

Parecoxib Enhances Resveratrol against Human Colorectal Cancer Cells through Akt and TXNDC5 Inhibition and MAPK Regulation

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Chemicals
2.2. Cell Culture
2.3. MTT Assay for Cell Viability
2.4. Isobologram Analysis for Synergistic Anti-Cancer Effect
2.5. Overexpression of TXNDC5 in Cancer Cells
2.6. DAPI Staining for Chromatin Condensation and Fragmented Nucleus
2.7. Western Blotting
2.8. Intracellular ROS Analysis
2.9. Statistical Analysis
3. Results
3.1. Parecoxib Enhances Resveratrol to Inhibit Cell Viability in DLD-1 Cells
3.2. Parecoxib Enhances Resveratrol to Induce Apoptosis in DLD-1 Cells
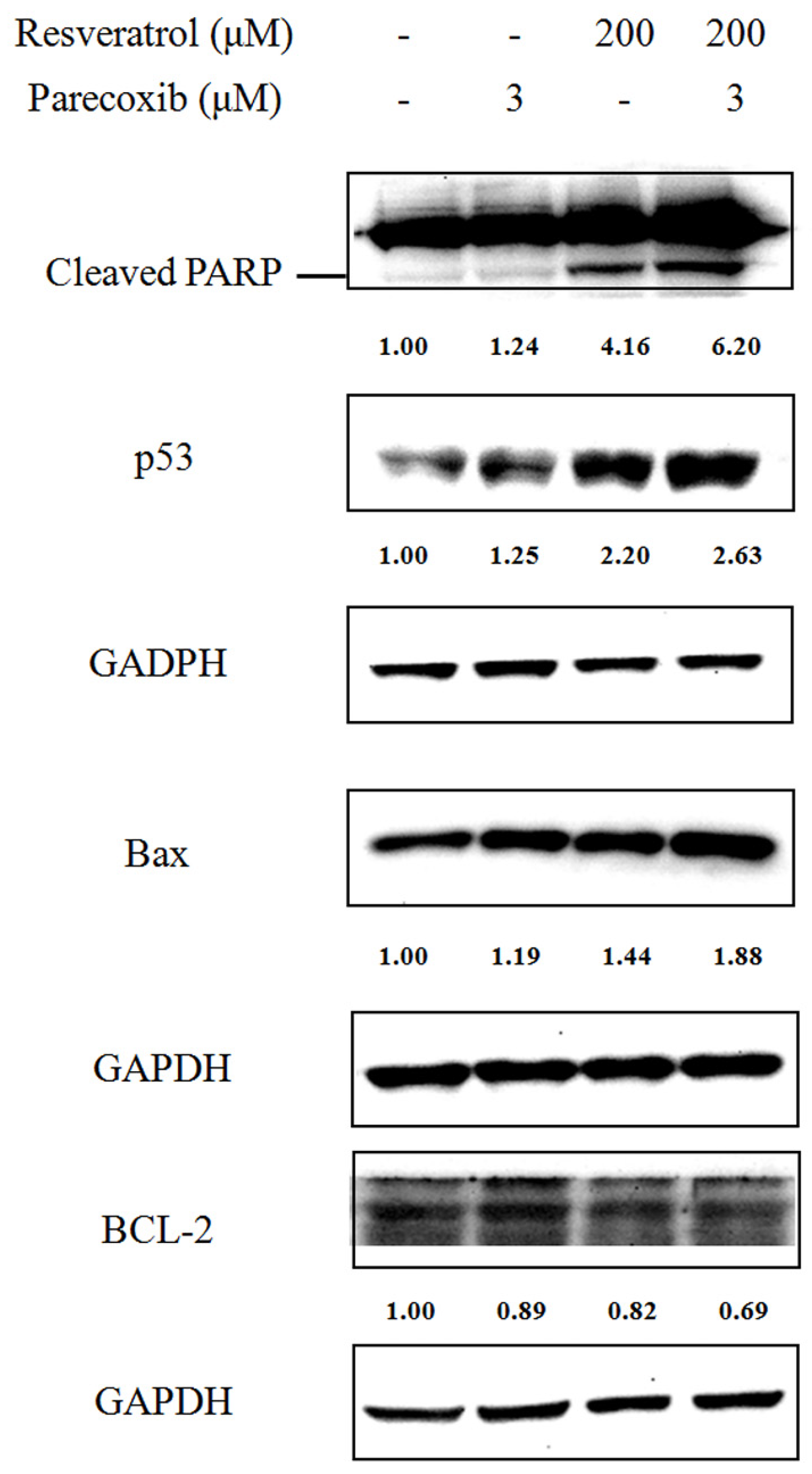
3.3. Parecoxib Enhances Resveratrol to Induce Apoptotic Proteins in DLD-1 Cells
3.4. Role of the PI3K/Akt Signaling Pathway in the Combination Effects of Parecoxib and Resveratrol
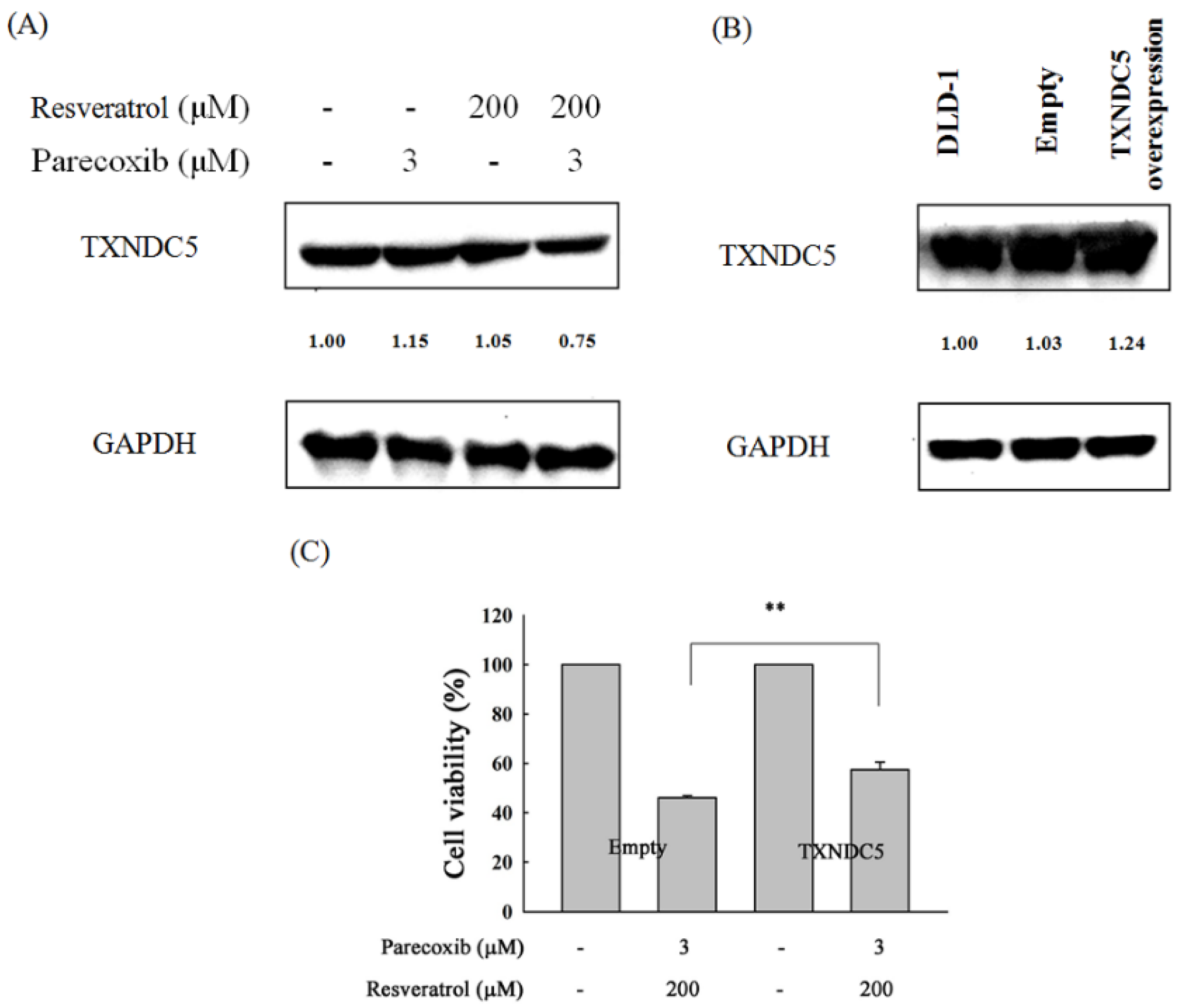
3.5. Role of TXNDC5 in the Combination Effects of Parecoxib and Resveratrol
3.6. Apoptotic Role of TXNDC5 in the Combination Effects of Parecoxib and Resveratrol
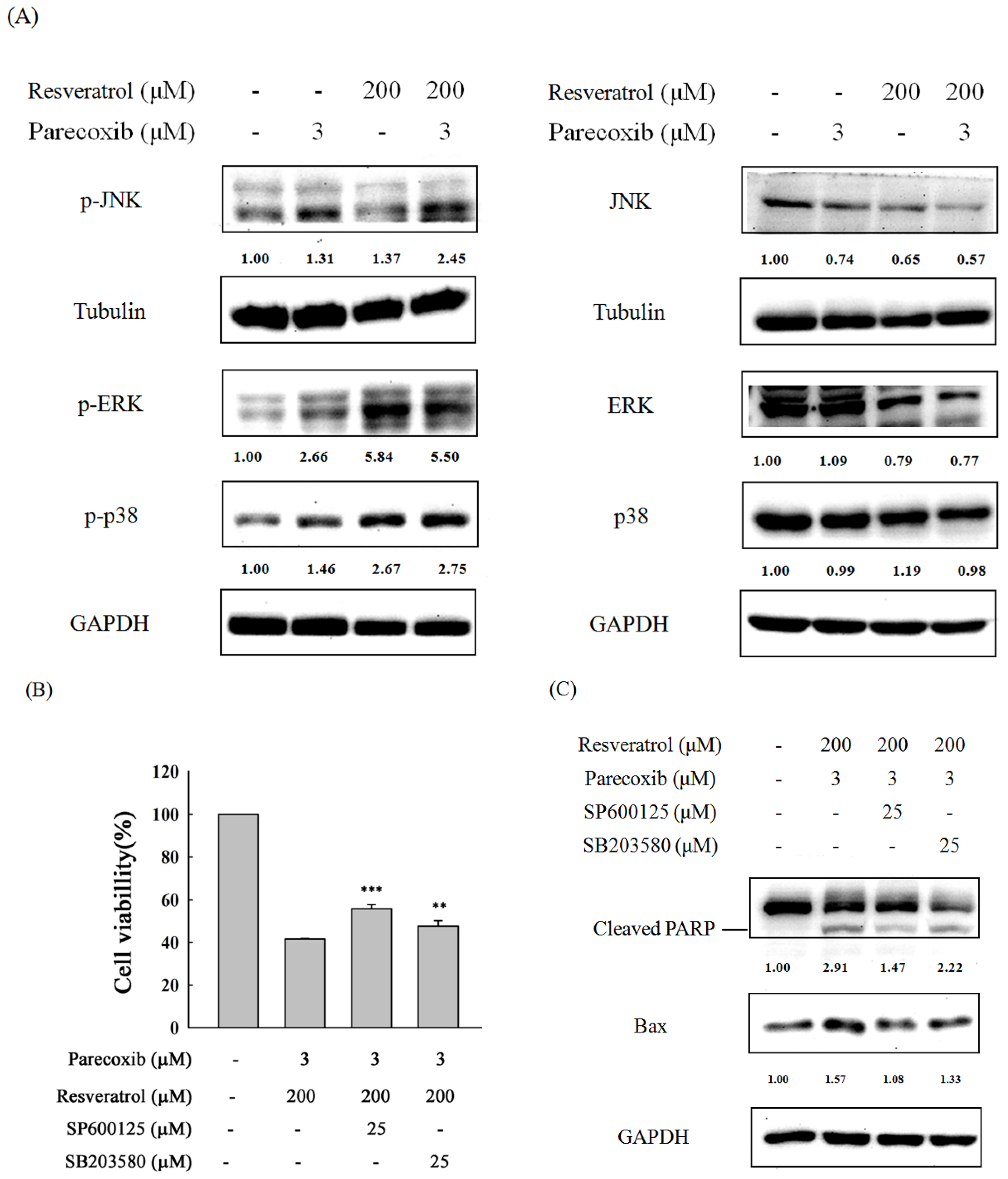
3.7. Role of MAPK Signaling in the Combination Effects of Parecoxib and Resveratrol
3.8. Effects of Parecoxib and Resveratrol on Intracellular ROS in DLD-1 Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ye, Y.; Ma, Y.; Kong, M.; Wang, Z.; Sun, K.; Li, F. Effects of dietary phytochemicals on DNA Damage in cancer cells. Nutr. Cancer 2023, 75, 761–775. [Google Scholar] [CrossRef] [PubMed]
- Tsai, M.C.; Chen, C.C.; Tseng, T.H.; Chang, Y.C.; Lin, Y.J.; Tsai, I.N.; Wang, C.C.; Wang, C.J. Hibiscus anthocyanins extracts induce apoptosis by activating AMP-activated protein kinase in human colorectal cancer cells. Nutrients 2023, 15, 3972. [Google Scholar] [CrossRef] [PubMed]
- Yao, C.J.; Chang, C.L.; Hu, M.H.; Liao, C.H.; Lai, G.M.; Chiou, T.J.; Ho, H.L.; Kio, H.C.; Yang, Y.Y.; Wang-Peng, J.; et al. Drastic synergy of lovastatin and Antrodia camphorata extract combination against PC3 androgen-refractory prostatecancer cells, accompanied by AXL and stemness molecules inhibition. Nutrients 2023, 15, 4493. [Google Scholar] [CrossRef] [PubMed]
- Weiskirchen, S.; Weiskirchen, R. Resveratrol: How much wine do you have to drink to stay healthy? Adv. Nutr. 2016, 7, 706–718. [Google Scholar] [CrossRef]
- Xu, Y.; Fang, M.; Li, X.; Wang, D.; Yu, L.; Ma, F.; Jiang, J.; Zhang, L.; Li, P. Contributions of common foods to resveratrol intake in the Chinese diet. Foods 2024, 13, 1267. [Google Scholar] [CrossRef]
- Walle, T. Bioavailability of resveratrol. Ann. N. Y. Acad. Sci. 2011, 1215, 9–15. [Google Scholar] [CrossRef]
- Li, Y.; Shin, Y.G.; Yu, C.; Kosmeder, J.W.; Hirschelman, W.H.; Pezzuto, J.M.; van Breemen, R.B. Increasing the throughput and productivity of Caco-2 cell permeability assays using liquid chromatography-mass spectrometry: Application to resveratrol absorption and metabolism. Comb. Chem. High. T Scr. 2003, 6, 757–767. [Google Scholar] [CrossRef]
- Fiod Riccio, B.V.; Fonseca-Santos, B.; Colerato Ferrari, P.; Chorilli, M. Characteristics, Biological properties and analytical methods of trans-resveratrol: A review. Crit. Rev. Anal. Chem. 2020, 50, 339–358. [Google Scholar] [CrossRef]
- Langcake, P.; Pryce, R.J. The production of resveratrol by Vitis vinifera and other members of the Vitaceae as a response to infection or injury. Plant Pathol. 1976, 9, 77–86. [Google Scholar] [CrossRef]
- Nonomura, S.; Kanagawa, H.; Makimoto, A. Chemical constituents of polygonaceous plants. I. studies on the components of ko-jo-kon. (Polygonum cuspidatum Sieb. et Zucc). Yakugaku Zasshi 1963, 83, 988–990. [Google Scholar] [CrossRef]
- Jin, Y.; Liu, X.; Liang, X.; Liu, J.; Liu, J.; Han, Z.; Lu, Q.; Wang, K.; Meng, B.; Zhang, C.; et al. Resveratrol rescues cutaneous radiation-induced DNA damage via a novel AMPK/SIRT7/HMGB1 regulatory axis. Cell Death Dis. 2023, 13, 847. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, M.; Ogasawara, Y.; Hayashi, H.; Inoue, K.; Sakashita, H. Resveratrol inhibits proliferation and induces autophagy by blocking SREBP1 expression in oral cancer cells. Molecules. 2022, 27, 8250. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.Y.; Im, E.; Kim, N.D. Mechanism of resveratrol-induced programmed cell death and new drug discovery against cancer: A Review. Int. J. Mol. Sci. 2022, 23, 13689. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Fan, Y.; Zhang, Y.; Liu, Y.; Yu, Y.; Ma, M. Resveratrol induces autophagy and apoptosis in non-small-cell lung cancer cells by activating the NGFR-AMPK-mTOR pathway. Nutrients 2022, 14, 2413. [Google Scholar] [CrossRef]
- Wang, J.; Huang, P.; Pan, X.; Xia, C.; Zhang, H.; Zhao, H.; Yuan, Z.; Liu, J.; Meng, C.; Liu, F. Resveratrol reverses TGF-β1-mediated invasion and metastasis of breast cancer cells via the SIRT3/AMPK/autophagy signal axis. Phytother. Res. 2023, 37, 211–230. [Google Scholar] [CrossRef]
- Ji, Q.; Liu, X.; Han, Z.; Zhou, L.; Sui, H.; Yan, L.; Jiang, H.; Ren, J.; Cai, J.; Li, Q. Resveratrol suppresses epithelial-to-mesenchymal transition in colorectal cancer through TGF-β1/Smads signaling pathway mediated Snail/E-cadherin expression. BMC Cancer 2015, 15, 97. [Google Scholar] [CrossRef]
- Duan, J.J.; Yue, W.; E, J.Y.; Malhotra, J.; Lu, S.; Gu, J.; Xu, F.; Tan, X.-L. In vitro comparative studies of resveratrol and triacetylresveratrol on cell proliferation, apoptosis, and STAT3 and NFκB signaling in pancertic cancer cells. Sci. Rep. 2016, 6, 31672. [Google Scholar] [CrossRef]
- Jackson, L.M.; Wu, K.C.; Mahida, Y.R.; Jenkins, D.; Hawkey, C.J. Cyclooxygenase (COX) 1 and 2 in normal, inflamed, and ulcerated human gastric mucosa. Gut 2000, 47, 762–770. [Google Scholar] [CrossRef]
- Lehmann, F.S.; Beglinger, C. Impact of COX-2 inhibitors in common clinical practice a gastroenterologist’s perspective. Curr. Top. Med. Chem. 2005, 5, 449–464. [Google Scholar] [CrossRef]
- Stiller, C.O.; Hjemdahl, P. Lessons from 20 years with COX-2 inhibitors: Importance of dose-response considerations and fair play in comparative trials. J. Intern. Med. 2022, 292, 557–574. [Google Scholar] [CrossRef]
- Kellett, E.; Berman, R.; Morgan, H.; Collins, J. Parecoxib for opioid-induced hyperalgesia. BMJ Support. Palliat. Care. 2021, 11, 126–127. [Google Scholar] [CrossRef]
- Chiu, S.C.; Livneh, H.; Chen, J.C.; Chang, C.M.; Hsu, H.; Chiang, T.I.; Tsai, T.Y. Parecoxib reduced postsurgical pain and facilitated movement more than patient controlled analgesia. Front. Surg. 2022, 9, 799795. [Google Scholar] [CrossRef] [PubMed]
- Ge, Z.; Li, M.; Chen, Y.; Sun, Y.; Zhang, R.; Zhang, J.; Bai, X.; Zhang, Y.; Chen, Q. The efficacy and safety of parecoxib multimodal preemptive analgesia in artificial joint replacement: A systematic review and meta-analysis of randomized controlled trials. Pain Ther. 2023, 12, 1065–1078. [Google Scholar] [CrossRef]
- Gao, J.; Zhao, G.; Wang, P.; Li, S.; Wang, X.; Zhao, H. Parecoxib protects against myocardial ischemia/reperfusion via targeting PKA-CREB signaling pathway. Panminerva Med. 2022, 64, 587–588. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.H.; Chang, W.L.; Lu, F.J.; Liu, Y.W.; Peng, J.Y.; Chen, C.H. Parecoxib expresses anti-metastasis effect through inhibition of epithelial-mesenchymal transition and the Wnt/β-catenin signaling pathway in human colon cancer DLD-1 cell line. Environ. Toxicol. 2022, 37, 2718–2727. [Google Scholar] [CrossRef] [PubMed]
- Li, L.Y.; Xiao, J.; Liu, Q.; Xia, K. Parecoxib inhibits glioblastoma cell proliferation, migration and invasion by upregulating miRNA-29c. Biol. Open. 2017, 6, 311–316. [Google Scholar] [CrossRef]
- Xu, S.; Li, X.; Li, W.; Ma, N.; Ma, H.; Cui, J.; You, X.; Chen, X. Sufentanil combined with parecoxib sodium inhibits proliferation and metastasis of HER2-positive breast cancer cells and regulates epithelial-mesenchymal transition. Clin. Exp. Metastasis. 2023, 40, 149–160. [Google Scholar] [CrossRef]
- Chang, W.L.; Peng, J.Y.; Hong, C.L.; Li, P.C.; Lu, F.J.; Chen, C.H. Parecoxib and 5-fluorouracil synergistically inhibit EMT and subsequent metastasis in colorectal cancer by targeting PI3K/Akt/NF-κB signaling. Biomedicines 2024, 12, 1526. [Google Scholar] [CrossRef]
- Horna-Terrón, E.; Pradilla-Dieste, A.; Sanchez-de-Diego, C.; Osada, J. TXNDC5, a newly discovered disulfide isomerase with a key role in cell physiology and pathology. Int. J. Mol. Sci. 2014, 15, 23501–23518. [Google Scholar] [CrossRef]
- Sullivan, D.C.; Huminiecki, L.; Moore, J.W.; Boyle, J.J.; Poulsom, R.; Creamer, D.; Barker, J.; Bicknell, R. EndoPDI. A novel protein-disulfide isomerase-like protein that is preferentially expressed in endothelial cells acts as a stress survival factor. J. Biol. Chem. 2003, 278, 47079–47088. [Google Scholar] [CrossRef]
- Chang, X.; Xu, B.; Wang, L.; Wang, Y.; Wang, Y.; Yan, S. Investigating a pathogenic role for TXNDC5 in tumors. Int. J. Oncol. 2013, 43, 1871–1884. [Google Scholar] [CrossRef]
- Tan, F.; Zhu, H.; He, X.; Yu, N.; Zhang, X.; Xu, H.; Pei, H. Role of TXNDC5 in tumorigenesis of colorectal cancer cells: In vivo and in vitro evidence. Int. J. Mol. Med. 2018, 42, 935–945. [Google Scholar] [CrossRef]
- Cheaib, B.; Auguste, A.; Leary, A. The PI3K/Akt/mTOR pathway in ovarian cancer: Therapeutic opportunities and challenges. R Chin. J. Cancer 2015, 34, 4–16. [Google Scholar] [CrossRef] [PubMed]
- Colakoglu, T.; Yildirim, S.; Kayaselcuk, F.; Nursal, T.Z.; Ezer, A.; Noyan, T.; Karakayali, H.; Haberal, M. Clinicopathological significance of PTEN loss and the phosphoinositide 3-kinase/Akt pathway in sporadic colorectal neoplasms: Is PTEN loss predictor of local recurrence? Am. J. Surg. 2008, 195, 719–725. [Google Scholar] [CrossRef]
- Pal, I.; Mandal, M. PI3K and Akt as molecular targets for cancer therapy: Current clinical outcomes. Acta Pharmacol. Sin. 2012, 33, 1441–1458. [Google Scholar] [CrossRef] [PubMed]
- Dhillon, A.S.; Hagan, S.; Rath, O.; Kolch, W. MAP kinase signalling pathways in cancer. Oncogene 2007, 26, 3279–3290. [Google Scholar] [CrossRef]
- Hankittichai, P.; Thaklaewphan, P.; Wikan, N.; Ruttanapattanakul, J.; Potikanond, S.; Smith, D.R.; Nimlamool, W. Resveratrol enhances cytotoxic effects of cisplatin by inducing cell cycle arrest and apoptosis in ovarian adenocarcinoma SKOV-3 cells through activating the p38 MAPK and suppressing AKT. Pharmaceuticals 2023, 16, 755. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.Y.; Pei, L.; Liu, Q.; Chen, S.; Dou, H.; Shu, G.; Yuan, Z.X.; Lin, J.; Peng, G.; Zhang, W.; et al. Isobologram analysis: A comprehensive review of nethodology and current research. Front. Pharmacol. 2019, 10, 1222. [Google Scholar] [CrossRef]
- Wu, Z.; Zhang, L.; Li, N.; Sha, L.; Zhang, K. An immunohistochemical study of thioredoxin domain-containing 5 expression in gastric adenocarcinoma. Oncol. Lett. 2015, 9, 1154–1158. [Google Scholar] [CrossRef]
- Peng, F.; Zhang, H.; Du, Y.; Tan, P. Cetuximab enhances cisplatin-induced endoplasmic reticulum stress-associated apoptosis in laryngeal squamous cell carcinoma cells by inhibiting expression of TXNDC5. Mol. Med. Rep. 2018, 17, 4767–4776. [Google Scholar] [CrossRef]
- Vincent, E.E.; Elder, D.J.; Phillips, L.; Heesom, K.J.; Pawade, J.; Luckett, M.; Sohail, M.; May, M.T.; Hetzel, M.R.; Tavaré, J.M. Overexpression of the TXNDC5 protein in non-small cell lung carcinoma. Anticancer Res. 2011, 31, 1577–1582. [Google Scholar]
- Lee, S.O.; Jin, U.H.; Kang, J.H.; Kim, S.B.; Guthrie, A.S.; Sreevalsan, S.; Lee, J.S.; Safe, S. The orphan nuclear receptor NR4A1 (Nur77) regulates oxidative and endoplasmic reticulum stress in pancreatic cancer cells. Mol. Cancer Res. 2014, 12, 527–538. [Google Scholar] [CrossRef]
- Zang, H.; Li, Y.; Zhang, X.; Huang, G. Circ_0000517 contributes to hepatocellular carcinoma progression by upregulating TXNDC5 via sponging miR-1296–5p. Cancer Manag. Res. 2020, 12, 3457–3468. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Song, G.; Chang, X.; Tan, W.; Pan, J.; Zhu, X.; Liu, Z.; Qi, M.; Yu, J.; Han, B. The role of TXNDC5 in castration-resistant prostate cancer—Involvement of androgen receptor signaling pathway. Oncogene 2014, 34, 4735–4745. [Google Scholar] [CrossRef]
- Mo, R.; Peng, J.; Xiao, J.; Ma, J.; Li, W.; Wang, J.; Ruan, Y.; Ma, S.; Hong, Y.; Wang, C.; et al. High TXNDC5 expression predicts poor prognosis in renal cell carcinoma. Tumor Biol. 2016, 37, 9797–9806. [Google Scholar] [CrossRef]
- Brockmueller, A.; Buhrmann, C.; Moravejolahkami, A.R.; Shakibaei, M. Resveratrol and p53: How are they involved in CRC plasticity and apoptosis? J. Adv. Res. 2024, 1232, 5. [Google Scholar] [CrossRef]
- Sangawa, A.; Shintani, M.; Yamao, N.; Kamoshida, S. Phosphorylation status of Akt and caspase-9 in gastric and colorectal carcinomas. Int. J. Clin. Exp. Pathol. 2014, 7, 3312–3317. [Google Scholar]
- Shen, B.; Chen, H.B.; Zhou, H.G.; Wu, M.H. Celastrol induces caspase-dependent apoptosis of hepatocellular carcinoma cells by suppression of mammalian target of rapamycin. J. Tradit. Chin. Med. 2021, 41, 381–389. [Google Scholar] [CrossRef]
- Li, D.; Wang, G.; Jin, G.; Yao, K.; Zhao, Z.; Bie, L.; Guo, Y.; Li, N.; Deng, W.; Chen, X.; et al. Resveratrol suppresses colon cancer growth by targeting the AKT/STAT3 signaling pathway. Int. J. Mol. Med. 2019, 43, 630–640. [Google Scholar] [CrossRef] [PubMed]
- Mohapatra, P.; Preet, R.; Choudhuri, M.; Choudhuri, T.; Kundu, C.N. 5-fluorouracil increases the chemopreventive potentials of resveratrol through DNA damage and MAPK signaling pathway in human colorectal cancer cells. Oncol. Res. 2011, 19, 311–321. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Xiong, M.; Xu, C.; Duan, L.; Dong, Y.; Luo, Y.; Niu, T.; Lu, C. Resveratrol induces apoptosis of human chronic myelogenous leukemia cells in vitro through p38 and JNK-regulated H2AX phosphorylation. Acta Pharmacol. Sin. 2015, 36, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Jia, Z.; Zhang, H.; Ma, C.; Li, N.; Wang, M. Celecoxib enhances apoptosis of the liver cancer cells via regulating ERK/JNK/P38 pathway. J. BUON. 2021, 26, 875–881. [Google Scholar] [PubMed]
- Wu, J.; Wang, D.; Zhou, J.; Li, J.; Xie, R.; Li, Y.; Huang, J.; Liu, B.; Qiu, J. Gambogenic acid induces apoptosis and autophagy through ROS-mediated endoplasmic reticulum stress via JNK pathway in prostate cancer cells. Phytother. Res. 2023, 37, 310–328. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Yu, K.; Hu, Y.; Su, F.; Gao, Z.; Hu, T.; Yang, Y.; Cao, X.; Qian, F. Schisantherin A induces cell apoptosis through ROS/JNK signaling pathway in human gastric cancer cells. Biochem. Pharmacol. 2020, 173, 113673. [Google Scholar] [CrossRef]
- Puckett, D.L.; Alquraishi, M.; Alani, D.; Chahed, S.; Donohoe, D.; Voy, B.; Whelan, J.; Bettaieb, A. Zyflamend induces apoptosis in pancreatic cancer cells via modulation of the JNK pathway. Cell Commun. Signal. 2020, 18, 126. [Google Scholar] [CrossRef]
- Woo, K.J.; Lee, T.J.; Lee, S.H.; Lee, J.M.; Seo, J.H.; Jeong, Y.J.; Park, J.W.; Kwon, T.K. Elevated gadd153/chop expression during resveratrol-induced apoptosis in human colon cancer cells. Biochem. Pharmacol. 2007, 73, 68–76. [Google Scholar] [CrossRef]
- Wang, X.; Li, H.; Chang, X. The role and mechanism of TXNDC5 in diseases. Eur. J. Med. Res. 2022, 27, 145. [Google Scholar] [CrossRef]
- De Amicis, F.; Adele Chimento, A.; Montalto, F.I.; Casaburi, I.; Rosa Sirianni, R.; Vincenzo Pezzi, V. Steroid receptor signallings as targets for resveratrol actions in breast and prostate cancer. Int. J. Mol. Sci. 2019, 20, 1087. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, W.-L.; Yang, K.-C.; Peng, J.-Y.; Hong, C.-L.; Li, P.-C.; Chye, S.M.; Lu, F.-J.; Shih, C.-W.; Chen, C.-H. Parecoxib Enhances Resveratrol against Human Colorectal Cancer Cells through Akt and TXNDC5 Inhibition and MAPK Regulation. Nutrients 2024, 16, 3020. https://doi.org/10.3390/nu16173020
Chang W-L, Yang K-C, Peng J-Y, Hong C-L, Li P-C, Chye SM, Lu F-J, Shih C-W, Chen C-H. Parecoxib Enhances Resveratrol against Human Colorectal Cancer Cells through Akt and TXNDC5 Inhibition and MAPK Regulation. Nutrients. 2024; 16(17):3020. https://doi.org/10.3390/nu16173020
Chicago/Turabian StyleChang, Wan-Ling, Kai-Chien Yang, Jyun-Yu Peng, Chain-Lang Hong, Pei-Ching Li, Soi Moi Chye, Fung-Jou Lu, Ching-Wei Shih, and Ching-Hsein Chen. 2024. "Parecoxib Enhances Resveratrol against Human Colorectal Cancer Cells through Akt and TXNDC5 Inhibition and MAPK Regulation" Nutrients 16, no. 17: 3020. https://doi.org/10.3390/nu16173020