Abstract
The ketogenic diet (KD) is a high-fat, low-carbohydrate diet that has been reported to have neuroprotective effects. The health effects of KD might be linked to an altered gut microbiome, which plays a major role in host health, leading to neuroprotective effects via the gut-brain axis. However, results from different studies, most often based on the 16S rRNA gene and metagenome sequencing, have been inconsistent. In this study, we assessed the effect of a 4-week KD compared to a western diet (WD) on the colonic microbiome of female C57Bl/6J mice by analyzing fecal samples using fluorescence in situ hybridization. Our results showed distinct changes in the total number of gut bacteria following the 4-week KD, in addition to changes in the composition of the microbiome. KD-fed mice showed higher absolute numbers of Actinobacteria (especially Bifidobacteria spp.) and lower absolute levels of Proteobacteria, often linked to gut inflammation, in comparison with WD-fed mice. Furthermore, an increased abundance of the typically rare genus Atopobium was observed. These changes may indicate the possible anti-inflammatory effects of the KD. However, since the overall changes in the microbiota seem low, the KD effects might be linked to the differential abundance of only a few key genera in mice.
1. Introduction
The ketogenic diet (KD) is characterized by high-fat and low-carbohydrate dietary intake, leading to a state of ketosis [1]. It has been mainly used as a therapeutic approach for pharmaco-resistant epilepsy in children [2], and it is increasingly considered for other neurological disorders [3]. Ingestion of KD forces the body to utilize fatty acids instead of glucose as its primary energy source, resulting in restricted glycolysis, increased fatty acid oxidation, and, ultimately, a significant increase in the generation of ketone bodies. As a consequence of the elevation of fat-derived ketones, a metabolic switch can occur, in which the body obtains energy from the metabolism of ketone bodies. In addition to these metabolic alterations, KD has also been shown to modulate the levels of hormones, neurotransmitters, and neuropeptides and to affect key signaling pathways, such as PPARs, AMP-activated kinase, and mammalian target of rapamycin [1,4,5]. Furthermore, several studies have indicated that KD has anti-inflammatory effects and profoundly affects mitochondria by stimulating mitochondrial biogenesis, improving mitochondrial function, and decreasing oxidative stress [6]. The mechanisms implicated in the neuroprotective effects of KD are particularly those involving the aforementioned alterations in cellular energy metabolism and increased mitochondrial activity due to this diet [7]. Moreover, KD and ketone bodies are increasingly being studied for their therapeutic efficacy in different non-neurological disorders, including obesity, non-alcoholic fatty liver disease, heart failure, and cancer [7]. KD is applied in a variety of protocols that differ in caloric and macronutrient contents. For instance, the classical ketogenic diet, which is normocaloric, has been used to treat epilepsy in children. Other forms of ketogenic diet differ in fat-to-protein and carbohydrate ratios, and these include modified Atkins diet, low glycemic index treatment, and medium-chain triglyceride diet [8]. KD has also been used as a weight loss strategy [9] and to reduce insulin resistance in both type 1 and type 2 diabetes [10,11]. In this context, KD can be adapted to be vegetarian by including plant proteins rather than animal proteins, giving diabetic or obese patients the flexibility to choose their preferred diets. However, other studies have reported the side effects of KD on renal function and lipid profiles due to its high content of protein and fat [11].
Interestingly, the therapeutic potential of KD to improve seizure activity has been linked to its effect on the gut microbiota and related metabolites, although KD has been reported to reduce gut microbial diversity [12,13]. The gut microbiota has been described as an endocrine organ. It has a major impact on host health and is associated with several diseases [14]. The gut microbiota is highly affected by dietary intake, which plays a major role in modulating host metabolism in addition to shaping the gut microbiome [15,16]. Gut bacteria play an important role in digestion in the gastrointestinal tract, and commensal bacteria are pivotal for the synthesis and absorption of nutrients and metabolites such as lipids, amino acids, bile acids, vitamins, and short-chain fatty acids (SCFAs). SCFAs result from the fermentation of dietary fiber and resistant starch by the gut bacteria [17,18]. Furthermore, studies have reported the crucial role of the gut microbiota in modulating the homeostasis and function of innate and adaptive immune cells [19]. It is important to mention that the gut microbiota functions differently in different individuals due to the unique bacterial combination in each individual, which is related to inter-and intra-individual variation in humans [18]. Studies in humans [20] and animals [21] have shown that KD is able to alter the function of the gut microbiota in health [16]. Ang et al. reported that KD-associated gut microbiota is able to reduce the accumulation of Th17 cells, indicating an anti-inflammatory role of KD [16]. In addition, Kong et al. reported that fecal microbial transplantation from donors with KD was able to alleviate colitis in DSS-treated mice recipients [12]. The authors suggested that their results showed that the anti-inflammatory effects of KD may open the door to a therapeutic approach for IBD patients [12]. Moreover, an animal study by Olson et al. found that specific KD-associated bacteria were responsible for the anti-seizure effects. This effect was achieved by the KD-associated microbiota being able to modulate amino acid γ-glutamylation and hippocampal GABA/glutamate [21]. Furthermore, KD could lead to genetic variation within the gut microbiome, and the microbiome diversity could be altered by increasing the ratio of Bacteroidetes to Firmicutes [22]. However, more studies are still needed for long-term (>2 years), medium-term (>6 months –2 years), and short-term (<3 months) KD studies are essential to investigate its effect on the microbiome and determine whether it is directly related to the resulting weight loss or not [8].
Most studies on KD have used 16S rRNA gene sequencing or metagenome sequencing to assess the relative abundance of bacteria, and the results from different studies, most often based on the 16S rRNA gene and metagenome sequencing, have been inconsistent. This difference is due to the fact that metagenomics is able to detect less abundant taxa, and sometimes, these less abundant genera could be more biologically meaningful than the highly abundant ones that are only detected by 16S rRNA sequencing [23]. For characterizing microbial changes and showing bacteria and host interactions, high-throughput sequencing techniques have emerged to be the most important. Nevertheless, data interpretation from these techniques is based on relative abundance, and it disregards the absolute abundance or total bacterial load. In specific cases, such as addressing biological problems like community interactions, absolute abundance is necessary. As such, absolute abundance is considered more important than relative abundance, and the interpretation of microbiota data based on only relative abundance can be ambiguous. The different approaches to absolute quantification are diverse. For instance, the absolute quantification of specific taxa can be calculated by multiplying the relative abundance of the taxa generated by the 16S rRNA amplicon sequencing by the total cell count [24]. However, bias could potentially occur when using different absolute and relative quantification tools, such as 16S rRNA copy number discrepancy and qPCR primer specificity [24,25]. Another PCR-independent method that can directly enumerate specific taxa is fluorescence in situ hybridization (FISH), which uses a fluorescent probe to hybridize complementary sequences in the target cells. Due to its high sensitivity, FISH is able to detect and quantify low-abundance microbes [24]. It can be used to calculate relative taxon abundance, and in an optimized methodological approach like flow cytometry or microscopy, it can even estimate absolute taxon abundance [24,25,26]. The aim of the present study was to determine the differences in gut microbiota between healthy C57/BL6J mice fed a Western diet (WD), and those fed a KD, using fluorescence in situ hybridization (FISH) to study the absolute and relative abundance of different bacterial taxa.
2. Materials and Methods
2.1. Mice and Dietary Intervention
C57BL/6J mice (N = 20, n = 10/group) were maintained under standard conditions in a specific pathogen-free environment at 12 h day-night cycles according to the FELASA recommendations with food and water ad libitum. Following a one-week acclimatization period, female mice were divided randomly into homogeneous groups according to their weight and age (10–12 weeks of age) and fed an ad libitum Western or ketogenic diet for four weeks. The Western diet contained 18% protein, 59% carbohydrates, and 23% fat in terms of energy content. The ketogenic diet contained 15% protein, 1.4% carbohydrates, and 83.6% fat. Health status, such as the observation of behavior, physical assessment, and body weight, was monitored twice per week throughout the intervention. At the end of the dietary intervention, fecal samples from individual mice were fixed in modified Carnoy’s solution (ethanol:glacial acetic acid: chloroform, 6:6:1, v:v:v) for at least 24 h at room temperature [27]. Fecal samples were embedded in paraffin, and a 4 µm thick section was cut from each sample for fluorescence in-situ hybridization (FISH). To evaluate the ketotic state of diet-fed mice, blood concentrations of β-hydroxybutyrate, as an indicator of ketone body levels as well as blood glucose levels, were measured at the end of the experiment using a ketone and glucose meter.
The experiments were approved by the State Animal Care Committee (Landesamt für Gesundheit und Soziales, Berlin, Germany; approval code G0047/15) and performed according to the guidelines for the care and use of laboratory animals adopted by the U.S. National Institutes of Health, and the ARRIVE guidelines.
2.2. Fluorescence In-Situ Hybridization (FISH)
The Fecal microbiota was investigated using FISH analysis with ribosomal RNA-derived probes. Hybridization was performed on 4 µM thick sections of Carnoy-fixed and paraffin-embedded stool samples [28]. The samples were collected after 4 weeks of feeding.
The bacteria were quantified using group-specific C3 probes. The FITC-marked universal probe was used in each hybridization to evaluate the number of all bacteria, and C5 marked probes with a different specificity to C3 probes were used to determine the spatial relationship between different bacterial groups. Only signals that hybridized with a specific FISH probe and the universal FISH probe that did not hybridize with specific FISH probes from unrelated bacterial groups were evaluated [29].
Bacterial concentrations in homogeneous populations were enumerated visually in one of the 10 × 10 fields of the ocular raster, corresponding to 10 µm × 10 µm of the section surface at a magnification of ×1000. This number was assigned to a concentration of ×109 bacteria/mL, which was equivalent to the formula used previously [28].
In the case of uneven distribution of bacteria over the microscopic field, the positive signals were enumerated in 10 fields of the ocular raster along the gradient of distribution, and an average was used after dividing by 10.
2.3. Investigated Bacterial Groups and FISH Probes
A total of 48 bacterial FISH probes were applied. Seven of these probes (ACI623, Bcv13b, CAP365, EUB338 II, Pce, Phasco741, Veil223) were excluded from the analysis because the bacteria detected by these probes were not specific to the mouse intestine. The names of the FISH probes are listed according to abbreviations of the probeBase online resource (https://probebase.csb.univie.ac.at (accessed on 16 February 2022)) [30]. Details of the FISH probe specificity and hybridization conditions are provided in probeBase.
The probes in Table 1 were ordered alphabetically into subgroups according to their abundance and specificity, as described in the Results section.

Table 1.
Applied FISH probes.
2.4. Statistical Analysis
Statistical analyses were performed using the GraphPad Prism 8.1.2 software (San Diego, CA, USA). Since some data were not normally distributed, statistical significance was determined using the non-parametric Mann-Whitney U test. Differences were considered statistically significant at p < 0.05. The data are expressed as individual values ± SEM.
3. Results
3.1. Efficacy of the Ketogenic Diet in Mice
Mice were fed for 4 weeks with a WD in the control group and with KD in the treatment group. As expected, a significant reduction in blood glucose concentration and a significant increase in ketone levels were observed in the KD-fed group compared with those in the WD-fed group (Figure 1a,b). However, no significant change in body weight was observed in either group (Figure 1c).
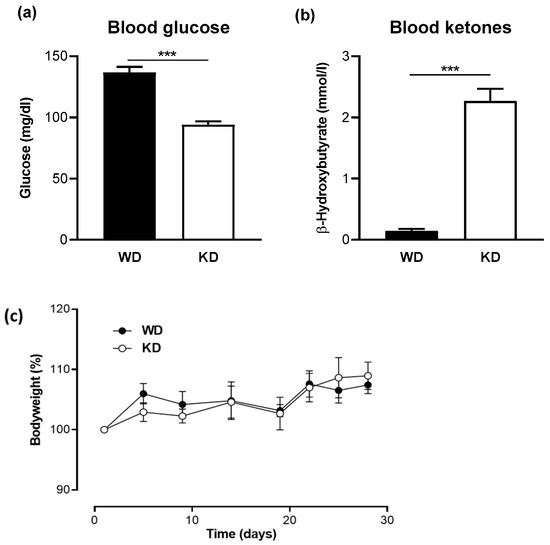
Figure 1.
A Ketogenic diet leads to low glucose and high ketone levels in the blood. (a) Blood glucose levels and (b) blood β-hydroxybutyrate concentrations in mice fed a western diet (WD) or a ketogenic diet (KD) for four weeks (n = 10/group). Data are represented as mean ± SEM. Statistical significance was assessed using the Mann-Whitney U test (*** p < 0.001). (c) No significant change in body weight was observed between the WD- and KD-fed mice.
3.2. Eligibile FISH Probes Used for Analysis of the Stool Microbiome
Depending on their prevalence and abundance, the investigated bacterial groups were divided into two categories: “substantial” and “marginal.” Substantial bacterial groups were found in at least 20% of the fecal samples at concentrations higher than 109 bacteria/mL. Marginal bacterial groups showed low occurrence as well as marginal concentrations. They occurred in less than 20% of fecal samples at concentrations below 109 bacteria/mL and, in most cases, lower than 108 bacteria/mL. According to this classification, 19 of the FISH probes represented substantial bacterial groups, and 22 of the probes presented marginal bacterial groups (Table 1). Because the marginal bacterial groups did not significantly contribute to the overall colonic biomass, they were excluded from the subsequent evaluation. This specific definition and division of bacterial groups, depending on their prevalence, were previously introduced and described by Swidsinski et al. [31].
The bacteria were subdivided into highly conserved and individual bacterial groups. Three bacteria detected using CF319a (most Flavobacteria, some Bacteroidetes), CFB560 (subgroup of Bacteroidetes, CFB division), and MIB661 (mouse intestinal bacteria) probes were consistently present in the fecal samples of mice fed the WD and KD at concentrations between 10 × 109 and 25 × 109 bacteria/mL, contributing to approximately half of the colonic microbiota in each mouse. The invariability and predominance of these three bacterial groups in C57BL/6J mice contributed to a major part of the microbiome and were designated as highly conserved bacteria. All other substantial bacterial groups were present only in a subset of the mice and were designated as individual bacteria. As their concentrations reached 1010 bacteria/mL, they contributed substantially to the fecal biomass, comprising approximately 50% of the biomass.
3.3. Effect of the Ketogenic Diet on the Fecal Microbiome in Mice
The fecal microbiota of WD- and KD-fed C57BL/6J mice were analyzed by FISH using 41 previously described probes (Table 1).
Feeding mice with the KD decreased the overall number of bacteria (Figure 2a). This observation was also reflected by the reduction in the total number of highly conserved bacterial groups (Table 2). The total numbers of the marginal bacterial group and all individual bacteria were not changed by the KD (Table 2). The decrease in the highly conserved bacterial groups in the stool samples of KD-fed mice was mainly due to a significant reduction in bacteria detected by the MIB661 probe, specific for several Bacteroides species. Bacteria detected by the CF319a and CFB560 probes had similarly high concentrations in mice fed the KD and WD.
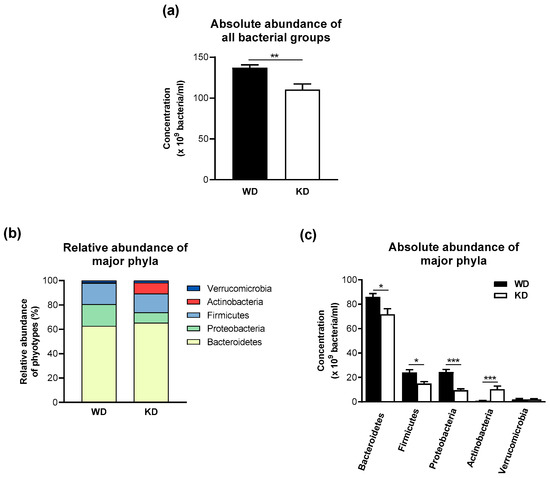
Figure 2.
The ketogenic diet-induced moderate compositional changes and lower numbers of bacteria in the fecal microbiota of wild-type mice. (a) Total bacterial number, (b) relative abundance of major phyla, and (c) absolute microbiota composition at the phylum level. Data are represented as mean ± SEM (n = 10/group). Statistical significance was assessed using the Mann-Whitney U test (* p < 0.05, ** p < 0.01, *** p < 0.001).
Table 2.
Mean microbial concentrations ± SD (×109 bacteria/mL) of all substantial bacterial groups in stool samples of mice fed a Western diet (WD) and a ketogenic diet (KD). The increase (↑) or decrease (↓) in bacterial % between the two groups is indicated.
Although the total number of individual bacterial subgroups did not change in the KD-fed group, the most distinct changes in single bacteria were found in this group, with statistically significant differences found for 7 of the 16 investigated individual bacterial groups (Table 2). The most abundant individual bacterial groups in WD-fed mice were Burkholderia cepacia (Burcep probe), followed by the Eubacterium hallii group (Ehal1469 probe), and Bacteroides putredinis (Bputre698 probe), and the numbers of these three bacterial groups were lower in KD-fed mice (Table 2). In addition, the number of Sphingomonas/Erythrobacter (SPH492 probe) was lower in the KD group (Table 2). The dominant individual bacterial groups in the KD-fed mice were Bacteroides distasonis (Bdis656 probe), Eubacterium cylindroides (Ecyl387 probe), and Bacteroides spp. (Bac303 probe) (Table 2). Three of the investigated individual bacterial groups showed an increase in KD-fed as compared to WD-fed mice by approximately 200 to 4600%: Atopobium cluster (Ato291 probe), Bifidobacterium spp. (Bif662 probe), and the Eubacterium cylindroides group (Ecyl387 probe) (Table 2).
As mentioned before, differences in single marginal bacterial groups were not statistically evaluated between WD- and KD-fed mice.
The three most abundant phyla in the stool of WD-fed mice were Bacteroidetes (62.8%), Proteobacteria (17.8%), and Firmicutes (17.3%), whereas Verrucomicrobia (1.5%) and Actinobacteria (0.6%) were only present in a minor proportion in the stool samples of mice consuming the WD (Figure 2b). In contrast, the KD-fed group showed a higher relative abundance of Actinobacteria (9.1%) and a lower abundance of Proteobacteria (8.5%) (Figure 2b). These compositional differences were confirmed at the level of absolute numbers, as KD-fed mice showed a higher absolute abundance of Actinobacteria and a lower absolute abundance of the phylum Proteobacteria (Figure 2c). Interestingly, the absolute abundances of Bacteroidetes and Firmicutes were lower in the KD-fed group in comparison to the WD-fed group, and the average ratio of Firmicutes/Bacteroidetes was not significantly different between the WD-fed (0.280 ± 0.084) and the KD-fed groups (0.220 ± 0.091).
4. Discussion
Diet plays a major role in causing 50% of gut microbial variations in mice and 20% in humans, making it a potential strategy for disease management via modulation of the gut microbiota [32,33,34]. Studies reporting the effects of KD on the gut microbiome of both humans and mice have shown that KD leads to a lower overall alpha diversity [8,35]. This effect leads to lower SCFAs production, which mainly results from the fermentation of dietary fibers and nondigestible carbohydrates by the gut microbiota [8,35]. Studies have demonstrated through high-throughput DNA sequencing technologies in large-scale 16S rRNA and shotgun metagenomics different changes in the composition of the colonic microbiome upon KD in both humans and mice [16,21,35,36,37,38,39]. These studies reported the influence of the KD on bacterial taxa, richness, and diversity. They mentioned the positive effect of the KD in reshaping the gut microbiota and its biological functions and its negative effects like decreased diversity and increased number of pro-inflammatory bacteria [11]. In addition, using the FISH technique, Swidsinski et al. showed that there was a decrease in the diversity and total concentration of bacteria in the gut of humans consuming KD [31], which was also observed in wild-type C57Bl/6J mice in our study. However, because most studies are carried out in the short term, they are disease-associated, which limits their generalizability to the overall population.
In our study, we also used the FISH technique, but we could not have a final conclusion on the whole biodiversity or composition of the microbiota, as FISH only detects bacteria at concentrations higher than 105 per ml. Nevertheless, Swidsinski et al. explained that even though the information provided by sequence analysis on the physical abundance and contribution of bacteria to bio-fermentation is substandard, their abundance within the fecal matter directly expresses their bio-fermenting power [40].
Our data quantifying bacterial participants demonstrated a clear difference in bacterial concentration and composition after KD compared with WD in C57BL/6J mice. For instance, the total number of bacteria in the fecal samples of mice consuming KD was reduced compared to those consuming WD, confirming previous data [41]. Feeding mice with the KD resulted in higher levels of bacteria belonging to the phylum Actinobacteria. Numerous reports have shown that members of Actinobacteria, such as Bifidobacterium spp., play a protective role against colitis and are able to reduce the overall level of systemic inflammation [42,43,44,45]. In addition, the significant decrease in blood glucose levels observed after 4 weeks of KD was consistent with the significant increase in Actinobacteria, and in particular Bifidobacterium spp., which have previously been reported to have lower blood glucose and act as an anti-diabetic agent [46]. This significant increase in the Bifidobacterium spp. after KD may indicate that KD can have a possible beneficial effect on the inflammatory status, as well as on glucose homeostasis.
On the other hand, in our study, KD in mice led to a decrease in the number of Proteobacteria. Members of the Proteobacteria phylotype have been shown to play a role in the onset and progression of ulcerative colitis and inflammatory diseases, such as Crohn’s disease [45,47,48,49,50]. Proteobacteria represent a minor part of the healthy gut microbiota, but their disproportionate increase leads to inflammation [51]; as such, Proteobacteria have been considered as a microbial signature of intestinal dysbiosis [52]. Similar to our results, Kong et al. showed that KD was able to reduce the abundance of Proteobacteria in mice with DSS-induced colitis [12]. This may be due to the low intake of carbohydrates and sugars in the KD, which can be fermented by Proteobacteria, leading to their increase [51]. However, in their systemic review, Kaviyarasan et al. found several studies in which KD was associated with an increase in the number of Proteobacteria [53].
Interestingly, we found no differences in the abundance of Verrucomicrobia between WD- and KD-fed mice. This is an important finding as this taxon contains members such as Akkermansia muciniphila, whose presence has been associated with gut health and reversal of experimental colitis [54,55]. Our results were not consistent with those of Ma et al., and Olson et al., who showed an increase in A. muciniphila after a KD in mice [8,21,36]. A. muciniphila has been reported to have anti-diabetic effects, leading to an improvement in insulin secretion and glucose homeostasis [56]. This was confirmed in the studies by Ma et al. and Olson et al., where a decrease in glucose levels was associated with an increase in A. muciniphila numbers after KD [8,21,36]. However, another study by Newell et al. found a significant decrease in A. muciniphila numbers in BTBR mice following a KD compared to a chow diet [41], possibly due to the very low carbohydrate content of the KD, which serves as a fuel source for microorganisms such as A. muciniphila [57]. This effect was only observed in BTBR mice and not wild-type mice, suggesting a disease-related mechanism [41]. Looking at the different results observed in C57BL/6 mice, it is noteworthy that A. muciniphila was significantly increased after a 16-week ketogenic diet [36], but not after 4 weeks (our study) or 14 days [41], suggesting that a longer duration of KD may be required to cause a significant increase in A. muciniphila.
Looking more closely at the individual bacteria detected by the FISH probes, we found a significant increase in the Atopobium cluster in the KD group compared to the WD group, which has been implicated in the pathogenesis of several diseases such as type 2 diabetes mellitus [58], and ulcerative colitis [59]. However, we also observed a significant decrease in the concentration of opportunistic pathogens associated with infections, such as Sphingomonas and Erythrobacter, in the KD group.
Furthermore, our results showed that Bacteroidetes were overrepresented and Firmicutes were underrepresented in comparison to previous studies based on 16S rRNA sequencing [37] or metagenome sequencing. This may be due to the low number of probes specifically designed for the detection of Firmicutes in the murine gut. Therefore, claims based on the Firmicutes/Bacteroides ratio should be carefully considered. Here, the 4-week KD in C57Bl/6J mice did not have a significant effect on the Firmicutes to Bacteroidetes ratio compared to the WD. As Firmicutes and Bacteroidetes represent more than 90% of the gut microbial community, the ratio of Firmicutes to Bacteroidetes is considered important for microbial balance and gut health [11]. This composition is relatively unaffected by acute perturbations but is affected by continuous exposure to various stress factors that can have an impact on host health [60].
Evidence from different studies has shown a clear effect of diet on the gut microbiome and a clear relationship between the state of the gut microbiome and chronic diseases such as type 2 diabetes mellitus (T2DM) and cardiovascular disease (CVD) [61]. For instance, the beneficial effects of the KD on the gut microbiota have been implicated in metabolic health [8]. Defeudis et al. stated that a very low carbohydrate ketogenic diet (VLCKD) might represent an effective strategy for treating T2DM and obesity as it results in ketosis due to low carbohydrate content, and this effect has been related to the impact of VLCKD on the gut microbiota [62]. In addition, ketosis has been associated with different biochemical and physiological mechanisms that exert a systemic anti-inflammatory effect, which in turn has a direct effect on cardiovascular diseases [63]. Furthermore, VLCKD is reported to improve the diversity of the microbiota by counteracting Proteobacteria leading to weight loss and favouring Firmicutes, Ruminococcaceae, and Mogibacteriaceae [62]. In our study, we were able to confirm a decrease in Proteobacteria in the KD-fed mice. However, we did not observe a corresponding increase in the other genera, which could be due to the short-term period of the diet followed in our study. Furthermore, the proven anti-inflammatory effects of KD reported in different studies make it a potential strategy for the prevention or treatment of CVD. Another anti-inflammatory factor in KD is the elimination of pro-inflammatory simple sugars, which has been reflected in CVD improvement because the restriction of carbohydrate content has shown anti-inflammatory benefits in the case of cardiometabolic health. As such, these studies recommend a high-fat, well-composed KD that is rich in omega-3 polyunsaturated fatty acids due to its anti-inflammatory and cardioprotective effects [63].
In conclusion, KD led to a lower total number of bacteria in the guts of mice. A limitation to consider is the duration of the dietary intervention, as longer durations have been shown to have a stronger effect on shaping the gut microbiota. Our study was also limited by the fact that the fecal microbiota analysis was performed using the FISH technique. This technique does not allow an in-depth study of all the major species involved in health and disease; therefore, we still lack data to make a definitive conclusion about the effect of KD at the species level. Another limitation of the study is that the effect of KD on the fecal microbiota was only investigated in female mice, and the estrous cycle was not determined, as recent studies have shown sex differences in response to KD [64,65]. Therefore, future studies are needed to evaluate possible sex-related effects on the gut microbiota associated with KD. However, the preliminary data we have obtained shows that the KD can alter the gut microbiota composition in female wild-type mice, which could potentially have anti-inflammatory and anti-diabetic effects. Accordingly, the gut microbiota may be considered a target for the prevention or treatment of diseases associated with a western diet and lifestyle. However, more preclinical and clinical studies should be conducted to further investigate the effect of the KD on different diseases and to reach a definitive conclusion, as the results are still controversial among different studies.
Author Contributions
Conceptualization: A.S., C.S., N.R. and K.-H.W.; Methodology: A.S. and C.S.; Formal analysis: N.R., R.E.H., S.O. and T.G. (Tobias Goris); Investigation: A.S., C.S. and N.R.; Data curation, T.G. (Tobias Goris) and S.O.; Writing—original draft preparation, R.E.H. and N.R.; Writing—review and editing, A.S., K.-H.W., N.R., R.E.H., S.O. and T.G. (Tobias Goris) and T.G. (Tilman Grune); Visualization, R.E.H. and N.R.; Supervision, K.-H.W.; Project administration, K.-H.W. and T.G. (Tilman Grune); Funding acquisition: K.-H.W. and T.G. (Tilman Grune). All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the German Research Foundation (DFG) under the grant (WE2908/13-1) awarded to K.-H.W. and T.G. (Tilman Grune) (GR1240/20-1). This study was also funded by the Ministry of Science, Research, and Culture of the State of Brandenburg and the Brandenburg Medical School Theodor Fontane (Medizinische Hochschule Brandenburg Theodor Fontane, MHB), publication fund supported by the German Research Foundation (Deutsche Forschungsgemeinschaft, DFG). The work of T.G. (Tobias Goris) was funded by the European Union’s Horizon 2020 Research and Innovation Programme under grant agreement number 814650 (SynBio4Flav).
Institutional Review Board Statement
The animal study protocol was approved by the State Animal Care Committee (Landesamt für Gesundheit und Soziales, Berlin, Germany; approval code G0047/15).
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Zhu, H.; Bi, D.; Zhang, Y.; Kong, C.; Du, J.; Wu, X.; Wei, Q.; Qin, H. Ketogenic diet for human diseases: The underlying mechanisms and potential for clinical implementations. Signal Transduct. Target. Ther. 2022, 7, 11. [Google Scholar] [CrossRef] [PubMed]
- Wijnen, B.F.M.; de Kinderen, R.J.A.; Lambrechts, D.; Postulart, D.; Aldenkamp, A.P.; Majoie, M.; Evers, S. Long-term clinical outcomes and economic evaluation of the ketogenic diet versus care as usual in children and adolescents with intractable epilepsy. Epilepsy Res. 2017, 132, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Gano, L.B.; Patel, M.; Rho, J.M. Ketogenic diets, mitochondria, and neurological diseases. J. Lipid Res. 2014, 55, 2211–2228. [Google Scholar] [CrossRef] [PubMed]
- Boison, D. New insights into the mechanisms of the ketogenic diet. Curr. Opin. Neurol. 2017, 30, 187–192. [Google Scholar] [CrossRef]
- Srivastava, S.; Pawar, V.A.; Tyagi, A.; Sharma, K.P.; Kumar, V.; Shukla, S.K. Immune Modulatory Effects of Ketogenic Diet in Different Disease Conditions. Immuno 2023, 3, 1–15. [Google Scholar] [CrossRef]
- Milder, J.; Patel, M. Modulation of oxidative stress and mitochondrial function by the ketogenic diet. Epilepsy Res. 2012, 100, 295–303. [Google Scholar] [CrossRef]
- Puchalska, P.; Crawford, P.A. Multi-dimensional Roles of Ketone Bodies in Fuel Metabolism, Signaling, and Therapeutics. Cell Metab. 2017, 25, 262–284. [Google Scholar] [CrossRef]
- Attaye, I.; van Oppenraaij, S.; Warmbrunn, M.V.; Nieuwdorp, M. The Role of the Gut Microbiota on the Beneficial Effects of Ketogenic Diets. Nutrients 2021, 14, 191. [Google Scholar] [CrossRef]
- Dashti, H.M.; Mathew, T.C.; Khadada, M.; Al-Mousawi, M.; Talib, H.; Asfar, S.K.; Behbahani, A.I.; Al-Zaid, N.S. Beneficial effects of ketogenic diet in obese diabetic subjects. Mol. Cell. Biochem. 2007, 302, 249–256. [Google Scholar] [CrossRef]
- Bolla, A.M.; Caretto, A.; Laurenzi, A.; Scavini, M.; Piemonti, L. Low-Carb and Ketogenic Diets in Type 1 and Type 2 Diabetes. Nutrients 2019, 11, 962. [Google Scholar] [CrossRef]
- Paoli, A.; Mancin, L.; Bianco, A.; Thomas, E.; Mota, J.F.; Piccini, F. Ketogenic Diet and Microbiota: Friends or Enemies? Genes 2019, 10, 534. [Google Scholar] [CrossRef] [PubMed]
- Kong, C.; Yan, X.; Liu, Y.; Huang, L.; Zhu, Y.; He, J.; Gao, R.; Kalady, M.F.; Goel, A.; Qin, H.; et al. Ketogenic diet alleviates colitis by reduction of colonic group 3 innate lymphoid cells through altering gut microbiome. Signal Transduct. Target. Ther. 2021, 6, 154. [Google Scholar] [CrossRef] [PubMed]
- Rawat, K.; Singh, N.; Kumari, P.; Saha, L. A review on preventive role of ketogenic diet (KD) in CNS disorders from the gut microbiota perspective. Rev. Neurosci. 2021, 32, 143–157. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.M.; Hazen, S.L. The gut microbial endocrine organ: Bacterially derived signals driving cardiometabolic diseases. Annu. Rev. Med. 2015, 66, 343–359. [Google Scholar] [CrossRef]
- Cani, P.D.; Van Hul, M.; Lefort, C.; Depommier, C.; Rastelli, M.; Everard, A. Microbial regulation of organismal energy homeostasis. Nat. Metab. 2019, 1, 34–46. [Google Scholar] [CrossRef]
- Ang, Q.Y.; Alexander, M.; Newman, J.C.; Tian, Y.; Cai, J.; Upadhyay, V.; Turnbaugh, J.A.; Verdin, E.; Hall, K.D.; Leibel, R.L.; et al. Ketogenic Diets Alter the Gut Microbiome Resulting in Decreased Intestinal Th17 Cells. Cell 2020, 181, 1263–1275.e1216. [Google Scholar] [CrossRef]
- Portincasa, P.; Bonfrate, L.; Vacca, M.; De Angelis, M.; Farella, I.; Lanza, E.; Khalil, M.; Wang, D.Q.; Sperandio, M.; Di Ciaula, A. Gut Microbiota and Short Chain Fatty Acids: Implications in Glucose Homeostasis. Int. J. Mol. Sci. 2022, 23, 1105. [Google Scholar] [CrossRef]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. What is the Healthy Gut Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and Diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef]
- Brestoff, J.R.; Artis, D. Commensal bacteria at the interface of host metabolism and the immune system. Nat. Immunol. 2013, 14, 676–684. [Google Scholar] [CrossRef]
- Mardinoglu, A.; Wu, H.; Bjornson, E.; Zhang, C.; Hakkarainen, A.; Räsänen, S.M.; Lee, S.; Mancina, R.M.; Bergentall, M.; Pietiläinen, K.H.; et al. An Integrated Understanding of the Rapid Metabolic Benefits of a Carbohydrate-Restricted Diet on Hepatic Steatosis in Humans. Cell Metab. 2018, 27, 559–571.e555. [Google Scholar] [CrossRef]
- Olson, C.A.; Vuong, H.E.; Yano, J.M.; Liang, Q.Y.; Nusbaum, D.J.; Hsiao, E.Y. The Gut Microbiota Mediates the Anti-Seizure Effects of the Ketogenic Diet. Cell 2018, 173, 1728–1741.e1713. [Google Scholar] [CrossRef]
- Dowis, K.; Banga, S. The Potential Health Benefits of the Ketogenic Diet: A Narrative Review. Nutrients 2021, 13, 1654. [Google Scholar] [CrossRef] [PubMed]
- Durazzi, F.; Sala, C.; Castellani, G.; Manfreda, G.; Remondini, D.; De Cesare, A. Comparison between 16S rRNA and shotgun sequencing data for the taxonomic characterization of the gut microbiota. Sci. Rep. 2021, 11, 3030. [Google Scholar] [CrossRef]
- Wang, X.; Howe, S.; Deng, F.; Zhao, J. Current Applications of Absolute Bacterial Quantification in Microbiome Studies and Decision-Making Regarding Different Biological Questions. Microorganisms 2021, 9, 1797. [Google Scholar] [CrossRef] [PubMed]
- Props, R.; Kerckhof, F.M.; Rubbens, P.; De Vrieze, J.; Hernandez Sanabria, E.; Waegeman, W.; Monsieurs, P.; Hammes, F.; Boon, N. Absolute quantification of microbial taxon abundances. ISME J. 2017, 11, 584–587. [Google Scholar] [CrossRef] [PubMed]
- Daims, H.; Ramsing, N.B.; Schleifer, K.H.; Wagner, M. Cultivation-independent, semiautomatic determination of absolute bacterial cell numbers in environmental samples by fluorescence in situ hybridization. Appl. Environ. Microbiol. 2001, 67, 5810–5818. [Google Scholar] [CrossRef] [PubMed]
- Dorffel, Y.; Swidsinski, A.; Loening-Baucke, V.; Wiedenmann, B.; Pavel, M. Common biostructure of the colonic microbiota in neuroendocrine tumors and Crohn’s disease and the effect of therapy. Inflamm. Bowel Dis. 2012, 18, 1663–1671. [Google Scholar] [CrossRef]
- Swidsinski, A.; Loening-Baucke, V.; Kirsch, S.; Doerffel, Y. Functional biostructure of colonic microbiota (central fermenting area, germinal stock area and separating mucus layer) in healthy subjects and patients with diarrhea treated with Saccharomyces boulardii. Gastroenterol. Clin. Biol. 2010, 34 (Suppl. S1), S79–S92. [Google Scholar] [CrossRef]
- Swidsinski, A. Standards for bacterial identification by fluorescence In situ hybridization within eukaryotic tissue using ribosomal rRNA-based probes. Inflamm. Bowel Dis. 2006, 12, 824–826. [Google Scholar] [CrossRef]
- Greuter, D.; Loy, A.; Horn, M.; Rattei, T. probeBase—An online resource for rRNA-targeted oligonucleotide probes and primers: New features 2016. Nucleic Acids Res. 2016, 44, D586–D589. [Google Scholar] [CrossRef]
- Swidsinski, A.; Dorffel, Y.; Loening-Baucke, V.; Gille, C.; Goktas, O.; Reisshauer, A.; Neuhaus, J.; Weylandt, K.H.; Guschin, A.; Bock, M. Reduced Mass and Diversity of the Colonic Microbiome in Patients with Multiple Sclerosis and Their Improvement with Ketogenic Diet. Front. Microbiol. 2017, 8, 1141. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Zhang, M.; Wang, S.; Han, R.; Cao, Y.; Hua, W.; Mao, Y.; Zhang, X.; Pang, X.; Wei, C.; et al. Interactions between gut microbiota, host genetics and diet relevant to development of metabolic syndromes in mice. ISME J. 2010, 4, 232–241. [Google Scholar] [CrossRef]
- David, L.A.; Materna, A.C.; Friedman, J.; Campos-Baptista, M.I.; Blackburn, M.C.; Perrotta, A.; Erdman, S.E.; Alm, E.J. Host lifestyle affects human microbiota on daily timescales. Genome Biol. 2014, 15, R89. [Google Scholar] [CrossRef] [PubMed]
- Leeming, E.R.; Johnson, A.J.; Spector, T.D.; Le Roy, C.I. Effect of Diet on the Gut Microbiota: Rethinking Intervention Duration. Nutrients 2019, 11, 2862. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhou, S.; Zhou, Y.; Yu, L.; Zhang, L.; Wang, Y. Altered gut microbiome composition in children with refractory epilepsy after ketogenic diet. Epilepsy Res. 2018, 145, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.; Wang, A.C.; Parikh, I.; Green, S.J.; Hoffman, J.D.; Chlipala, G.; Murphy, M.P.; Sokola, B.S.; Bauer, B.; Hartz, A.M.S.; et al. Ketogenic diet enhances neurovascular function with altered gut microbiome in young healthy mice. Sci. Rep. 2018, 8, 6670. [Google Scholar] [CrossRef]
- Murtaza, N.; Burke, L.M.; Vlahovich, N.; Charlesson, B.; O’ Neill, H.; Ross, M.L.; Campbell, K.L.; Krause, L.; Morrison, M. The Effects of Dietary Pattern during Intensified Training on Stool Microbiota of Elite Race Walkers. Nutrients 2019, 11, 261. [Google Scholar] [CrossRef]
- Lindefeldt, M.; Eng, A.; Darban, H.; Bjerkner, A.; Zetterström, C.K.; Allander, T.; Andersson, B.; Borenstein, E.; Dahlin, M.; Prast-Nielsen, S. The ketogenic diet influences taxonomic and functional composition of the gut microbiota in children with severe epilepsy. NPJ Biofilms Microbiomes 2019, 5, 5. [Google Scholar] [CrossRef]
- Xie, G.; Zhou, Q.; Qiu, C.Z.; Dai, W.K.; Wang, H.P.; Li, Y.H.; Liao, J.X.; Lu, X.G.; Lin, S.F.; Ye, J.H.; et al. Ketogenic diet poses a significant effect on imbalanced gut microbiota in infants with refractory epilepsy. World J. Gastroenterol. 2017, 23, 6164–6171. [Google Scholar] [CrossRef]
- Swidsinski, A.; Dorffel, Y.; Loening-Baucke, V.; Gille, C.; Reisshauer, A.; Goktas, O.; Kruger, M.; Neuhaus, J.; Schrodl, W. Impact of humic acids on the colonic microbiome in healthy volunteers. World J. Gastroenterol. 2017, 23, 885–890. [Google Scholar] [CrossRef]
- Newell, C.; Bomhof, M.R.; Reimer, R.A.; Hittel, D.S.; Rho, J.M.; Shearer, J. Ketogenic diet modifies the gut microbiota in a murine model of autism spectrum disorder. Mol. Autism 2016, 7, 37. [Google Scholar] [CrossRef]
- Duranti, S.; Gaiani, F.; Mancabelli, L.; Milani, C.; Grandi, A.; Bolchi, A.; Santoni, A.; Lugli, G.A.; Ferrario, C.; Mangifesta, M.; et al. Elucidating the gut microbiome of ulcerative colitis: Bifidobacteria as novel microbial biomarkers. FEMS Microbiol. Ecol. 2016, 92, fiw191. [Google Scholar] [CrossRef]
- Hidalgo-Cantabrana, C.; Algieri, F.; Rodriguez-Nogales, A.; Vezza, T.; Martinez-Camblor, P.; Margolles, A.; Ruas-Madiedo, P.; Galvez, J. Effect of a Ropy Exopolysaccharide-Producing Bifidobacterium animalis subsp. lactis Strain Orally Administered on DSS-Induced Colitis Mice Model. Front. Microbiol. 2016, 7, 868. [Google Scholar] [CrossRef]
- Srutkova, D.; Schwarzer, M.; Hudcovic, T.; Zakostelska, Z.; Drab, V.; Spanova, A.; Rittich, B.; Kozakova, H.; Schabussova, I. Bifidobacterium longum CCM 7952 Promotes Epithelial Barrier Function and Prevents Acute DSS-Induced Colitis in Strictly Strain-Specific Manner. PLoS ONE 2015, 10, e0134050. [Google Scholar] [CrossRef]
- Zhang, D.; Wei, C.; Yao, J.; Cai, X.; Wang, L. Interleukin-10 gene-carrying bifidobacteria ameliorate murine ulcerative colitis by regulating regulatory T cell/T helper 17 cell pathway. Exp. Biol. Med. 2015, 240, 1622–1629. [Google Scholar] [CrossRef]
- Le, T.K.; Hosaka, T.; Nguyen, T.T.; Kassu, A.; Dang, T.O.; Tran, H.B.; Pham, T.P.; Tran, Q.B.; Le, T.H.; Pham, X.D. Bifidobacterium species lower serum glucose, increase expressions of insulin signaling proteins, and improve adipokine profile in diabetic mice. Biomed. Res. 2015, 36, 63–70. [Google Scholar] [CrossRef]
- Atherly, T.; Mosher, C.; Wang, C.; Hostetter, J.; Proctor, A.; Brand, M.W.; Phillips, G.J.; Wannemuehler, M.; Jergens, A.E. Helicobacter bilis Infection Alters Mucosal Bacteria and Modulates Colitis Development in Defined Microbiota Mice. Inflamm. Bowel Dis. 2016, 22, 2571–2581. [Google Scholar] [CrossRef]
- Mirsepasi-Lauridsen, H.C.; Halkjaer, S.I.; Mortensen, E.M.; Lydolph, M.C.; Nordgaard-Lassen, I.; Krogfelt, K.A.; Petersen, A.M. Extraintestinal pathogenic Escherichia coli are associated with intestinal inflammation in patients with ulcerative colitis. Sci. Rep. 2016, 6, 31152. [Google Scholar] [CrossRef]
- Carvalho, F.A.; Koren, O.; Goodrich, J.K.; Johansson, M.E.; Nalbantoglu, I.; Aitken, J.D.; Su, Y.; Chassaing, B.; Walters, W.A.; González, A.; et al. Transient inability to manage proteobacteria promotes chronic gut inflammation in TLR5-deficient mice. Cell Host Microbe 2012, 12, 139–152. [Google Scholar] [CrossRef]
- Vester-Andersen, M.K.; Mirsepasi-Lauridsen, H.C.; Prosberg, M.V.; Mortensen, C.O.; Träger, C.; Skovsen, K.; Thorkilgaard, T.; Nøjgaard, C.; Vind, I.; Krogfelt, K.A.; et al. Increased abundance of proteobacteria in aggressive Crohn’s disease seven years after diagnosis. Sci. Rep. 2019, 9, 13473. [Google Scholar] [CrossRef]
- Satokari, R. High Intake of Sugar and the Balance between Pro- and Anti-Inflammatory Gut Bacteria. Nutrients 2020, 12, 1348. [Google Scholar] [CrossRef] [PubMed]
- Rizzatti, G.; Lopetuso, L.R.; Gibiino, G.; Binda, C.; Gasbarrini, A. Proteobacteria: A Common Factor in Human Diseases. Biomed. Res. Int. 2017, 2017, 9351507. [Google Scholar] [CrossRef] [PubMed]
- Kaviyarasan, S.; Chung Sia, E.L.; Retinasamy, T.; Arulsamy, A.; Shaikh, M.F. Regulation of gut microbiome by ketogenic diet in neurodegenerative diseases: A molecular crosstalk. Front. Aging Neurosci. 2022, 14, 1015837. [Google Scholar] [CrossRef] [PubMed]
- Kang, C.S.; Ban, M.; Choi, E.J.; Moon, H.G.; Jeon, J.S.; Kim, D.K.; Park, S.K.; Jeon, S.G.; Roh, T.Y.; Myung, S.J.; et al. Extracellular vesicles derived from gut microbiota, especially Akkermansia muciniphila, protect the progression of dextran sulfate sodium-induced colitis. PLoS ONE 2013, 8, e76520. [Google Scholar] [CrossRef]
- Zhang, Z.; Wu, X.; Cao, S.; Wang, L.; Wang, D.; Yang, H.; Feng, Y.; Wang, S.; Li, L. Caffeic acid ameliorates colitis in association with increased Akkermansia population in the gut microbiota of mice. Oncotarget 2016, 7, 31790–31799. [Google Scholar] [CrossRef]
- Yan, X.; Liu, X.Y.; Zhang, D.; Zhang, Y.D.; Li, Z.H.; Liu, X.; Wu, F.; Chen, G.Q. Construction of a sustainable 3-hydroxybutyrate-producing probiotic Escherichia coli for treatment of colitis. Cell. Mol. Immunol. 2021, 18, 2344–2357. [Google Scholar] [CrossRef]
- Flint, H.J.; Scott, K.P.; Duncan, S.H.; Louis, P.; Forano, E. Microbial degradation of complex carbohydrates in the gut. Gut Microbes 2012, 3, 289–306. [Google Scholar] [CrossRef]
- Das, T.; Jayasudha, R.; Chakravarthy, S.; Prashanthi, G.S.; Bhargava, A.; Tyagi, M.; Rani, P.K.; Pappuru, R.R.; Sharma, S.; Shivaji, S. Alterations in the gut bacterial microbiome in people with type 2 diabetes mellitus and diabetic retinopathy. Sci. Rep. 2021, 11, 2738. [Google Scholar] [CrossRef]
- Swidsinski, A.; Loening-Baucke, V.; Vaneechoutte, M.; Doerffel, Y. Active Crohn’s disease and ulcerative colitis can be specifically diagnosed and monitored based on the biostructure of the fecal flora. Inflamm. Bowel Dis. 2008, 14, 147–161. [Google Scholar] [CrossRef]
- Magne, F.; Gotteland, M.; Gauthier, L.; Zazueta, A.; Pesoa, S.; Navarrete, P.; Balamurugan, R. The Firmicutes/Bacteroidetes Ratio: A Relevant Marker of Gut Dysbiosis in Obese Patients? Nutrients 2020, 12, 1474. [Google Scholar] [CrossRef]
- Sikalidis, A.K.; Maykish, A. The Gut Microbiome and Type 2 Diabetes Mellitus: Discussing a Complex Relationship. Biomedicines 2020, 8, 8. [Google Scholar] [CrossRef] [PubMed]
- Defeudis, G.; Rossini, M.; Khazrai, Y.M.; Pipicelli, A.M.V.; Brucoli, G.; Veneziano, M.; Strollo, F.; Bellia, A.; Bitterman, O.; Lauro, D.; et al. The gut microbiome as possible mediator of the beneficial effects of very low calorie ketogenic diet on type 2 diabetes and obesity: A narrative review. Eat. Weight Disord. 2022, 27, 2339–2346. [Google Scholar] [CrossRef] [PubMed]
- Dyńka, D.; Kowalcze, K.; Charuta, A.; Paziewska, A. The Ketogenic Diet and Cardiovascular Diseases. Nutrients 2023, 15, 3368. [Google Scholar] [CrossRef] [PubMed]
- Cochran, J.; Taufalele, P.V.; Lin, K.D.; Zhang, Y.; Dale Abel, E. Sex Differences in the Response of C57BL/6 Mice to Ketogenic Diets. Diabetes 2018, 67, 1884-P. [Google Scholar] [CrossRef]
- Pontifex, M.G.; Vauzour, D.; Muller, M. Sexual dimorphism in the context of nutrition and health. In Proceedings of the Nutrition Society; Cambridge University Press: Cambridge, UK, 2023; pp. 1–11. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).