1. Introduction
Aging is characterized by a progressive decline in physiological functions over time. The etiology of aging is multifactorial, including genetic, hormonal, free radical, and autoimmune factors. The free radical theory of aging has been widely supported by previous studies, as the accumulation of oxidative stress is closely related to aging. The brain is particularly vulnerable to the detrimental effects of oxidative stress, which can ultimately result in impaired cognitive function and the development of dementia [
1]. Previous studies have demonstrated that intraperitoneal (i.p.) administration of high doses of D-galactose can result in dysfunction of the mitochondrial electron transport chain, leading to elevated levels of advanced glycation end products (AGEs) and superoxide radicals. This, in turn, triggers the generation of reactive oxygen species (ROS) and subsequent oxidative stress. Oxidative stress is known to promote the expression of inflammatory cytokine markers, including cyclooxygenase-2 (COX-2), inducible nitric oxide synthase (iNOS), tumor necrosis factor-alpha (TNF-α), interleukin-1β (IL-1β), interleukin-6 (IL-6), and nuclear factor kappa-light-chain-enhancer of activated B (NF-κB). As a consequence, mitochondrial function is compromised and the activity of the JNK pathway is heightened, ultimately leading to apoptosis and nerve damage in the brain [
2].
Deep ocean water (DOW) is seawater found below the thermocline of the ocean, typically at depths greater than 200 m. This water is characterized by its low temperature, cleanliness, and high concentration of minerals and salt nutrients. Past studies have demonstrated the potential for DOW to prevent obesity [
3], cardiovascular disease [
4], and anti-atherosclerosis [
5] developments. In a previous study, it was discovered that the use of DOW in the fermentation process of
Cordyceps militaris resulted in enhanced efficacious ingredients, including adenosine and cordycepin. These compounds were found to reduce the expression of fibrosis-related factors by inhibiting thioacetamide (TAA)-induced inflammatory factor expression and increasing PPAR-γ activity [
3]. Animal experiments were conducted to investigate the effects of DOW on liver fibrosis using
Antrodia camphorata-fermented products cultured with DOW or ultrapure water. The group fed with DOW-cultured products showed a significantly greater inhibitory effect on lipid peroxidation, ROS, iNOS, and TNF-α compared to the group fed with
Antrodia camphorata fermented products cultured with ultrapure water [
6].
Cordyceps cicadae is a traditional Chinese medicinal fungus that can be cultured in a silkworm, cereal, and potato glucose medium. Previous studies have identified several effective components of
Cordyceps cicadae, including adenosine, N6-(2-hydroxyethyl)-adenosine (HEA), polysaccharides, and ergosterol, which possess antioxidant, anti-inflammatory, and antiaging properties. In an experiment using LPS-induced inflammation in RAW 264.7 macrophages, treatment with the methanol extract of
C. cicadae effectively reduced the inflammatory factors COX-2, Prostaglandin E2 (PGE2), IL-1β, and TNF-α. Furthermore, HEA extracted from
C. cicadae inhibited the NF-κB signaling pathway to achieve anti-inflammatory effects [
7]. Prior research has demonstrated that in H
2O
2-induced oxidative damage experiments in PC12 cells, HEA improves cell viability and reduces LDH release, MMP deficiency, and ROS generation caused by H
2O
2 toxicity. HEA also enhances the activity of antioxidant enzymes and suppresses the production of inflammatory factors by inhibiting lipid peroxidation [
8]. A previous study used DOW-fermented
C. cicadae (DCC) mycelium to induce Alzheimer’s disease by intracerebral injection of amyloid β-protein 40 in rats. After 28 days of testing, the rats showed improvements in memory and learning, as well as brain damage. DCC can significantly improve the absorption of magnesium ions in the brain by increasing the expression of magnesium transporters, thereby achieving the effect of improving Alzheimer’s disease [
9].
Based on the above, it is evident that C. cicadae possess potential anti-inflammatory and antioxidant properties. Co-culturing with DOW can enhance its preventive and ameliorating effects on brain injury. However, oxidative stress is a critical factor in brain aging and cognitive decline. This study aims to investigate the efficacy of using C. cicadae mycelium products cultured in DOW to prevent D-galactose-induced brain injury and memory impairment in rats, with the aim of developing fermentation products that can enhance brain neuroprotection.
2. Materials and Methods
2.1. Chemicals
Potato dextrose agar (PDA) and potato dextrose broth (PDB) were purchased from Difco Co. (Detroit, MI, USA). D-galactose and N(6)-(2-Hydroxyethyl) adenosine (HEA) (>95% purity) were purchased from Sigma Chemical Co. (St Louis, MO, USA). Mouse TNF-α protein (50349-MNAE), mouse IL-6 protein (50349-MNAE), mouse IL-1β protein (50349-MNAE), and rat iNOS protein (Q06518) were purchased from SinoBiological Inc. (North Wales, PA, USA). The mouse anti-rat IL-6 monoclonal antibody (sc-57315) and the mouse anti-human IL-1β monoclonal antibody (sc-32294) were purchased from Santa Cruz Biotechnology, Inc. (Dallas, TX, USA). The rabbit iNOS antibody (FNab04325), the rabbit anti-rat COX-2 polyclonal antibody (RPD284Ra01), and the rat COX-2 protein (RPD284Ra01) were purchased from Cloud-Clone Corp. (Wuhan, China).The rRabbit anti-mouse TNF-α polyclonal antibody (AB2148P) was purchased from the EMD Millipore Corporation (Temecula, CA, USA). Peroxidase conjugated antibodies (31460) were purchased from Pierce Biotechnology (Rockford, IL, USA).
2.2. The Source of DOW
The concentrated deep ocean water (DOW) utilized in this study was obtained from the astern Taiwan Deep Seawater Innovation Research Center located in Taitung, Taiwan. The DOW was extracted from a depth of 670 m in the Pacific Ocean near eastern Taiwan. In a previous study conducted by our research group, it was determined that the DOW containing 20.65 mg/L Mg
2+ would be classified as double-strength DOW. For this present study, a 15-fold DOW was prepared by diluting concentrated DOW (82640 mg/L Mg
2+) with ultrapure water (UPW). The 15X DOW contained 309.75 mg/L Mg
2+, 24.08 mg/L Na
+, 36.56 mg/L K
+, 104.95 mg/L Ca
2+, 0.0012 mg/L Zn
2+, and 0.0024 mg/L Cu
2+. These concentrations of trace elements and minerals were analyzed and reported in a study conducted by Wu and Lee in 2021 [
9].
2.3. Sample Preparation
The UPW medium with 75.25 mg/L of Mg2+ concentration was made by dissolving a potato dextrose broth (PDB) medium in ultrapure water. The DOW medium with 385 mg/L of Mg2+ concentration was made by dissolving a PDB medium in 15X-DOW with 309.75 mg/L of Mg2+. The MgCl2 medium with 385 mg/L of Mg2+ concentration was made by dissolving a PDB medium in a MgCl2 solution with 309.75 mg/L of Mg2+.
The C. cicadae NTTU 868 fermentation product was obtained from a 1.5 L submerged fermentation. C. cicadae NTTU 868 was seed cultured in PDB at 23 °C for 3 days, then inoculated (10%) into the UPW medium, DOW medium, or MgCl2 medium and cultured at 23 °C for 10 days. After fermentation, C. cicadae fermentation products cultured with UPW (UCC), DOW (DCC), and magnesium chloride solution (MgCC) were obtained, respectively.
To obtain the mycelium, the fermentation product was centrifuged at 10,000× g for 15 min at +4 °C. The mycelium was dried at 50 °C, ground into powder, and stored at room temperature. The supernatant (culture fluid) obtained after centrifugation was stored at +4 °C. All submerged fermentation products (including mycelium and culture fluid) were homogenized to prepare animal test samples, the dry weight of the mycelium was calculated proportionally, and the weight per milliliter of mycelium was determined before preparing and administering the test samples.
Previous experiments have shown that different
C. cicadae NTTU 868 fermentation products have varying levels of HEA, polysaccharides, and intracellular magnesium content. UCC had 0.88 mg/L of HEA, 648 mg/L of polysaccharides and 10.1 mg/L of intracellular magnesium content. DCC had 1.04 mg/L of HEA, 591 mg/L of polysaccharides and 161 mg/L of intracellular magnesium content. MgCC had 1.32 mg/L of HEA, 628 mg/L of polysaccharides and 132 mg/L of intracellular magnesium content [
9]. The main difference between DCC and UCC was the higher accumulation of intracellular and extracellular deep seawater minerals in DCC. MgCC had the same amount of magnesium ion (Mg
2+) as DCC but a lower intracellular magnesium ion (Mg
2+) content than DCC.
2.4. Animals Grouping and Experiment Schedule
Individually housed male Sprague Dawley (SD) rats, aged 8 weeks, were purchased from the BioLasco Co. in Taipei, Taiwan. They were provided with free access to standard laboratory chow (Ralston Purina, St. Louis, MO, USA) and water, and were kept in a temperature-controlled room at 24 °C with a 12 h light–dark cycle (lights on at 6:00). The rats were given food ad libitum throughout the experiment. To form the control group, 42 rats were randomly divided into 6 groups and fed a standard diet (NOR; 4.5% fat, 3.34 kcal/g).
During the experiment, all groups except for the control group (NOR group) were subcutaneously injected with 500 mg/kg b.w. of D-galactose per day and fed with the test substance for 56 days. The test substance was C. cicadae after 10 days of liquid fermentation; the mycelium was obtained, dried at 50 °C and ground into powder. An adult (60 kg) was suggested to consume 250 mL of C. cicadae fermented product. The DCC group was fed with the total C. cicadae product of DOW liquid state fermentation (26 mL/kg b.w./day) every day, and the MgCC group was fed with the DCC group with the same magnesium ion (Mg2+) content as the DCC group. The C. cicadae total product of liquid state fermentation (26 mL/kg b.w./day). The UCC group was fed with the total product of C. cicadae (26 mL/kg b.w./day) from UPW fermentation every day, and the HEA group was fed with the functional ingredient standard product of the same HEA concentration as the DCC group every day. Rat body weights were recorded weekly.
After 9 weeks of feeding, the rats were humanely euthanized for tissue collection. The collected mouse serum was immediately stored at −80 °C until further analysis. The brain tissue was dissected into hippocampus and cortex, and the cortex was further divided into left and right hemispheres, and then stored at −80 °C until protein analysis. This study was conducted in accordance with the ethical guidelines established by the Animal Care and Research Ethics Committee of National Taitung University and was approved by the same committee.
2.5. Animal Learning and Memory Tests
2.5.1. Morris Water Maze Task
The water maze test was pioneered by the British psychologist Morris in 1981 to study the learning, memory, and spatial cognition abilities of rats with damaged brain regions such as the hippocampus. In this test, the animals’ abilities can be evaluated by observing and recording their swimming time for searching the resting platform under the water after entering the water. The test apparatus is a circular pool with a diameter of 168 cm and a water depth of 40 cm, and it contains a movable resting platform that is 10 cm in diameter and 5 cm above the water surface. The swimming pool is divided into quadrants I, II, III, and IV, with 5 starting points. The rest platform is located in the center of the third quadrant to avoid animals seeing the platform and affecting the experiment. During the experiment, a video camera (DCM-LX5, Panasonic Co, Osaka, Japan) was set up above the center of the swimming pool to record the time when the experimental animals were looking for the rest platform and swimming lane [
10].
2.5.2. Passive Avoidance Task
The passive avoidance test assesses the memory and learning abilities of rats. It consists of two equally sized rooms, one dark and one light, separated by a movable door. A metal rod connected to an electric current supply is present in the dark room to deliver an electric shock. Rats are placed in the bright room and allowed to acclimate before the door is opened to enter the dark room. The time spent by the rats in the bright room is recorded, and they receive an electric shock upon entering the dark room. The test is repeated thrice daily for three consecutive days [
11].
2.6. Preparation of Brains
The method for brain processing was adapted from a 2021 study by Wu and Lee. The cerebral cortex and hippocampus were carefully isolated from the whole brains on ice and any blood was removed. The isolated tissues were then snap-frozen using liquid nitrogen and stored at −80 °C until further use. For protein extraction in preparation for immunoblotting, 100 mg of tissue was homogenized in 1.0 mL of lysis buffer containing various reagents, including Triton X-100, Tris, NaCl, NaF, SDS, deoxycholate, EDTA, EGTA, and Na3VO4. The homogenate was then briefly sonicated for 10 s before being centrifuged at 100,000× g for 30 min. The resulting supernatant was utilized for the immunoblotting assay.
2.7. Enzyme-Linked Immunosorbent Assay
The protein content of the samples was analyzed with a commercially available BCA protein assay kit (23225, Thermo Fisher Scientific Inc., Rockford, IL, USA). The protein expression analysis steps are as follows. To perform the assay, 100 μL of either the sample or a protein standard (TNF-α, IL-6, IL-1β, COX-2, iNOS) was added to each well of a 96-well plate and incubated at 37 °C for 90 min. Next, the samples or protein standards were removed, and 100 μL of biotinylated detection antibody was added to each well. The plate was then incubated at 37 °C for 60 min. After removing the sample and washing the plate, 100 μL of HRP conjugate was added to each well and the plate was incubated at 37 °C for 30 min. Following the removal of the sample and further washing of the plate, 90 μL of the subtract reagent was added to each well and the plate was incubated at 37 °C for 15 min. Finally, 50 μL of 2 N sulfuric acid was added to the plate to terminate the reaction, and the absorbance was measured at 450 nm using a full-spectrum microplate analyzer (51119300, MultiskanTW Go, Thermo Fisher Scientific Inc., Waltham, MA, USA).
GPx, GRd, and SOD activities were analyzed with commercial kits (Randox Laboratories Ltd., Antrim, UK). Presenilin1 and GFAP protein content were analyzed with commercially available ELISA kits (Wuhan Fine Biotech Co., Ltd., Wuhan, China).
2.8. Statistical Analysis
Data are presented as mean ± standard deviation. Analysis of variance with Duncan’s test and the one-way ANOVA test was determined using SPSS version 12.0 software (SPSS Institute, Inc., Chicago, IL, USA). Differences at p < 0.05 were considered statistically significant.
4. Discussion
The prolonged excessive intake of sugar can lead to oxidative stress, which, if not effectively mitigated, can result in inflammation and abnormal mitochondrial function. In turn, both inflammation and mitochondrial dysfunction can cause nerve cell apoptosis, leading to cognitive impairment [
2]. Additionally, excessive intake of DG (a sugar alcohol) has been shown to increase the expression of proteins related to inflammation, such as COX-2, iNOS, NOS-2, IL-1β, IL-6, TNF-α, and NF-κB, in the brains of Kunming mice or Sprague Dawley rats. These effects were observed after the administration of DG at doses ranging from 100 to 500 mg/kg/day for 6 to 8 weeks. Moreover, the DG-treated group exhibited higher levels of reactive oxygen species (ROS) and malondialdehyde (MDA), as well as reduced antioxidant enzymes (e.g., SOD, CAT, and GSH) and total antioxidant capacity. Consequently, the synthesis of respiratory chain enzymes and ATP decreased, and mitochondrial DNA was mutated, leading to structural damage [
21].
The current study revealed that the negative control group treated with D-galactose (DG) exhibited less exploration time in the target quadrant and wandering paths than the NOR group during the spatial exploration test, indicating impaired memory and learning ability. Moreover, the negative control DG group showed a significant increase in the time to find a plateau compared to the NOR group during the working memory test, which evaluates short-term memory. Additionally, the activities of antioxidant enzymes such as SOD, GRd, and GPx were found to be decreased, while the levels of inflammatory factors, including TNF-α, IL-6, and IL-1β, showed an upward trend. The inflammatory response stimulated the production of iNOS and COX-2 oxidizing factors. Furthermore, the expression levels of aging factors GFAP and Presenilin 1 were also significantly increased after the administration of D-galactose. Overall, excessive intake of D-galactose can diminish antioxidant enzyme activity, promote inflammation, and accelerate the aging process.
Prior studies have demonstrated that
C. cicadae ethanolic extract enhances cell survival in glutamate-induced PC12 cell senescence experiments. This is achieved by reducing intracellular superoxide free radicals, inhibiting lactate dehydrogenase release, and increasing GSH-Px and SOD activity.
C. cicadae have been shown to possess neuroprotective and antiaging properties [
22]. Additionally,
C. cicadae exhibit in vitro antioxidant activity by scavenging DPPH and superoxide anion (O
2−) free radicals, thereby possessing antioxidant capacity and antiaging potential. In vitro studies also reveal that the polysaccharide of
C. cicadae significantly enhances cell survival and reduces intracellular ROS generation in glutamic acid-induced oxidative damage in PC12 cells [
23]. Previous research indicates that HEA scavenges ROS to protect PC12 cells from oxidative damage caused by H
2O
2, primarily by enhancing GSH-Px and SOD performance, highlighting its antioxidant capacity [
8]. Our study demonstrates significant improvement in the activities of GPx and GRd in both DCC and HEA groups, with the MgCC group displaying better effects on SOD and GRd activity. Conversely, the UCC group exhibited no significant antioxidant capacity. It is hypothesized that CC fermented with DOW can enhance the regulation of biological efficacy components.
Studies have demonstrated that TNF-α upregulates the expression of IL-6 in CCl4-induced liver fibrosis. Treatment with
C. cicadae NTTU 868 mycelium and its bioactive components, HEA and
C. cicadae NTTU 868 mycelium, inhibits the TNF-α/IL-6 pathway [
24]. In previous studies, magnesium sulfate at concentrations of 5 and 10 mmol/L significantly inhibited nitric oxide expression in LPS-activated microglia, as well as the expression of prostaglandin E2, IL-1β, TNF-α, and inducible nitric oxide synthase mRNA [
25].
C. cicadae fermented with DOW can reduce the production of inflammatory factors in the cerebral cortex, downregulate TNF-α expression, and prevent inflammation-induced apoptosis and nerve damage. MgCC and HEA had a more pronounced effect, followed by the DCC group.
C. cicadae and HEA have well-known anti-inflammatory properties, and magnesium ions are among the ions that effectively inhibit the production of inflammatory factors. Following an inflammatory response, additional cytokines, such as iNOS, COX-2, etc., are induced. The MgCC and HEA groups demonstrated significantly downregulated expression of iNOS, while COX-2 expression was downregulated in the DCC, MgCC, and HEA groups. Effective regulation of inflammatory factors indeed reduces the generation of subsequent oxidative factors.
Previous studies have shown that mutations in presenilin 1 lead to abnormal γ-secretase activity and increased deposition of Aβ42, which is a causative protein in Alzheimer’s disease [
26].
C. cicadae and HEA fermented with DOW can reduce the production of GFAP and presenilin 1 in the cerebral cortex, slowing brain and nerve damage caused by aging factors and reducing the occurrence of senile diseases. MgCC showed superior regulation ability. Magnesium ions have been shown to promote synaptic plasticity and improve learning and memory [
27], and they are effective in inhibiting aging-related factors.
C. cicadae have neuroprotective properties and slow the production of aging-related factors.
This study investigates how minerals in DOW enhance the antiaging effects of
C. cicadae fermentation products. Minerals in DOW such as magnesium, calcium, selenium, manganese, and others have important roles in human health and disease prevention. In recent years, more and more studies have shown that minerals in DOW can regulate cellular metabolism, antioxidant stress, inflammation inhibition, and neural function improvement among other physiological processes. Magnesium, calcium, selenium, and manganese are essential minerals for the human body, and they play important roles in the function and health of the nervous system. Magnesium is an important ion that participates in regulating neurotransmission, energy metabolism, antioxidant, and anti-inflammatory processes. Magnesium deficiency can cause neuronal damage and death, increasing the risk of neurodegenerative diseases such as Alzheimer’s disease. Magnesium supplementation may be beneficial for protecting neurons, reducing chronic inflammation, and improving cognitive function [
28]. Calcium is an important signaling molecule that participates in regulating neuronal excitability, neurotransmitter release, activating signal transduction, and other processes. Calcium imbalance can cause neuronal apoptosis or necrosis, promoting protein aggregation and oxidative stress, increasing the risk of neurodegenerative diseases such as Parkinson’s disease. Calcium supplementation may be beneficial for maintaining the normal neuronal function, inhibiting neurotoxicity, and improving learning and memory [
29]. Selenium is a powerful antioxidant that participates in synthesizing selenoproteins and glutathione and other molecules, protecting neurons from free radical attacks. Selenium deficiency or excess can both cause nervous system damage, increasing the risk of dementia and other diseases. Selenium supplementation may be beneficial for enhancing antioxidant capacity, reducing protein aggregation, and improving cognitive function [
30,
31]. Manganese is a trace element that has important functions for the nervous system under normal conditions, such as synthesizing dopamine and other neurotransmitters, regulating energy metabolism and so on. However, manganese excess can cause manganese poisoning, resulting in neurodegenerative changes such as Parkinsonian syndrome. Manganese poisoning affects protein folding, cytochrome c oxidase activity, neurotransmitter metabolisms, and other processes, leading to neuronal damage and death. Manganese supplementation may have some help in preventing or treating some neurodegenerative diseases, but more research and evidence are needed to support this hypothesis.
In a previous study of DOW, minerals in DOW such as magnesium and calcium can promote mitochondrial biogenesis, increasing mitochondrial DNA (mtDNA) content and cytochrome c oxidase activity through the AMPK activation signaling pathway [
32]. Mitochondria are key components of cellular energy metabolism and apoptosis, and their dysfunction is related to neurodegenerative diseases. Our previous studies have suggested that the level of magnesium ions in the brain is related to the formation of amyloid and phosphorylated tau proteins [
9]. This study used minerals in DOW to cultivate
C. cicadae mycelium, and found that these minerals not only promoted mycelial growth, but also were absorbed by mycelia and increased bioavailability. This study compared the fermentation products of
C. cicadae cultivated with magnesium chloride and ultrapure water, and found that the former had better antioxidant and anti-inflammatory effects. Therefore, this study speculated that organic magnesium might help aging animals absorb more magnesium ions, thereby improving brain aging. In addition, this study also confirmed that minerals in DOW could be absorbed through
C. cicadae fermentation products, and had a positive effect on alleviating brain damage and aging caused by Aβ40 [
9].
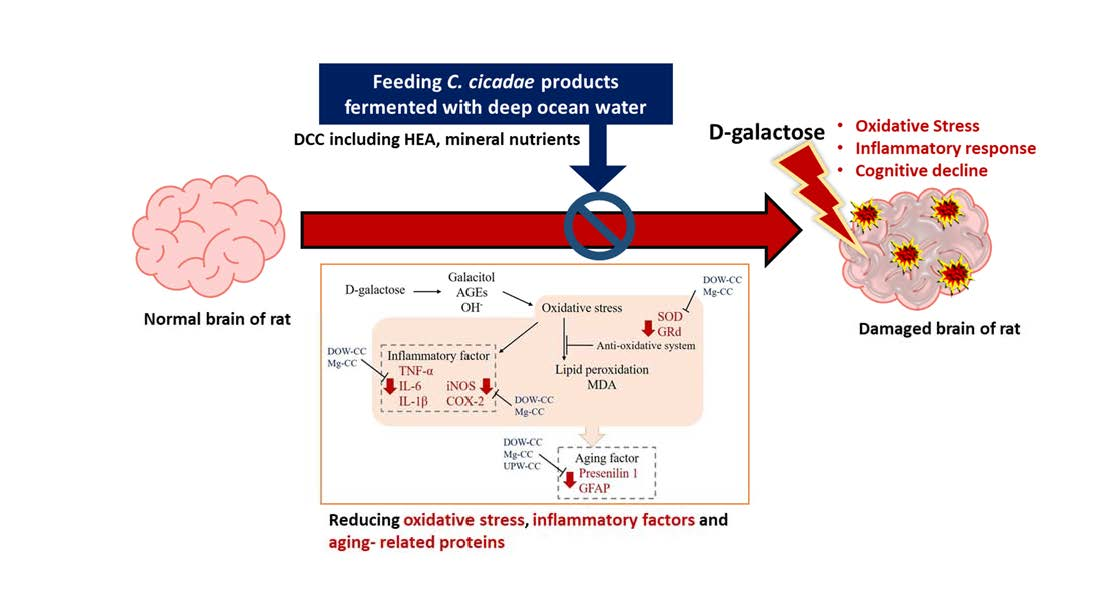
In conclusion, DOW-fermented
C. cicadae and HEA were found to increase SOD and GRd activity, scavenge free radicals, and reduce the inflammatory response caused by oxidative stress (
Figure 8). MgCC had superior efficacy. MgCC also inhibited TNF-α to reduce the expression of downstream inflammatory response mediators iNOS and COX-2, and had a regulatory effect on the expression of aging-related factors GFAP and presenilin 1. These findings demonstrate that the regulation of the expression of anti-inflammatory, antioxidant, and aging-related factors of
C. cicadae products fermented with DOW can prevent aging and show significant improvement in efficacy.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







