
Effect of a Higher-Protein Nut versus Higher-Carbohydrate Cereal Enriched Diet on the Gut Microbiomes of Chinese Participants with Overweight and Normoglycaemia or Prediabetes in the Tū Ora Study

 , ,
, ,  , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Faecal Sample Collection
2.2. DNA Extraction
2.3. 16S rRNA Gene-Targeted PCR and Sequencing
2.4. Shotgun Metagenome Sequencing
2.5. Bioinformatic and Statistical Analysis of Sequence Datasets
3. Results
3.1. Participant Demographics at Baseline
3.2. Taxonomic Composition of 16S rRNA Gene Amplicon Sequencing Data
3.3. Factors Affecting 16S rRNA Gene-Based Microbiota Composition
3.4. Taxonomic Composition and Functional Potential of the Faecal Microbiome Based on Shotgun Metagenomic Sequencing
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Therdtatha, P.; Shinoda, A.; Nakayama, J. Crisis of the Asian gut: Associations among diet, microbiota, and metabolic diseases. Biosci. Microbiota Food Health 2022, 41, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Pingali, P. Westernization of Asian diets and the transformation of food systems: Implications for research and policy. Food Policy 2007, 32, 281–298. [Google Scholar] [CrossRef]
- Graciliano, N.G.; Silveira, J.; Oliveira, A.C.M. The consumption of ultra-processed foods reduces overall quality of diet in pregnant women. Cad. Saude Publica 2021, 37, e00030120. [Google Scholar] [CrossRef]
- Ojeda-Granados, C.; Barchitta, M.; La Rosa, M.C.; La Mastra, C.; Roman, S.; Panduro, A.; Agodi, A.; Maugeri, A. Evaluating dietary patterns in women from Southern Italy and Western Mexico. Nutrients 2022, 14, 1603. [Google Scholar] [CrossRef] [PubMed]
- Di Novi, C.; Marenzi, A. Improving health and sustainability: Patterns of red and processed meat consumption across generations. Health Policy 2022, 126, 1324–1330. [Google Scholar] [CrossRef]
- Magliona, D.; Boyko, E.; Scientific Committee. IDF Diabetes Atlas, 10th ed.; International Diabetes Federation: Brussels, Belgium, 2021. [Google Scholar]
- Obesity and Overweight: Fact Sheet. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 24 March 2024).
- Yang, H.K.; Han, K.; Kwon, H.S.; Park, Y.M.; Cho, J.H.; Yoon, K.H.; Kang, M.I.; Cha, B.Y.; Lee, S.H. Obesity, metabolic health, and mortality in adults: A nationwide population-based study in Korea. Sci. Rep. 2016, 6, 30329. [Google Scholar] [CrossRef]
- Sigit, F.S.; Tahapary, D.L.; Trompet, S.; Sartono, E.; Willems van Dijk, K.; Rosendaal, F.R.; de Mutsert, R. The prevalence of metabolic syndrome and its association with body fat distribution in middle-aged individuals from Indonesia and the Netherlands: A cross-sectional analysis of two population-based studies. Diabetol. Metab. Syndr. 2020, 12, 2. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Li, W.; Lun, Z.; Zhang, H.; Sun, Z.; Kanu, J.S.; Qiu, S.; Cheng, Y.; Liu, Y. Prevalence of metabolic syndrome in Mainland China: A meta-analysis of published studies. BMC Public Health 2016, 16, 296. [Google Scholar] [CrossRef] [PubMed]
- Lear, S.A.; Humphries, K.H.; Kohli, S.; Chockalingam, A.; Frohlich, J.J.; Birmingham, C.L. Visceral adipose tissue accumulation differs according to ethnic background: Results of the Multicultural Community Health Assessment Trial (M-CHAT). Am. J. Clin. Nutr. 2007, 86, 353–359. [Google Scholar] [CrossRef] [PubMed]
- Nazare, J.A.; Smith, J.D.; Borel, A.L.; Haffner, S.M.; Balkau, B.; Ross, R.; Massien, C.; Alméras, N.; Després, J.P. Ethnic influences on the relations between abdominal subcutaneous and visceral adiposity, liver fat, and cardiometabolic risk profile: The International Study of Prediction of Intra-Abdominal Adiposity and Its Relationship with Cardiometabolic Risk/Intra-Abdominal Adiposity. Am. J. Clin. Nutr. 2012, 96, 714–726. [Google Scholar] [PubMed]
- Ramachandran, A.; Chamukuttan, S.; Shetty, S.A.; Arun, N.; Susairaj, P. Obesity in Asia—Is it different from rest of the world. Diabetes Metab. Res. Rev. 2012, 28 (Suppl. S2), 47–51. [Google Scholar] [CrossRef] [PubMed]
- Thomas, E.L.; Parkinson, J.R.; Frost, G.S.; Goldstone, A.P.; Doré, C.J.; McCarthy, J.P.; Collins, A.L.; Fitzpatrick, J.A.; Durighel, G.; Taylor-Robinson, S.D.; et al. The missing risk: MRI and MRS phenotyping of abdominal adiposity and ectopic fat. Obesity 2012, 20, 76–87. [Google Scholar] [CrossRef] [PubMed]
- Sniderman, A.D.; Bhopal, R.; Prabhakaran, D.; Sarrafzadegan, N.; Tchernof, A. Why might South Asians be so susceptible to central obesity and its atherogenic consequences? The adipose tissue overflow hypothesis. Int. J. Epidemiol. 2007, 36, 220–225. [Google Scholar] [CrossRef] [PubMed]
- West, J.; Lawlor, D.A.; Fairley, L.; Bhopal, R.; Cameron, N.; McKinney, P.A.; Sattar, N.; Wright, J. UK-born Pakistani-origin infants are relatively more adipose than white British infants: Findings from 8704 mother-offspring pairs in the Born-in-Bradford prospective birth cohort. J. Epidemiol. Community Health 2013, 67, 544–551. [Google Scholar] [CrossRef] [PubMed]
- Tota-Maharaj, R.; Blaha, M.J.; Zeb, I.; Katz, R.; Blankstein, R.; Blumenthal, R.S.; Budoff, M.J.; Nasir, K. Ethnic and sex differences in fatty liver on cardiac computed tomography: The multi-ethnic study of atherosclerosis. Mayo Clin. Proc. 2014, 89, 493–503. [Google Scholar] [CrossRef] [PubMed]
- Zamboni, M.; Rossi, A.P.; Fantin, F.; Budui, S.L.; Zoico, E.; Zamboni, G.A.; Mazzali, G. Predictors of ectopic fat in humans. Curr. Obes. Rep. 2014, 3, 404–413. [Google Scholar] [CrossRef] [PubMed]
- Tillin, T.; Sattar, N.; Godsland, I.F.; Hughes, A.D.; Chaturvedi, N.; Forouhi, N.G. Ethnicity-specific obesity cut-points in the development of Type 2 diabetes—A prospective study including three ethnic groups in the United Kingdom. Diabet. Med. 2015, 32, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Saxena, A.; Tiwari, P.; Wahi, N.; Kumar, A.; Mathur, S.K. The common pathophysiologic threads between Asian Indian diabetic’s ‘Thin Fat Phenotype’ and partial lipodystrophy: The peripheral adipose tissue transcriptomic evidences. Adipocyte 2020, 9, 253–263. [Google Scholar] [CrossRef] [PubMed]
- Sequeira, I.R.; Yip, W.C.; Lu, L.W.W.; Jiang, Y.; Murphy, R.; Plank, L.D.; Cooper, G.J.S.; Peters, C.N.; Lu, J.; Hollingsworth, K.G.; et al. Pancreas fat, an early marker of metabolic risk? A magnetic resonance study of Chinese and Caucasian women: TOFI_Asia Study. Front. Physiol. 2022, 13, 819606. [Google Scholar] [CrossRef]
- Luo, C.; Zhang, Y.; Ding, Y.; Shan, Z.; Chen, S.; Yu, M.; Hu, F.B.; Liu, L. Nut consumption and risk of type 2 diabetes, cardiovascular disease, and all-cause mortality: A systematic review and meta-analysis. Am. J. Clin. Nutr. 2014, 100, 256–269. [Google Scholar] [CrossRef]
- Lattimer, J.M.; Haub, M.D. Effects of dietary fiber and its components on metabolic health. Nutrients 2010, 2, 1266–1289. [Google Scholar] [CrossRef] [PubMed]
- Andoh, A.; Tsujikawa, T.; Fujiyama, Y. Role of dietary fiber and short-chain fatty acids in the colon. Curr. Pharm. Des. 2003, 9, 347–358. [Google Scholar] [CrossRef]
- Wong, J.M.; de Souza, R.; Kendall, C.W.; Emam, A.; Jenkins, D.J. Colonic health: Fermentation and short chain fatty acids. J. Clin. Gastroenterol. 2006, 40, 235–243. [Google Scholar] [CrossRef]
- Sorensen, M.D.; Hsi, R.S.; Chi, T.; Shara, N.; Wactawski-Wende, J.; Kahn, A.J.; Wang, H.; Hou, L.; Stoller, M.L. Dietary intake of fiber, fruit and vegetables decreases the risk of incident kidney stones in women: A Women’s Health Initiative report. J. Urol. 2014, 192, 1694–1699. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.K.; Alasalvar, C.; Shahidi, F. Review of dried fruits: Phytochemicals, antioxidant efficacies, and health benefits. J. Funct. Foods 2016, 21, 113–132. [Google Scholar] [CrossRef]
- Hernández-Alonso, P.; Camacho-Barcia, L.; Bulló, M.; Salas-Salvadó, J. Nuts and dried fruits: An update of their beneficial effects on type 2 diabetes. Nutrients 2017, 9, 673. [Google Scholar] [CrossRef] [PubMed]
- Da Porto, A.; Cavarape, A.; Colussi, G.; Casarsa, V.; Catena, C.; Sechi, L.A. Polyphenols rich diets and risk of type 2 diabetes. Nutrients 2021, 13, 1445. [Google Scholar] [CrossRef] [PubMed]
- Cohen, A.E.; Johnston, C.S. Almond ingestion at mealtime reduces postprandial glycemia and chronic ingestion reduces hemoglobin A(1c) in individuals with well-controlled type 2 diabetes mellitus. Metabolism 2011, 60, 1312–1317. [Google Scholar] [CrossRef] [PubMed]
- Jiang, R.; Manson, J.E.; Stampfer, M.J.; Liu, S.; Willett, W.C.; Hu, F.B. Nut and peanut butter consumption and risk of type 2 diabetes in women. JAMA 2002, 288, 2554–2560. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.Y.; Ojo, O.; Wang, L.L.; Wang, Q.; Jiang, Q.; Shao, X.Y.; Wang, X.H. A randomized controlled trial to compare the effect of peanuts and almonds on the cardio-metabolic and inflammatory parameters in patients with type 2 diabetes mellitus. Nutrients 2018, 10, 1565. [Google Scholar] [CrossRef] [PubMed]
- Li, S.C.; Liu, Y.H.; Liu, J.F.; Chang, W.H.; Chen, C.M.; Chen, C.Y. Almond consumption improved glycemic control and lipid profiles in patients with type 2 diabetes mellitus. Metabolism 2011, 60, 474–479. [Google Scholar] [CrossRef] [PubMed]
- Nowrouzi-Sohrabi, P.; Hassanipour, S.; Sisakht, M.; Daryabeygi-Khotbehsara, R.; Savardashtaki, A.; Fathalipour, M. The effectiveness of pistachio on glycemic control and insulin sensitivity in patients with type 2 diabetes, prediabetes and metabolic syndrome: A systematic review and meta-analysis. Diabetes Metab. Syndr. 2020, 14, 1589–1595. [Google Scholar] [CrossRef] [PubMed]
- Kendall, C.W.; Josse, A.R.; Esfahani, A.; Jenkins, D.J. The impact of pistachio intake alone or in combination with high-carbohydrate foods on post-prandial glycemia. Eur. J. Clin. Nutr. 2011, 65, 696–702. [Google Scholar] [CrossRef] [PubMed]
- Kendall, C.W.; West, S.G.; Augustin, L.S.; Esfahani, A.; Vidgen, E.; Bashyam, B.; Sauder, K.A.; Campbell, J.; Chiavaroli, L.; Jenkins, A.L.; et al. Acute effects of pistachio consumption on glucose and insulin, satiety hormones and endothelial function in the metabolic syndrome. Eur. J. Clin. Nutr. 2014, 68, 370–375. [Google Scholar] [CrossRef]
- King, J.C.; Blumberg, J.; Ingwersen, L.; Jenab, M.; Tucker, K.L. Tree nuts and peanuts as components of a healthy diet. J. Nutr. 2008, 138, 1736s–1740s. [Google Scholar] [CrossRef] [PubMed]
- Vassiliou, E.K.; Gonzalez, A.; Garcia, C.; Tadros, J.H.; Chakraborty, G.; Toney, J.H. Oleic acid and peanut oil high in oleic acid reverse the inhibitory effect of insulin production of the inflammatory cytokine TNF-α both in vitro and in vivo systems. Lipids Health Dis. 2009, 8, 25. [Google Scholar] [CrossRef] [PubMed]
- Becerra-Tomás, N.; Paz-Graniel, I.; Hernández-Alonso, P.; Jenkins, D.J.A.; Kendall, C.W.C.; Sievenpiper, J.L.; Salas-Salvadó, J. Nut consumption and type 2 diabetes risk: A systematic review and meta-analysis of observational studies. Am. J. Clin. Nutr. 2021, 113, 960–971. [Google Scholar] [CrossRef] [PubMed]
- LeBlanc, J.G.; Milani, C.; de Giori, G.S.; Sesma, F.; van Sinderen, D.; Ventura, M. Bacteria as vitamin suppliers to their host: A gut microbiota perspective. Curr. Opin. Biotechnol. 2013, 24, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Pedersen, O. Gut microbiota in human metabolic health and disease. Nat. Rev. Microbiol. 2021, 19, 55–71. [Google Scholar] [CrossRef] [PubMed]
- Corbin, K.D.; Carnero, E.A.; Dirks, B.; Igudesman, D.; Yi, F.; Marcus, A.; Davis, T.L.; Pratley, R.E.; Rittmann, B.E.; Krajmalnik-Brown, R.; et al. Reprogramming the human gut microbiome reduces dietary energy harvest. Res. Sq. 2023, preprint. [Google Scholar] [CrossRef] [PubMed]
- Aziz, T.; Hussain, N.; Hameed, Z.; Lin, L. Elucidating the role of diet in maintaining gut health to reduce the risk of obesity, cardiovascular and other age-related inflammatory diseases: Recent challenges and future recommendations. Gut Microbes 2024, 16, 2297864. [Google Scholar] [CrossRef] [PubMed]
- Aziz, T.; Khan, A.A.; Tzora, A.; Voidarou, C.C.; Skoufos, I. Dietary implications of the bidirectional relationship between the gut microflora and inflammatory diseases with special emphasis on irritable bowel disease: Current and future perspective. Nutrients 2023, 15, 2956. [Google Scholar] [CrossRef] [PubMed]
- McNabney, S.M.; Henagan, T.M. Short chain fatty acids in the colon and peripheral tissues: A focus on butyrate, colon cancer, obesity and insulin resistance. Nutrients 2017, 9, 1348. [Google Scholar] [CrossRef] [PubMed]
- Canani, R.B.; Costanzo, M.D.; Leone, L.; Pedata, M.; Meli, R.; Calignano, A. Potential beneficial effects of butyrate in intestinal and extraintestinal diseases. World J. Gastroenterol. 2011, 17, 1519–1528. [Google Scholar] [CrossRef] [PubMed]
- Bultman, S.J. Butyrate consumption of differentiated colonocytes in the upper crypt promotes homeostatic proliferation of stem and progenitor cells near the crypt base. Transl. Cancer Res. 2016, 5 (Suppl. S3), S526–S528. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Liu, Z.; Zhu, C.; Mou, H.; Kong, Q. Nondigestible carbohydrates, butyrate, and butyrate-producing bacteria. Crit. Rev. Food Sci. Nutr. 2019, 59, S130–S152. [Google Scholar] [CrossRef] [PubMed]
- Sapp, P.; Lamendella, R.; Kris-Etherton, P.; Petersen, K. Peanut Intake Enriches Butyrate Producing Bacteria and Expression of a Gene Associated with Butyrate Production in Adults with Elevated Fasting Glucose: An RCT. Curr. Dev. Nutr. 2021, 5 (Suppl. S2), 1178. [Google Scholar] [CrossRef]
- Zhu, L.B.; Zhang, Y.C.; Huang, H.H.; Lin, J. Prospects for clinical applications of butyrate-producing bacteria. World J. Clin. Pediatr. 2021, 10, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Li, Y.; Cai, Z.; Li, S.; Zhu, J.; Zhang, F.; Liang, S.; Zhang, W.; Guan, Y.; Shen, D.; et al. A metagenome-wide association study of gut microbiota in type 2 diabetes. Nature 2012, 490, 55–60. [Google Scholar] [CrossRef]
- Lu, L.W.; Silvestre, M.P.; Sequeira, I.R.; Plank, L.D.; Foster, M.; Middleditch, N.; Acevedo-Fani, A.; Hollingsworth, K.G.; Poppitt, S.D. A higher-protein nut-based snack product suppresses glycaemia and decreases glycaemic response to co-ingested carbohydrate in an overweight prediabetic Asian Chinese cohort: The Tū Ora postprandial RCT. J. Nutr. Sci. 2021, 10, e30. [Google Scholar] [CrossRef] [PubMed]
- Sequeira-Bisson, I.R.L.L.; Silvestre, M.P.; Plank, L.D.; Middleditch, N.; Acevedo-Fani, A.; Parry-Strong, A.; Hollingsworth, K.G.; Tups, A.; Rizwan, M.Z.; Boucsein, A.; et al. Glycemic response to a higher-protein nut-enriched diet in Asian Chinese with overweight plus normal or high glycaemia and ectopic organ fat. Nutrients 2024, submitted.
- Guasch-Ferré, M.; Li, J.; Hu, F.B.; Salas-Salvadó, J.; Tobias, D.K. Effects of walnut consumption on blood lipids and other cardiovascular risk factors: An updated meta-analysis and systematic review of controlled trials. Am. J. Clin. Nutr. 2018, 108, 174–187. [Google Scholar] [CrossRef] [PubMed]
- Costea, P.I.; Zeller, G.; Sunagawa, S.; Pelletier, E.; Alberti, A.; Levenez, F.; Tramontano, M.; Driessen, M.; Hercog, R.; Jung, F.E.; et al. Towards standards for human fecal sample processing in metagenomic studies. Nat. Biotechnol. 2017, 35, 1069–1076. [Google Scholar] [CrossRef] [PubMed]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2, High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Wright, E. Using DECIPHER v2.0 to Analyze Big Biological Sequence Data in R. R J. 2016, 8, 352–359. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed]
- Heidrich, V.; Karlovsky, P.; Beule, L. ‘SRS’R Package and ‘q2-srs’ QIIME 2 Plugin: Normalization of microbiome data using scaling with ranked subsampling (SRS). Appl. Sci. 2021, 11, 11473. [Google Scholar] [CrossRef]
- BBMap. Available online: https://sourceforge.net/projects/bbmap/ (accessed on 14 January 2024).
- Bağcı, C.; Patz, S.; Huson, D.H. DIAMOND+MEGAN: Fast and easy taxonomic and functional analysis of short and long microbiome sequences. Curr. Protoc. 2021, 1, e59. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Simpson, G.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.; O’Hara, R.; Solymos, P.; Stevens, M.H.H.; Szoecs, E.; et al. Vegan: Community Ecology Package. R package version 2.6-6. 2024. Available online: https://CRAN.R-project.org/package=vegan (accessed on 24 March 2024).
- Wickham, H.; Wickham, H. Data Analysis; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Louis, P.; Flint, H.J. Diversity, metabolism and microbial ecology of butyrate-producing bacteria from the human large intestine. FEMS Microbiol. Lett. 2009, 294, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.; Lee, G.; Son, H.; Koh, H.; Kim, E.S.; Unno, T.; Shin, J.H. Butyrate producers, “The Sentinel of Gut”: Their intestinal significance with and beyond butyrate, and prospective use as microbial therapeutics. Front. Microbiol. 2022, 13, 1103836. [Google Scholar] [CrossRef] [PubMed]
- Fujio-Vejar, S.; Vasquez, Y.; Morales, P.; Magne, F.; Vera-Wolf, P.; Ugalde, J.A.; Navarrete, P.; Gotteland, M. The gut microbiota of healthy Chilean subjects reveals a high abundance of the Phylum Verrucomicrobia. Front. Microbiol. 2017, 8, 1221. [Google Scholar] [CrossRef] [PubMed]
- Ukhanova, M.; Wang, X.; Baer, D.J.; Novotny, J.A.; Fredborg, M.; Mai, V. Effects of almond and pistachio consumption on gut microbiota composition in a randomised cross-over human feeding study. Br. J. Nutr. 2014, 111, 2146–2152. [Google Scholar] [CrossRef] [PubMed]
- Holscher, H.D.; Guetterman, H.M.; Swanson, K.S.; An, R.; Matthan, N.R.; Lichtenstein, A.H.; Novotny, J.A.; Baer, D.J. Walnut consumption alters the gastrointestinal microbiota, microbially derived secondary bile acids, and health markers in healthy adults: A randomized controlled trial. J. Nutr. 2018, 148, 861–867. [Google Scholar] [CrossRef] [PubMed]
- Vinolo, M.A.R.; Rodrigues, H.G.; Nachbar, R.T.; Curi, R. Regulation of inflammation by short chain fatty acids. Nutrients 2011, 3, 858–876. [Google Scholar] [CrossRef] [PubMed]
- Campos, S.B.; de Oliveira Filho, J.G.; Dos Santos, D.C.; Egea, M.B. Daily intake of baru (Dipteryx alata Vog.), a Cerrado native almond, promotes changes in the intestinal microbiota of individuals with obesity: A pilot study. Food Res. Int. 2024, 187, 114370. [Google Scholar] [CrossRef] [PubMed]


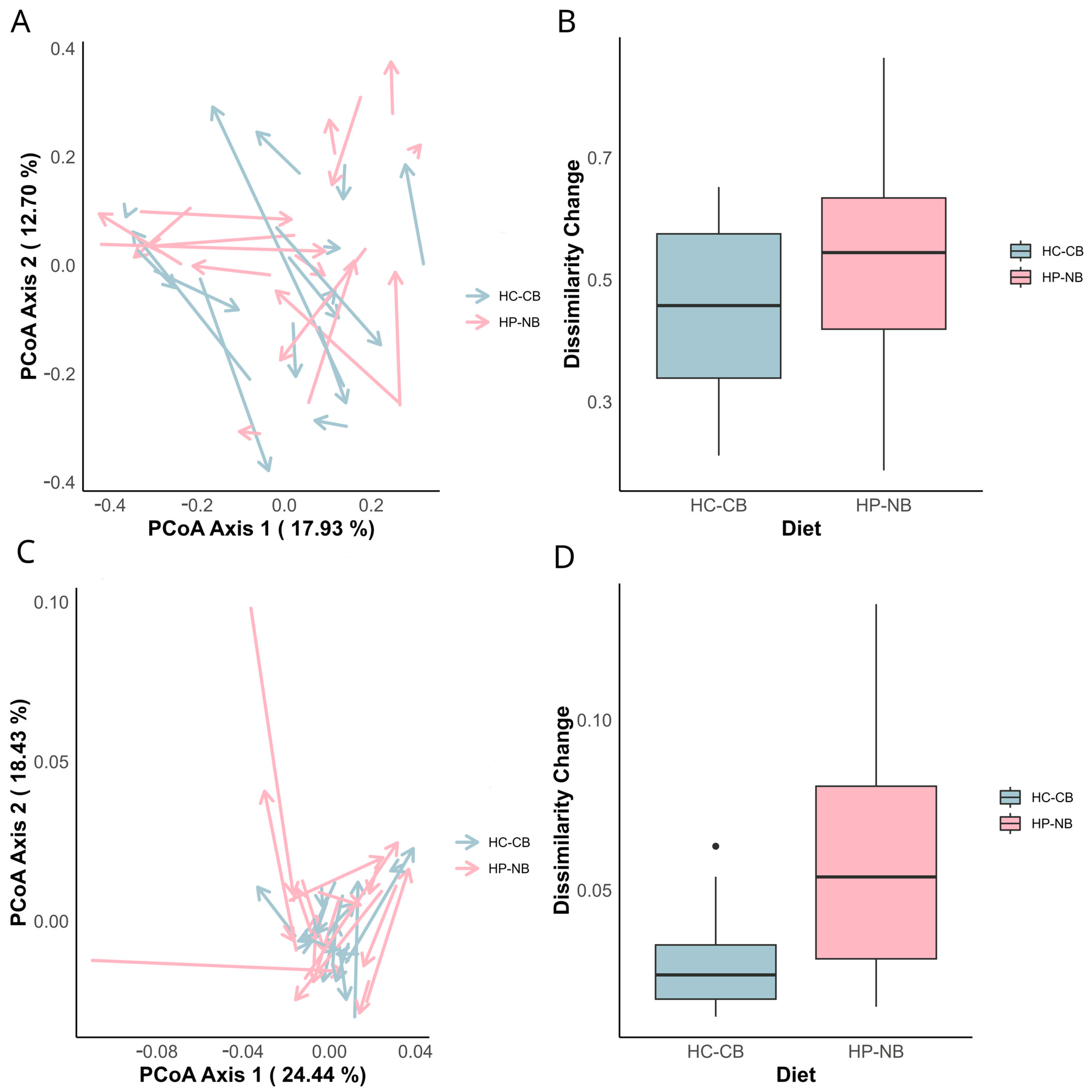
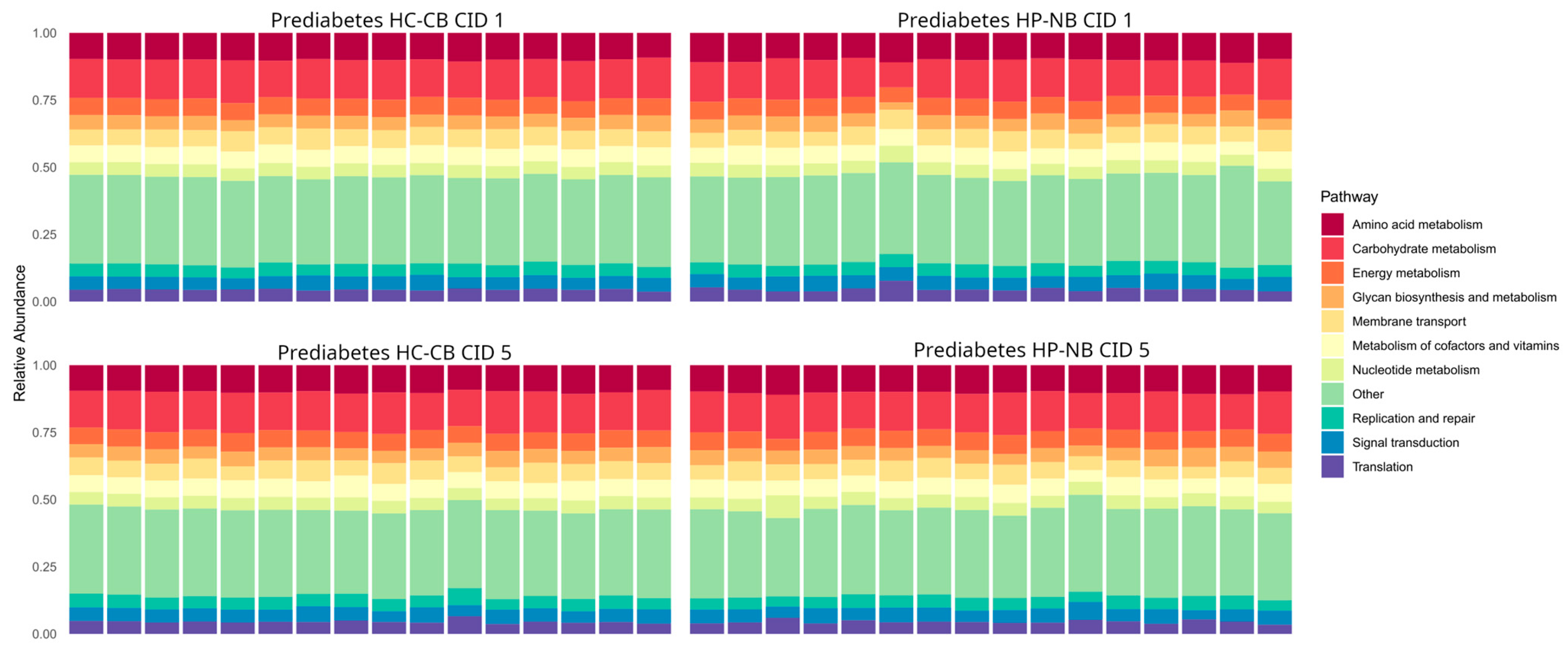

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | All | Diet | p-Value | Glycaemia | p-Value | ||
|---|---|---|---|---|---|---|---|
| HP-NB | HC-CB | Normoglycaemia | Prediabetes | ||||
| n | 84 | 38 | 46 | 52 | 32 | ||
| Age (y) | 47.11, 11.04 | 46.81, 10.44 | 47.35, 11.93 | 0.83 | 45.63, 10.92 | 49.51, 11.06 | 0.12 |
| Sex (M:F) | (38:46) | (22:24) | (14:24) | 0.19 | (17:35) | (21:11) | <0.0001 |
| Height (m) | 1.66, 0.86 | 1.65, 0.09 | 1.67, 0.07 | 0.22 | 1.66, 0.09 | 1.67, 0.07 | 0.61 |
| Body weight (kg) | 76.3, 14.6 | 73.6, 16.3 | 78.4, 11.3 | 0.11 | 75.3, 14.4 | 77.8, 14.5 | 0.46 |
| BMI (kg/m2) | 27.4, 3.63 | 27.0, 3.74 | 27.8, 3.27 | 0.3 | 27.2, 3.27 | 27.8, 3.98 | 0.46 |
| Waist circumf. (cm) | 91.9, 10.9 | 92.8, 11.3 | 90.8, 10.2 | 0.38 | 89.6, 9.3 | 95.6, 11.7 | 0.018 |
| Hip circumf. (cm) | 102.9, 8.2 | 103.8, 8.6 | 101.9, 7.8 | 0.27 | 102.7, 7.5 | 103.3, 9.0 | 0.77 |
| FPG (mmol/L) | 5.51, 0.50 | 5.47, 0.56 | 5.54, 0.43 | 0.48 | 5.21, 0.28 | 6.00, 0.39 | <0.0001 |
| Insulin (μU/mL) | 12.48, 9.29 | 12.12, 8.37 | 12.91, 10.24 | 0.69 | 9.87, 9.85 | 16.71, 11.30 | 0.0015 |
| Sex | Diet | Glycaemic Status | Diet × Glycaemic Status | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| HP-NB | HC-CB | ||||||||||
| All | Male | Female | HP-NB | HC-CB | N | P | N | P | N | P | |
| n | 84 | 38 | 46 | 38 | 46 | 52 | 32 | 23 | 15 | 29 | 17 |
| R2 | 0.004 | 0.012 | 0.007 | 0.007 | 0.007 | 0.008 | 0.007 | 0.012 | 0.014 | 0.019 | 0.021 |
| p-value | 0.447 | 0.017 | 0.73 | 0.31 | 0.821 | 0.107 | 0.995 | 0.471 | 0.903 | 0.077 | 0.548 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Faraj, S.; Sequeira-Bisson, I.R.; Lu, L.; Miles-Chan, J.L.; Hoggard, M.; Barnett, D.; Parry-Strong, A.; Foster, M.; Krebs, J.D.; Poppitt, S.D.; et al. Effect of a Higher-Protein Nut versus Higher-Carbohydrate Cereal Enriched Diet on the Gut Microbiomes of Chinese Participants with Overweight and Normoglycaemia or Prediabetes in the Tū Ora Study. Nutrients 2024, 16, 1971. https://doi.org/10.3390/nu16121971
Faraj S, Sequeira-Bisson IR, Lu L, Miles-Chan JL, Hoggard M, Barnett D, Parry-Strong A, Foster M, Krebs JD, Poppitt SD, et al. Effect of a Higher-Protein Nut versus Higher-Carbohydrate Cereal Enriched Diet on the Gut Microbiomes of Chinese Participants with Overweight and Normoglycaemia or Prediabetes in the Tū Ora Study. Nutrients. 2024; 16(12):1971. https://doi.org/10.3390/nu16121971
Chicago/Turabian StyleFaraj, Saif, Ivana R. Sequeira-Bisson, Louise Lu, Jennifer L. Miles-Chan, Michael Hoggard, Daniel Barnett, Amber Parry-Strong, Meika Foster, Jeremy D. Krebs, Sally D. Poppitt, and et al. 2024. "Effect of a Higher-Protein Nut versus Higher-Carbohydrate Cereal Enriched Diet on the Gut Microbiomes of Chinese Participants with Overweight and Normoglycaemia or Prediabetes in the Tū Ora Study" Nutrients 16, no. 12: 1971. https://doi.org/10.3390/nu16121971
APA StyleFaraj, S., Sequeira-Bisson, I. R., Lu, L., Miles-Chan, J. L., Hoggard, M., Barnett, D., Parry-Strong, A., Foster, M., Krebs, J. D., Poppitt, S. D., Taylor, M. W., & Mathrani, A. (2024). Effect of a Higher-Protein Nut versus Higher-Carbohydrate Cereal Enriched Diet on the Gut Microbiomes of Chinese Participants with Overweight and Normoglycaemia or Prediabetes in the Tū Ora Study. Nutrients, 16(12), 1971. https://doi.org/10.3390/nu16121971