
The Effects of Testosterone on Hypothalamic and Serum Oxytocin Levels Are Affected by the Estrogen Milieu in Female Rats
 ,
, {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Effects of Chronic Testosterone Administration on Ovariectomized Rats That Did Not Receive Estradiol Supplementation (OVX)
2.3. Effects of Chronic Testosterone Administration on Ovariectomized Rats That Received Estradiol Supplementation (OVX + E2)
2.4. Hormone Assay
2.5. Quantitative Real-Time Polymerase Chain Reaction
2.6. Statistical Analyses
3. Results
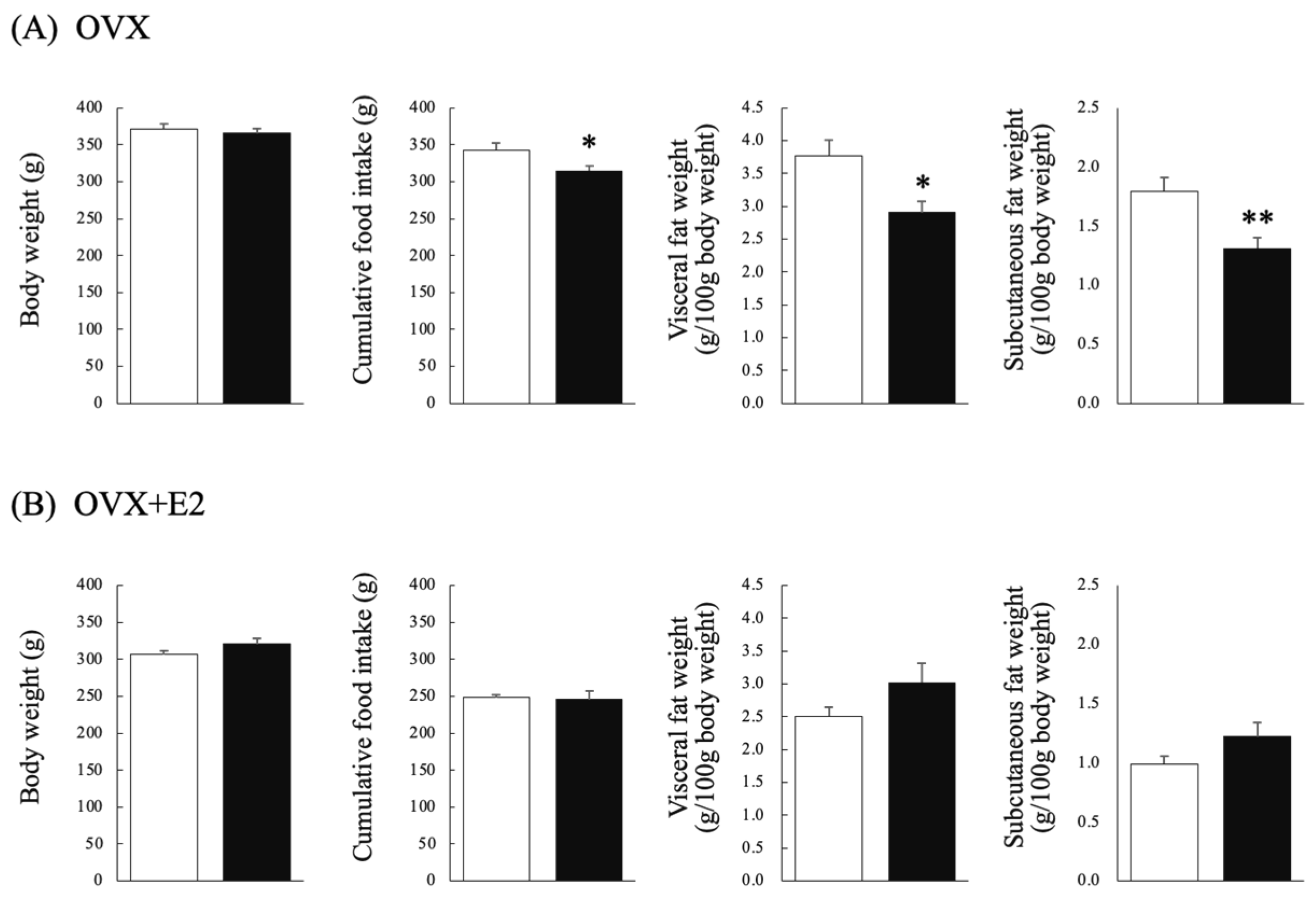
3.1. Effects of Chronic Testosterone Administration on BW and FI in Ovariectomized Rats That Did (OVX + E2) or Did Not (OVX) Receive Estradiol Supplementation
3.2. Effects of Chronic Testosterone Administration on Visceral Fat Weight and Subcutaneous Fat Weight in Ovariectomized Rats That Did (OVX + E2) or Did Not (OVX) Receive Estradiol Supplementation
3.3. Effects of Chronic Testosterone Administration on Serum OT Levels in Ovariectomized Rats That Did (OVX + E2) or Did Not (OVX) Receive Estradiol Supplementation
3.4. Effects of Chronic Testosterone Administration on Hypothalamic OT mRNA Levels, and Hypothalamic, Visceral Fat, and Subcutaneous Fat OTR mRNA Levels in Ovariectomized Rats That Did (OVX + E2) or Did Not (OVX) Receive Estradiol Supplementation
3.5. Correlations between Serum Testosterone Levels and Serum or Hypothalamic OT Levels in Ovariectomized Rats That Did (OVX + E2) or Did Not (OVX) Receive Estradiol Supplementation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Takeshi, I.; Matsuzaki, T.; Yano, K.; Yanagihara, R.; Tungalagsuvd, A.; Munkhzaya, M.; Mayila, Y.; Kuwahara, A.; Irahara, M. The Effects of Chronic Testosterone Administration on Body Weight, Food Intake, and Adipose Tissue Are Changed by Estrogen Treatment in Female Rats. Horm. Behav. 2017, 93, 53–61. [Google Scholar]
- Butera Peter, C.; Beikirch, R.J. Central Implants of Diluted Estradiol: Independent Effects on Ingestive and Reproductive Behaviors of Ovariectomized Rats. Brain Res. 1989, 491, 266–273. [Google Scholar] [CrossRef] [PubMed]
- Palmer, K.; Gray, J.M. Central vs. Peripheral Effects of Estrogen on Food Intake and Lipoprotein Lipase Activity in Ovariectomized Rats. Physiol. Behav. 1986, 37, 187–189. [Google Scholar] [CrossRef] [PubMed]
- Lindén Hirschberg, A.; Naessén, S.; Stridsberg, M.; Byström, B.; Holte, J. Impaired Cholecystokinin Secretion and Disturbed Appetite Regulation in Women with Polycystic Ovary Syndrome. Gynecol. Endocrinol. 2004, 19, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Naessén, S.; Carlström, K.; Byström, B.; Pierre, Y.; Hirschberg, A.L. Effects of an Antiandrogenic Oral Contraceptive on Appetite and Eating Behavior in Bulimic Women. Psychoneuroendocrinology 2007, 32, 548–554. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.S.; Norman, R.J.; Clifton, P.M.; Noakes, M. Hyperandrogenemia, Psychological Distress, and Food Cravings in Young Women. Physiol. Behav. 2009, 98, 276–280. [Google Scholar] [CrossRef] [PubMed]
- Hirschberg, A.L. Sex Hormones, Appetite and Eating Behaviour in Women. Maturitas 2012, 71, 248–256. [Google Scholar] [CrossRef]
- Hannan, F.M.; Elajnaf, T.; Vandenberg, L.N.; Kennedy, S.H.; Thakker, R.V. Hormonal Regulation of Mammary Gland Development and Lactation. Nat. Rev. Endocrinol. 2023, 19, 46–61. [Google Scholar] [CrossRef] [PubMed]
- Takayanagi, Y.; Kasahara, Y.; Onaka, T.; Takahashi, N.; Kawada, T.; Nishimori, K. Oxytocin Receptor-Deficient Mice Developed Late-Onset Obesity. NeuroReport 2008, 19, 951. [Google Scholar] [CrossRef]
- Yamamoto, S.; Noguchi, H.; Takeda, A.; Arakaki, R.; Uchishiba, M.; Imaizumi, J.; Minato, S.; Kamada, S.; Kagawa, T.; Yoshida, A.; et al. Changes in Endogenous Oxytocin Levels and the Effects of Exogenous Oxytocin Administration on Body Weight Changes and Food Intake in Polycystic Ovary Syndrome Model Rats. Int. J. Mol. Sci. 2022, 23, 8207. [Google Scholar] [CrossRef]
- Roberts, Z.S.; Wolden-Hanson, T.; Matsen, M.E.; Ryu, V.; Vaughan, C.H.; Graham, J.L.; Havel, P.J.; Chukri, D.W.; Schwartz, M.W.; Morton, G.J.; et al. Chronic Hindbrain Administration of Oxytocin Is Sufficient to Elicit Weight Loss in Diet-Induced Obese Rats. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2017, 313, R357–R371. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Cai, D. Circadian Intervention of Obesity Development via Resting-Stage Feeding Manipulation or Oxytocin Treatment. Am. J. Physiol.-Endocrinol. Metab. 2011, 301, E1004–E1012. [Google Scholar] [CrossRef] [PubMed]
- Maejima, Y.; Aoyama, M.; Sakamoto, K.; Jojima, T.; Aso, Y.; Takasu, K.; Takenosihita, S.; Shimomura, K. Impact of Sex, Fat Distribution and Initial Body Weight on Oxytocin’s Body Weight Regulation. Sci. Rep. 2017, 7, 8599. [Google Scholar] [CrossRef] [PubMed]
- Deblon, N.; Veyrat-Durebex, C.; Bourgoin, L.; Caillon, A.; Bussier, A.L.; Petrosino, S.; Piscitelli, F.; Legros, J.-J.; Geenen, V.; Foti, M.; et al. Mechanisms of the Anti-Obesity Effects of Oxytocin in Diet-Induced Obese Rats. PLoS ONE 2011, 6, e25565. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Gobrogge, K. Neurodevelopmental Model Explaining Associations between Sex Hormones, Personality, and Eating Pathology. Brain Sci. 2023, 13, 859. [Google Scholar] [CrossRef] [PubMed]
- Nikolaenko, L.; Jia, Y.; Wang, C.; Diaz-Arjonilla, M.; Yee, J.K.; French, S.W.; Liu, P.Y.; Laurel, S.; Chong, C.; Lee, K.; et al. Testosterone Replacement Ameliorates Nonalcoholic Fatty Liver Disease in Castrated Male Rats. Endocrinology 2014, 155, 417–428. [Google Scholar] [CrossRef] [PubMed]
- Erdenebayar, O.; Kato, T.; Kawakita, T.; Kasai, K.; Kadota, Y.; Yoshida, K.; Iwasa, T.; Irahara, M. Effects of Peripheral Oxytocin Administration on Body Weight, Food Intake, Adipocytes, and Biochemical Parameters in Peri- and Postmenopausal Female Rats. Endocr. J. 2021, 68, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, S.; Arakaki, R.; Noguchi, H.; Takeda, A.; Uchishiba, M.; Kamada, S.; Mineda, A.; Kon, M.; Kawakita, T.; Kinouchi, R.; et al. New Discoveries on the Interaction between Testosterone and Oxytocin in Male Rats—Testosterone-Mediated Effects of Oxytocin in the Prevention of Obesity. Physiol. Behav. 2023, 266, 114199. [Google Scholar] [CrossRef]
- Iwasa, T.; Noguchi, H.; Tanano, R.; Yamanaka, E.; Takeda, A.; Tamura, K.; Aoki, H.; Sugimoto, T.; Sasada, H.; Maeda, T.; et al. Age-Dependent Changes in the Effects of Androgens on Female Metabolic and Body Weight Regulation Systems in Humans and Laboratory Animals. Int. J. Mol. Sci. 2023, 24, 16567. [Google Scholar] [CrossRef]
- Tokui, T.; Kawakita, T.; Yanagihara, R.; Kamada, S.; Minato, S.; Takeda, A.; Imaizumi, J.; Yamamoto, Y.; Yoshida, K.; Kato, T.; et al. Effects of Gonadal Status and the Estrogen Milieu on Hypothalamic Oxytocin Gene Expression and Serum Oxytocin Levels in Female Rats. Horm. Behav. 2021, 133, 105005. [Google Scholar] [CrossRef]
- Tachibana, A.; Yamamoto, Y.; Noguchi, H.; Takeda, A.; Tamura, K.; Aoki, H.; Minato, S.; Uchishiba, M.; Yamamoto, S.; Kamada, S.; et al. Changes in Serum Oxytocin Levels under Physiological and Supraphysiological Gonadal Steroid Hormone Conditions in Women of Reproductive Age: A Preliminary Study. Nutrients 2022, 14, 5350. [Google Scholar] [CrossRef] [PubMed]
- De Vries, G.J.; Wang, Z.; Bullock, N.A.; Numan, S. Sex Differences in the Effects of Testosterone and Its Metabolites on Vasopressin Messenger RNA Levels in the Bed Nucleus of the Stria Terminalis of Rats. J. Neurosci. 1994, 14, 1789–1794. [Google Scholar] [CrossRef] [PubMed]
- Le, T.Y.L.; Ashton, A.W.; Mardini, M.; Stanton, P.G.; Funder, J.W.; Handelsman, D.J.; Mihailidou, A.S. Role of Androgens in Sex Differences in Cardiac Damage During Myocardial Infarction. Endocrinology 2014, 155, 568–575. [Google Scholar] [CrossRef]
- Asarian, L.; Geary, N. Sex Differences in the Physiology of Eating. Am. J. Physiol. -Regul. Integr. Comp. Physiol. 2013, 305, R1215–R1267. [Google Scholar] [CrossRef] [PubMed]
- Blaustein, J.D.; Wade, G.N. Ovarian Influences on the Meal Patterns of Female Rats. Physiol. Behav. 1976, 17, 201–208. [Google Scholar] [CrossRef]
- Heine, P.A.; Taylor, J.A.; Iwamoto, G.A.; Lubahn, D.B.; Cooke, P.S. Increased Adipose Tissue in Male and Female Estrogen Receptor-α Knockout Mice. Proc. Natl. Acad. Sci. USA 2000, 97, 12729–12734. [Google Scholar] [CrossRef] [PubMed]
- Okura, T.; Koda, M.; Ando, F.; Niino, N.; Ohta, S.; Shimokata, H. Association of Polymorphisms in the Estrogen Receptor α Gene with Body Fat Distribution. Int. J. Obes. 2003, 27, 1020–1027. [Google Scholar] [CrossRef]
- Gao, Q.; Mezei, G.; Nie, Y.; Rao, Y.; Choi, C.S.; Bechmann, I.; Leranth, C.; Toran-Allerand, D.; A Priest, C.; Roberts, J.L.; et al. Anorectic Estrogen Mimics Leptin’s Effect on the Rewiring of Melanocortin Cells and Stat3 Signaling in Obese Animals. Nat. Med. 2007, 13, 89–94. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arata, M.; Tamura, K.; Aoki, H.; Noguchi, H.; Takeda, A.; Minato, S.; Yamamoto, S.; Kinouchi, R.; Yoshida, K.; Yamamoto, Y.; et al. The Effects of Testosterone on Hypothalamic and Serum Oxytocin Levels Are Affected by the Estrogen Milieu in Female Rats. Nutrients 2024, 16, 2533. https://doi.org/10.3390/nu16152533
Arata M, Tamura K, Aoki H, Noguchi H, Takeda A, Minato S, Yamamoto S, Kinouchi R, Yoshida K, Yamamoto Y, et al. The Effects of Testosterone on Hypothalamic and Serum Oxytocin Levels Are Affected by the Estrogen Milieu in Female Rats. Nutrients. 2024; 16(15):2533. https://doi.org/10.3390/nu16152533
Chicago/Turabian StyleArata, Moeka, Kou Tamura, Hidenori Aoki, Hiroki Noguchi, Asuka Takeda, Saki Minato, Shota Yamamoto, Riyo Kinouchi, Kanako Yoshida, Yuri Yamamoto, and et al. 2024. "The Effects of Testosterone on Hypothalamic and Serum Oxytocin Levels Are Affected by the Estrogen Milieu in Female Rats" Nutrients 16, no. 15: 2533. https://doi.org/10.3390/nu16152533