Abstract
Branched-chain amino acids (BCAAs), comprising leucine (Leu), isoleucine (Ile), and valine (Val), are essential nutrients vital for protein synthesis and metabolic regulation via specialized signaling networks. Their association with cardiovascular diseases (CVDs) has become a focal point of scientific debate, with emerging evidence suggesting both beneficial and detrimental roles. This review aims to dissect the multifaceted relationship between BCAAs and cardiovascular health, exploring the molecular mechanisms and clinical implications. Elevated BCAA levels have also been linked to insulin resistance (IR), type 2 diabetes mellitus (T2DM), inflammation, and dyslipidemia, which are well-established risk factors for CVD. Central to these processes are key pathways such as mammalian target of rapamycin (mTOR) signaling, nuclear factor kappa-light-chain-enhancer of activate B cells (NF-κB)-mediated inflammation, and oxidative stress. Additionally, the interplay between BCAA metabolism and gut microbiota, particularly the production of metabolites like trimethylamine-N-oxide (TMAO), adds another layer of complexity. Contrarily, some studies propose that BCAAs may have cardioprotective effects under certain conditions, contributing to muscle maintenance and metabolic health. This review critically evaluates the evidence, addressing the biological basis and signal transduction mechanism, and also discusses the potential for BCAAs to act as biomarkers versus active mediators of cardiovascular pathology. By presenting a balanced analysis, this review seeks to clarify the contentious roles of BCAAs in CVD, providing a foundation for future research and therapeutic strategies required because of the rising prevalence, incidence, and total burden of CVDs.
1. Introduction
Branched-chain amino acids (BCAAs), including leucine, isoleucine, and valine, are essential nutrients that play critical roles in protein synthesis, energy production, and metabolic regulation. While BCAAs are essential for normal physiological processes, emerging research has highlighted their complex involvement in chronic cardiovascular diseases (CVDs), including heart failure (HF), atherosclerosis (AS), coronary artery disease (CAD), and hypertension [1]. The function of BCAAs in chronic CVD is multifaceted and involves intricate interactions with various metabolic pathways, cellular signaling, and mechanisms [2].
Alterations in metabolism and circulating BCAA levels have been observed and are increasingly recognized as potential biomarkers, and they are associated with insulin resistance, dyslipidemia, and endothelial dysfunction [3,4]. Moreover, the role of BCAAs in chronic CVD extends beyond their traditional functions in protein metabolism. BCAAs can modulate cellular signaling pathways, such as the mammalian target of rapamycin (mTOR) pathway, which plays a crucial role in regulating cell growth, proliferation, and metabolism [5]. Dysregulated mTOR signaling, influenced by BCAA availability, has been implicated in adverse cardiac remodeling, fibrosis, and hypertrophy observed in heart failure, as well as other cardiovascular conditions [5].
Furthermore, the interaction between BCAAs and lipid metabolism, particularly through sterol regulatory element-binding proteins (SREBPs) and peroxisome proliferator-activated receptors (PPARs), underscores the intricate metabolic cross-talk in CVD pathogenesis [6,7]. Dysregulation of these transcriptional regulators can lead to lipid accumulation, inflammation, and oxidative stress, all of which contribute to the development and progression of atherosclerosis and coronary artery disease. Besides their role in CVD pathology, recent studies have correlated BCAAs with type 2 diabetes (T2D) [6,7], not only showing their involvement in glycemic control but also their involvement in insulin resistance [3,4].
In this context, the involvement of BCAAs in chronic CVD represents a dynamic interplay between nutrient metabolism, cellular signaling, and disease pathophysiology. Understanding the nuanced roles of BCAAs in cardiovascular health and disease holds promise for identifying novel therapeutic targets and interventions aimed at mitigating metabolic dysfunction and improving outcomes for individuals with chronic cardiovascular conditions. This review aims to explore and elucidate the multifaceted interplay of BCAAs in chronic CVD, highlighting the latest studies about their impact on metabolic regulation, cellular signaling, and disease pathogenesis. By dissecting these complex interactions, we can advance our understanding of CVD mechanisms and identify new avenues for personalized treatment strategies targeting BCAA metabolism and associated pathways.
2. Branched-Chain Amino Acid Synthesis, Metabolism, and Catabolites
BCAAs, specifically leucine, isoleucine, and valine, have gained interest since the mid-1970s and 1980s due to their observed immunomodulatory properties subsequent to their broad implementation as dietary supplements in individuals affected by chronic liver and kidney disease, sepsis or trauma, and muscle-wasting disorders [2,8,9]. As stimulators of the mTOR pathway, notably so in the case of leucine, BCAAs indirectly interfere with numerous other signaling pathways that mediate insulin action, crucial for maintaining glucose homeostasis, protein synthesis, mitochondrial biogenesis, inflammation, and lipid metabolism [10,11]. Despite the numerous in vitro research studies conducted on cellular cultures and human and animal model studies, the results have been contradictory regarding the biological activity of BCAAs [8,12]. Consequently, the intricate mechanisms underlying their effects still need to be more adequately understood [8,12].
Although they constitute about 35% of essential amino acids in mammals, playing a significant role in protein structure, BCAAs cannot be synthesized and must be obtained from dietary sources, representing approximately 20–25% of daily dietary protein intake [13,14]. In order to regulate catabolism in mammals, a tightly controlled metabolic system has evolved, consisting of a cascade of processes that include transamination, irreversible oxidative decarboxylation, and adenosine triphosphate (ATP) generation [15,16].
2.1. Transamination
The initial event in the breakdown of the three aforementioned BCAAs is multifaceted. It involves the conversion of the amino acids to branched-chain α-keto acids (BCKAs) through transamination, a process catalyzed by the cytosolic or mitochondrial enzyme BCAA aminotransferase (BCAT) [17]. The transamination process involves the transfer of specific amino groups to α-ketoglutarate (α-KG), resulting in glutamate and the formation of corresponding BCKAs. The BCKAs derived from leucine, isoleucine, and valine are α-ketoisocaproate (α-KIC), α-keto-β-methylvalerate (α-KMV), and α-ketoisovalerate (α-KIV), respectively [18,19,20,21]. Further, glutamate can be used either as the amino component necessary to form alanine (ALA) or it can be used for ammonia detoxification, resulting in glutamine (GLN), and all of the above-mentioned three components involved in transamination (BCKAs, ALA, and GLN) are released into the bloodstream via muscles [18,19,20,21].
The entire transamination process cannot occur without the action of the BCAT enzyme, along with its two isoforms. BCAT1, the cytosolic one, is predominantly expressed in the brain, while BCAT2, the mitochondrial one, is found in most tissues and particularly skeletal muscle [14,21,22,23]. Due to the reduced activity of BCAT in the liver as opposed to other amino acids, the first step in the catabolism of BCAAs does not occur at this level, and they rapidly reach elevated levels in the systemic circulation after protein ingestion and are even more quickly available to extrahepatic tissues [22]. Transamination is subsequently followed by the decarboxylation and dehydrogenation of the BCKAs to branched-chain acyl-CoA esters, along with the production of CO2 and NADH by the BCKA dehydrogenase complex (BCKDC/BCKDH). This step converts α-KIC, α-KMV, and α-KIV to isovaleryl-CoA, 2-methyl-butyryl-CoA, and isobutyryl-CoA, respectively [14,20,24].
2.2. Decarboxylation
Following the transamination pattern described above, the catalyst of the oxidative decarboxylation reaction is the BCKDH complex, which is a multienzyme complex located in the inner mitochondrial membrane [25,26]. The complex consists of three catalytic components: E1 (thiamine-dependent decarboxylase) for the oxidative decarboxylation of BCKAs, E2 (dihydrolipoyltransacylase) for transferring acyl groups to CoA, and E3 (FAD-dependent dihydrolipoyl dehydrogenase) for electron transfer to NAD+ [25,26].
E1 uses a reduced coenzyme-A substrate for decarboxylation, while E2 uses lipoic acid as an acceptor and transfers it to acetyl-CoA [13,27]. The phosphorylation status of E1 is changed by the different levels of BCKDH kinase (BCKDK) and protein phosphatase Mg2+/Mn2+-dependent 1K (Ppm1k), which then regulates the inhibitory phosphorylation of BCKDH E2 and impacts the flow of substances in the pathway [13,24,25]. E3 acts as a lipoamide dehydrogenase, transferring its hydrogen to NAD+ via FAD [13,27].
The phosphorylation–dephosphorylation mechanism regulates the BCKDH complex, with functional alterations in kinase activity producing a significant impact. BCKDK causes inactivation through phosphorylation, while mitochondrial phosphatase 2C (PP2Cm) catalyzes dephosphorylation, which activates the BCKDH complex [17,28]. The highest BCKDK activity is exhibited by the liver, followed by the kidneys and heart, while the muscles, adipose tissue, and brain have lower activity levels [13,23].
2.3. ATP Generation
Following the decarboxylation of BCKAs induced by the BCKDH complex, the subsequent stages of degradation resemble the process of fatty acid oxidation in terms of adenosine triphosphate (ATP) production [29]. However, each reaction is distinct from the three BCAAs, and the sole location in which these reactions to occur is the mitochondrial matrix. The end result of valine is propionyl-CoA, which eventually enters the tricarboxylic acid (TCA) cycle and has the potential to be used in gluconeogenesis [29]. On the other hand, leucine produces acetyl-CoA, making it a ketogenic source. Isoleucine, however, produces both propionyl-CoA and acetyl-CoA [8,23,30,31].
Even though their metabolism involves multiple tissues, the levels of BCAAs in systemic circulation vary depending on dietary intake and general protein turnover [32]. Through their metabolic particularity of not being broken down in the liver as other amino acids, BCAAs are absorbed at the intestinal level and enter the bloodstream directly; therefore, the levels of BCAAs in the diet directly impact the levels in the blood [16,33]. Excess amino acids are stored in proteins in the liver and skeletal muscle, acting as major reservoirs for amino acids in mammals, and they are utilized during fasting periods [8,31]. After being released from proteins, BCAAs are metabolized in various peripheral tissues that express BCAT enzymes, such as skeletal muscle, as described above [8,31]. This leads to the generation of BCKAs, which are released into the bloodstream. Although the liver lacks BCAT expression, it plays a role in the metabolism of circulating BCKAs, using their carbon group for gluconeogenesis, ketogenesis, or fatty acid synthesis via the BCKDH complex [23,34].
Isotopic tracer studies that examined the uptake and breakdown of BCAAs in the whole body sought to uncover tissue-specific differences in BCAA metabolism and maximum oxidation occurring in skeletal muscle [25]. In addition, skeletal muscle tissues such as the liver and pancreas contribute the most to protein synthesis from BCAAs, and metabolic alteration in these tissues, as seen in neoplasms, may be responsible for consistent alterations in BCAA levels [25].
3. Branched-Chain Amino Acid-Regulated Signaling Pathways
Furthermore, apart from their roles as structural and metabolic components, BCAAs also have a functional role, acting as a promoter of various cellular signaling pathways. Among these pathways, the most extensively studied is the activation of the mTOR signaling pathway by leucine [26,35]. mTOR, a serine–threonine protein kinase, is a member of the PI3K-related kinase (PIKK) family and functions as the catalytic component for two separate complexes known as mTORC1 and mTORC2 [36,37,38]. Besides controlling the production of proteins by overseeing the messenger ribonucleic acid (mRNA) translation process, the intracellular mTOR molecule serves as a critical regulator in various essential cellular processes, including cell growth, cell division, autophagy, and glucose balance [39].
3.1. mTOR and Upstream Regulators
mTORC1 is composed of five elements: mTOR, the regulatory protein linked to mTOR (Raptor); the G-protein beta-subunit-like protein (GbL or mammalian lethal with Sec13 protein 8—mLST8); and two inhibitory components, pleckstrin (DEP) domain-containing mTOR-interacting protein (Deptor, Egl-10) and proline-rich AKT substrate 40 kDa (PRAS40) (Figure 1) [10,16,35,39]. Conversely, mTORC2 consists of mTOR, Deptor, mLST8, Rictor (rapamycin-insensitive companion of mTOR), protein observed with Rictor-1 (Protor-1), and mammalian stress-activated protein kinase-interacting protein 1 (mSIN1) [10,16,35,39].
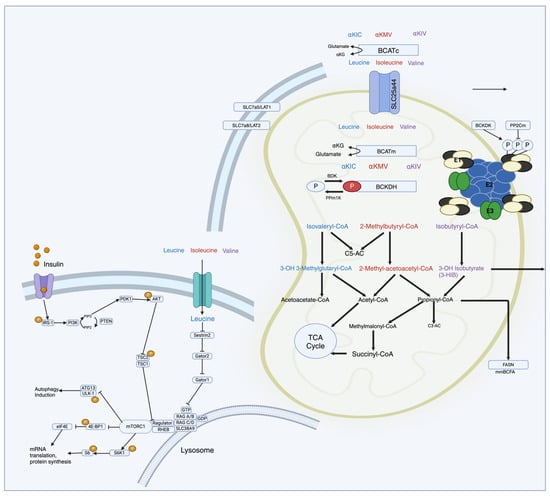
Figure 1.
Schematic representation of molecular pathways of BCAA metabolism in relation to mTOR regulation. Upon ingestion, BCAAs are transported into cells and undergo metabolism. The first step involves transamination, where BCAAs are converted into BCKAs by the enzyme BCAT. Subsequently, the BCKAs are further metabolized in mitochondria through a series of enzymatic reactions, ultimately generating intermediates like acetyl-CoA and succinyl-CoA, which enter central metabolic pathways. Leu acts as a direct activator of mTORC1 that stimulates protein synthesis by phosphorylating downstream effectors like S6 kinase 1 (S6K1) and eukaryotic translation initiation factor 4E-binding protein 1 (4E-BP1), leading to the enhanced translation of messenger ribonucleic acid (mRNA) into proteins involved in cell growth and proliferation.
Upon ingestion, BCAAs are transported into cells and undergo metabolism, primarily in skeletal muscle. The first step involves transamination, where BCAAs are converted into BCKAs by the enzyme BCAT. Subsequently, the BCKAs are further metabolized in mitochondria through a series of enzymatic reactions, ultimately generating intermediates like acetyl-CoA and succinyl-CoA, which enter central metabolic pathways [10,16,35,39]. The mTOR pathway is particularly sensitive to changes in BCAA levels, especially leucine. Leucine acts as a direct activator of mTORC1 (mTOR complex 1) by binding to and activating the protein kinase mTOR [11,26,32]. The activation of mTORC1 stimulates protein synthesis by phosphorylating downstream effectors like S6 kinase 1 (S6K1) and eukaryotic translation initiation factor 4E-binding protein 1 (4E-BP1), leading to the enhanced translation of mRNA into proteins involved in cell growth and proliferation [11,26,32].
The activation of the mTORC1 signaling pathway is initiated by a variety of factors, including cytokines, hormones, growth factors, and nutrients, further leading to the phosphorylation of translational regulators, including S6K, 4E-BP1 (eukaryotic translation initiation factor 4E (eIF4E)-binding protein 1), and SREBP1 [11,26,32]. This gamut of pathways further oversees the regulation of new lipid production, aerobic glycolysis, and the pentose phosphate metabolic pathway, also bringing equity among cell growth commensurate with nutrient availability [11,26,32]. Conversely, mTORC2 contributes to cell survival, cell proliferation, and the regulation of the cytoskeleton, being indirectly subjected to the control of the mTORC1 signaling pathway [11,13].
The tuberous sclerosis complex (TSC), composed of TSC1 (hamartin) and TSC2 (tuberin), is the primary mechanism for the upstream regulation of mTORC1 [40]. The stimulation of mTORC1 by growth factors like insulin and insulin-like growth factor 1 (IGF-1) triggers the activation of the phosphatidylinositol 3-kinase (PI3K)/protein kinase B (Akt) complex and extracellular-signal-regulated kinase 1 and 2 (ERK1/ERK2) [40]. These, in turn, phosphorylate and inhibit the activity of the TSC1–TSC2 complex, leading to the formation of a suppressor complex that acts as a guanine triphosphatase (GTPase)-activating protein (GAP) for the Ras homolog enriched in brain (Rheb), activating mTORC1 [40,41,42]. On the contrary, the AMP-activated protein kinase (AMPK) pathway activates TSC1/2, resulting in the inactivation of Rhed and mTORC1 inhibition with energy restriction. In particular, the AMPK pathway acts as an inhibitor, regardless of the TSC1/2 complex, due to direct Raptor phosphorylation [40,41]
Another pathway of mTORC1 activation that acts independently of TSC1/2 runs through Rag GTPases due to elevated levels of amino acids, particularly leucine. A complex between Rag GTPases and mTORC1 is formed in the presence of amino acids translocating from the cytoplasm to the lysosomal membranes to further activate by Rheb [43]. The direct interaction with the Raptor part of mTORC1 is facilitated by leucyl-tRNA synthetase (LRS)’s interaction with Rag, resulting in the assembly of the RagA/B–RagC/D complex in their GTP/GDP-loading states (RagA/B-GTP and RagC/D-GDP) [39,44]. This explains why low levels or the absence of amino acids promote mTORC1 inactivation secondary to Rag transitions into an inactive state [45]. This mTORC1-independent pathway activation, being primarily impacted by leucine levels compared to other amino acids, justifies why the reduction in leucine alone is just as effective as completely depriving the body of all amino acids and, conversely, why leucine alone is enough to facilitate the activation [14].
Two GAP complexes, GATOR1 and Folliculin (FLCN)–folliculin interacting protein 2 (FNIP2), are responsible for the downregulation of the Rag GTPases. GATOR1 acts on RagA and RagB, while FLCN-FNIP2 acts on RagC and RagD [14,44,46]. The GATOR2 complex, composed of Sestrin2 and other interacting proteins, negatively regulates GATOR1 [14,44,46]. Sestrin2, being a leucine sensor upstream of mTORC1, acts as a negative regulator, whereas LRS acts as a positive regulator of mTORC [14,44,46].
Although the mechanisms of mTORC1 activation are extensively studied, the mechanisms of mTORC2 activation still need to be better understood [14]. Recently issued hypotheses posit that its activation is induced by growth factors through the binding of mSin1 in response to PI3K being stimulated by growth factors like insulin [14]. When stimulated by amino acids, mTORC2 is involved in various cellular functions, such as adhesion, differentiation, proliferation, and apoptosis, functions mediated by the activation of Akt (Serine473), protein kinase C (PKC) alpha, and serum- and glucocorticoid-inducible kinase (SGK) [47].
3.2. Downstream Regulators
In addition to the above-described upstream regulators of mTORC, the downstream regulators appear to have greater involvement in insulin resistance [48].
The phosphorylation of the eukaryotic initiation factor (eIF) 4E-binding protein 1 (4E-BP1), an inhibitory protein, enhances the formation of the elF4F complex, which promotes mRNA translation [11,26,32]. Moreover, the activation of ribosomal S6K, also facilitated by insulin and amino acids, provides an additional trigger for translation initiation [32]. The simultaneous activation of mTOR and S6K1 negatively affects Akt by phosphorylating serine residues on IRS-1, disrupting the interaction between insulin and its receptor [2,47]. Consistently elevated BCAA and mTORC1 activation levels trigger a negative feedback loop of IRS-1, resulting in insulin resistance [2,47].
The impact of mTORC2 on metabolism, stress responses, and apoptosis is also mediated by phosphorylating several AGC kinases, such as Akt, serum- and glucocorticoid-induced protein kinase (SGK), protein kinase C-a (PKCa), and Rho1 GDP-GTP exchange protein-2 (Rom2), with Akt subsequently inhibiting TSC1/2 [49]. Furthermore, mTORC2 promotes the negative feedback of IRS-1 and prevents its inactive form from cytosol buildup [49].
PPARs, with their three subtypes (PPAR α, β/δ, γ), are nuclear receptors that play a significant role in regulating the metabolism of BCAAs in relation to mTOR in insulin resistance. The interaction between PPARs and BCAAs is complex and can impact various aspects of metabolic homeostasis [50,51,52]. PPARα is primarily involved in regulating fatty acid metabolism and energy homeostasis, being highly expressed in tissues such as the heart, liver, and skeletal muscle. The activation of PPARα promotes the uptake of fatty acids into cardiomyocytes and enhances mitochondrial fatty acid oxidation [50,51,52]. Studies have suggested that PPARα activation may indirectly influence BCAA metabolism through its effects on mitochondrial function and energy utilization, therefore increasing the risk of lipid peroxidation toxicity and the susceptibility of the heart to ischemia/reperfusion damage via the GCN2/ATF6 pathway [53]. PPARδ is predominantly expressed in adipose tissue and plays a key role in adipocyte differentiation and insulin sensitivity. While less studied in the context of BCAA metabolism compared to PPARα, in one study, PPARδ inhibition modulated cardiomyocyte proliferation, while the overexpression via the agonist GW5015116 reduced the proliferation following ischemic lesions and significantly reduced BCAA and secondary metabolite levels, alleviating insulin resistance in the diabetic milieu [50,51].
Significantly, these modifications replicate the first metabolic shifts associated with systemic insulin resistance [52]. It is unclear whether elevated levels of fatty acid oxidation are preserved or reduced in subsequent stages of diabetic cardiomyopathy, as happens in the ischemic heart [52].
4. Branched-Chain Amino Acid in Cardiovascular Disease
Being one of the vital organs requiring major energy consumption on a daily basis, when subjected to pathological energetic states such as pressure overload, hypertrophy, and ischemia, a metabolic shift must occur to enable adaptation to different physiological and nutritional circumstances [54].
The majority of the energy is normally produced through fatty acid oxidation in the mitochondria, with a return to the fetal pattern of cardiac development in a pathological state, in which anaerobic glycolytic metabolism, with its high use of ketones and lactate and a decrease in fatty acid use, is characteristic [50,55]. This alternative adaptive mechanism is only beneficial in the short-term, extended use of glucose, causing a deficiency in ATP production commonly observed in various cardiac diseases [37,56,57,58]. ATP generation through the tricarboxylic acid (TCA) cycle in the mitochondria after fatty acid breakdown, together with the electrons generated and transferred through the electron transport chain (ETC), leads to the creation of a proton gradient and energy production [50,59,60]. This explains why fatty acids are the primary source of energy for the adult heart and why glucose is a secondary energy source [50,59,60].
The cardiac muscle is only a minor site of BCAA oxidation in terms of mass compared with other tissues like skeletal muscle and fat. Therefore, impairments in non-cardiac BCAA oxidation are more likely to be responsible for the increased levels of circulating BCAAs in CVD [60,61]. A potential association could be explained by a decrease in the breakdown of BCAAs due to obesity or insulin resistance cases in which the metabolism exhibits inflexible alterations characterized by a resistance to shift from oxidizing fatty acids to oxidizing glucose, subsequently leading to an increase in triglyceride levels, particularly in the heart tissue [26,62]. Dysfunctional BCAA breakdown and a deficiency in the enzymes responsible for their metabolism lead to increased levels in both the heart tissue and liver, with the subsequent mTORC-1 activation leading to cardiac hypertrophy [16,63]. Additional proof that alterations in the metabolism of BCAAs are connected to CVD was observed in the Prevención con Dieta Mediterránea (PREDIMED) study, a case–cohort study involving 226 newly diagnosed CVD cases and 781 individuals without CVD which concluded that elevated levels of BCAAs were linked to a higher risk of CVDs, particularly stroke (Table 1) [64]. However, the significance of BCAA metabolism in relation to CVD is still not well understood [65].

Table 1.
Summary of references for studies on BCAAs as potential biomarkers.
Understanding the mechanisms involved in the pathogenesis of CVD and the role of BCAA may provide valuable insights for developing new strategies, especially when they coexist with type 2 diabetes mellitus (T2DM) encompassing the cardiometabolic disease [34,79,80,81].
4.1. Heart Failure
A decrease in energy production from the breakdown of fatty acids at the expense of increased glucose utilization marks the defining metabolic substrate for heart failure. This metabolic inflexibility contributes to the progression of both systolic and diastolic dysfunctions [50,82].
In order to gain a deeper understanding of the role of amino acid metabolism in heart failure, particular attention has been given to BCAAs and their different utilization patterns [50,82]. The changes observed in failing hearts mainly encompass the accumulation of BCAAs secondary to multiple probable mechanisms, such as the downregulation of encoding genes from the catabolic pathway, mitochondrial dysfunction, mTOR activation, and glucose oxidation impairment [36,60,83,84].
Several animal models of heart failure were studied in correlation with BCAA levels, but the results were conflicting, giving scope for further research in the future. In rodents with HF induced by myocardial infarction (MI), BCAA systemic levels increase progressively, while both cardiac BCAA levels and catabolic enzyme abundance decrease secondary to a decreased metabolism over time [85,86].
mTOR activation and reactive oxygen species (ROS) lead to the accumulation of BCAAs and their catabolic enzymes, which is a hallmark of cardiac dysfunction and remodeling [87]. While BCKDH subunits are downregulated in heart failure patients, BCAAs accumulate in the myocardium, but these do not always correspond to plasma concentrations [16,60,88,89]. Consequently, therapeutic methods for promoting BCAA breakdown in myocardial tissue have been tested, but the depletion of plasma levels has also led to exacerbations of cardiac dysfunction, as seen in a murine model of left ventricular failure [63]. Another model of pressure overload-induced HF in transaortic constriction (TAC) mice resulted in increased cardiac BCAA catabolic intermediates, which was corroborated by subsequent verification from a human cohort with HF with reduced ejection fraction (HfrEF) that displayed increased levels of a-keto-b-methylvalerate (the BCKA of isoleucine) [90,91]. Another comparative study on TAC- and MI-induced HF revealed changes in cardiac BCAA levels with no alteration regarding plasmatic values but with a new insight into the correlation with cardiac insulin resistance [16,92].
In 2022, a novel ‘chemogenetic’ mouse model of dilated cardiomyopathy-induced heart failure (HF) was introduced utilizing hydrogen peroxide in cardiomyocytes [93]. After feeding animals with the d-alanine enzyme, oxidative stress was induced [93]. BCAA catabolic enzymes were downregulated with increased cardiac levels but without a change in the plasma concentrations of the BCAAs [93]. In a different study based on dilated cardiomyopathy but in human patients, higher BCAA cardiac levels outlined a deficiency in BCKDH phosphorylation and Ppm1k and BCAT2 activity [66]. These results show model-dependent differences in systemic circulating BCAA levels and point to higher cardiac BCAA concentrations and impaired cardiac BCAA metabolism as consistent hallmarks of cardiac dysfunction [58,94,95].
To differentiate the extent to which intrinsic disturbances in cardiac amino acid metabolism versus fluctuating plasma amino acid levels influence the physiopathology of HF, further studies were carried out employing dietary BCAA supplementation, pharmacological interventions targeting BCAA metabolism, and genetic animal models [17,85].
A novel mice model lacking Ppm1k (or Pp2cm), a gene encoding a protein phosphatase crucial for BCAA catabolism, exhibits heightened levels of BCAAs and related metabolites in both plasma and cardiac tissue, alongside a mild decline in cardiac function over time [28,50]. Notably, when subjected to cardiac stress via transaortic constriction (TAC), Ppm1k-deficient mice display accelerated HF progression [90]. In Ppm1k-deficient mice, the allosteric inhibitor of BDK, known as 3,6 dichlorobenzo[b]thiophene-2-carboxylic acid (BT2), was used in an attempt to uncover the cardiac-specific metabolic BCAA dysfunction [56]. The administration of BT2 for one week resulted in the dephosphorylation and maximal activation of BCKDH in multiple sites, including the heart, skeletal muscle, kidneys, and liver of mice and rats, leading to normalizing plasma BCAA concentrations [17,90,96,97]. A fresh perspective was observed regarding Pp2cm and its interplay with the microbiome metabolite butyrate in the early stages of obesity-related heart failure with preserved ejection fraction (HfpEF) [98]. It has been noted that a decrease in the expression of the Ppm1k gene, responsible for encoding PP2Cm, leads to elevated levels of inactive p-BCKDH, prompting the necessity for additional investigation [14,98,99,100].
Furthermore, BT2 administration in TAC-induced HF delayed progression in mice. These findings suggest that impaired cardiac BCAA catabolism contributes to HF pathogenesis and progression, with BT2 treatment being a possible potent amino acid catabolic restaurateur [66,101,102]. However, the precise impairment in cardiac-specific BCAA dysfunction remains unclear, as these models impact systemic BCAA metabolism, altering both circulating and cardiac BCAA concentrations (Table 2) [56,84,103].
Table 2.
Summary of references for relevant studies related to BCAAs and heart failure.
This downregulation is consistent with the disruption of BCAA breakdown. Enhancing BCAA catabolism could be a therapeutic strategy for improving cardiac insulin sensitivity and limiting pathological remodeling after a myocardial infarction [87,108].
After attempting to determine the potential mechanisms underlying fluctuations in plasma and cardiac amino acid concentrations in HF without conclusive results, attention shifted to unraveling the intricate interplay between BCAA delivery to the heart and intrinsic cardiac metabolism in HF pathogenesis regarding mTOR activation [109,110].
Oral supplementation with BCAAs in mice after MI exacerbated cardiac dysfunction by the increased phosphorylation of mTOR, quantified by measuring its hallmark downstream targets, such as ribosomal protein S6 kinase-b1 (also known as P70S6K) and insulin receptor substrate [111], which could be prevented by concurrent treatment with the mTOR inhibitor rapamycin [39,86,112]. Elevated cardiac BCAA concentrations are also linked to mTOR activation in cardiac tissue from humans with dilated cardiomyopathy, associated with the reduced phosphorylation of enzymes involved in insulin signaling, potentially contributing to cardiac insulin resistance via AKT and glycogen synthase kinase 3 [66,99].
The metabolic circadian pattern to which mTOR is subjected could not establish a causal relationship that would explain the daily metabolic fluctuations of BCAAs during heart failure but drew attention to KLF15 [113], a major regulator of the circadian metabolism of BCAAs [114,115]. KLF15, a member of the KLF family, is responsible for activating the mitochondrial branched-chain aminotransferase (BCATm), which is the first step in the breakdown of BCAA [115]. Furthermore, KLF15 can influence the hypertrophic signaling of mTOR in skeletal muscle, causing cardiac insulin resistance and hypertrophy [115]. However, the activity of KLF15 is reduced by the activation of p38 mitogen-activated protein kinase (p38 MAPK), and it is also downregulated in both murine models of heart failure (HF) and patients with cardiomyopathy, leading to BCKA intramyocardial accumulation and BCAA catabolic gene suppression [14,111]. The high concentrations of BCAAs impact mitochondrial ATP production and inhibit pyruvate and 2-KG dehydrogenases, creating a proarrhythmic substrate secondary to impaired contractility, as highlighted in a mice model lacking KLF15 [14]. The information surrounding the regulatory molecules of KLF15 is limited, and previous research has indicated that the activation of p38 MAPK through TGFβ-activated kinase 1 (TAK1) has an inhibitory role by stimulating the expression of transforming growth factor beta (TGFβ) in myocytes, contributing to the impaired BCAA catabolism observed in human DCM hearts (Figure 2) [5,66,105].
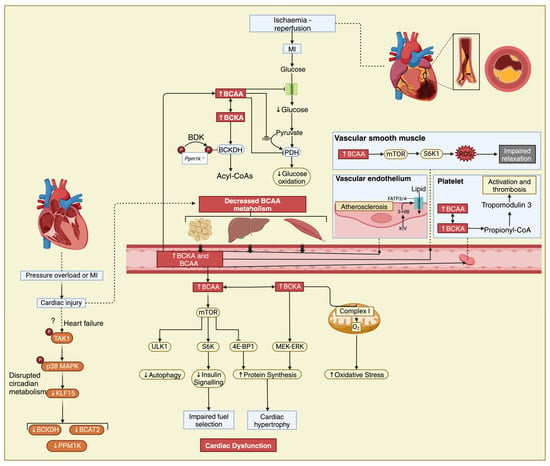
Figure 2.
The interplay between BCAAs, heart failure, and atherosclerosis. Elevated circulating levels of BCAAs have been observed in individuals with heart failure and have been linked to insulin resistance, which is a common feature of both heart failure and atherosclerosis. In the setting of myocardial infarction, disrupted BCAA metabolism can impact mitochondrial function in cardiac muscle cells, potentially exacerbating tissue damage and impairing recovery post-MI. ?—Cardiac injury also potentially impairs BCAA metabolism via a mechanism involving transforming growth factor-ß-activated kinase 1 (TAK1) and p38 mitogen-activated protein kinase (MAPK) signalling and downregulation of KLF15 as seen in heart failure, resulting in increased circulating BCAA and BCKA levels. Additionally, BCAAs may play a role in promoting inflammation and oxidative stress, contributing to the progression of atherosclerosis. Unv-51-like kinase 1, ULK1; ribosomal protein S6 kinase, S6K; 4E-binding protein 1, 4E-BP1; mitogen-activated protein kinase ERK kinase/extracellular-signal-regulated kinase, MEK-ERK.
There is still more research to be conducted on the topic of myocardial insulin resistance mediated by BCAAs because no clear mechanism has been found. Although research on pancreatic islet cells from mice lacking BCAT2 suggested that BCAAs and their derivatives, BCKAs, would have similar effects, they showed that KIC stimulation of insulin production depends on the conversion of KIC and glutamate to leucine and α-ketoglutarate [111]. Furthermore, mice deficient in BCAT2 showed enhanced insulin resistance and suppressed glucose oxidation in response to BCKA stimulation [111].
4.2. Cardiometabolic Disease
Diabetes has emerged as a widespread health crisis, placing a significant strain on healthcare systems globally due to its substantial impact on morbidity and mortality rates. BCAA and their related metabolites are now recognized as highly reliable biomarkers for various cardiometabolic diseases and associated conditions, such as obesity, insulin resistance, and T2D [4]. Several prospective studies have consistently shown a link between circulating levels of BCAAs and the onset of T2DM across different age groups [116].
Current research suggests that BCAAs hold promise in predicting the development of diabetes, with alterations in plasma-free amino acid (AA) profiles observed prior to the onset of T2DM and significantly correlated with future diabetes diagnoses [117]. These changes in plasma-free AA profiles levels likely stem from metabolic shifts triggered by the early stages of diabetes progression. A study analyzing metabolite profiles in 2422 normoglycemic individuals revealed that 201 participants developed diabetes over a 12-year period, highlighting robust associations between the three aforementioned BCAAs, two aromatic AAs (tyrosine and phenylalanine), and diabetic outcomes [117]. These results underscore the potential value of AA profiles in enhancing future risk assessment tools for diabetes.
For instance, a large cohort study involving over 50,000 individuals from the UK Biobank showed that adding plasma nuclear magnetic resonance-measured metabolites, including BCAAs, improved the prediction of T2DM risk when combined with basic clinical parameters [118]. Another prospective cohort study in the Chinese population demonstrated enhanced risk estimation for T2DM prediction by including specific metabolites over an average follow-up period of eight years [119].
A significant study utilizing the Framingham cohort revealed that initial BCAA levels were identified as robust predictors of developing diabetes, with the patients included being three times more likely to develop diabetes within a 12-year monitoring period [77]. This correlation has been consistently verified across various cohorts; however, most studies have primarily examined the risk of developing diabetes based on single-point measurements of circulating BCAA levels [77]. One example is a group of men from South Africa from the SABRE (Southall and Brent Revisited) Study, wherein elevated circulating BCAA levels were linked to around a 40% higher relative risk of incident diabetes [78]. Recently, a longitudinal study in a diverse cohort of young Black and White adults over more than 35 years old indicated that circulating BCAA levels remain stable over a period of 28 years for most young adults, highlighting the necessity of successive metabolomic assessments to identify specific subgroups exhibiting an elevated risk of developing diabetes later in life [120].
Additionally, an updated meta-analysis incorporating data from 61 prospective cohort reports on T2DM risk underscored strong associations not only with BCAAs but also other amino acids and lipid metabolites [121]. This analysis evaluated metabolite concentrations in various biological samples, such as plasma, serum, and urine samples, using high-throughput metabolomic platforms [121].
Also elevated levels of 3-HIB (3- hydroxyisobutyrate) in plasma serve as an indicator for future T2DM risk and may play a crucial role in regulating metabolic flexibility in the heart and muscles [12,117]. While diabetic conditions lead to increased circulating levels of branched-chain amino acids (BCAA), ketones, glucose, and fatty acids, their oxidation rates do not proportionately increase this imbalance, contributing to cardiac dysfunction among diabetic individuals [12].
A meta-analysis of 19 prospective studies revealed an association between branched-chain amino acids (BCAAs) and aromatic AAs with prediabetes and T2DM, which are mechanistically related through their potential impact on insulin resistance [122]. The activation of mTOR/p70S6 kinase by increased BCAAs, particularly leucine, has been proposed as the most prevalent mechanism that inhibits insulin signaling by phosphorylating the IRS-1 [32]. However, it remains unclear whether BCAA-induced mTORC1 activation is sufficient to trigger insulin resistance or if further research is needed to understand their roles fully [32].
Recently, attention has been directed towards trimethylamine-N-oxide (TMAO), a metabolite produced as a result of gut microbiota fermentation that has garnered attention due to its suspected involvement in the progression of ischemic heart disease [123], complications associated with T2DM, and premature mortality in the general population [124,125]. While some studies have linked elevated plasma levels of TMAO with adverse cardiovascular outcomes, findings among individuals with underlying health conditions vary [126]. Notably, the association between TMAO and cardiovascular mortality in individuals with T2DM seems to be particularly significant among high-risk populations [126]. Recent research has indicated that BCAA concentrations are also influenced by the microbiota, therefore raising the question of how circulating BCAA concentrations correlate with TMAO levels in diabetic milieu [126,127].
Furthermore, research suggests that excessive BCAA catabolism may play a role in the pathway from obesity and insulin resistance to T2DM by influencing inflammation and genetically predicted insulin resistance [128].
4.3. Hypertension
The prevalence of high blood pressure and the negative effects it has on health, as well as the associated economic burdens, are well known risk factors for CVDs [31,129,130,131]. While studies on heart failure and coronary artery disease are more frequent and several pathophysiological mechanisms have been identified regarding hypertension, the results are inconclusive, pointing only to an association between BCAAs and this condition [31,129,130,131].
Since Akt/mTORC1 was highlighted in connection to heart failure, researchers aimed to uncover its role in hypertension [132,133]. Potential interplay between BCAAs and the increased activation of angiotensin-converting enzymes through binding amino acid residues has been proposed [132,133]. A study published by Chen et al. reinforced this supposition by uncovering elevated levels of mTOR in hypertensive rats with cardiac hypertrophy [134]. Consequently, the mTOR/S6K1 pathway was found to be activated by angiotensin II (ANG II), further promoting oxidative stress and decreased levels of nitric oxide (NO) [134].
While the particular metabolic pathways remain under scrutiny, a few studies have tried to underline a correlation between plasma BCAA levels and dietary intake with hypertension [135]. Although attempts have sought to determine precisely which amino acid is linked to hypertension risk, studies have yielded conflicting results [135]. For example, in an Iranian study involving 4288 non-hypertensive patients, dietary Val intake turned out to have a stronger correlation with hypertension over a monitoring period of three years [136,137], while Mahbub et al. discovered the involvement of Leu in Japanese participants [132].
When measuring 24 h urine levels of BCAAs in older hypertensive patients, isoleucine was linked with diastolic blood pressure, while valine was found to be negatively correlated with both systolic and diastolic blood pressure [138]. However, Siomkajło et al. [131] found significant positive associations between plasma levels and both systolic and diastolic blood pressure findings reinforced by a large-scale prospective cohort study on middle-aged men and women conducted by Flores-Guerrero et al. [139]. On the other hand, Liu et al. found no particular association between a specific BCAA with hypertensive risk, thus raising the hypothesis of population-based variability and genetic background [135].
The PREVEND study, enrolling over 4100 participants over nine years of follow-up, predicted the impact of incident hypertension on BCAA plasma levels independent of traditional risk factors [139]. Similar results were observed in cross-sectional analyses of two Asian cohorts (81 out of 8589 Japanese individuals and 82 out of 472 Chinese individuals) [140]. On the other hand, a lower incidence of arterial stiffness and incident hypertension was linked to a higher dietary intake of BCAAs in research involving 1898 female twins from the TwinsUK registry, with the caveat that plasma levels were also influenced by intrinsic BCAA metabolic impairment, not only by dietary consumption [130].
The precise mechanisms underlying the connection between each BCAA and hypertension have yet to be clarified. Regarding leucine, a partial mechanism responsible for a stronger association with high blood pressure was proposed via the inhibition of NO synthesis from L-arginine and alanine generation secondary to glutamate production after the transamination process [132,136].
While the aforementioned studies found discrepant results, it has been consistently shown that BCAA concentrations are positively associated with high blood pressure, with the reserve that other nutrients or yet unknown pathways may counterbalance their effects (Table 3) [141].
Table 3.
Summary of references on relevant studies related to BCAAs and hypertension.
4.4. Atherosclerosis and Coronary Artery Disease
Peripheral artery disease, ischemic stroke, and atherosclerotic coronary heart disease are only a few of the major cardiovascular events that are caused by atherosclerosis (AS), and the significantly increase the morbidity and mortality rates associated with CVD [144,145]. Besides the traditional risk factors extensively researched in AS pathogenesis, BCAAs have also sparked interest; however, the exact mechanisms are still unknown [1,146,147].
Two of the initial steps in the development of atherosclerotic plaque are represented by lipid and cholesterol deposition at the endothelial level, followed by inflammatory cell infiltration, which facilitates platelet activation and thrombus formation after a plaque rupture [148]. A comparative study between human platelets and platelets from Ppm1k−/− mice showed that BCAA consumption in human volunteers led to platelet activation and aggregation because of collagen [149]. On the other hand, platelets from Ppm1k−/− mice exhibited lower susceptibility to activation and thrombus formation acting indirectly upon the thrombotic process via intermediate catabolic products, especially α-ketoisovalerate [149]. Another aspect worth mentioning is that Ppm1k−/− mice with increased BCAA plasma levels had an inauspicious response to ischemia–reperfusion injury at the expense of downregulated glucose oxidation [1,28,150].
Zhao et al. discovered that increased BCAA levels lead to proinflammatory macrophage activation and AS progression [151]. Mitochondrial H2O2 (mtH2O2) levels observed in activated macrophages induced the formation of high-mobility group box 1 protein (HMGB1) and the activation of the Toll-like receptor 4 (TLR4)/nuclear factor kappa-light-chain-enhancer of activate B cells (NF-κB) pathway, which is modulated by BCAA catabolism [151]. Notably, recent research by Zhenyukh et al. has suggested that elevated levels of BCAAs stimulate the PI3K-Akt/mTORC1 signaling pathway, triggering the formation of reactive oxygen species (ROS) and proinflammatory proteins [38,152,153]. Nevertheless, the underlying molecular pathways are yet to be understood, and it is unclear whether an increased expression of BCAAs is linked to macrophage-mediated atherosclerotic inflammation [35,38,154].
mTOR activation was also linked to reduced endothelial relaxation secondary to leucine levels in mouse aortic rings, in which an increased phosphorylation of 40S ribosomal protein S6 and overexpression of ribosomal protein S6 kinase (S6K) were observed (Figure 2) [155].
Concerning coronary artery disease, BCAAs have highlighted themselves as potential biomarkers and a severity predictor independently or in association with T2DM [144,156,157]. Being one of the primary causes of cardiovascular death, the coexistence with T2DM increases the severity of lesions, worsening the prognosis and reducing patient survival due to severe complications [1].
Baseline plasma levels of BCAAs and the risk of acute HF complication in patients with ST segment elevation MI (STEMI) necessitating percutaneous coronary intervention were first observed by Shah et al. in 2012 [158] and were associated with a worse prognosis [86,109]. Later, the PREDIMED study concluded that an elevated risk of myocardial infarction was associated with leucine and isoleucine levels [64], validating earlier findings from a European cohort in which isoleucine was linked to atherosclerosis progression (Table 4) [70,159].
Table 4.
Summary of references for relevant studies related to BCAAs, atherosclerosis, and coronary artery disease.
A Women’s Health Study that included over 27,000 participants with 19 years of follow-up reinforced the predictive role of BCAA levels in identifying patients that developed CVD in association with T2DM [69,164]. A potential limitation was based on race since the analysis from the Diabetes Heart Study showed a difference between European Americans and African Americans in BCAA levels, which showed a correlation with coronary artery calcium [165]. Another particularity in African Americans was observed in the Jackson Heart Study, where leucine was associated with a lower incidence of coronary artery disease [166].
5. Discussion
The molecular ties between BCAAs and chronic CVDs represent a complex and evolving area of research with important implications for disease pathogenesis and therapeutic strategies. Throughout this exploration, several key insights and conclusions can be drawn.
The balance of free circulating BCAAs is influenced by both their intake (from diet and proteolysis) and their utilization (for protein synthesis and catabolism for energy). As essential amino acids that can be obtained only from food sources, their homeostasis must be maintained by catabolism [1,8]. The first two steps of this process are the same for all three BCAAs: BCAT catalyzes reversible transamination, which forms BCKAs. The irreversible oxidative decarboxylation of BCKAs, which is subsequently catalyzed by the BCKDH complex, represents the rate-limiting step in the entire BCAA catabolic complex process (Figure 1) [1,8].
Elevated circulating BCAA levels are commonly observed in individuals with chronic CVDs, including heart failure, atherosclerosis, and hypertension. This dysregulation of BCAA metabolism and accumulation in the bloodstream are closely linked to insulin resistance, dyslipidemia, oxidative stress, and endothelial dysfunction, which are critical factors in CVD development and progression [65,70]. Although they are crucial in numerous essential processes, their deleterious effect is secondary to imbalances occurring in one of the phases of the complex metabolism, thus altering their plasmatic level secondary to local cardiac-level fluctuations in different pathologic states [65,70]. The molecular links between BCAAs and CVD can be understood through several mechanisms.
Elevated BCAA levels are associated with insulin resistance, a condition that often precedes type 2 diabetes [48]. Insulin resistance itself is a significant risk factor for cardiovascular diseases [48]. BCAAs may impair insulin signaling pathways, particularly through the mTOR pathway and serine phosphorylation of insulin receptor substrate (IRS) [11,26,32]. Chronic activation of the mTOR pathway desensitizes cells to insulin, contributing to insulin resistance, especially secondary to leucine [2,47]. BCAAs can also promote the phosphorylation of IRS on serine residues rather than tyrosine residues, disrupting the insulin signaling cascade and further sustaining insulin resistance [11,26,32]. Dysregulated mTOR signaling, influenced by BCAA availability, can contribute to adverse cardiac remodeling, fibrosis, and hypertrophy, ultimately impacting cardiovascular function. BCAAs interact with transcriptional regulators like SREBPs and PPARs, affecting lipid metabolism and inflammatory processes in the cardiovascular system [1,50,51,52]. Inflammatory pathways such as the NF-κB pathway, which is known to contribute to atherosclerosis, are upregulated by elevated levels of BCAAs [151]. This can also lead to oxidative stress by altering mitochondrial function and promoting the production of reactive oxygen species (ROS). Oxidative stress damages the vascular endothelium, contributing to atherosclerosis and other cardiovascular issues [38,152,153].
Regarding dyslipidemia, a known risk factor for cardiovascular diseases, BCAAs modulate lipid metabolism by affecting the expression of genes involved in lipid synthesis and degradation, leading to increased levels of circulating free fatty acids, triglycerides, and low-density lipoprotein (LDL) cholesterol, promoting atherosclerosis and coronary artery disease [167].
Besides their association with the traditional cardiovascular risk factors, gut microbiota play a significant role in metabolizing BCAAs [126]. Dysbiosis, or an imbalance in gut microbiota, can lead to altered BCAA metabolism and increased levels of harmful metabolites such as TMAO, which has been linked to atherosclerosis and CVDs [126].
In heart failure, there is a decrease in the central circadian regulator of BCAA metabolism, KLF15. This downregulation is mediated by the TAK1 and p38 MAPK signaling pathway, resulting in a reduced expression of BCAA metabolic enzymes such as mitochondrial BCAT2, BCKDH, and Ppm1k [5,66,105]. Consequently, there is an accumulation of BCAAs and BCKAs at cardiac level, while cardiomyocyte injury may also disrupt BCAA metabolism in peripheral tissues, leading to elevated circulating levels of BCAAs and BCKAs [111]. As already mentioned, Leu plays a role in activating mTOR, inhibiting autophagy through ULK1, inducing insulin resistance via IRS-1 phosphorylation by S6K, and promoting protein synthesis through 4E-BP1 phosphorylation. BCKAs contribute to increased 4E-BP1 phosphorylation and activate the MEK-ERK pathway [11,26,32]. Exposure to BCKAs impairs mitochondrial complex I function in the heart, causing oxidative stress due to superoxide generation [11,26,32].
During ischemia–reperfusion injury, deleting the Ppm1k gene in mice leads to elevated levels of both circulating and cardiac BCAAs and BCKAs [1,28,150]. This increase hinders glucose transport and oxidation by reducing the O-linked N-acetylglucosaminylation of pyruvate dehydrogenase (PDH), exacerbating ischemic damage [1,28,150]. Through an mTOR-dependent mechanism involving ROS production, BCAAs interfere with vascular relaxation [155]. Platelet oxidation of BCAAs promotes thrombosis by propionylating tropomodulin 3, a potent platelet activator [35,38,154].
In models of obesity and insulin resistance in animals, accumulated BCAAs hinder fatty acid oxidation while boosting triglyceride storage. Elevated levels of BCKAs seen in obesity and type 2 diabetes inhibit AKT and PDH, affecting energy substrate utilization, but the impact of these alterations on cardiac function remains uncertain at present [28,150]. It is also unclear whether changes within the cardiac muscle trigger shifts in BCAA metabolism within other tissues like the liver, skeletal muscle, or adipose tissue, and this is a question that persists regarding TMAO also [123].
Numerous human epidemiological studies have indicated a correlation between circulating BCAAs and the onset of cardiometabolic diseases [120]. However, most of these studies relied on single-time measurements of BCAAs in populations of European ancestry, and fasting levels of BCAAs exhibit significant variations between men and women, suggesting potential sex-specific differences in how BCAAs relate to the risk of cardiometabolic diseases [77]. This complexity underscores the importance of quantifying the associated risks more effectively by utilizing repeated measures and exploring these connections across diverse cohorts consisting of both sexes and different races [126]. Adopting such an approach will not only help in assessing the dynamic nature of circulating BCAAs but also in identifying the factors influencing the relationships between BCAAs and specific cardiometabolic conditions at molecular levels.
6. Conclusions
In summary, the role of BCAAs in chronic CVD underscores their multifaceted involvement in metabolic regulation, cellular signaling, and disease pathophysiology [168]. Despite significant advances, many questions remain regarding the precise mechanisms underlying BCAA-mediated effects on cardiovascular health.
By unraveling the complexities of BCAA metabolism and its interactions with cardiovascular health, researchers can identify novel therapeutic targets and dietary interventions aimed at improving outcomes for individuals with chronic cardiovascular conditions. However, it is essential to note that many additional factors, such as overall diet, genetics, and lifestyle, influence individual risk profile. Continued investigation into the role of BCAAs in CVDs holds promise for advancing our understanding of disease mechanisms and developing more effective strategies for prevention and treatment.
Author Contributions
Conceptualization, D.M.T. and E.V.; methodology, C.F.C.; software, D.V.S.; validation, A.O., M.F. and D.M.T.; formal analysis, C.C.T.; investigation, L.L.H.; resources, M.C.; data curation, L.G.B.; writing—original draft preparation, D.M.T., M.F. and E.V.; writing—review and editing, D.E.F. and D.V.S.; visualization, C.F.C.; supervision, A.O.; project administration, D.M.T.; funding acquisition, L.L.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Jiang, W.; Lu, K.; Zhuang, Z.; Wang, X.; Tang, X.; Huang, T.; Gao, P.; Wang, Y.; Du, J. Mendelian Randomization Analysis Provides Insights into the Pathogenesis of Serum Levels of Branched-Chain Amino Acids in Cardiovascular Disease. Metabolites 2023, 13, 403. [Google Scholar] [CrossRef]
- Cuomo, P.; Capparelli, R.; Iannelli, A.; Iannelli, D. Role of Branched-Chain Amino Acid Metabolism in Type 2 Diabetes, Obesity, Cardiovascular Disease and Non-Alcoholic Fatty Liver Disease. Int. J. Mol. Sci. 2022, 23, 4325. [Google Scholar] [CrossRef] [PubMed]
- Kubacka, J.; Cembrowska, P.; Sypniewska, G.; Stefanska, A. The Association between Branched-Chain Amino Acids (BCAAs) and Cardiometabolic Risk Factors in Middle-Aged Caucasian Women Stratified According to Glycemic Status. Nutrients 2021, 13, 3307. [Google Scholar] [CrossRef]
- White, P.J.; McGarrah, R.W.; Herman, M.A.; Bain, J.R.; Shah, S.H.; Newgard, C.B. Insulin Action, Type 2 Diabetes, and Branched-Chain Amino Acids: A Two-Way Street. Mol. Metab. 2021, 52, 101261. [Google Scholar] [CrossRef]
- Walejko, J.M.; Christopher, B.A.; Crown, S.B.; Zhang, G.-F.; Pickar-Oliver, A.; Yoneshiro, T.; Foster, M.W.; Page, S.; Van Vliet, S.; Ilkayeva, O.; et al. Branched-Chain α-Ketoacids Are Preferentially Reaminated and Activate Protein Synthesis in the Heart. Nat. Commun. 2021, 12, 1680. [Google Scholar] [CrossRef] [PubMed]
- Alfaqih, M.A.; Melhem, N.Y.; Khabour, O.F.; Al-Dwairi, A.; Elsalem, L.; Alsaqer, T.G.; Allouh, M.Z. Normalization of Vitamin D Serum Levels in Patients with Type Two Diabetes Mellitus Reduces Levels of Branched Chain Amino Acids. Medicina 2022, 58, 1267. [Google Scholar] [CrossRef] [PubMed]
- Okekunle, A.P.; Wu, X.; Duan, W.; Feng, R.; Li, Y.; Sun, C. Dietary Intakes of Branched-Chained Amino Acid and Risk for Type 2 Diabetes in Adults: The Harbin Cohort Study on Diet, Nutrition and Chronic Non-Communicable Diseases Study. Can. J. Diabetes 2018, 42, 484–492.e7. [Google Scholar] [CrossRef]
- Bonvini, A.; Coqueiro, A.Y.; Tirapegui, J.; Calder, P.C.; Rogero, M.M. Immunomodulatory Role of Branched-Chain Amino Acids. Nutr. Rev. 2018, 76, 840–856. [Google Scholar] [CrossRef]
- Neinast, M.D.; Jang, C.; Hui, S.; Murashige, D.S.; Chu, Q.; Morscher, R.J.; Li, X.; Zhan, L.; White, E.; Anthony, T.G.; et al. Quantitative Analysis of the Whole-Body Metabolic Fate of Branched-Chain Amino Acids. Cell Metab. 2019, 29, 417–429.e4. [Google Scholar] [CrossRef]
- Bar-Tana, J. Type 2 Diabetes—Unmet Need, Unresolved Pathogenesis, mTORC1-Centric Paradigm. Rev. Endocr. Metab. Disord. 2020, 21, 613–629. [Google Scholar] [CrossRef]
- Cheon, S.Y.; Cho, K. Lipid Metabolism, Inflammation, and Foam Cell Formation in Health and Metabolic Disorders: Targeting mTORC1. J. Mol. Med. 2021, 99, 1497–1509. [Google Scholar] [CrossRef] [PubMed]
- Nie, C.; He, T.; Zhang, W.; Zhang, G.; Ma, X. Branched Chain Amino Acids: Beyond Nutrition Metabolism. Int. J. Mol. Sci. 2018, 19, 954. [Google Scholar] [CrossRef]
- Rohini, A.; Agrawal, N.; Kumar, H.; Kumar, V. Emerging Role of Branched Chain Amino Acids in Metabolic Disorders: A Mechanistic Review. PharmaNutrition 2018, 6, 47–54. [Google Scholar] [CrossRef]
- Dimou, A.; Tsimihodimos, V.; Bairaktari, E. The Critical Role of the Branched Chain Amino Acids (BCAAs) Catabolism-Regulating Enzymes, Branched-Chain Aminotransferase (BCAT) and Branched-Chain α-Keto Acid Dehydrogenase (BCKD), in Human Pathophysiology. Int. J. Mol. Sci. 2022, 23, 4022. [Google Scholar] [CrossRef]
- Le Couteur, D.G.; Solon-Biet, S.M.; Cogger, V.C.; Ribeiro, R.; De Cabo, R.; Raubenheimer, D.; Cooney, G.J.; Simpson, S.J. Branched Chain Amino Acids, Aging and Age-Related Health. Ageing Res. Rev. 2020, 64, 101198. [Google Scholar] [CrossRef] [PubMed]
- Neinast, M.; Murashige, D.; Arany, Z. Branched Chain Amino Acids. Annu. Rev. Physiol. 2018, 81, 139–164. [Google Scholar] [CrossRef]
- Bai, X.; Long, X.; Song, F.; Chen, B.; Sheng, C.; Tang, C.; Li, L.; Zhang, J.; Zhang, R.; Zhang, J.; et al. High Doses of Rosuvastatin Induce Impaired Branched-chain Amino Acid Catabolism and Lead to Insulin Resistance. Exp. Physiol. 2023, 108, 961–974. [Google Scholar] [CrossRef]
- Bloomgarden, Z. Diabetes and Branched-Chain Amino Acids: What Is the Link? J. Diabetes 2018, 10, 350–352. [Google Scholar] [CrossRef] [PubMed]
- Du, C.; Liu, W.-J.; Yang, J.; Zhao, S.-S.; Liu, H.-X. The Role of Branched-Chain Amino Acids and Branched-Chain α-Keto Acid Dehydrogenase Kinase in Metabolic Disorders. Front. Nutr. 2022, 9, 932670. [Google Scholar] [CrossRef]
- Holeček, M. Branched-Chain Amino Acids in Health and Disease: Metabolism, Alterations in Blood Plasma, and as Supplements. Nutr. Metab. 2018, 15, 33. [Google Scholar] [CrossRef]
- Liang, Y.-F.; Long, Z.-X.; Zhang, Y.-J.; Luo, C.-Y.; Yan, L.-T.; Gao, W.-Y.; Li, H. The Chemical Mechanisms of the Enzymes in the Branched-Chain Amino Acids Biosynthetic Pathway and Their Applications. Biochimie 2021, 184, 72–87. [Google Scholar] [CrossRef] [PubMed]
- Holeček, M. Why Are Branched-Chain Amino Acids Increased in Starvation and Diabetes? Nutrients 2020, 12, 3087. [Google Scholar] [CrossRef] [PubMed]
- Mann, G.; Mora, S.; Madu, G.; Adegoke, O.A.J. Branched-Chain Amino Acids: Catabolism in Skeletal Muscle and Implications for Muscle and Whole-Body Metabolism. Front. Physiol. 2021, 12, 702826. [Google Scholar] [CrossRef] [PubMed]
- Ye, Z.; Wang, S.; Zhang, C.; Zhao, Y. Coordinated Modulation of Energy Metabolism and Inflammation by Branched-Chain Amino Acids and Fatty Acids. Front. Endocrinol. 2020, 11, 617. [Google Scholar] [CrossRef] [PubMed]
- Sivanand, S.; Vander Heiden, M.G. Emerging Roles for Branched-Chain Amino Acid Metabolism in Cancer. Cancer Cell 2020, 37, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Supruniuk, E.; Żebrowska, E.; Chabowski, A. Branched Chain Amino Acids—Friend or Foe in the Control of Energy Substrate Turnover and Insulin Sensitivity? Crit. Rev. Food Sci. Nutr. 2023, 63, 2559–2597. [Google Scholar] [CrossRef] [PubMed]
- Yahsi, B.; Gunaydin, G. Immunometabolism—The Role of Branched-Chain Amino Acids. Front. Immunol. 2022, 13, 886822. [Google Scholar] [CrossRef] [PubMed]
- Lian, K.; Guo, X.; Wang, Q.; Liu, Y.; Wang, R.-T.; Gao, C.; Li, C.-Y.; Li, C.-X.; Tao, L. PP2Cm Overexpression Alleviates MI/R Injury Mediated by a BCAA Catabolism Defect and Oxidative Stress in Diabetic Mice. Eur. J. Pharmacol. 2020, 866, 172796. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Wang, M.; Ning, F.; Zhou, S.; Hu, X.; Xin, H.; Reilly, S.; Zhang, X. Emerging Role for Branched-Chain Amino Acids Metabolism in Fibrosis. Pharmacol. Res. 2023, 187, 106604. [Google Scholar] [CrossRef]
- Arany, Z.; Neinast, M. Branched Chain Amino Acids in Metabolic Disease. Curr. Diab Rep. 2018, 18, 76. [Google Scholar] [CrossRef]
- Zhang, Z.-Y.; Monleon, D.; Verhamme, P.; Staessen, J.A. Branched-Chain Amino Acids as Critical Switches in Health and Disease. Hypertension 2018, 72, 1012–1022. [Google Scholar] [CrossRef] [PubMed]
- Vanweert, F.; Schrauwen, P.; Phielix, E. Role of Branched-Chain Amino Acid Metabolism in the Pathogenesis of Obesity and Type 2 Diabetes-Related Metabolic Disturbances BCAA Metabolism in Type 2 Diabetes. Nutr. Diabetes 2022, 12, 35. [Google Scholar] [CrossRef] [PubMed]
- Trautman, M.E.; Richardson, N.E.; Lamming, D.W. Protein Restriction and Branched-chain Amino Acid Restriction Promote Geroprotective Shifts in Metabolism. Aging Cell 2022, 21, e13626. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Wang, Y. Branched Chain Amino Acid Metabolic Reprogramming in Heart Failure. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2016, 1862, 2270–2275. [Google Scholar] [CrossRef] [PubMed]
- Poznyak, A.V.; Sukhorukov, V.N.; Zhuravlev, A.; Orekhov, N.A.; Kalmykov, V.; Orekhov, A.N. Modulating mTOR Signaling as a Promising Therapeutic Strategy for Atherosclerosis. Int. J. Mol. Sci. 2022, 23, 1153. [Google Scholar] [CrossRef] [PubMed]
- Hinkle, J.S.; Rivera, C.N.; Vaughan, R.A. Branched-Chain Amino Acids and Mitochondrial Biogenesis: An Overview and Mechanistic Summary. Mol. Nutr. Food Res. 2022, 66, 2200109. [Google Scholar] [CrossRef] [PubMed]
- Kogot-Levin, A.; Riahi, Y.; Abramovich, I.; Mosenzon, O.; Agranovich, B.; Kadosh, L.; Ben-Haroush Schyr, R.; Kleiman, D.; Hinden, L.; Cerasi, E.; et al. Mapping the Metabolic Reprogramming Induced by Sodium-Glucose Cotransporter 2 Inhibition. JCI Insight 2023, 8, e164296. [Google Scholar] [CrossRef] [PubMed]
- Satomi, S.; Morio, A.; Miyoshi, H.; Nakamura, R.; Tsutsumi, R.; Sakaue, H.; Yasuda, T.; Saeki, N.; Tsutsumi, Y.M. Branched-Chain Amino Acids-Induced Cardiac Protection against Ischemia/Reperfusion Injury. Life Sci. 2020, 245, 117368. [Google Scholar] [CrossRef] [PubMed]
- Sciarretta, S.; Forte, M.; Frati, G.; Sadoshima, J. New Insights into the Role of mTOR Signaling in the Cardiovascular System. Circ. Res. 2018, 122, 489–505. [Google Scholar] [CrossRef]
- Gargalionis, A.N.; Papavassiliou, K.A.; Papavassiliou, A.G. mTOR Signaling: Recent Progress. Int. J. Mol. Sci. 2024, 25, 2587. [Google Scholar] [CrossRef]
- Suhara, T.; Baba, Y.; Shimada, B.K.; Higa, J.K.; Matsui, T. The mTOR Signaling Pathway in Myocardial Dysfunction in Type 2 Diabetes Mellitus. Curr. Diab Rep. 2017, 17, 38. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.; Ye, W.-R.; Shi, W.; Yin, P.; Chen, C.; He, Y.-B.; Chen, M.-F.; Zu, X.-B.; Cai, Y. Perfect Match: mTOR Inhibitors and Tuberous Sclerosis Complex. Orphanet J. Rare Dis. 2022, 17, 106. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Sowers, J.R.; Ren, J. Targeting Autophagy in Obesity: From Pathophysiology to Management. Nat. Rev. Endocrinol. 2018, 14, 356–376. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Ji, L.; Hu, J.; Zhao, Y.; Johnston, L.J.; Zhang, X.; Ma, X. Functional Amino Acids and Autophagy: Diverse Signal Transduction and Application. Int. J. Mol. Sci. 2021, 22, 11427. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Ming, X.-F. mTOR Signalling: The Molecular Interface Connecting Metabolic Stress, Aging and Cardiovascular Diseases: mTOR in Metabolic and Age-Related Cardiovascular Diseases. Obes. Rev. 2012, 13, 58–68. [Google Scholar] [CrossRef] [PubMed]
- Yoon, M.-S. The Emerging Role of Branched-Chain Amino Acids in Insulin Resistance and Metabolism. Nutrients 2016, 8, 405. [Google Scholar] [CrossRef] [PubMed]
- Sciarretta, S.; Forte, M.; Frati, G.; Sadoshima, J. The Complex Network of mTOR Signalling in the Heart. Cardiovasc. Res. 2022, 118, 424–439. [Google Scholar] [CrossRef] [PubMed]
- Yao, H.; Li, K.; Wei, J.; Lin, Y.; Liu, Y. The Contradictory Role of Branched-Chain Amino Acids in Lifespan and Insulin Resistance. Front. Nutr. 2023, 10, 1189982. [Google Scholar] [CrossRef] [PubMed]
- Kaur, A.; Sharma, S. Mammalian Target of Rapamycin (mTOR) as a Potential Therapeutic Target in Various Diseases. Inflammopharmacology 2017, 25, 293–312. [Google Scholar] [CrossRef]
- Bae, J.; Paltzer, W.G.; Mahmoud, A.I. The Role of Metabolism in Heart Failure and Regeneration. Front. Cardiovasc. Med. 2021, 8, 702920. [Google Scholar] [CrossRef]
- Xiang, L.; Nie, J.; Wang, L.; Wang, Y.; Shi, J.; Wei, J.; Lau, C.-W.; Cai, Z.; Huang, Y. Integrated Metabolomics Analysis of the Effect of PPARδ Agonist GW501516 on Catabolism of BCAAs and Carboxylic Acids in Diabetic Mice. Chin. Chem. Lett. 2021, 32, 2197–2202. [Google Scholar] [CrossRef]
- Montaigne, D.; Butruille, L.; Staels, B. PPAR Control of Metabolism and Cardiovascular Functions. Nat. Rev. Cardiol. 2021, 18, 809–823. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xiong, Z.; Yan, W.; Gao, E.; Cheng, H.; Wu, G.; Liu, Y.; Zhang, L.; Li, C.; Wang, S.; et al. Branched Chain Amino Acids Exacerbate Myocardial Ischemia/Reperfusion Vulnerability via Enhancing GCN2/ATF6/PPAR-α Pathway-Dependent Fatty Acid Oxidation. Theranostics 2020, 10, 5623–5640. [Google Scholar] [CrossRef] [PubMed]
- Castagneto-Gissey, L.; Angelini, G.; Mingrone, G.; Cavarretta, E.; Tenori, L.; Licari, C.; Luchinat, C.; Tiepner, A.L.; Basso, N.; Bornstein, S.R.; et al. The Early Reduction of Left Ventricular Mass after Sleeve Gastrectomy Depends on the Fall of Branched-Chain Amino Acid Circulating Levels. eBioMedicine 2022, 76, 103864. [Google Scholar] [CrossRef] [PubMed]
- Manolis, A.S.; Manolis, T.A.; Manolis, A.A. Ketone Bodies and Cardiovascular Disease: An Alternate Fuel Source to the Rescue. Int. J. Mol. Sci. 2023, 24, 3534. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Song, J.; Hu, S. Metabolic Remodeling of Substrate Utilization during Heart Failure Progression. Heart Fail. Rev. 2019, 24, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Felker, G.M.; Buttrick, P.; Rosenzweig, A.; Abel, E.D.; Allen, L.A.; Bristow, M.; Das, S.; DeVore, A.D.; Drakos, S.G.; Fang, J.C.; et al. Heart Failure Strategically Focused Research Network: Summary of Results and Future Directions. J. Am. Heart Assoc. 2022, 11, e025517. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.-Y.; Marrachelli, V.G.; Yang, W.-Y.; Trenson, S.; Huang, Q.-F.; Wei, F.-F.; Thijs, L.; Van Keer, J.; Monleon, D.; Verhamme, P.; et al. Diastolic Left Ventricular Function in Relation to Circulating Metabolic Biomarkers in a Population Study. Eur. J. Prev. Cardiol. 2019, 26, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Lopaschuk, G.D.; Karwi, Q.G.; Tian, R.; Wende, A.R.; Abel, E.D. Cardiac Energy Metabolism in Heart Failure. Circ. Res. 2021, 128, 1487–1513. [Google Scholar] [CrossRef]
- Ng, S.M.; Neubauer, S.; Rider, O.J. Myocardial Metabolism in Heart Failure. Curr. Heart Fail. Rep. 2023, 20, 63–75. [Google Scholar] [CrossRef]
- Siddik, M.A.B.; Shin, A.C. Recent Progress on Branched-Chain Amino Acids in Obesity, Diabetes, and Beyond. Endocrinol. Metab. 2019, 34, 234. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Huang, K.; Liu, F.; Lu, X.; Huang, J.; Gu, D. Association of Circulating Branched-Chain Amino Acids with Risk of Cardiovascular Disease: A Systematic Review and Meta-Analysis. Atherosclerosis 2022, 350, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Müller, J.; Bertsch, T.; Volke, J.; Schmid, A.; Klingbeil, R.; Metodiev, Y.; Karaca, B.; Kim, S.-H.; Lindner, S.; Schupp, T.; et al. Narrative Review of Metabolomics in Cardiovascular Disease. J. Thorac. Dis. 2021, 13, 2532–2550. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Canela, M.; Toledo, E.; Clish, C.B.; Hruby, A.; Liang, L.; Salas-Salvadó, J.; Razquin, C.; Corella, D.; Estruch, R.; Ros, E.; et al. Plasma Branched-Chain Amino Acids and Incident Cardiovascular Disease in the PREDIMED Trial. Clin. Chem. 2016, 62, 582–592. [Google Scholar] [CrossRef] [PubMed]
- Cosentino, R.G.; Churilla, J.R.; Josephson, S.; Molle-Rios, Z.; Hossain, M.J.; Prado, W.L.; Balagopal, P.B. Branched-Chain Amino Acids and Relationship with Inflammation in Youth With Obesity: A Randomized Controlled Intervention Study. J. Clin. Endocrinol. Metab. 2021, 106, 3129–3139. [Google Scholar] [CrossRef] [PubMed]
- Uddin, G.M.; Zhang, L.; Shah, S.; Fukushima, A.; Wagg, C.S.; Gopal, K.; Al Batran, R.; Pherwani, S.; Ho, K.L.; Boisvenue, J.; et al. Impaired Branched Chain Amino Acid Oxidation Contributes to Cardiac Insulin Resistance in Heart Failure. Cardiovasc. Diabetol. 2019, 18, 86. [Google Scholar] [CrossRef]
- Tharrey, M.; Mariotti, F.; Mashchak, A.; Barbillon, P.; Delattre, M.; Huneau, J.-F.; Fraser, G.E. Patterns of Amino Acids Intake Are Strongly Associated with Cardiovascular Mortality, Independently of the Sources of Protein. Int. J. Epidemiol. 2019, 49, 312–321. [Google Scholar] [CrossRef]
- Jacob, K.J.; Chevalier, S.; Lamarche, M.; Morais, J.A. Leucine Supplementation Does Not Alter Insulin Sensitivity in Prefrail and Frail Older Women Following a Resistance Training Protocol. J. Nutr. 2019, 149, 959–967. [Google Scholar] [CrossRef]
- Tobias, D.K.; Lawler, P.R.; Harada, P.H.; Demler, O.V.; Ridker, P.M.; Manson, J.E.; Cheng, S.; Mora, S. Circulating Branched-Chain Amino Acids and Incident Cardiovascular Disease in a Prospective Cohort of US Women. Circ. Genom. Precis. Med. 2018, 11, e002157. [Google Scholar] [CrossRef]
- Magnusson, M.; Lewis, G.D.; Ericson, U.; Orho-Melander, M.; Hedblad, B.; Engström, G.; Östling, G.; Clish, C.; Wang, T.J.; Gerszten, R.E.; et al. A Diabetes-Predictive Amino Acid Score and Future Cardiovascular Disease. Eur. Heart J. 2013, 34, 1982–1989. [Google Scholar] [CrossRef]
- Würtz, P.; Havulinna, A.S.; Soininen, P.; Tynkkynen, T.; Prieto-Merino, D.; Tillin, T.; Ghorbani, A.; Artati, A.; Wang, Q.; Tiainen, M.; et al. Metabolite Profiling and Cardiovascular Event Risk: A Prospective Study of 3 Population-Based Cohorts. Circulation 2015, 131, 774–785. [Google Scholar] [CrossRef] [PubMed]
- Welsh, P.; Rankin, N.; Li, Q.; Mark, P.B.; Würtz, P.; Ala-Korpela, M.; Marre, M.; Poulter, N.; Hamet, P.; Chalmers, J.; et al. Circulating Amino Acids and the Risk of Macrovascular, Microvascular and Mortality Outcomes in Individuals with Type 2 Diabetes: Results from the ADVANCE Trial. Diabetologia 2018, 61, 1581–1591. [Google Scholar] [CrossRef] [PubMed]
- Ottosson, F.; Smith, E.; Fernandez, C.; Melander, O. Plasma Metabolites Associate with All-Cause Mortality in Individuals with Type 2 Diabetes. Metabolites 2020, 10, 315. [Google Scholar] [CrossRef] [PubMed]
- Le Couteur, D.G.; Ribeiro, R.; Senior, A.; Hsu, B.; Hirani, V.; Blyth, F.M.; Waite, L.M.; Simpson, S.J.; Naganathan, V.; Cumming, R.G.; et al. Branched Chain Amino Acids, Cardiometabolic Risk Factors and Outcomes in Older Men: The Concord Health and Ageing in Men Project. J. Gerontol. Ser. A 2020, 75, 1805–1810. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.C.; Watkins, S.M.; Lorenzo, C.; Wagenknecht, L.E.; Il’yasova, D.; Chen, Y.-D.I.; Haffner, S.M.; Hanley, A.J. Branched-Chain Amino Acids and Insulin Metabolism: The Insulin Resistance Atherosclerosis Study (IRAS). Diabetes Care 2016, 39, 582–588. [Google Scholar] [CrossRef] [PubMed]
- Lotta, L.A.; Scott, R.A.; Sharp, S.J.; Burgess, S.; Luan, J.; Tillin, T.; Schmidt, A.F.; Imamura, F.; Stewart, I.D.; Perry, J.R.B.; et al. Genetic Predisposition to an Impaired Metabolism of the Branched-Chain Amino Acids and Risk of Type 2 Diabetes: A Mendelian Randomisation Analysis. PLoS Med. 2016, 13, e1002179. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.J.; Larson, M.G.; Vasan, R.S.; Cheng, S.; Rhee, E.P.; McCabe, E.; Lewis, G.D.; Fox, C.S.; Jacques, P.F.; Fernandez, C.; et al. Metabolite Profiles and the Risk of Developing Diabetes. Nat. Med. 2011, 17, 448–453. [Google Scholar] [CrossRef] [PubMed]
- Tillin, T.; Hughes, A.D.; Wang, Q.; Würtz, P.; Ala-Korpela, M.; Sattar, N.; Forouhi, N.G.; Godsland, I.F.; Eastwood, S.V.; McKeigue, P.M.; et al. Diabetes Risk and Amino Acid Profiles: Cross-Sectional and Prospective Analyses of Ethnicity, Amino Acids and Diabetes in a South Asian and European Cohort from the SABRE (Southall and Brent REvisited) Study. Diabetologia 2015, 58, 968–979. [Google Scholar] [CrossRef] [PubMed]
- Cardelo, M.M.P. Estrategias Dietéticas en el Manejo de la Diabetes Mellitus tipo 2 en Pacientes con Enfermedad Coronaria Establecida: Estudio Cordioprev. Ph.D. Thesis, University of Córdoba, Córdoba, Spain, 2023. [Google Scholar]
- Santos-Gallego, C.G.; Requena-Ibanez, J.A.; San Antonio, R.; Ishikawa, K.; Watanabe, S.; Picatoste, B.; Flores, E.; Garcia-Ropero, A.; Sanz, J.; Hajjar, R.J.; et al. Empagliflozin Ameliorates Adverse Left Ventricular Remodeling in Nondiabetic Heart Failure by Enhancing Myocardial Energetics. J. Am. Coll. Cardiol. 2019, 73, 1931–1944. [Google Scholar] [CrossRef]
- Fine, K.S.; Wilkins, J.T.; Sawicki, K.T. Circulating Branched Chain Amino Acids and Cardiometabolic Disease. J. Am. Heart Assoc. 2024, 13, e031617. [Google Scholar] [CrossRef]
- Chen, M.; Gao, C.; Yu, J.; Ren, S.; Wang, M.; Wynn, R.M.; Chuang, D.T.; Wang, Y.; Sun, H. Therapeutic Effect of Targeting Branched-Chain Amino Acid Catabolic Flux in Pressure-Overload Induced Heart Failure. J. Am. Heart Assoc. 2019, 8, e011625. [Google Scholar] [CrossRef]
- Birkenfeld, A.L.; Jordan, J.; Dworak, M.; Merkel, T.; Burnstock, G. Myocardial Metabolism in Heart Failure: Purinergic Signalling and Other Metabolic Concepts. Pharmacol. Ther. 2019, 194, 132–144. [Google Scholar] [CrossRef]
- Ragni, M.; Greco, C.M.; Felicetta, A.; Ren, S.V.; Kunderfranco, P.; Ruocco, C.; Carullo, P.; Larcher, V.; Tedesco, L.; Severi, I.; et al. Dietary Essential Amino Acids for the Treatment of Heart Failure with Reduced Ejection Fraction. Cardiovasc. Res. 2023, 119, 982–997. [Google Scholar] [CrossRef]
- Li, R.; He, H.; Fang, S.; Hua, Y.; Yang, X.; Yuan, Y.; Liang, S.; Liu, P.; Tian, Y.; Xu, F.; et al. Time Series Characteristics of Serum Branched-Chain Amino Acids for Early Diagnosis of Chronic Heart Failure. J. Proteome Res. 2019, 18, 2121–2128. [Google Scholar] [CrossRef]
- Wang, W.; Zhang, F.; Xia, Y.; Zhao, S.; Yan, W.; Wang, H.; Lee, Y.; Li, C.; Zhang, L.; Lian, K.; et al. Defective Branched Chain Amino Acid Catabolism Contributes to Cardiac Dysfunction and Remodeling Following Myocardial Infarction. Am. J. Physiol. Heart Circ. Physiol. 2016, 311, H1160–H1169. [Google Scholar] [CrossRef]
- Morio, A.; Tsutsumi, R.; Satomi, S.; Kondo, T.; Miyoshi, H.; Kato, T.; Kuroda, M.; Kitamura, T.; Hara, K.; Saeki, N.; et al. Leucine Imparts Cardioprotective Effects by Enhancing mTOR Activity and Mitochondrial Fusion in a Myocardial Ischemia/Reperfusion Injury Murine Model. Diabetol. Metab. Syndr. 2021, 13, 139. [Google Scholar] [CrossRef] [PubMed]
- Kimura, Y.; Okumura, T.; Kazama, S.; Shibata, N.; Oishi, H.; Arao, Y.; Kuwayama, T.; Kato, H.; Yamaguchi, S.; Hiraiwa, H.; et al. Usefulness of Plasma Branched-Chain Amino Acid Analysis in Predicting Outcomes of Patients with Nonischemic Dilated Cardiomyopathy. Int. Heart J. 2020, 61, 739–747. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Cao, N.; Rau, C.D.; Lee, R.-P.; Yang, J.; Flach, R.J.R.; Petersen, L.; Zhu, C.; Pak, Y.-L.; Miller, R.A.; et al. Cell-Autonomous Effect of Cardiomyocyte Branched-Chain Amino Acid Catabolism in Heart Failure in Mice. Acta Pharmacol. Sin. 2023, 44, 1380–1390. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Olson, K.C.; Gao, C.; Prosdocimo, D.A.; Zhou, M.; Wang, Z.; Jeyaraj, D.; Youn, J.-Y.; Ren, S.; Liu, Y.; et al. Catabolic Defect of Branched-Chain Amino Acids Promotes Heart Failure. Circulation 2016, 133, 2038–2049. [Google Scholar] [CrossRef]
- Xie, H.; Zhang, B.; Xie, M.; Li, T. Circulating Metabolic Signatures of Heart Failure in Precision Cardiology. Precis. Clin. Med. 2023, 6, pbad005. [Google Scholar] [CrossRef]
- Sansbury, B.E.; DeMartino, A.M.; Xie, Z.; Brooks, A.C.; Brainard, R.E.; Watson, L.J.; DeFilippis, A.P.; Cummins, T.D.; Harbeson, M.A.; Brittian, K.R.; et al. Metabolomic Analysis of Pressure-Overloaded and Infarcted Mouse Hearts. Circ. Heart Fail. 2014, 7, 634–642. [Google Scholar] [CrossRef] [PubMed]
- Spyropoulos, F.; Sorrentino, A.; Van Der Reest, J.; Yang, P.; Waldeck-Weiermair, M.; Steinhorn, B.; Eroglu, E.; Saeedi Saravi, S.S.; Yu, P.; Haigis, M.; et al. Metabolomic and Transcriptomic Signatures of Chemogenetic Heart Failure. Am. J. Physiol. Heart Circ. Physiol. 2022, 322, H451–H465. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Xie, X.; Cao, F.; Wang, Y. Mitochondrial Metabolism in Myocardial Remodeling and Mechanical Unloading: Implications for Ischemic Heart Disease. Front. Cardiovasc. Med. 2021, 8, 789267. [Google Scholar] [CrossRef]
- Lerman, J.B.; Giamberardino, S.N.; Hernandez, A.F.; Felker, G.M.; Shah, S.H.; McGarrah, R.W. Plasma Metabolites Associated with Functional and Clinical Outcomes in Heart Failure with Reduced Ejection Fraction with and without Type 2 Diabetes. Sci. Rep. 2022, 12, 9183. [Google Scholar] [CrossRef] [PubMed]
- Tso, S.-C.; Gui, W.-J.; Wu, C.-Y.; Chuang, J.L.; Qi, X.; Skvorak, K.J.; Dorko, K.; Wallace, A.L.; Morlock, L.K.; Lee, B.H.; et al. Benzothiophene Carboxylate Derivatives as Novel Allosteric Inhibitors of Branched-Chain α-Ketoacid Dehydrogenase Kinase. J. Biol. Chem. 2014, 289, 20583–20593. [Google Scholar] [CrossRef] [PubMed]
- White, P.J.; McGarrah, R.W.; Grimsrud, P.A.; Tso, S.-C.; Yang, W.-H.; Haldeman, J.M.; Grenier-Larouche, T.; An, J.; Lapworth, A.L.; Astapova, I.; et al. The BCKDH Kinase and Phosphatase Integrate BCAA and Lipid Metabolism via Regulation of ATP-Citrate Lyase. Cell Metab. 2018, 27, 1281–1293.e7. [Google Scholar] [CrossRef]
- Hatahet, J.; Cook, T.M.; Bonomo, R.R.; Elshareif, N.; Gavini, C.K.; White, C.R.; Jesse, J.; Mansuy-Aubert, V.; Aubert, G. Fecal Microbiome Transplantation and Tributyrin Improves Early Cardiac Dysfunction and Modifies the BCAA Metabolic Pathway in a Diet Induced Pre-HFpEF Mouse Model. Front. Cardiovasc. Med. 2023, 10, 1105581. [Google Scholar] [CrossRef] [PubMed]
- Caragnano, A.; Aleksova, A.; Bulfoni, M.; Cervellin, C.; Rolle, I.G.; Veneziano, C.; Barchiesi, A.; Mimmi, M.C.; Vascotto, C.; Finato, N.; et al. Autophagy and Inflammasome Activation in Dilated Cardiomyopathy. J. Clin. Med. 2019, 8, 1519. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, T.; Yamashita, T.; Takahashi, T.; Tabata, T.; Watanabe, H.; Gotoh, Y.; Shinohara, M.; Kami, K.; Tanaka, H.; Matsumoto, K.; et al. Uncovering the Role of Gut Microbiota in Amino Acid Metabolic Disturbances in Heart Failure through Metagenomic Analysis. Front. Cardiovasc. Med. 2021, 8, 789325. [Google Scholar] [CrossRef]
- Biswas, D.; Duffley, L.; Pulinilkunnil, T. Role of Branched-chain Amino Acid–Catabolizing Enzymes in Intertissue Signaling, Metabolic Remodeling, and Energy Homeostasis. FASEB J. 2019, 33, 8711–8731. [Google Scholar] [CrossRef]
- Narita, K.; Amiya, E. Is Branched-Chain Amino Acid Nutritional Supplementation Beneficial or Detrimental in Heart Failure? World J. Cardiol. 2021, 13, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Voronova, V.; Sokolov, V.; Morias, Y.; Boezelman, M.J.; Wågberg, M.; Henricsson, M.; Hansson, K.; Goltsov, A.; Peskov, K.; Sundqvist, M. Evaluation of Therapeutic Strategies Targeting BCAA Catabolism Using a Systems Pharmacology Model. Front. Pharmacol. 2022, 13, 993422. [Google Scholar] [CrossRef] [PubMed]
- Tanada, Y.; Shioi, T.; Kato, T.; Kawamoto, A.; Okuda, J.; Kimura, T. Branched-Chain Amino Acids Ameliorate Heart Failure with Cardiac Cachexia in Rats. Life Sci. 2015, 137, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Lim, L.; Lau, E.S.H.; Fung, E.; Lee, H.; Ma, R.C.W.; Tam, C.H.T.; Wong, W.K.K.; Ng, A.C.W.; Chow, E.; Luk, A.O.Y.; et al. Circulating Branched-chain Amino Acids and Incident Heart Failure in Type 2 Diabetes: The Hong Kong Diabetes Register. Diabetes Metab. Res. Rev. 2020, 36, e3253. [Google Scholar] [CrossRef] [PubMed]
- Uchino, Y.; Watanabe, M.; Takata, M.; Amiya, E.; Tsushima, K.; Adachi, T.; Hiroi, Y.; Funazaki, T.; Komuro, I. Effect of Oral Branched-Chain Amino Acids on Serum Albumin Concentration in Heart Failure Patients with Hypoalbuminemia: Results of a Preliminary Study. Am. J. Cardiovasc. Drugs 2018, 18, 327–332. [Google Scholar] [CrossRef] [PubMed]
- Lanfear, D.E.; Gibbs, J.J.; Li, J.; She, R.; Petucci, C.; Culver, J.A.; Tang, W.H.W.; Pinto, Y.M.; Williams, L.K.; Sabbah, H.N.; et al. Targeted Metabolomic Profiling of Plasma and Survival in Heart Failure Patients. JACC Heart Fail. 2017, 5, 823–832. [Google Scholar] [CrossRef] [PubMed]
- Karwi, Q.G.; Zhang, L.; Wagg, C.S.; Wang, W.; Ghandi, M.; Thai, D.; Yan, H.; Ussher, J.R.; Oudit, G.Y.; Lopaschuk, G.D. Targeting the Glucagon Receptor Improves Cardiac Function and Enhances Insulin Sensitivity Following a Myocardial Infarction. Cardiovasc. Diabetol. 2019, 18, 1. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; You, H.; Li, Y.; Wang, Y.; Hui, P.; Qiao, B.; Lu, J.; Zhang, W.; Zhou, S.; Zheng, Y.; et al. Relationships between Circulating Branched Chain Amino Acid Concentrations and Risk of Adverse Cardiovascular Events in Patients with STEMI Treated with PCI. Sci. Rep. 2018, 8, 15809. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Li, Y.; Wang, Y.; You, H.; Hui, P.; Zheng, Y.; Du, J. Increased Branched-Chain Amino Acid Levels Are Associated with Long-Term Adverse Cardiovascular Events in Patients with STEMI and Acute Heart Failure. Life Sci. 2018, 209, 167–172. [Google Scholar] [CrossRef]
- Uddin, G.M.; Karwi, Q.G.; Pherwani, S.; Gopal, K.; Wagg, C.S.; Biswas, D.; Atnasious, M.; Wu, Y.; Wu, G.; Zhang, L.; et al. Deletion of BCATm Increases Insulin-Stimulated Glucose Oxidation in the Heart. Metabolism 2021, 124, 154871. [Google Scholar] [CrossRef]
- Asghari, G.; Farhadnejad, H.; Teymoori, F.; Mirmiran, P.; Tohidi, M.; Azizi, F. High Dietary Intake of Branched-Chain Amino Acids Is Associated with an Increased Risk of Insulin Resistance in Adults. J. Diabetes 2018, 10, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Latimer, M.N.; Sonkar, R.; Mia, S.; Frayne, I.R.; Carter, K.J.; Johnson, C.A.; Rana, S.; Xie, M.; Rowe, G.C.; Wende, A.R.; et al. Branched Chain Amino Acids Selectively Promote Cardiac Growth at the End of the Awake Period. J. Mol. Cell. Cardiol. 2021, 157, 31–44. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; Hsieh, P.N.; Sweet, D.R.; Jain, M.K. Krüppel-like Factor 15: Regulator of BCAA Metabolism and Circadian Protein Rhythmicity. Pharmacol. Res. 2018, 130, 123–126. [Google Scholar] [CrossRef]
- Shao, D.; Villet, O.; Zhang, Z.; Choi, S.W.; Yan, J.; Ritterhoff, J.; Gu, H.; Djukovic, D.; Christodoulou, D.; Kolwicz, S.C.; et al. Glucose Promotes Cell Growth by Suppressing Branched-Chain Amino Acid Degradation. Nat. Commun. 2018, 9, 2935. [Google Scholar] [CrossRef] [PubMed]
- Flores-Guerrero, J.; Osté, M.; Kieneker, L.; Gruppen, E.; Wolak-Dinsmore, J.; Otvos, J.; Connelly, M.; Bakker, S.; Dullaart, R. Plasma Branched-Chain Amino Acids and Risk of Incident Type 2 Diabetes: Results from the PREVEND Prospective Cohort Study. J. Clin. Med. 2018, 7, 513. [Google Scholar] [CrossRef]
- Mardinoglu, A.; Gogg, S.; Lotta, L.A.; Stančáková, A.; Nerstedt, A.; Boren, J.; Blüher, M.; Ferrannini, E.; Langenberg, C.; Wareham, N.J.; et al. Elevated Plasma Levels of 3-Hydroxyisobutyric Acid Are Associated with Incident Type 2 Diabetes. EBioMedicine 2018, 27, 151–155. [Google Scholar] [CrossRef]
- Bragg, F.; Trichia, E.; Aguilar-Ramirez, D.; Bešević, J.; Lewington, S.; Emberson, J. Predictive Value of Circulating NMR Metabolic Biomarkers for Type 2 Diabetes Risk in the UK Biobank Study. BMC Med. 2022, 20, 159. [Google Scholar] [CrossRef]
- Bragg, F.; Kartsonaki, C.; Guo, Y.; Holmes, M.; Du, H.; Yu, C.; Pei, P.; Yang, L.; Jin, D.; Chen, Y.; et al. The Role of NMR-Based Circulating Metabolic Biomarkers in Development and Risk Prediction of New Onset Type 2 Diabetes. Sci. Rep. 2022, 12, 15071. [Google Scholar] [CrossRef]
- Sawicki, K.T.; Ning, H.; Allen, N.B.; Carnethon, M.R.; Wallia, A.; Otvos, J.D.; Ben-Sahra, I.; McNally, E.M.; Snell-Bergeon, J.K.; Wilkins, J.T. Longitudinal Trajectories of Branched Chain Amino Acids through Young Adulthood and Diabetes in Later Life. JCI Insight 2023, 8, e166956. [Google Scholar] [CrossRef]
- Morze, J.; Wittenbecher, C.; Schwingshackl, L.; Danielewicz, A.; Rynkiewicz, A.; Hu, F.B.; Guasch-Ferré, M. Metabolomics and Type 2 Diabetes Risk: An Updated Systematic Review and Meta-Analysis of Prospective Cohort Studies. Diabetes Care 2022, 45, 1013–1024. [Google Scholar] [CrossRef]
- Guasch-Ferré, M.; Hruby, A.; Toledo, E.; Clish, C.B.; Martínez-González, M.A.; Salas-Salvadó, J.; Hu, F.B. Metabolomics in Prediabetes and Diabetes: A Systematic Review and Meta-Analysis. Diabetes Care 2016, 39, 833–846. [Google Scholar] [CrossRef] [PubMed]
- Gencer, B.; Li, X.S.; Gurmu, Y.; Bonaca, M.P.; Morrow, D.A.; Cohen, M.; Bhatt, D.L.; Steg, P.G.; Storey, R.F.; Johanson, P.; et al. Gut Microbiota-Dependent Trimethylamine N-oxide and Cardiovascular Outcomes in Patients with Prior Myocardial Infarction: A Nested Case Control Study From the PEGASUS-TIMI 54 Trial. J. Am. Heart Assoc. 2020, 9, e015331. [Google Scholar] [CrossRef] [PubMed]
- Croyal, M.; Saulnier, P.-J.; Aguesse, A.; Gand, E.; Ragot, S.; Roussel, R.; Halimi, J.-M.; Ducrocq, G.; Cariou, B.; Montaigne, D.; et al. Plasma Trimethylamine N-Oxide and Risk of Cardiovascular Events in Patients with Type 2 Diabetes. J. Clin. Endocrinol. Metab. 2020, 105, 2371–2380. [Google Scholar] [CrossRef] [PubMed]
- Cardona, A.; O’Brien, A.; Bernier, M.C.; Somogyi, A.; Wysocki, V.H.; Smart, S.; He, X.; Ambrosio, G.; Hsueh, W.A.; Raman, S.V. Trimethylamine N-Oxide and Incident Atherosclerotic Events in High-Risk Individuals with Diabetes: An ACCORD Trial Post Hoc Analysis. BMJ Open Diab Res. Care 2019, 7, e000718. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.H.W.; Wang, Z.; Li, X.S.; Fan, Y.; Li, D.S.; Wu, Y.; Hazen, S.L. Increased Trimethylamine N-Oxide Portends High Mortality Risk Independent of Glycemic Control in Patients with Type 2 Diabetes Mellitus. Clin. Chem. 2017, 63, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Flores-Guerrero, J.L.; Van Dijk, P.R.; Connelly, M.A.; Garcia, E.; Bilo, H.J.G.; Navis, G.; Bakker, S.J.L.; Dullaart, R.P.F. Circulating Trimethylamine N-Oxide Is Associated with Increased Risk of Cardiovascular Mortality in Type-2 Diabetes: Results from a Dutch Diabetes Cohort (ZODIAC-59). J. Clin. Med. 2021, 10, 2269. [Google Scholar] [CrossRef]
- Wang, W.; Liu, Z.; Liu, L.; Han, T.; Yang, X.; Sun, C. Genetic Predisposition to Impaired Metabolism of the Branched Chain Amino Acids, Dietary Intakes, and Risk of Type 2 Diabetes. Genes. Nutr. 2021, 16, 20. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Sun, L.; Gong, Y.; Zhou, Y.; Yang, P.; Ye, Z.; Fu, J.; Huang, A.; Fu, Z.; Yu, W.; et al. Relationship between Branched-Chain Amino Acids, Metabolic Syndrome, and Cardiovascular Risk Profile in a Chinese Population: A Cross-Sectional Study. Int. J. Endocrinol. 2016, 2016, 1–10. [Google Scholar] [CrossRef]
- Jennings, A.; MacGregor, A.; Pallister, T.; Spector, T.; Cassidy, A. Associations between Branched Chain Amino Acid Intake and Biomarkers of Adiposity and Cardiometabolic Health Independent of Genetic Factors: A Twin Study. Int. J. Cardiol. 2016, 223, 992–998. [Google Scholar] [CrossRef]
- Siomkajło, M.; Rybka, J.; Mierzchała-Pasierb, M.; Gamian, A.; Stankiewicz-Olczyk, J.; Bolanowski, M.; Daroszewski, J. Specific Plasma Amino Acid Disturbances Associated with Metabolic Syndrome. Endocrine 2017, 58, 553–562. [Google Scholar] [CrossRef]
- Mahbub, M.H.; Yamaguchi, N.; Hase, R.; Takahashi, H.; Ishimaru, Y.; Watanabe, R.; Saito, H.; Shimokawa, J.; Yamamoto, H.; Kikuchi, S.; et al. Plasma Branched-Chain and Aromatic Amino Acids in Relation to Hypertension. Nutrients 2020, 12, 3791. [Google Scholar] [CrossRef] [PubMed]
- Martin, M.; Deussen, A. Effects of Natural Peptides from Food Proteins on Angiotensin Converting Enzyme Activity and Hypertension. Crit. Rev. Food Sci. Nutr. 2019, 59, 1264–1283. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Yu, J.; Yuan, R.; Li, N.; Li, C.; Zhang, X. mTOR Inhibitor Improves Testosterone-Induced Myocardial Hypertrophy in Hypertensive Rats. J. Endocrinol. 2022, 252, 179–193. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, C.; Zhang, Y.; Jiang, X.; Liang, Y.; Wang, H.; Li, Y.; Sun, G. Association between Excessive Dietary Branched-Chain Amino Acids Intake and Hypertension Risk in Chinese Population. Nutrients 2022, 14, 2582. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Wu, Y.; Ye, L. The Role of Amino Acids in Endothelial Biology and Function. Cells 2022, 11, 1372. [Google Scholar] [CrossRef] [PubMed]
- Mirmiran, P.; Teymoori, F.; Asghari, G.; Azizi, F. Dietary Intakes of Branched Chain Amino Acids and the Incidence of Hypertension: A Population-Based Prospective Cohort Study. Arch. Iran. Med. 2019, 22, 182–188. [Google Scholar] [PubMed]
- Wang, F.; Wang, B.; Chen, X.; Liu, W.; Wang, G.; Li, X.; Liu, X.; Li, N.; Zhang, J.; Yin, T.; et al. Association Between Blood Pressure and Branched-Chain/Aromatic Amino Acid Excretion Rate in 24-Hour Urine Samples from Elderly Hypertension Patients. Diabetes Metab. Syndr. Obes. 2021, 14, 3965–3973. [Google Scholar] [CrossRef] [PubMed]
- Flores-Guerrero, J.L.; Groothof, D.; Connelly, M.A.; Otvos, J.D.; Bakker, S.J.L.; Dullaart, R.P.F. Concentration of Branched-Chain Amino Acids Is a Strong Risk Marker for Incident Hypertension. Hypertension 2019, 74, 1428–1435. [Google Scholar] [CrossRef]
- Yamaguchi, N.; Mahbub, M.; Takahashi, H.; Hase, R.; Ishimaru, Y.; Sunagawa, H.; Amano, H.; Kobayashi- Miura, M.; Kanda, H.; Fujita, Y.; et al. Plasma Free Amino Acid Profiles Evaluate Risk of Metabolic Syndrome, Diabetes, Dyslipidemia, and Hypertension in a Large Asian Population. Environ. Health Prev. Med. 2017, 22, 35. [Google Scholar] [CrossRef]
- Poggiogalle, E.; Fontana, M.; Giusti, A.M.; Pinto, A.; Iannucci, G.; Lenzi, A.; Donini, L.M. Amino Acids and Hypertension in Adults. Nutrients 2019, 11, 1459. [Google Scholar] [CrossRef]
- Jennings, A.; MacGregor, A.; Welch, A.; Chowienczyk, P.; Spector, T.; Cassidy, A. Amino Acid Intakes Are Inversely Associated with Arterial Stiffness andCentral Blood Pressure in Women. J. Nutr. 2015, 145, 2130–2138. [Google Scholar] [CrossRef] [PubMed]
- Teymoori, F.; Asghari, G.; Mirmiran, P.; Azizi, F. Dietary Amino Acids and Incidence of Hypertension: A Principle Component Analysis Approach. Sci. Rep. 2017, 7, 16838. [Google Scholar] [CrossRef] [PubMed]
- Amin, A.M. Metabolomics Applications in Coronary Artery Disease Personalized Medicine. In Advances in Clinical Chemistry; Elsevier: Amsterdam, The Netherlands, 2021; Volume 102, pp. 233–270. ISBN 978-0-12-824614-6. [Google Scholar]
- Xu, H.; Wang, X.; Geng, G.; Xu, X.; Liu, L.; Zhang, Y.; Wang, Z.; Wang, L.; Li, Y. Association of Circulating Branched-Chain Amino Acids with Cardiovascular Diseases: A Mendelian Randomization Study. Nutrients 2023, 15, 1580. [Google Scholar] [CrossRef] [PubMed]
- Grajeda-Iglesias, C.; Aviram, M. Specific Amino Acids Affect Cardiovascular Diseases and Atherogenesis via Protection against Macrophage Foam Cell Formation: Review Article. Rambam Maimonides Med. J. 2018, 9, e0022. [Google Scholar] [CrossRef] [PubMed]
- Zaric, B.L.; Radovanovic, J.N.; Gluvic, Z.; Stewart, A.J.; Essack, M.; Motwalli, O.; Gojobori, T.; Isenovic, E.R. Atherosclerosis Linked to Aberrant Amino Acid Metabolism and Immunosuppressive Amino Acid Catabolizing Enzymes. Front. Immunol. 2020, 11, 551758. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhang, R.; Mu, H.; Zhang, W.; Zeng, J.; Li, H.; Wang, S.; Zhao, X.; Chen, W.; Dong, J.; et al. Oral Administration of Branched-Chain Amino Acids Attenuates Atherosclerosis by Inhibiting the Inflammatory Response and Regulating the Gut Microbiota in ApoE-Deficient Mice. Nutrients 2022, 14, 5065. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Jiang, H.; Li, L.; Chen, F.; Liu, Y.; Zhou, M.; Wang, J.; Jiang, J.; Li, X.; Fan, X.; et al. Branched-Chain Amino Acid Catabolism Promotes Thrombosis Risk by Enhancing Tropomodulin-3 Propionylation in Platelets. Circulation 2020, 142, 49–64. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Zhang, Z.; Kolwicz, S.C.; Abell, L.; Roe, N.D.; Kim, M.; Zhou, B.; Cao, Y.; Ritterhoff, J.; Gu, H.; et al. Defective Branched-Chain Amino Acid Catabolism Disrupts Glucose Metabolism and Sensitizes the Heart to Ischemia-Reperfusion Injury. Cell Metab. 2017, 25, 374–385. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Zhou, L.; Wang, Q.; Cao, J.-H.; Chen, Y.; Wang, W.; Zhu, B.-D.; Wei, Z.-H.; Li, R.; Li, C.-Y.; et al. Elevated Branched-Chain Amino Acid Promotes Atherosclerosis Progression by Enhancing Mitochondrial-to-Nuclear H2O2-Disulfide HMGB1 in Macrophages. Redox Biol. 2023, 62, 102696. [Google Scholar] [CrossRef]
- Jiang, Y.-J.; Sun, S.-J.; Cao, W.-X.; Lan, X.-T.; Ni, M.; Fu, H.; Li, D.-J.; Wang, P.; Shen, F.-M. Excessive ROS Production and Enhanced Autophagy Contribute to Myocardial Injury Induced by Branched-Chain Amino Acids: Roles for the AMPK-ULK1 Signaling Pathway and α7nAChR. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2021, 1867, 165980. [Google Scholar] [CrossRef]
- Zhenyukh, O.; González-Amor, M.; Rodrigues-Diez, R.R.; Esteban, V.; Ruiz-Ortega, M.; Salaices, M.; Mas, S.; Briones, A.M.; Egido, J. Branched-chain Amino Acids Promote Endothelial Dysfunction through Increased Reactive Oxygen Species Generation and Inflammation. J. Cell Mol. Med. 2018, 22, 4948–4962. [Google Scholar] [CrossRef] [PubMed]
- Liao, M.; Hu, F.; Qiu, Z.; Li, J.; Huang, C.; Xu, Y.; Cheng, X. Pim-2 Kinase Inhibits Inflammation by Suppressing the mTORC1 Pathway in Atherosclerosis. Aging 2021, 13, 22412–22431. [Google Scholar] [CrossRef] [PubMed]
- Reho, J.J.; Guo, D.; Rahmouni, K. Mechanistic Target of Rapamycin Complex 1 Signaling Modulates Vascular Endothelial Function Through Reactive Oxygen Species. J. Am. Heart Assoc. 2019, 8, e010662. [Google Scholar] [CrossRef] [PubMed]
- Ottosson, F.; Smith, E.; Melander, O.; Fernandez, C. Altered Asparagine and Glutamate Homeostasis Precede Coronary Artery Disease and Type 2 Diabetes. J. Clin. Endocrinol. Metab. 2018, 103, 3060–3069. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, Y.; Ma, Z.; Zhu, Y.; Chen, Z. Altered Amino Acid Metabolism between Coronary Heart Disease Patients with and without Type 2 Diabetes by Quantitative 1H NMR Based Metabolomics. J. Pharm. Biomed. Anal. 2021, 206, 114381. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.H.; Sun, J.-L.; Stevens, R.D.; Bain, J.R.; Muehlbauer, M.J.; Pieper, K.S.; Haynes, C.; Hauser, E.R.; Kraus, W.E.; Granger, C.B.; et al. Baseline Metabolomic Profiles Predict Cardiovascular Events in Patients at Risk for Coronary Artery Disease. Am. Heart J. 2012, 163, 844–850.e1. [Google Scholar] [CrossRef]
- Fan, Y.; Li, Y.; Chen, Y.; Zhao, Y.-J.; Liu, L.-W.; Li, J.; Wang, S.-L.; Alolga, R.N.; Yin, Y.; Wang, X.-M.; et al. Comprehensive Metabolomic Characterization of Coronary Artery Diseases. J. Am. Coll. Cardiol. 2016, 68, 1281–1293. [Google Scholar] [CrossRef] [PubMed]
- Tzoulaki, I.; Castagné, R.; Boulangé, C.L.; Karaman, I.; Chekmeneva, E.; Evangelou, E.; Ebbels, T.M.D.; Kaluarachchi, M.R.; Chadeau-Hyam, M.; Mosen, D.; et al. Serum Metabolic Signatures of Coronary and Carotid Atherosclerosis and Subsequent Cardiovascular Disease. Eur. Heart J. 2019, 40, 2883–2896. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Granger, C.B.; Craig, D.; Haynes, C.; Bain, J.; Stevens, R.D.; Hauser, E.R.; Newgard, C.B.; Kraus, W.E.; Newby, L.K.; et al. Validation of the Association between a Branched Chain Amino Acid Metabolite Profile and Extremes of Coronary Artery Disease in Patients Referred for Cardiac Catheterization. Atherosclerosis 2014, 232, 191–196. [Google Scholar] [CrossRef]
- Vaarhorst, A.A.M.; Verhoeven, A.; Weller, C.M.; Böhringer, S.; Göraler, S.; Meissner, A.; Deelder, A.M.; Henneman, P.; Gorgels, A.P.M.; Van Den Brandt, P.A.; et al. A Metabolomic Profile Is Associated with the Risk of Incident Coronary Heart Disease. Am. Heart J. 2014, 168, 45–52.e7. [Google Scholar] [CrossRef]
- Paynter, N.P.; Balasubramanian, R.; Giulianini, F.; Wang, D.D.; Tinker, L.F.; Gopal, S.; Deik, A.A.; Bullock, K.; Pierce, K.A.; Scott, J.; et al. Metabolic Predictors of Incident Coronary Heart Disease in Women. Circulation 2018, 137, 841–853. [Google Scholar] [CrossRef] [PubMed]
- Hamaya, R.; Mora, S.; Lawler, P.R.; Cook, N.R.; Buring, J.E.; Lee, I.-M.; Manson, J.E.; Tobias, D.K. Association of Modifiable Lifestyle Factors with Plasma Branched-Chain Amino Acid Metabolites in Women. J. Nutr. 2022, 152, 1515–1524. [Google Scholar] [CrossRef] [PubMed]
- Chevli, P.A.; Freedman, B.I.; Hsu, F.-C.; Xu, J.; Rudock, M.E.; Ma, L.; Parks, J.S.; Palmer, N.D.; Shapiro, M.D. Plasma Metabolomic Profiling in Subclinical Atherosclerosis: The Diabetes Heart Study. Cardiovasc. Diabetol. 2021, 20, 231. [Google Scholar] [CrossRef] [PubMed]
- Cruz, D.E.; Tahir, U.A.; Hu, J.; Ngo, D.; Chen, Z.-Z.; Robbins, J.M.; Katz, D.; Balasubramanian, R.; Peterson, B.; Deng, S.; et al. Metabolomic Analysis of Coronary Heart Disease in an African American Cohort from the Jackson Heart Study. JAMA Cardiol. 2022, 7, 184–194. [Google Scholar] [CrossRef] [PubMed]
- De La, O.V.; Zazpe, I.; Ruiz-Canela, M. Effect of Branched-Chain Amino Acid Supplementation, Dietary Intake and Circulating Levels in Cardiometabolic Diseases: An Updated Review. Curr. Opin. Clin. Nutr. Metab. Care 2020, 23, 35–50. [Google Scholar] [CrossRef]
- Doestzada, M.; Zhernakova, D.V.; Van Den Munckhof, I.C.L.; Wang, D.; Kurilshikov, A.; Chen, L.; Bloks, V.W.; Van Faassen, M.; Rutten, J.H.W.; Joosten, L.A.B.; et al. Systematic Analysis of Relationships between Plasma Branched-Chain Amino Acid Concentrations and Cardiometabolic Parameters: An Association and Mendelian Randomization Study. BMC Med. 2022, 20, 485. [Google Scholar] [CrossRef]
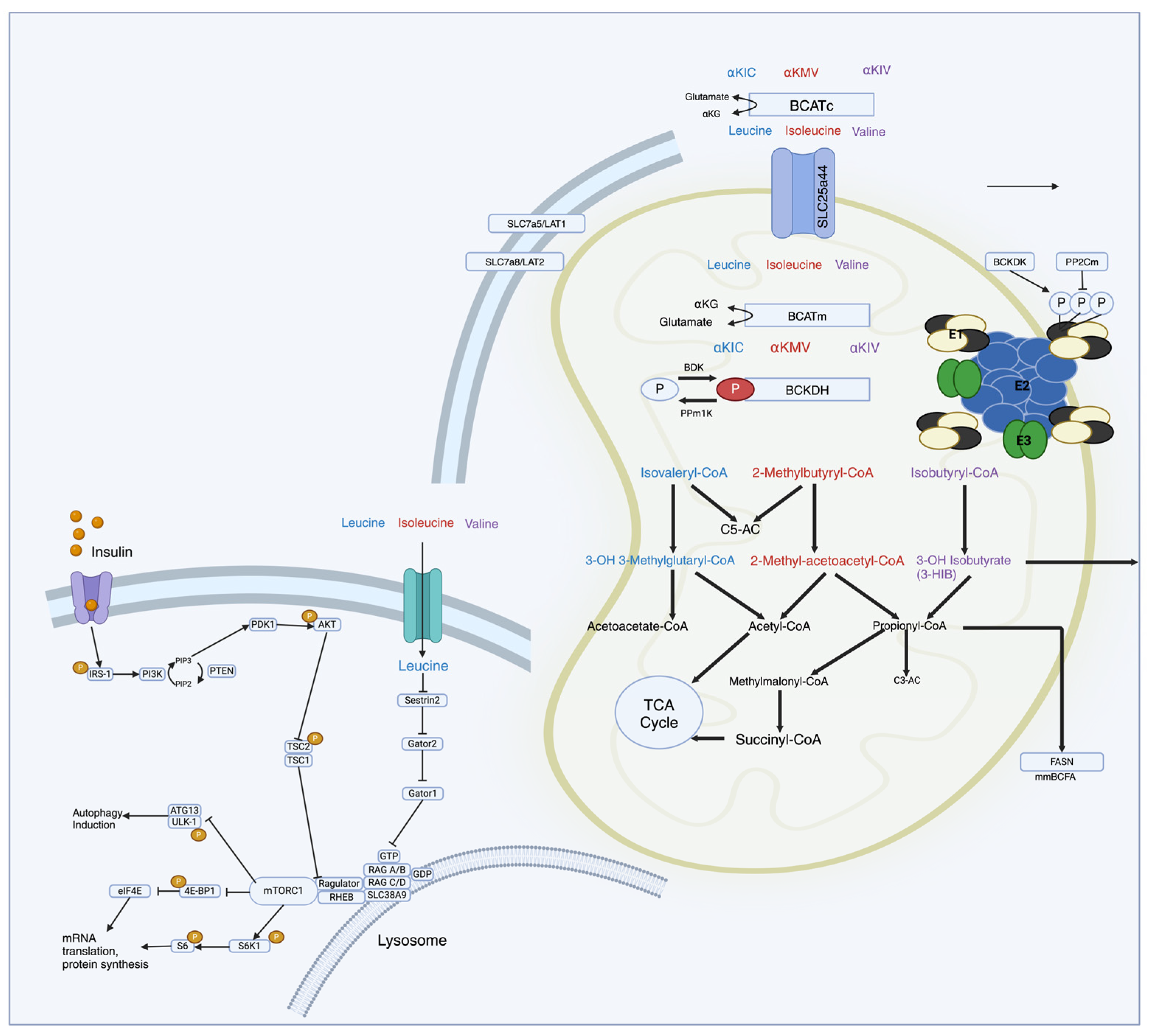
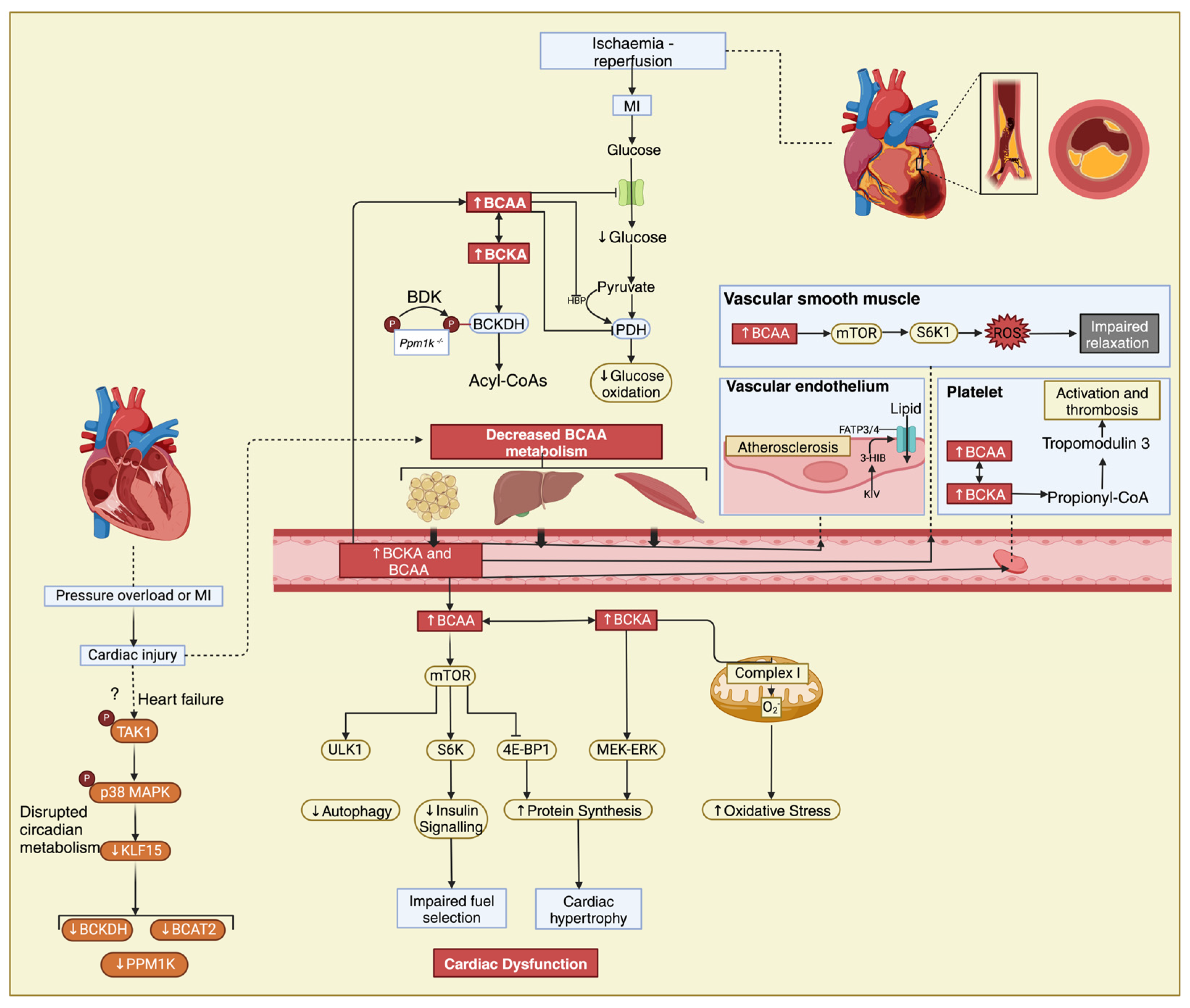
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).