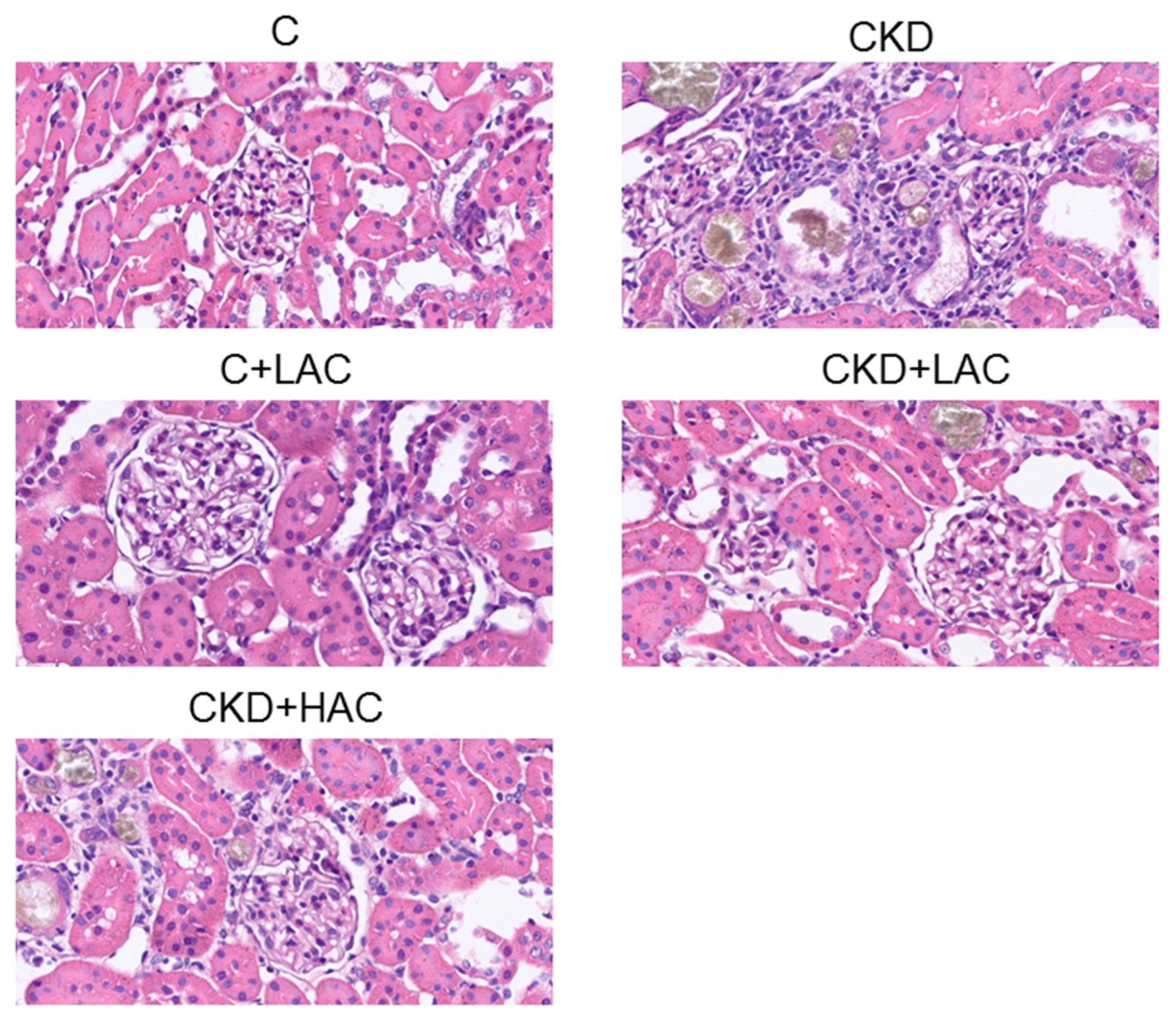
2.2. Animal Model
2.3. Measurement of Creatinine Concentration by HPLC
2.4. Measurement of SCFAs by GC-MS
2.5. Measurement of TMAO by LC–MS
2.6. 16S rRNA Gene Sequencing and Analysis
2.7. Analysis of Renal Gene Expression by qPCR
2.8. Statistics
3. Results
3.1. Anthropometrics, Renal Outcome, and Blood Pressure
3.2. Plasma SCFA Levels and Renal SCFA Receptors
3.3. DMA, TMA, and TMAO
3.4. Gut Microbiota Composition
3.5. RAS
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wasser, S.P. Medicinal mushroom science: Current perspectives, advances, evidences, and challenges. Biomed. J. 2014, 7, 345–356. [Google Scholar] [CrossRef]
- Ganesan, N.; Baskaran, R.; Velmurugan, B.K.; Thanh, N.C. Antrodia cinnamomea-An updated minireview of its bioactive components and biological activity. J. Food Biochem. 2019, 43, e12936. [Google Scholar] [CrossRef] [PubMed]
- Li, H.X.; Wang, J.J.; Lu, C.L.; Gao, Y.J.; Gao, L.; Yang, Z.Q. Review of Bioactivity, Isolation, and Identification of Active Compounds from Antrodia cinnamomea. Bioengineering 2022, 9, 494. [Google Scholar] [CrossRef] [PubMed]
- Tain, Y.L.; Hsu, C.N. Developmental Origins of Chronic Kidney Disease: Should We Focus on Early Life? Int. J. Mol. Sci. 2017, 18, 381. [Google Scholar] [CrossRef] [PubMed]
- James, S.L.; Abate, D.; Abate, K.H.; Abay, S.M.; Abbafati, C.; Abbasi, N.; Abdollahpour, I. Global, regional, and national incidence, prevalence, and years lived with disability for 354 diseases and injuries for 195 countries and territories, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2018, 392, 1789–1858. [Google Scholar] [CrossRef] [PubMed]
- Ingelfinger, J.R.; Kalantar-Zadeh, K.; Schaefer, F.; World Kidney Day Steering Committee. World Kidney Day 2016: Averting the legacy of kidney disease-focus on childhood. Pediatr. Nephrol. 2016, 31, 343–348. [Google Scholar] [CrossRef]
- Hsu, C.N.; Hou, C.Y.; Chang, C.I.; Tain, Y.L. Resveratrol Butyrate Ester Protects Adenine-Treated Rats against Hypertension and Kidney Disease by Regulating the Gut-Kidney Axis. Antioxidants 2021, 11, 83. [Google Scholar] [CrossRef]
- Yang, T.; Richards, E.M.; Pepine, C.J.; Raizada, M.K. The gut microbiota and the brain-gut-kidney axis in hypertension and chronic kidney disease. Nat. Rev. Nephrol. 2018, 14, 442–456. [Google Scholar] [CrossRef]
- Hobby, G.P.; Karaduta, O.; Dusio, G.F.; Singh, M.; Zybailov, B.L.; Arthur, J.M. Chronic kidney disease and the gut microbiome. Am. J. Physiol.-Ren. Physiol. 2019, 316, F1211–F1217. [Google Scholar] [CrossRef]
- Hsu, C.N.; Chang-Chien, G.P.; Lin, S.; Hou, C.Y.; Tain, Y.L. Targeting on Gut Microbial Metabolite Trimethylamine-N-Oxide and Short-Chain Fatty Acid to Prevent Maternal High-Fructose-Diet-Induced Developmental Programming of Hypertension in Adult Male Offspring. Mol. Nutr. Food Res. 2019, 63, e1900073. [Google Scholar] [CrossRef]
- Al Khodor, S.; Shatat, I.F. Gut microbiome and kidney disease: A bidirectional relationship. Pediatr. Nephrol. 2017, 32, 921–931. [Google Scholar] [CrossRef]
- Liu, X.; Yu, Z.; Jia, W.; Wu, Y.; Wu, D.; Zhang, H.; Liu, Z.; Yang, Y.; Zhang, J.; Liu, Y.; et al. A Review on Linking the Medicinal Functions of Mushroom Prebiotics with Gut Microbiota. Int. J. Med. Mushrooms 2020, 22, 943–951. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.Z.; Liang, Y.C.; Lin, S.Y.; Lin, Y.S.; Wu, W.C.; Hou, W.C.; Su, C.H. Antihypertensive activities of a solid-state culture of Taiwanofungus camphoratus (Chang-chih) in spontaneously hypertensive rats. Biosci. Biotechnol. Biochem. 2007, 71, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.C.; Yang, C.H.; Grumezescu, A.M.; Lin, Y.M.; Huang, K.S.; Wang, W.T.; Su, H.Y.; Jhang, C.Y.; Chung, R.Y.; Chou, J.H. Renoprotective Effects of Shout Camphor Medicinal Mushroom (Taiwanofungus camphorates, Basidiomycetes) Mycelia on Several Media in Mice with Chronic Kidney Disease. Int. J. Med. Mushrooms 2016, 18, 1105–1114. [Google Scholar] [CrossRef] [PubMed]
- Chung, C.H.; Yeh, S.C.; Chen, C.J.; Lee, K.T. Coenzyme Q0 from Antrodia cinnamomea in Submerged Cultures Induces Reactive Oxygen Species-Mediated Apoptosis in A549 Human Lung Cancer Cells. Evid.-Based Complement. Altern. Med. 2014, 2014, 246748. [Google Scholar] [CrossRef] [PubMed]
- Reckelhoff, J.F. Gender differences in the regulation of blood pressure. Hypertension 2001, 37, 1199–1208. [Google Scholar] [CrossRef]
- Tain, Y.L.; Freshour, G.; Dikalova, A.; Griendling, K.; Baylis, C. Vitamin E reduces glomerulosclerosis, restores renal neuronal NOS, and suppresses oxidative stress in the 5/6 nephrectomized rat. Am. J. Physiol.-Ren. Physiol. 2007, 292, F1404–F1410. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Diwan, V.; Brown, L.; Gobe, G.C. Adenine-induced chronic kidney disease in rats. Nephrology 2018, 23, 5–11. [Google Scholar] [CrossRef]
- Xu, J.; Moore, B.N.; Pluznick, J.L. Short-Chain Fatty Acid Receptors and Blood Pressure Regulation: Council on Hypertension Mid-Career Award for Research Excellence 2021. Hypertension 2022, 79, 2127–2137. [Google Scholar] [CrossRef]
- Tang, W.H.; Wang, Z.; Kennedy, D.J.; Wu, Y.; Buffa, J.A.; Agatisa-Boyle, B.; Li, X.S.; Levison, B.S.; Hazen, S.L. Gut microbiota-dependent trimethylamine N-oxide (TMAO) pathway contributes to both development of renal insufficiency and mortality risk in chronic kidney disease. Circ. Res. 2015, 116, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Morris, E.K.; Caruso, T.; Buscot, F.; Fischer, M.; Hancock, C.; Maier, T.S.; Meiners, T.; Müller, C.; Obermaier, E.; Prati, D.; et al. Choosing and using diversity indices: Insights for ecological applications from the German Biodiversity Exploratories. Ecol. Evol. 2014, 4, 3514–3524. [Google Scholar] [CrossRef] [PubMed]
- Geethangili, M.; Tzeng, Y.M. Review of Pharmacological Effects of Antrodia camphorata and Its Bioactive Compounds. Evid.-Based Complement. Alternat. Med. 2011, 2011, 212641. [Google Scholar] [CrossRef] [PubMed]
- Fusco, W.; Lorenzo, M.B.; Cintoni, M.; Porcari, S.; Rinninella, E.; Kaitsas, F.; Lener, E.; Mele, M.C.; Gasbarrini, A.; Collado, M.C.; et al. Short-Chain Fatty-Acid-Producing Bacteria: Key Components of the Human Gut Microbiota. Nutrients 2023, 15, 2211. [Google Scholar] [CrossRef]
- Xie, D.; Zhang, M.; Wang, B.; Lin, H.; Wu, E.; Zhao, H.; Li, S. Differential Analysis of Hypertension-Associated Intestinal Microbiota. Int. J. Med. Sci. 2019, 16, 872–881. [Google Scholar]
- Palmu, J.; Lahti, L.; Niiranen, T. Targeting Gut Microbiota to Treat Hypertension: A Systematic Review. Int. J. Environ. Res. Public Health 2021, 18, 1248. [Google Scholar] [CrossRef]
- Mukohda, M.; Yano, T.; Matsui, T.; Nakamura, S.; Miyamae, J.; Toyama, K.; Mitsui, R.; Mizuno, R.; Ozaki, H. Treatment with Ligilactobacillus murinus lowers blood pressure and intestinal permeability in spontaneously hypertensive rats. Sci. Rep. 2023, 13, 15197. [Google Scholar] [CrossRef]
- Guerrero Sanchez, M.; Passot, S.; Campoy, S.; Olivares, M.; Fonseca, F. Ligilactobacillus salivarius functionalities, applications, and manufacturing challenges. Appl. Microbiol. Biotechnol. 2022, 106, 57–80. [Google Scholar] [CrossRef]
- Te Riet, L.; van Esch, J.H.; Roks, A.J.; van den Meiracker, A.H.; Danser, A.H. Hypertension: Renin-angiotensin-aldosterone system alterations. Circ. Res. 2015, 116, 960–975. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward | Reverse |
|---|---|---|
| GPR41 | 5 tcttcaccaccgtctatctcac 3 | 5 cacaagtcctgccaccctc 3 |
| GPR43 | 5 ctgcctgggatcgtctgtg 3 | 5 cataccctcggccttctgg 3 |
| GPR109A | 5 cggtggtctactatttctcc 3 | 5 cccctggaatacttctgatt 3 |
| OLFR78 | 5 gaggaagctcacttttggtttgg 3 | 5 cagcttcaatgtccttgtcacag 3 |
| Renin | 5 aacattaccagggcaactttcact 3 | 5 acccccttcatggtgatctg 3 |
| PRR | 5 gaggcagtgaccctcaacat 3 | 5 ccctcctcacacaacaaggt 3 |
| ACE | 5 caccggcaaggtctgctt 3 | 5 cttggcatagtttcgtgaggaa 3 |
| AT1R | 5 gctgggcaacgagtttgtct 3 | 5 cagtccttcagctggatcttca 3 |
| R18S | 5 gccgcggtaattccagctcca 3 | 5 cccgcccgctcccaagatc 3 |
| Group | C | CKD | C + LAC | CKD + LAC | CKD + HAC |
|---|---|---|---|---|---|
| Mortality | 0% | 0% | 12.5% | 0% | 0% |
| Body weight (BW) (g) | 225 ± 9 b | 266 ± 6 a | 212 ± 7 b | 276 ± 7 a | 248 ± 9 a |
| Left kidney weight (KW) (g) | 1.16 ± 0.05 c | 1.65 ± 0.05 b | 1.1 ± 0.04 c | 1.98 ± 0.12 a | 1.69 ± 0.1 a |
| Left KW/100 g BW | 0.52 ± 0.01 b | 0.62 ± 0.01 a | 0.52 ± 0.01 b | 0.72 ± 0.05 a | 0.69 ± 0.04 a |
| Creatinine (μM/L) | 11 ± 10.6 b | 16.4 ± 1.1 a | 12.1 ± 0.9 b | 11.7 ± 0.2 b | 14.1 ± 0.7 a |
| CCr (ml/min) | 2.2 ± 0.2 a | 1.5 ± 0.2 b | 3.6 ± 0.5 a | 3.2 ± 0.2 a | 2.4 ± 0.3 a |
| 24 h urinary protein (mg/100 g BW) | 36.4 ± 2.2 b | 51.3 ± 7.8 a | 34.3 ± 2.1 b | 34 ± 3.7 b | 41.9 ± 4.3 a |
| Glomerular injury score | 0.2 ± 0.1 b | 1.9 ± 0.5 a | 0.3 ± 0.2 b | 0.4 ± 0.4 b | 0.5 ± 0.4 a |
| Tubulointerstitial injury score | 0.2 ± 0.2 c | 2.5 ± 1.1 a | 0.2 ± 0.1 c | 0.6 ± 0.5 b | 0.8 ± 0.4 b |
| Group | C | CKD | C + LAC | CKD + LAC | CKD + HAC |
|---|---|---|---|---|---|
| Acetic acid (μM) | 641 ± 51 b | 639 ± 39 b | 1240 ± 57 a | 636 ± 25 b | 734 ± 34 b |
| Propionic acid (μM) | 4.2 ± 0.3 b | 4.2 ± 0.2 b | 8.6 ± 0.5 a | 4.1 ± 0.1 b | 4.6 ± 0.3 b |
| Isobutyric acid (μM) | 1.0 ± 0.2 b | 0.9 ± 0.1 b | 1.9 ± 0.3 a | 0.9 ± 0.1 b | 1.4 ± 0.2 a |
| Butyric acid (μM) | 10.4 ± 1.1 b | 10.6 ± 0.6 b | 21.5 ± 1.2 a | 10.3 ± 0.3 b | 11.9 ± 0.6 b |
| Isovaleric acid (μM) | 22.5 ± 1.6 b | 22.5 ± 0.8 b | 46.4 ± 1.8 a | 21.7 ± 0.6 b | 23.6 ± 0.9 b |
| Valeric acid (μM) | 1.8 ± 0.4 | 1.9 ± 0.2 | 2.9 ± 0.5 | 1.7 ± 0.1 | 1.6 ± 0.4 |
| Group | C | CKD | C + LAC | CKD + LAC | CKD + HAC |
|---|---|---|---|---|---|
| DMA | 85 ± 7 | 89 ± 2 | 103 ± 11 | 94 ± 4 | 86 ± 3 |
| TMA | 5 ± 0.6 | 3.5 ± 0.2 | 5.3 ± 1 | 4.2 ± 0.5 | 3.4 ± 0.4 |
| TMAO | 264 ± 11 | 246 ± 8 | 269 ± 14 | 284 ± 24 | 280 ± 15 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tain, Y.-L.; Chang-Chien, G.-P.; Lin, S.; Hou, C.-Y.; Hsu, C.-N. Renoprotective Effects of Solid-State Cultivated Antrodia cinnamomea in Juvenile Rats with Chronic Kidney Disease. Nutrients 2023, 15, 4626. https://doi.org/10.3390/nu15214626
Tain Y-L, Chang-Chien G-P, Lin S, Hou C-Y, Hsu C-N. Renoprotective Effects of Solid-State Cultivated Antrodia cinnamomea in Juvenile Rats with Chronic Kidney Disease. Nutrients. 2023; 15(21):4626. https://doi.org/10.3390/nu15214626
Chicago/Turabian StyleTain, You-Lin, Guo-Ping Chang-Chien, Sufan Lin, Chih-Yao Hou, and Chien-Ning Hsu. 2023. "Renoprotective Effects of Solid-State Cultivated Antrodia cinnamomea in Juvenile Rats with Chronic Kidney Disease" Nutrients 15, no. 21: 4626. https://doi.org/10.3390/nu15214626