
Effect of Amino Acid Supplementation on Iron Regulation after Endurance Exercise


Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
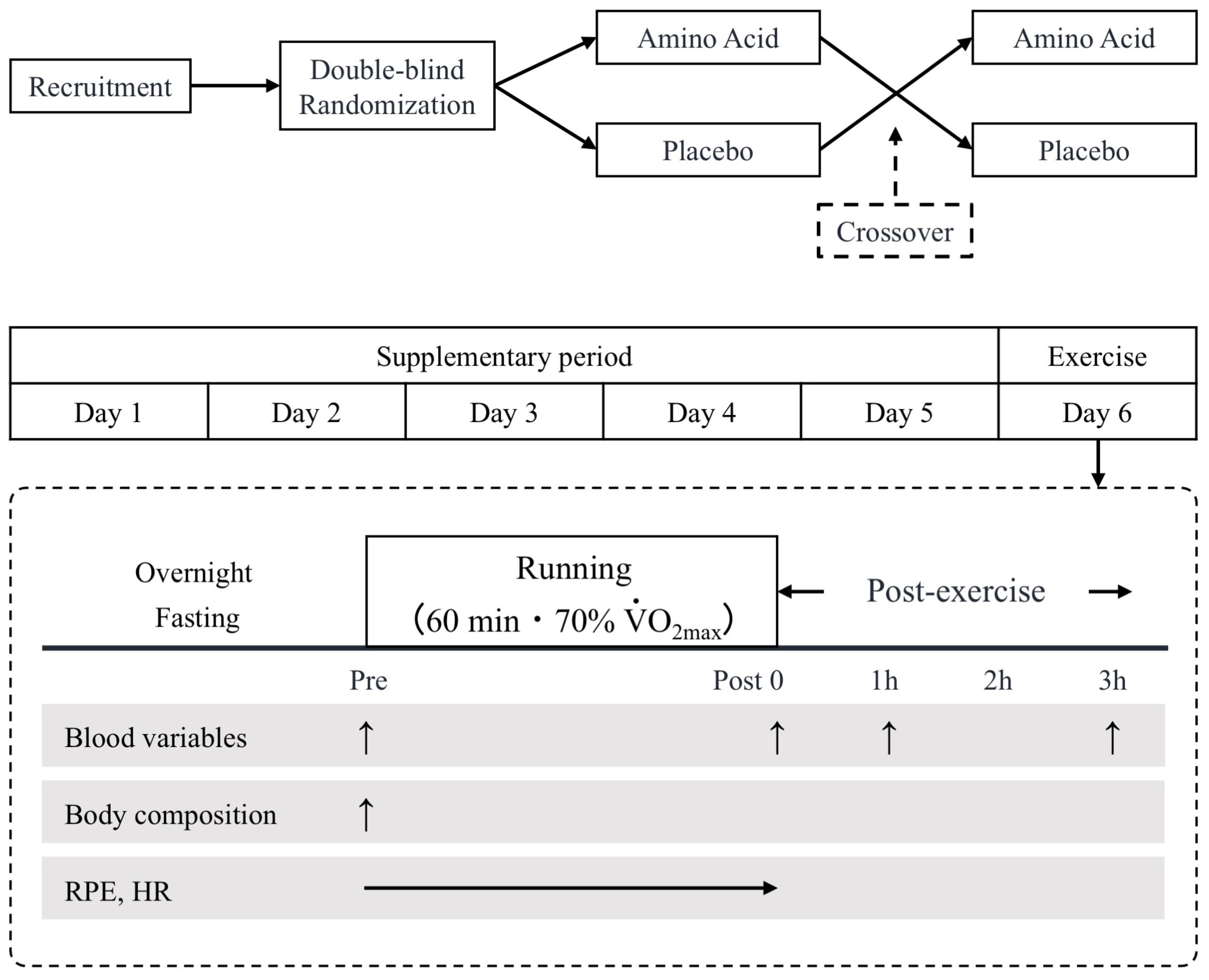
2.2. Experimental Design
2.3. Amino Acid Supplementation
2.4. Experimental Protocol
2.4.1. Maximal Oxygen Uptake (O2max)
2.4.2. Supplement and Dietary Prescriptions
2.4.3. Blood Sample Collection and Analyses
2.5. Statistical Analysis
3. Results
3.1. Body Composition and Running Speed during Exercise
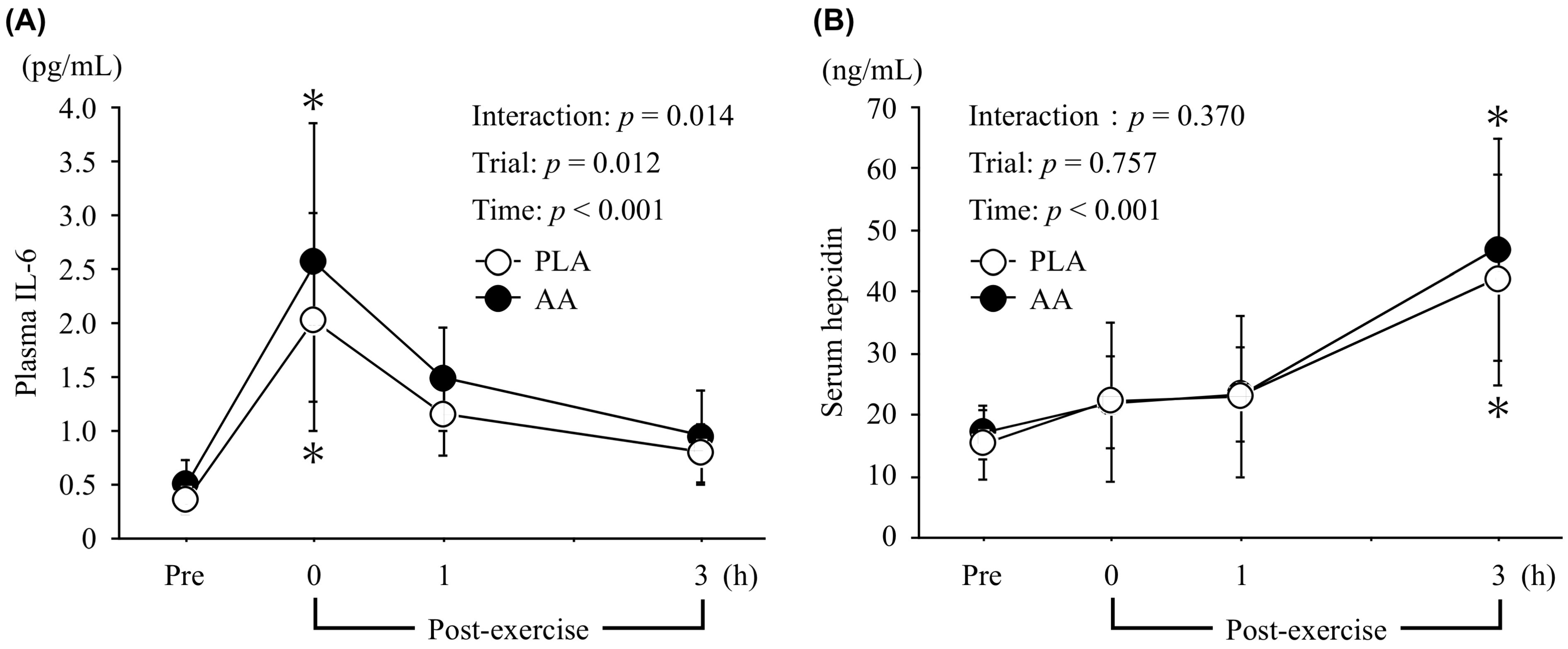
3.2. IL-6 and Hepcidin
3.3. Serum Ferritin, Iron, Blood Glucose, and Lactate Levels
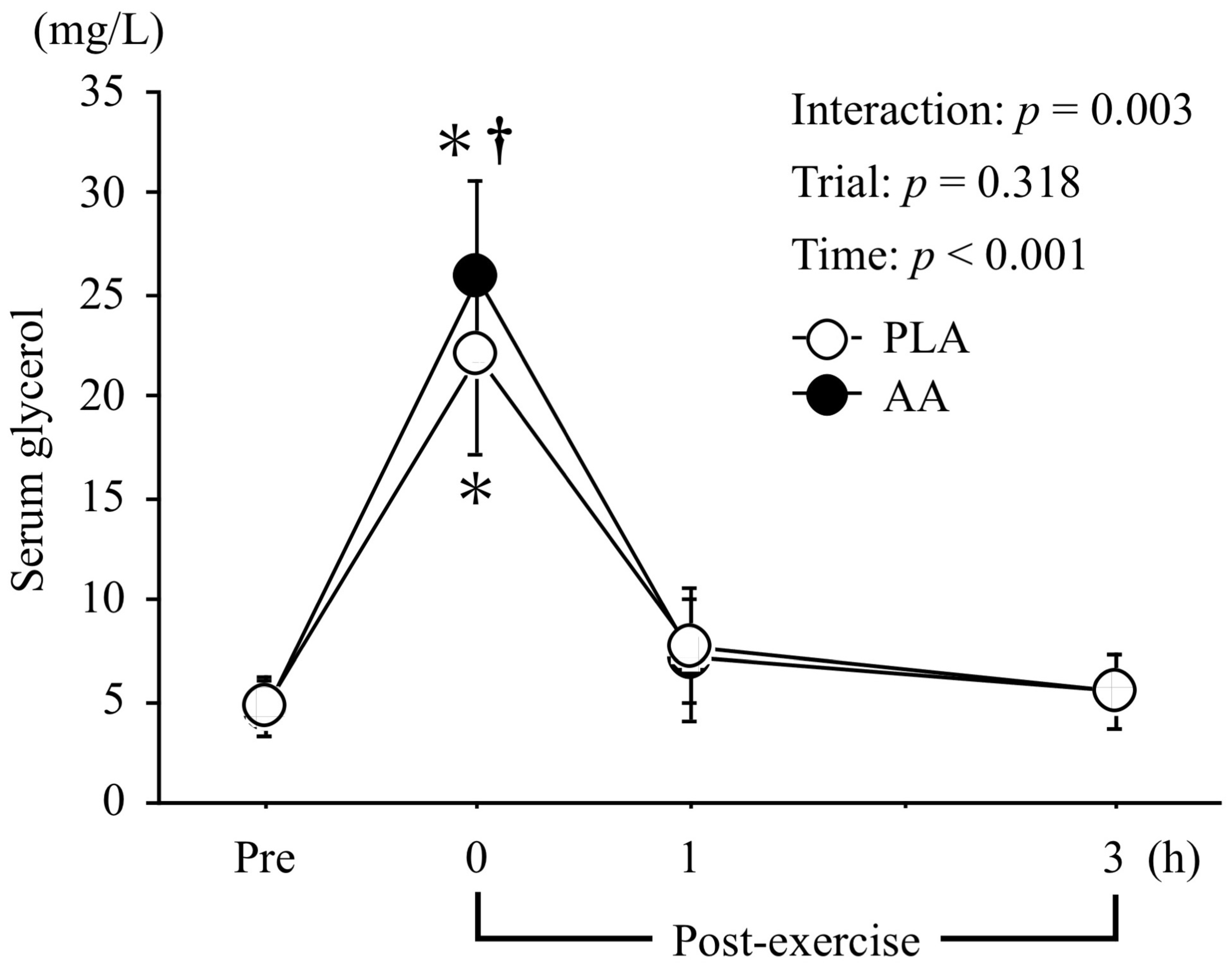
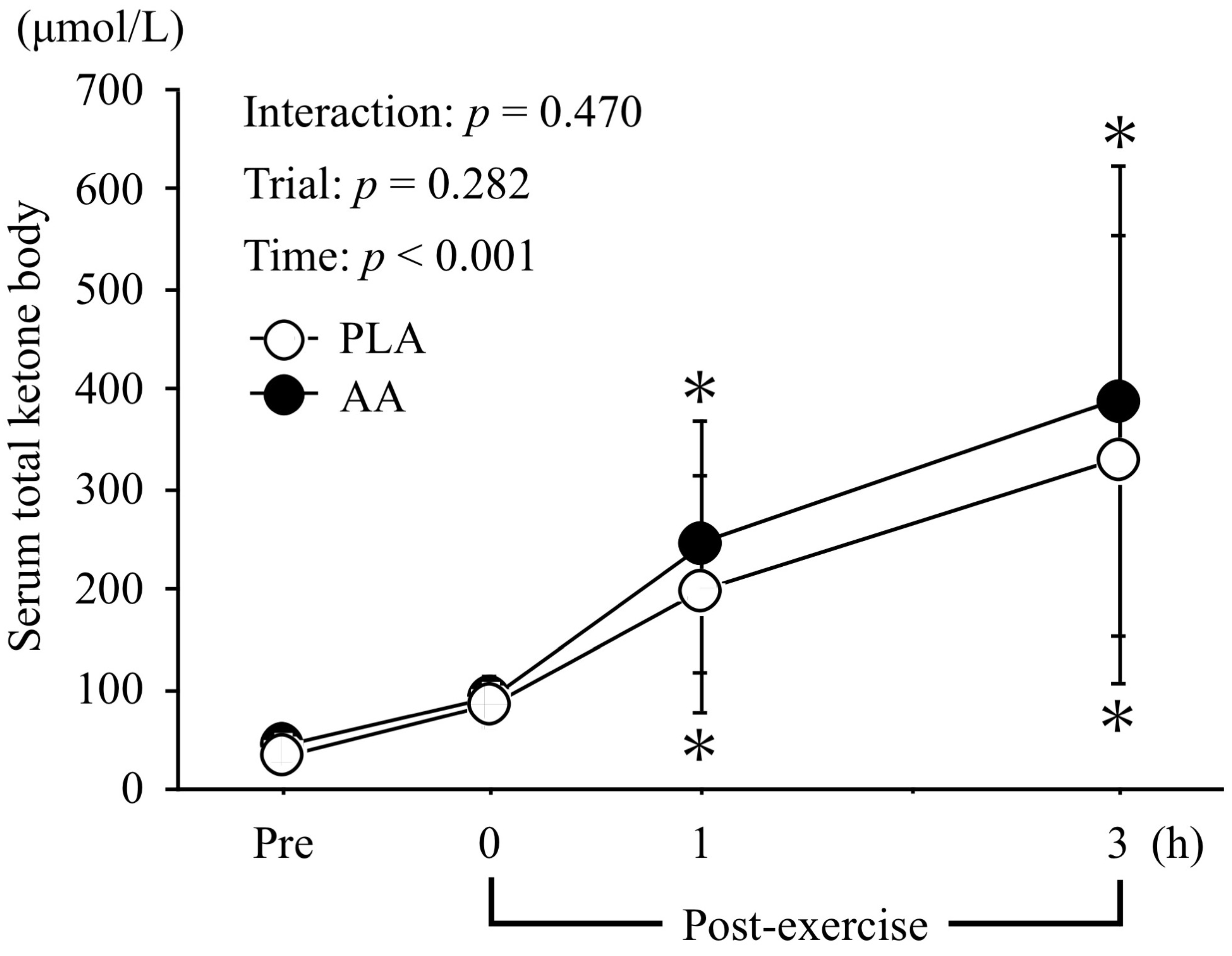
3.4. Serum Glycerol and Total Ketone Body Levels
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Latunde-Dada, G.O. Iron metabolism in athletes—Achieving a gold standard. Eur. J. Haematol. 2013, 90, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Dellavalle, D.M.; Haas, J.D. Iron status is associated with endurance performance and training in female rowers. Med. Sci. Sports Exerc. 2012, 44, 1552–1559. [Google Scholar] [CrossRef] [PubMed]
- Rineau, E.; Gueguen, N.; Procaccio, V.; Geneviève, F.; Reynier, P.; Henrion, D.; Lasocki, S. Iron deficiency without anemia decreases physical endurance and mitochondrial complex I activity of oxidative skeletal muscle in the mouse. Nutrients 2021, 13, 1056. [Google Scholar] [CrossRef] [PubMed]
- Coates, A.; Mountjoy, M.; Burr, J. Incidence of iron deficiency and iron deficient anemia in elite runners and triathletes. Clin. J. Sport Med. 2017, 27, 493–498. [Google Scholar] [CrossRef] [PubMed]
- Ishibashi, A.; Maeda, N.; Sumi, D.; Goto, K. Elevated serum hepcidin levels during an intensified training period in well-trained female long-distance runners. Nutrients 2017, 9, 277. [Google Scholar] [CrossRef] [PubMed]
- Peeling, P.; Dawson, B.; Goodman, C.; Landers, G.; Trinder, D. Athletic induced iron deficiency: New insights into the role of inflammation, cytokines and hormones. Eur. J. Appl. Physiol. 2008, 103, 381–391. [Google Scholar] [CrossRef] [PubMed]
- Peeling, P.; McKay, A.K.A.; Pyne, D.B.; Guelfi, K.J.; McCormick, R.H.; Laarakkers, C.M.; Swinkels, D.W.; Garvican-Lewis, L.A.; Ross, M.L.R.; Sharma, A.P.; et al. Factors influencing the post-exercise hepcidin-25 response in elite athletes. Eur. J. Appl. Physiol. 2017, 117, 1233–1239. [Google Scholar] [CrossRef]
- Ganz, T. Hepcidin, a key regulator of iron metabolism and mediator of anemia of inflammation. Blood 2003, 102, 783–788. [Google Scholar] [CrossRef]
- Nemeth, E.; Tuttle, M.S.; Powelson, J.; Vaughn, M.B.; Donovan, A.; Ward, D.M.; Ganz, T.; Kaplan, J. Hepcidin regulates cellular iron efflux by binding to ferroportin and inducing its internalization. Science 2004, 306, 2090–2093. [Google Scholar] [CrossRef]
- Nemeth, E.; Ganz, T. Hepcidin-Ferroportin Interaction Controls Systemic Iron Homeostasis. Int. J. Mol. Sci. 2021, 22, 6493. [Google Scholar] [CrossRef]
- Newlin, M.K.; Williams, S.; McNamara, T.; Tjalsma, H.; Swinkels, D.W.; Haymes, E.M. The effects of acute exercise bouts on hepcidin in women. Int. J. Sport Nutr. Exerc. Metab. 2012, 22, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Sim, M.; Dawson, B.; Landers, G.; Swinkels, D.W.; Tjalsma, H.; Trinder, D.; Peeling, P. Effect of exercise modality and intensity on post-exercise interleukin-6 and hepcidin levels. Int. J. Sport Nutr. Exerc. Metab. 2013, 23, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Peeling, P.; Dawson, B.; Goodman, C.; Landers, G.; Wiegerinck, E.T.; Swinkels, D.W.; Trinder, D. Effects of exercise on hepcidin response and iron metabolism during recovery. Int. J. Sport Nutr. Exerc. Metab. 2009, 19, 583–597. [Google Scholar] [CrossRef] [PubMed]
- Steensberg, A.; Febbraio, M.A.; Osada, T.; Schjerling, P.; van Hall, G.; Saltin, B.; Pedersen, B.K. Interleukin-6 production in contracting human skeletal muscle is influenced by pre-exercise muscle glycogen content. J. Physiol. 2001, 537, 633–639. [Google Scholar] [CrossRef] [PubMed]
- Hennigar, S.R.; McClung, J.P.; Pasiakos, S.M. Nutritional interventions and the IL-6 response to exercise. FASEB J. 2017, 31, 3719–3728. [Google Scholar] [CrossRef] [PubMed]
- Robson-Ansley, P.; Walshe, I.; Ward, D. The effect of carbohydrate ingestion on plasma interleukin-6, hepcidin and iron concentrations following prolonged exercise. Cytokine 2011, 53, 196–200. [Google Scholar] [CrossRef] [PubMed]
- Plotkin, D.L.; Delcastillo, K.; Van Every, D.W.; Tipton, K.D.; Aragon, A.A.; Schoenfeld, B.J. Isolated leucine and branched-chain amino acid supplementation for enhancing muscular strength and hypertrophy: A narrative review. Int. J. Sport Nutr. Exerc. Metab. 2021, 31, 292–301. [Google Scholar] [CrossRef]
- Fouré, A.; Bendahan, D. Is branched-chain amino acids supplementation an efficient nutritional strategy to alleviate skeletal muscle damage? A systematic review. Nutrients 2017, 9, 1047. [Google Scholar] [CrossRef]
- Doma, K.; Singh, U.; Boullosa, D.; Connor, J.D. The effect of branched-chain amino acid on muscle damage markers and performance following strenuous exercise: A systematic review and meta-analysis. Appl. Physiol. Nutr. Metab. 2021, 46, 1303–1313. [Google Scholar] [CrossRef]
- Kato, H.; Miura, K.; Nakano, S.; Suzuki, K.; Bannai, M.; Inoue, Y. Leucine-enriched essential amino acids attenuate inflammation in rat muscle and enhance muscle repair after eccentric contraction. Amino Acids 2016, 48, 2145–2155. [Google Scholar] [CrossRef]
- An, Y.H.; Kim, J.; Kim, H.J.; Lim, K. Effects of leucine-enriched essential amino acid supplementation on muscular fatigue and inflammatory cytokines in wheelchair basketball players. Phys. Act. Nutr. 2020, 24, 38–46. [Google Scholar] [CrossRef]
- Tataka, Y.; Haramura, M.; Hamada, Y.; Ono, M.; Toyoda, S.; Yamada, T.; Hiratsu, A.; Suzuki, K.; Miyashita, M. Effects of oral cystine and glutamine on exercise-induced changes in gastrointestinal permeability and damage markers in young men. Eur. J. Nutr. 2022, 61, 2331–2339. [Google Scholar] [CrossRef]
- Nilsonne, G.; Lekander, M.; Åkerstedt, T.; Axelsson, J.; Ingre, M. Diurnal Variation of Circulating Interleukin-6 in Humans: A Meta-Analysis. PLoS ONE 2016, 11, e0165799. [Google Scholar] [CrossRef]
- McCormick, R.; Moretti, D.; McKay, A.K.A.; Laarakkers, C.M.; Vanswelm, R.; Trinder, D.; Cox, G.R.; Zimmerman, M.B.; Sim, M.; Goodman, C.; et al. The Impact of Morning versus Afternoon Exercise on Iron Absorption in Athletes. Med. Sci. Sports Exerc. 2019, 51, 2147–2155. [Google Scholar] [CrossRef]
- Sumi, D.; Kojima, C.; Goto, K. Impact of Endurance Exercise in Hypoxia on Muscle Damage, Inflammatory and Performance Responses. J. Strength Cond. Res. 2018, 32, 1053–1062. [Google Scholar] [CrossRef]
- Badenhorst, C.E.; Black, K.E.; O’Brien, W.J. Hepcidin as a Prospective Individualized Biomarker for Individuals at Risk of Low Energy Availability. Int. J. Sport Nutr. Exerc. Metab. 2019, 29, 671–681. [Google Scholar] [CrossRef]
- Ishibashi, A.; Kojima, C.; Tanabe, Y.; Iwayama, K.; Hiroyama, T.; Tsuji, T.; Kamei, A.; Goto, K.; Takahashi, H. Effect of low energy availability during three consecutive days of endurance training on iron metabolism in male long distance runners. Physiol. Rep. 2020, 8, e14494. [Google Scholar] [CrossRef]
- Larsuphrom, P.; Latunde-Dada, G.O. Association of serum hepcidin levels with aerobic and resistance exercise: A systematic review. Nutrients 2021, 13, 393. [Google Scholar] [CrossRef]
- Ostrowski, K.; Schjerling, P.; Pedersen, B.K. Physical activity and plasma interleukin-6 in humans--effect of intensity of exercise. Eur. J. Appl. Physiol. 2000, 83, 512–515. [Google Scholar] [CrossRef]
- Jones, S.A.; Jenkins, B.J. Recent insights into targeting the IL-6 cytokine family in inflammatory diseases and cancer. Nat. Rev. Immunol. 2018, 18, 773–789. [Google Scholar] [CrossRef]
- Wallberg, L.; Mattsson, C.M.; Enqvist, J.K.; Ekblom, B. Plasma IL-6 concentration during ultra-endurance exercise. Eur. J. Appl. Physiol. 2011, 111, 1081–1088. [Google Scholar] [CrossRef] [PubMed]
- Kell, D.B.; Pretorius, E. Serum ferritin is an important inflammatory disease marker, as it is mainly a leakage product from damaged cells. Metallomics 2014, 6, 748–773. [Google Scholar] [CrossRef]
- Wang, W.; Knovich, M.A.; Coffman, L.G.; Torti, F.M.; Torti, S.V. Serum ferritin: Past, present and future. Biochimi. Biophys. Acta 2010, 1800, 760–769. [Google Scholar] [CrossRef]
- Kistner, T.M.; Pedersen, B.K.; Lieberman, D.E. Interleukin 6 as an energy allocator in muscle tissue. Nat. Metab. 2022, 4, 170–179. [Google Scholar] [CrossRef]
- van Hall, G.; Steensberg, A.; Sacchetti, M.; Fischer, C.; Keller, C.; Schjerling, P.; Hiscock, N.; Møller, K.; Saltin, B.; Febbraio, M.A.; et al. Interleukin-6 stimulates lipolysis and fat oxidation in humans. J. Clin. Endocrinol. Metab. 2003, 88, 3005–3010. [Google Scholar] [CrossRef]
- Górecka, M.; Krzemiński, K.; Mikulski, T.; Ziemba, A.W. ANGPTL4, IL-6 and TNF-α as regulators of lipid metabolism during a marathon run. Sci. Rep. 2022, 12, 19940. [Google Scholar] [CrossRef]
- Ma, S.; Ono, M.; Mizugaki, A.; Kato, H.; Miyashita, M.; Suzuki, K. Cystine/glutamine mixture supplementation attenuated fatigue during endurance exercise in healthy young men by enhancing fatty acid utilization. Sports 2022, 10, 147. [Google Scholar] [CrossRef]
- Cullen, T.; Thomas, G.; Wadley, A.J. Sleep Deprivation: Cytokine and Neuroendocrine Effects on Perception of Effort. Med. Sci. Sports Exerc. 2020, 52, 909–918. [Google Scholar] [CrossRef]
- Goto, K.; Mamiya, A.; Ito, H.; Maruyama, T.; Hayashi, N.; Badenhorst, C.E. Partial sleep deprivation after an acute exercise session does not augment hepcidin levels the following day. Physiol. Rep. 2020, 8, e14450. [Google Scholar] [CrossRef]
- Badenhorst, C.E.; Dawson, B.; Cox, G.R.; Laarakkers, C.M.; Swinkels, D.W.; Peeling, P. Acute dietary carbohydrate manipulation and the subsequent inflammatory and hepcidin responses to exercise. Eur. J. Appl. Physiol. 2015, 115, 2521–2530. [Google Scholar] [CrossRef]
- Sim, M.; Garvican-Lewis, L.A.; Cox, G.R.; Govus, A.; McKay, A.K.A.; Stellingwerff, T.; Peeling, P. Iron considerations for the athlete: A narrative review. Eur. J. Appl. Physiol. 2019, 119, 1463–1478. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | Amino Acid (AA) | Placebo (PLA) | ||
|---|---|---|---|---|
| Compound Ratio (%) | Amount Per Serving (g) | Compound Ratio (%) | Amount Per Serving (g) | |
| L-Leucine | 23.37 | 1.03 | 0.00 | 0.00 |
| L-Isoleucine | 6.22 | 0.27 | 0.00 | 0.00 |
| L-Valine | 6.43 | 0.28 | 0.00 | 0.00 |
| L-Glutamine | 22.73 | 1.00 | 0.00 | 0.00 |
| L-Cystine | 5.23 | 0.23 | 0.00 | 0.00 |
| Other essential amino acid | 22.41 | 0.99 | 0.00 | 0.00 |
| Other ingredients (e.g., carbohydrate, water) | 13.61 | 0.60 | 100.00 | 4.00 |
| Trial | Pre | Post-Exercise | |||
|---|---|---|---|---|---|
| Post 0 | Post 1 | Post 3 (h) | |||
| Ferritin (ng/mL) | AA | 101.1 ± 59.5 | 107.1 ± 61.1 | 105.7 ± 60.2 | 105.9 ± 60.5 |
| PLA | 92.2 ± 56.9 | 96.9 ± 59.4 | 95.7 ± 60.5 | 97.1 ± 57.8 | |
| Iron (μg/dL) | AA | 136 ± 30 | 157 ± 30 | 159 ± 28 | 152 ± 25 |
| PLA | 139 ± 33 | 158 ± 36 | 150 ± 30 | 148 ± 30 | |
| Glucose (mg/dL) | AA | 84 ± 4 | 86 ± 6 | 82 ± 6 | 81 ± 3 |
| PLA | 84 ± 8 | 85 ± 10 | 84 ± 8 | 84 ± 6 | |
| Lactate (mmol/L) | AA | 1.9 ± 0.4 | 2.1 ± 1.1 | 1.7 ± 0.4 | 1.7 ± 0.4 |
| PLA | 1.9 ± 0.5 | 2.1 ± 0.9 | 1.8 ± 0.3 | 1.7 ± 0.2 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, C.-A.; Hayashi, N.; Badenhorst, C.E.; Goto, K. Effect of Amino Acid Supplementation on Iron Regulation after Endurance Exercise. Nutrients 2023, 15, 4924. https://doi.org/10.3390/nu15234924
Lin C-A, Hayashi N, Badenhorst CE, Goto K. Effect of Amino Acid Supplementation on Iron Regulation after Endurance Exercise. Nutrients. 2023; 15(23):4924. https://doi.org/10.3390/nu15234924
Chicago/Turabian StyleLin, Chao-An, Nanako Hayashi, Claire E. Badenhorst, and Kazushige Goto. 2023. "Effect of Amino Acid Supplementation on Iron Regulation after Endurance Exercise" Nutrients 15, no. 23: 4924. https://doi.org/10.3390/nu15234924
APA StyleLin, C.-A., Hayashi, N., Badenhorst, C. E., & Goto, K. (2023). Effect of Amino Acid Supplementation on Iron Regulation after Endurance Exercise. Nutrients, 15(23), 4924. https://doi.org/10.3390/nu15234924