
Lactoferrin: A Promising Therapeutic Molecule against Human Papillomavirus
 ,
,  ,
,  ,
,  , ,
, ,  and
and 
Abstract

1. Introduction
2. Lactoferrin: Structure and Biological Functions
3. Antiviral Effect of Lactoferrin
3.1. Lactoferrin against HPV
3.2. Lactoferrin against Other Viral Infections
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Sarıtaş, S.; Portocarrero, A.C.M.; Miranda López, J.M.; Lombardo, M.; Koch, W.; Raposo, A.; El-Seedi, H.R.; de Brito Alves, J.L.; Esatbeyoglu, T.; Karav, S.; et al. The Impact of Fermentation on the Antioxidant Activity of Food Products. Molecules 2024, 29, 3941. [Google Scholar] [CrossRef]
- Kaplan, M.; Baydemir, B.; Günar, B.B.; Arslan, A.; Duman, H.; Karav, S. Benefits of A2 Milk for Sports Nutrition, Health and Performance. Front. Nutr. 2022, 9, 935344. [Google Scholar] [CrossRef]
- Sorensen, M.; Sorensen, S.P.L. The Proteins in Whey. Compte Rendu Des Trav. Du Lab. De Carlsberg Ser. Chim. 1940, 23, 55–99. [Google Scholar]
- Vega-Bautista, A.; de la Garza, M.; Carrero, J.C.; Campos-Rodríguez, R.; Godínez-Victoria, M.; Drago-Serrano, M.E. The Impact of Lactoferrin on the Growth of Intestinal Inhabitant Bacteria. Int. J. Mol. Sci. 2019, 20, 4707. [Google Scholar] [CrossRef] [PubMed]
- Rascón-Cruz, Q.; Espinoza-Sánchez, E.A.; Siqueiros-Cendón, T.S.; Nakamura-Bencomo, S.I.; Arévalo-Gallegos, S.; Iglesias-Figueroa, B.F. Lactoferrin: A Glycoprotein Involved in Immunomodulation, Anticancer, and Antimicrobial Processes. Molecules 2021, 26, 205. [Google Scholar] [CrossRef]
- Lang, J.; Yang, N.; Deng, J.; Liu, K.; Yang, P.; Zhang, G.; Jiang, C. Inhibition of SARS Pseudovirus Cell Entry by Lactoferrin Binding to Heparan Sulfate Proteoglycans. PLoS ONE 2011, 6, e23710. [Google Scholar] [CrossRef]
- Frese, S.; Freeman-Sharkey, S.; Kyle, D.; Karav, S.; Duar, R.M. Nutritive Compositions with Bioactive Proteins. US20220273733A1, 2020. Available online: https://patents.google.com/patent/WO2021021765A1/en (accessed on 3 February 2024).
- Hu, Y.; Meng, X.; Zhang, F.; Xiang, Y.; Wang, J. The in Vitro Antiviral Activity of Lactoferrin against Common Human Coronaviruses and SARS-CoV-2 Is Mediated by Targeting the Heparan Sulfate Co-Receptor. Emerg. Microbes Infect. 2021, 10, 317–330. [Google Scholar] [CrossRef]
- Cumberbatch, M.; Dearman, R.J.; Uribe-luna, S.; Headon, D.R.; Ward, P.P.; Conneely, O.M.; Kimber, I. Regulation of Epidermal Langerhans Cell Migration by Lactoferrin. Immunology 2000, 100, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, C.E.M.; Cumberbatch, M.; Tucker, S.C.; Dearman, R.J.; Andrew, S.; Headon, D.R.; Kimber, I. Exogenous Topical Lactoferrin Inhibits Allergen-Induced Langerhans Cell Migration and Cutaneous Inflammation in Humans. Br. J. Dermatol. 2001, 144, 715–725. [Google Scholar] [CrossRef]
- Togawa, J.-I.; Nagase, H.; Tanaka, K.; Inamori, M.; Umezawa, T.; Nakajima, A.; Naito, M.; Sato, S.; Saito, T.; Sekihara, H. Lactoferrin Reduces Colitis in Rats via Modulation of the Immune System and Correction of Cytokine Imbalance. Am. J. Physiol. -Gastrointest. Liver Physiol. 2002, 283, G187–G195. [Google Scholar] [CrossRef]
- Legrand, D. Overview of Lactoferrin as a Natural Immune Modulator. J. Pediatr. 2016, 173, S10–S15. [Google Scholar] [CrossRef]
- Eker, F.; Akdaşçi, E.; Duman, H.; Yalçıntaş, Y.M.; Canbolat, A.A.; Kalkan, A.E.; Karav, S.; Šamec, D. Antimicrobial Properties of Colostrum and Milk. Antibiotics 2024, 13, 251. [Google Scholar] [CrossRef] [PubMed]
- Andersson, S.; Rylander, E.; Larsson, B.; Strand, A.; Silfversvärd, C.; Wilander, E. The Role of Human Papillomavirus in Cervical Adenocarcinoma Carcinogenesis. Eur. J. Cancer 2001, 37, 246–250. [Google Scholar] [CrossRef]
- de Sanjosé, S.; Diaz, M.; Castellsagué, X.; Clifford, G.; Bruni, L.; Muñoz, N.; Bosch, F.X. Worldwide Prevalence and Genotype Distribution of Cervical Human Papillomavirus DNA in Women with Normal Cytology: A Meta-Analysis. Lancet Infect. Dis. 2007, 7, 453–459. [Google Scholar] [CrossRef] [PubMed]
- Halec, G.; Schmitt, M.; Dondog, B.; Sharkhuu, E.; Wentzensen, N.; Gheit, T.; Tommasino, M.; Kommoss, F.; Bosch, F.X.; Franceschi, S.; et al. Biological Activity of Probable/Possible High-risk Human Papillomavirus Types in Cervical Cancer. Int. J. Cancer 2013, 132, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Wakabayashi, H.; Oda, H.; Yamauchi, K.; Abe, F. Lactoferrin for Prevention of Common Viral Infections. J. Infect. Chemother. 2014, 20, 666–671. [Google Scholar] [CrossRef] [PubMed]
- Puddu, P.; Borghi, P.; Gessani, S.; Valenti, P.; Belardelli, F.; Seganti, L. Antiviral Effect of Bovine Lactoferrin Saturated with Metal Ions on Early Steps of Human Immunodeficiency Virus Type 1 Infection. Int. J. Biochem. Cell Biol. 1998, 30, 1055–1063. [Google Scholar] [CrossRef]
- Li, W.Y.; Li, Q.W.; Han, Z.S.; Jiang, Z.L.; Yang, H.; Li, J.; Zhang, X.B. Growth Suppression Effects of Recombinant Adenovirus Expressing Human Lactoferrin on Cervical Cancer In Vitro and In Vivo. Cancer Biother. Radiopharm. 2011, 26, 477–483. [Google Scholar] [CrossRef]
- Zhang, Y.; Nicolau, A.; Lima, C.F.; Rodrigues, L.R. Bovine Lactoferrin Induces Cell Cycle Arrest and Inhibits Mtor Signaling in Breast Cancer Cells. Nutr. Cancer 2014, 66, 1371–1385. [Google Scholar] [CrossRef]
- Masson, P.L.; Heremans, J.F.; Dive, C.H. An Iron-Binding Protein Common to Many External Secretions. Clin. Chim. Acta 1966, 14, 735–739. [Google Scholar] [CrossRef]
- Vorland, L.H. Lactoferrin: A Multifunctional Glycoprotein. APMIS 1999, 107, 971–981. [Google Scholar] [CrossRef] [PubMed]
- Johanson, B. Isolation of an Iron-Containing Red Protein from Milk. Acta Chem. Scand. 1960, 14, 510–512. [Google Scholar] [CrossRef]
- Levay, P.F.; Viljoen, M. Lactoferrin: A General Review. Haematologica 1995, 80, 252–267. [Google Scholar] [PubMed]
- Legrand, D.; Pierce, A.; Elass, E.; Carpentier, M.; Mariller, C.; Mazurier, J. Lactoferrin Structure and Functions. In Bioactive Components of Milk; Bösze, Z., Ed.; Springer: New York, NY, USA, 2008; pp. 163–194. [Google Scholar] [CrossRef]
- Maciel, K.S.; Santos, L.S.; Bonomo, R.C.F.; Verissimo, L.A.A.; Minim, V.P.R.; Minim, L.A. Purification of Lactoferrin from Sweet Whey Using Ultrafiltration Followed by Expanded Bed Chromatography. Sep. Purif. Technol. 2020, 251, 117324. [Google Scholar] [CrossRef]
- Eker, F.; Bolat, E.; Pekdemir, B.; Duman, H.; Karav, S. Lactoferrin: Neuroprotection against Parkinson’s Disease and Secondary Molecule for Potential Treatment. Front. Aging Neurosci. 2023, 15, 1204149. [Google Scholar] [CrossRef]
- Baker, E.N.; Baker, H.M. Molecular Structure, Binding Properties and Dynamics of Lactoferrin. Cell. Mol. Life Sci. 2005, 62, 2531–2539. [Google Scholar] [CrossRef]
- MacGillivray, R.T.A.; Moore, S.A.; Chen, J.; Anderson, B.F.; Baker, H.; Luo, Y.; Bewley, M.; Smith, C.A.; Murphy, M.E.P.; Wang, Y.; et al. Two High-Resolution Crystal Structures of the Recombinant N-Lobe of Human Transferrin Reveal a Structural Change Implicated in Iron Release. Biochemistry 1998, 37, 7919–7928. [Google Scholar] [CrossRef] [PubMed]
- Baker, E.N. Structure and Reactivity of Transferrins. Adv. Inorg. Chem. 1994, 41, 389–463. [Google Scholar] [CrossRef]
- Baker, E.N.; Baker, H.M.; Kidd, R.D. Lactoferrin and Transferrin: Functional Variations on a Common Structural Framework. Biochem. Cell Biol. 2002, 80, 27–34. [Google Scholar] [CrossRef]
- Andreu, S.; Ripa, I.; Bello-Morales, R.; López-Guerrero, J.A. Liposomal Lactoferrin Exerts Antiviral Activity against HCoV-229E and SARS-CoV-2 Pseudoviruses In Vitro. Viruses 2023, 15, 972. [Google Scholar] [CrossRef] [PubMed]
- Hutchens, T.W.; Lönnerdal, B.; Rumball, S. Lactoferrin Structure and Function: Remaining Questions, Methodological Considerations and Future Directions. In Lactoferrin: Structure and Function; Hutchens, T.W., Rumball, S.V., Lönnerdal, B., Eds.; Springer: Boston, MA, USA, 1994; pp. 287–291. [Google Scholar] [CrossRef]
- Karav, S. Selective Deglycosylation of Lactoferrin to Understand Glycans’ Contribution to Antimicrobial Activity of Lactoferrin. Cell. Mol. Biol. 2018, 64, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Yalçıntaş, Y.M.; Duman, H.; Rocha, J.M.; Bartkiene, E.; Karav, S.; Ozogul, F. Role of Bovine Colostrum against Various Diseases. Food Biosci. 2024, 61, 104818. [Google Scholar] [CrossRef]
- Spadaro, M.; Caorsi, C.; Ceruti, P.; Varadhachary, A.; Forni, G.; Pericle, F.; Giovarelli, M. Lactoferrin, a Major Defense Protein of Innate Immunity, Is a Novel Maturation Factor for Human Dendritic Cells. FASEB J. 2008, 22, 2747–2757. [Google Scholar] [CrossRef]
- Siqueiros-Cendón, T.; Arévalo-Gallegos, S.; Iglesias-Figueroa, B.F.; García-Montoya, I.A.; Salazar-Martínez, J.; Rascón-Cruz, Q. Immunomodulatory Effects of Lactoferrin. Acta Pharmacol. Sin. 2014, 35, 557–566. [Google Scholar] [CrossRef]
- Perdijk, O.; Van Neerven, R.J.J.; Van den Brink, E.; Savelkoul, H.F.J.; Brugman, S. Bovine Lactoferrin Modulates Dendritic Cell Differentiation and Function. Nutrients 2018, 10, 848. [Google Scholar] [CrossRef]
- Di Biase, A.M.; Tinari, A.; Pietrantoni, A.; Antonini, G.; Valenti, P.; Conte, M.P.; Superti, F. Effect of Bovine Lactoferricin on Enteropathogenic Yersinia Adhesion and Invasion in HEp-2 Cells. J. Med. Microbiol. 2004, 53, 407–412. [Google Scholar] [CrossRef]
- Longhi, C.; Conte, M.; Seganti, L.; Polidoro, M.; Alfsen, A.; Valenti, P. Influence of Lactoferrin on the Entry Process of Escherichia Coli HB101(PRI203) in HeLa Cells. Med. Microbiol. Immunol. 1993, 182, 25–35. [Google Scholar] [CrossRef]
- Rose, J.E.; Meyer, D.H.; Fives-Taylor, P.M. Aae, an Autotransporter Involved in Adhesion of Actinobacillus actinomycetemcomitans to Epithelial Cells. Infect. Immun. 2003, 71, 2384–2393. [Google Scholar] [CrossRef]
- Valenti, P.; Antonini, G. Lactoferrin: An Important Host Defence against Microbial and Viral Attack. Cell. Mol. Life Sci. 2005, 62, 2576–2587. [Google Scholar] [CrossRef]
- Boyaka, P.N.; Sfeir, R.M.; Dubarry, M.; Rautureau, M.; Tomé, D. The Mode of Oral Bovine Lactoferrin Administration Influences Mucosal and Systemic Immune Responses in Mice. J. Nutr. 2004, 134, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Artym, J.; Zimecki, M.; Paprocka, M.; Kruzel, M.L. Orally Administered Lactoferrin Restores Humoral Immune Response in Immunocompromised Mice. Immunol. Lett. 2003, 89, 9–15. [Google Scholar] [CrossRef]
- Dhennin-Duthille, I.; Masson, M.; Damiens, E.; Fillebeen, C.; Spik, G.; Mazurier, J. Lactoferrin Upregulates the Expression of CD4 Antigen through the Stimulation of the Mitogen-Activated Protein Kinase in the Human Lymphoblastic T Jurkat Cell Line. J. Cell Biochem. 2000, 79, 583–593. [Google Scholar] [CrossRef] [PubMed]
- Zimecki, M.; Mazurier, J.; Machnicki, M.; Wieczorek, Z.; Montreuil, J.; Spik, G. Immunostimulatory Activity of Lactotransferrin and Maturation of CD4− CD8− Murine Thymocytes. Immunol. Lett. 1991, 30, 119–123. [Google Scholar] [CrossRef] [PubMed]
- Zimecki, M.; Mazurier, J.; Spik, G.; Kapp, J.A. Human Lactoferrin Induces Phenotypic and Functional Changes in Murine Splenic B Cells. Immunology 1995, 86, 122–127. [Google Scholar]
- Britigan, B.E.; Lewis, T.S.; Waldschmidt, M.; McCormick, M.L.; Krieg, A.M. Lactoferrin Binds CpG-Containing Oligonucleotides and Inhibits Their Immunostimulatory Effects on Human B Cells. J. Immunol. 2001, 167, 2921–2928. [Google Scholar] [CrossRef]
- Damiens, E.; Mazurier, J.; El Yazidi, I.; Masson, M.; Duthille, I.; Spik, G.; Boilly-Marer, Y. Effects of Human Lactoferrin on NK Cell Cytotoxicity against Haematopoietic and Epithelial Tumour Cells. Biochim. Et Biophys. Acta (BBA)-Mol. Cell Res. 1998, 1402, 277–287. [Google Scholar] [CrossRef]
- Fischer, R.; Debbabi, H.; Dubarry, M.; Boyaka, P.; Tomé, D. Regulation of Physiological and Pathological Th1 and Th2 Responses by LactoferrinThis Paper Is One of a Selection of Papers Published in This Special Issue, Entitled 7th International Conference on Lactoferrin: Structure, Function, and Applications, and Has Undergone the Journal’s Usual Peer Review Process. Biochem. Cell Biol. 2006, 84, 303–311. [Google Scholar] [CrossRef]
- Legrand, D.; Van Berkel, P.H.C.; Salmon, V.; Van Veen, A.H.; Slomianny, M.-C.; Nuijens, H.J.; Spik, G. The N-Terminal Arg2, Arg3 and Arg4 of Human Lactoferrin Interact with Sulphated Molecules but Not with the Receptor Present on Jurkat Human Lymphoblastic T-Cells. Biochem. J. 1997, 327, 841–846. [Google Scholar] [CrossRef]
- Curran, C.S.; Demick, K.P.; Mansfield, J.M. Lactoferrin Activates Macrophages via TLR4-Dependent and -Independent Signaling Pathways. Cell Immunol. 2006, 242, 23–30. [Google Scholar] [CrossRef]
- Rastogi, N.; Singh, A.; Pandey, S.N.; Sinha, M.; Bhushan, A.; Kaur, P.; Sharma, S.; Singh, T.P. Structure of the Iron-free True C-terminal Half of Bovine Lactoferrin Produced by Tryptic Digestion and Its Functional Significance in the Gut. FEBS J. 2014, 281, 2871–2882. [Google Scholar] [CrossRef] [PubMed]
- Teng, C.T. Factors Regulating Lactoferrin Gene ExpressionThis Paper Is One of a Selection of Papers Published in This Special Issue, Entitled 7th International Conference on Lactoferrin: Structure, Function, and Applications, and Has Undergone the Journal’s Usual Peer Review Process. Biochem. Cell Biol. 2006, 84, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Campione, E.; Lanna, C.; Cosio, T.; Rosa, L.; Conte, M.P.; Iacovelli, F.; Romeo, A.; Falconi, M.; Del Vecchio, C.; Franchin, E.; et al. Lactoferrin Against SARS-CoV-2: In Vitro and In Silico Evidences. Front. Pharmacol. 2021, 12, 666600. [Google Scholar] [CrossRef]
- Berlutti, F.; Pantanella, F.; Natalizi, T.; Frioni, A.; Paesano, R.; Polimeni, A.; Valenti, P. Antiviral Properties of Lactoferrin—A Natural Immunity Molecule. Molecules 2011, 16, 6992–7018. [Google Scholar] [CrossRef]
- Duman, H.; Karav, S. Bovine Colostrum and Its Potential Contributions for Treatment and Prevention of COVID-19. Front. Immunol. 2023, 14, 1214514. [Google Scholar] [CrossRef]
- Drobni, P.; Näslund, J.; Evander, M. Lactoferrin Inhibits Human Papillomavirus Binding and Uptake In Vitro. Antiviral Res. 2004, 64, 63–68. [Google Scholar] [CrossRef]
- Hasegawa, K.; Motsuchi, W.; Tanaka, S.; Dosako, S. Inhibition with Lactoferrin of In Vitro Infection with Human Herpes Virus. Jpn. J. Med. Sci. Biol. 1994, 47, 73–85. [Google Scholar] [CrossRef] [PubMed]
- Marchetti, M.; Trybala, E.; Superti, F.; Johansson, M.; Bergström, T. Inhibition of Herpes Simplex Virus Infection by Lactoferrin Is Dependent on Interference with the Virus Binding to Glycosaminoglycans. Virology 2004, 318, 405–413. [Google Scholar] [CrossRef]
- Hara, K. Lactoferrin Inhibits Hepatitis B Virus Infection in Cultured Human Hepatocytes. Hepatol. Res. 2002, 24, 228–235. [Google Scholar] [CrossRef]
- Redwan, E.M.; EL-Fakharany, E.M.; Uversky, V.N.; Linjawi, M.H. Screening the Anti Infectivity Potentials of Native N- and C-Lobes Derived from the Camel Lactoferrin against Hepatitis C Virus. BMC Complement. Altern. Med. 2014, 14, 219. [Google Scholar] [CrossRef]
- Groot, F.; Geijtenbeek, T.B.H.; Sanders, R.W.; Baldwin, C.E.; Sanchez-Hernandez, M.; Floris, R.; van Kooyk, Y.; de Jong, E.C.; Berkhout, B. Lactoferrin Prevents Dendritic Cell-Mediated Human Immunodeficiency Virus Type 1 Transmission by Blocking the DC-SIGN—Gp120 Interaction. J. Virol. 2005, 79, 3009–3015. [Google Scholar] [CrossRef] [PubMed]
- Bolat, E.; Eker, F.; Kaplan, M.; Duman, H.; Arslan, A.; Saritaş, S.; Şahutoğlu, A.S.; Karav, S. Lactoferrin for COVID-19 Prevention, Treatment, and Recovery. Front. Nutr. 2022, 9, 992733. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, M.; Şahutoğlu, A.S.; Sarıtaş, S.; Duman, H.; Arslan, A.; Pekdemir, B.; Karav, S. Role of Milk Glycome in Prevention, Treatment, and Recovery of COVID-19. Front. Nutr. 2022, 9, 1033779. [Google Scholar] [CrossRef]
- Santos, J.G.d.O.; Migueis, D.P.; Amaral, J.B.d.; Bachi, A.L.L.; Boggi, A.C.; Thamboo, A.; Voegels, R.L.; Pezato, R. Impact of SARS-CoV-2 on Saliva: TNF-⍺, IL-6, IL-10, Lactoferrin, Lysozyme, IgG, IgA, and IgM. J. Oral. Biosci. 2022, 64, 108–113. [Google Scholar] [CrossRef]
- Kaito, M.; Iwasa, M.; Fujita, N.; Kobayashi, Y.; Kojima, Y.; Ikoma, J.; Imoto, I.; Adachi, Y.; Hamano, H.; Yamauchi, K. Effect of Lactoferrin in Patients with Chronic Hepatitis C: Combination Therapy with Interferon and Ribavirin. J. Gastroenterol. Hepatol. 2007, 22, 1894–1897. [Google Scholar] [CrossRef]
- Hadidi, N.; Saffari, M.; Faizi, M. Optimized Transferosomal Bovine Lactoferrin (BLF) as a Promising Novel Non-Invasive Topical Treatment for Genital Warts Caused by Human Papiluma Virus (HPV). Iran. J. Pharm. Res. 2018, 17 (Suppl. S2), 12–23. [Google Scholar] [PubMed]
- Abbas, Z.H.; Doosh, K.S.; Yaseen, N.Y. Effect of Purified Goat Milk Lactoferrin on Cancer Cell Growth (AMN-3) in Vitro. Biochem. Cell Arch. 2019, 19, 2661–2667. [Google Scholar] [CrossRef]
- Gheit, T. Mucosal and Cutaneous Human Papillomavirus Infections and Cancer Biology. Front. Oncol. 2019, 9, 355. [Google Scholar] [CrossRef]
- Andrei, E.C.; Baniță, I.M.; Munteanu, M.C.; Busuioc, C.J.; Mateescu, G.O.; Mălin, R.D.; Pisoschi, C.G. Oral Papillomatosis: Its Relation with Human Papilloma Virus Infection and Local Immunity—An Update. Medicina 2022, 58, 1103. [Google Scholar] [CrossRef]
- Buck, C.B.; Pastrana, D.V.; Lowy, D.R.; Schiller, J.T. Efficient Intracellular Assembly of Papillomaviral Vectors. J. Virol. 2004, 78, 751–757. [Google Scholar] [CrossRef]
- Buck, C.B.; Cheng, N.; Thompson, C.D.; Lowy, D.R.; Steven, A.C.; Schiller, J.T.; Trus, B.L. Arrangement of L2 within the Papillomavirus Capsid. J. Virol. 2008, 82, 5190–5197. [Google Scholar] [CrossRef] [PubMed]
- Smotkin, D.; Wettstein, F.O. Transcription of Human Papillomavirus Type 16 Early Genes in a Cervical Cancer and a Cancer-Derived Cell Line and Identification of the E7 Protein. Proc. Natl. Acad. Sci. USA 1986, 83, 4680–4684. [Google Scholar] [CrossRef] [PubMed]
- Thierry, F.; Heard, J.M.; Dartmann, K.; Yaniv, M. Characterization of a Transcriptional Promoter of Human Papillomavirus 18 and Modulation of Its Expression by Simian Virus 40 and Adenovirus Early Antigens. J. Virol. 1987, 61, 134–142. [Google Scholar] [CrossRef]
- Pal, A.; Kundu, R. Human Papillomavirus E6 and E7: The Cervical Cancer Hallmarks and Targets for Therapy. Front. Microbiol. 2020, 10, 3116. [Google Scholar] [CrossRef]
- Becker, K.A.; Florin, L.; Sapp, C.; Sapp, M. Dissection of Human Papillomavirus Type 33 L2 Domains Involved in Nuclear Domains (ND) 10 Homing and Reorganization. Virology 2003, 314, 161–167. [Google Scholar] [CrossRef]
- Giroglou, T.; Florin, L.; Schäfer, F.; Streeck, R.E.; Sapp, M. Human Papillomavirus Infection Requires Cell Surface Heparan Sulfate. J. Virol. 2001, 75, 1565–1570. [Google Scholar] [CrossRef]
- Kumar, A.; Jacob, T.; Abban, C.Y.; Meneses, P.I. Intermediate Heparan Sulfate Binding During HPV-16 Infection in HaCaTs. Am. J. Ther. 2014, 21, 331–342. [Google Scholar] [CrossRef]
- Abban, C.Y.; Meneses, P.I. Usage of Heparan Sulfate, Integrins, and FAK in HPV16 Infection. Virology 2010, 403, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Porter, S.S.; Stepp, W.H.; Stamos, J.D.; McBride, A.A. Host Cell Restriction Factors That Limit Transcription and Replication of Human Papillomavirus. Virus Res. 2017, 231, 10–20. [Google Scholar] [CrossRef]
- Chiang, C.M.; Ustav, M.; Stenlund, A.; Ho, T.F.; Broker, T.R.; Chow, L.T. Viral E1 and E2 Proteins Support Replication of Homologous and Heterologous Papillomaviral Origins. Proc. Natl. Acad. Sci. USA 1992, 89, 5799–5803. [Google Scholar] [CrossRef]
- McKinney, C.C.; Kim, M.J.; Chen, D.; McBride, A.A. Brd4 Activates Early Viral Transcription upon Human Papillomavirus 18 Infection of Primary Keratinocytes. mBio 2016, 7, 10–1128. [Google Scholar] [CrossRef]
- McBride, A.A. The Papillomavirus E2 Proteins. Virology 2013, 445, 57–79. [Google Scholar] [CrossRef]
- Darnell, G.A.; Schroder, W.A.; Antalis, T.M.; Lambley, E.; Major, L.; Gardner, J.; Birrell, G.; Cid-Arregui, A.; Suhrbier, A. Human Papillomavirus E7 Requires the Protease Calpain to Degrade the Retinoblastoma Protein. J. Biol. Chem. 2007, 282, 37492–37500. [Google Scholar] [CrossRef] [PubMed]
- Yugis, A.R.; Saputra, A.; Noviana, R.; Pamungkas, J.; Suparto, I.H. Comparison of Methods for The Purification of Goat Lactoferrin and Antiviral Activity to Human Papillomavirus. Am. J. Microbiol. Res. 2015, 3, 171–175. [Google Scholar] [CrossRef]
- Mistry, N.; Drobni, P.; Näslund, J.; Sunkari, V.G.; Jenssen, H.; Evander, M. The Anti-Papillomavirus Activity of Human and Bovine Lactoferricin. Antiviral Res. 2007, 75, 258–265. [Google Scholar] [CrossRef]
- Luzi, C.; Brisdelli, F.; Iorio, R.; Bozzi, A.; Carnicelli, V.; Di Giulio, A.; Lizzi, A.R. Apoptotic Effects of Bovine Apo-Lactoferrin on HeLa Tumor Cells. Cell Biochem. Funct. 2017, 35, 33–41. [Google Scholar] [CrossRef]
- Zhang, X.; Du, Z.; Wang, N.; Meng, Y. Effects of Recombinant Adenovirus Vector Expressing Human Lactoferrin on Proliferation and Apoptosis of Cervical Cancer Stem-like Cells. Chin. J. Tissue Eng. Res. 2017, 21, 2009–2014. [Google Scholar] [CrossRef]
- Stragier, I.; Snoeck, R.; De Clercq, E.; Van den Oord, J.J.; Van Ranst, M.; De Greef, H. Local Treatment of HPV-induced Skin Lesions by Cidofovir. J. Med. Virol. 2002, 67, 241–245. [Google Scholar] [CrossRef] [PubMed]
- Fusconi, M.; Grasso, M.; Greco, A.; Gallo, A.; Campo, F.; Remacle, M.; Turchetta, R.; Pagliuca, G.; DE Vincentiis, M. Recurrent Respiratory Papillomatosis by HPV: Review of the Literature and Update on the Use of Cidofovir. Acta Otorhinolaryngol. Ital. 2014, 34, 375–381. [Google Scholar]
- Yang, J.; Pu, Y.; Zeng, Z.; Yu, Z.; Huang, N.; Deng, Q. Interferon for the Treatment of Genital Warts: A Systematic Review. BMC Infect. Dis. 2009, 9, 156. [Google Scholar] [CrossRef]
- Song, S.-H.; Lee, J.-K.; Lee, N.-W.; Saw, H.-S.; Kang, J.-S.; Lee, K.-W. Interferon-γ (IFN-γ): A Possible Prognostic Marker for Clearance of High-Risk Human Papillomavirus (HPV). Gynecol. Oncol. 2008, 108, 543–548. [Google Scholar] [CrossRef]
- Kenter, G.G.; Welters, M.J.P.; Valentijn, A.R.P.M.; Lowik, M.J.G.; Berends-van der Meer, D.M.A.; Vloon, A.P.G.; Essahsah, F.; Fathers, L.M.; Offringa, R.; Drijfhout, J.W.; et al. Vaccination against HPV-16 Oncoproteins for Vulvar Intraepithelial Neoplasia. N. Engl. J. Med. 2009, 361, 1838–1847. [Google Scholar] [CrossRef]
- Krzyzowska, M.; Janicka, M.; Tomaszewska, E.; Ranoszek-Soliwoda, K.; Celichowski, G.; Grobelny, J.; Szymanski, P. Lactoferrin-Conjugated Nanoparticles as New Antivirals. Pharmaceutics 2022, 14, 1862. [Google Scholar] [CrossRef] [PubMed]
- Krzyzowska, M.; Chodkowski, M.; Janicka, M.; Dmowska, D.; Tomaszewska, E.; Ranoszek-Soliwoda, K.; Bednarczyk, K.; Celichowski, G.; Grobelny, J. Lactoferrin-Functionalized Noble Metal Nanoparticles as New Antivirals for HSV-2 Infection. Microorganisms 2022, 10, 110. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Qin, H.; Guan, L.; Liu, K.; Hong, B.; Zhang, X.; Lou, F.; Li, M.; Lin, W.; Chen, Y.; et al. Bovine Lactoferrin Inhibits SARS-CoV-2 and SARS-CoV-1 by Targeting the RdRp Complex and Alleviates Viral Infection in the Hamster Model. J. Med. Virol. 2023, 95, e28281. [Google Scholar] [CrossRef]
- Kobayashi-Sakamoto, M.; Maeda, T.; Kimura, M.; Yusa, J.; Ito, H.; Tani, H.; Kato, Y.; Hirose, K. Bovine Lactoferrin Increases the Poly(I:C)-Induced Antiviral Response In Vitro. Biochem. Cell Biol. 2022, 100, 338–348. [Google Scholar] [CrossRef]
- Graikini, D.; Conesa, C.; Abad, I.; Pérez, M.D.; Sánchez, L. Evaluation of in Vitro Antirotaviral Activity of Lactoferrin from Different Species Using a Human Intestinal Model. Int. Dairy J. 2024, 149, 105818. [Google Scholar] [CrossRef]
- Zhou, H.; Zhu, Y.; Liu, N.; Zhang, W.; Han, J. Effect of Iron Saturation of Bovine Lactoferrin on the Inhibition of Hepatitis B Virus In Vitro. PeerJ 2024, 12, e17302. [Google Scholar] [CrossRef]
- Mann, J.K.; Reddy, T.; van der Stok, M.; Ngubane, A.; Mulaudzi, T.; Mchunu, N.; Nevhungoni, P.; Manickchund, N.; Manickchund, P.; Louise Cairns, C.H.; et al. Hen Egg White Bovine Colostrum Supplement Reduces Symptoms of Mild/Moderate COVID-19: A Randomized Control Trial. Future Sci. OA 2023, 9, FSO882. [Google Scholar] [CrossRef]
- Kobayashi-Sakamoto, M.; Maeda, T.; Yusa, J.; Shimada, T.; Tani, H.; Kato, Y.; Hirose, K. Bovine Lactoferrin Suppresses the Cathepsin-Dependent Pathway of SARS-CoV-2 Entry in Vitro. Int. Dairy J. 2024, 148, 105805. [Google Scholar] [CrossRef]
- Ramírez-Rico, G.; Ruiz-Mazón, L.; Reyes-López, M.; Rivillas Acevedo, L.; Serrano-Luna, J.; de la Garza, M. Apo-Lactoferrin Inhibits the Proteolytic Activity of the 110 KDa Zn Metalloprotease Produced by Mannheimia Haemolytica A2. Int. J. Mol. Sci. 2024, 25, 8232. [Google Scholar] [CrossRef]
- Abad, I.; Sangüesa, A.; Ubieto, M.; Carramiñana, J.J.; Pérez, M.D.; Buey, B.; Mesonero, J.E.; Grasa, L.; Sánchez, L. Protective Effect of Bovine Lactoferrin against Cronobacter Sakazakii in Human Intestinal Caco-2/TC7 Cells. Int. Dairy J. 2022, 133, 105428. [Google Scholar] [CrossRef]
- Wróbel, M.; Małaczewska, J.; Kaczorek-Łukowska, E. Antiviral Effect of Bovine Lactoferrin against Enterovirus E. Molecules 2022, 27, 5569. [Google Scholar] [CrossRef]
- Baram-Pinto, D.; Shukla, S.; Gedanken, A.; Sarid, R. Inhibition of HSV-1 Attachment, Entry, and Cell-to-Cell Spread by Functionalized Multivalent Gold Nanoparticles. Small 2010, 6, 1044–1050. [Google Scholar] [CrossRef] [PubMed]
- Lara, H.H.; Garza-Treviño, E.N.; Ixtepan-Turrent, L.; Singh, D.K. Silver Nanoparticles Are Broad-Spectrum Bactericidal and Virucidal Compounds. J. Nanobiotechnology 2011, 9, 30. [Google Scholar] [CrossRef] [PubMed]
- Johnston, H.J.; Hutchison, G.; Christensen, F.M.; Peters, S.; Hankin, S.; Stone, V. A Review of the in Vivo and in Vitro Toxicity of Silver and Gold Particulates: Particle Attributes and Biological Mechanisms Responsible for the Observed Toxicity. Crit. Rev. Toxicol. 2010, 40, 328–346. [Google Scholar] [CrossRef]
- Mironava, T.; Hadjiargyrou, M.; Simon, M.; Jurukovski, V.; Rafailovich, M.H. Gold Nanoparticles Cellular Toxicity and Recovery: Effect of Size, Concentration and Exposure Time. Nanotoxicology 2010, 4, 120–137. [Google Scholar] [CrossRef]
- Baram-Pinto, D.; Shukla, S.; Perkas, N.; Gedanken, A.; Sarid, R. Inhibition of Herpes Simplex Virus Type 1 Infection by Silver Nanoparticles Capped with Mercaptoethane Sulfonate. Bioconjug Chem. 2009, 20, 1497–1502. [Google Scholar] [CrossRef]
- Nayak, P.S.; Borah, S.M.; Gogoi, H.; Asthana, S.; Bhatnagar, R.; Jha, A.N.; Jha, S. Lactoferrin Adsorption onto Silver Nanoparticle Interface: Implications of Corona on Protein Conformation, Nanoparticle Cytotoxicity and the Formulation Adjuvanticity. Chem. Eng. J. 2019, 361, 470–484. [Google Scholar] [CrossRef]
- Kowdley, K.V. Iron, Hemochromatosis, and Hepatocellular Carcinoma. Gastroenterology 2004, 127, S79–S86. [Google Scholar] [CrossRef]
- Lepanto, M.S.; Rosa, L.; Paesano, R.; Valenti, P.; Cutone, A. Lactoferrin in Aseptic and Septic Inflammation. Molecules 2019, 24, 1323. [Google Scholar] [CrossRef] [PubMed]
- Rosa, L.; Cutone, A.; Lepanto, M.; Paesano, R.; Valenti, P. Lactoferrin: A Natural Glycoprotein Involved in Iron and Inflammatory Homeostasis. Int. J. Mol. Sci. 2017, 18, 1985. [Google Scholar] [CrossRef]
- Mancinelli, R.; Rosa, L.; Cutone, A.; Lepanto, M.S.; Franchitto, A.; Onori, P.; Gaudio, E.; Valenti, P. Viral Hepatitis and Iron Dysregulation: Molecular Pathways and the Role of Lactoferrin. Molecules 2020, 25, 1997. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.-H.; Wang, J.-Y.; Liu, P.-Y.; Sun, J.; Wang, X.-M.; Wu, R.-H.; He, X.-T.; Tu, Z.-K.; Wang, C.-G.; Xu, H.-Q.; et al. Iron Metabolism Disorders in Patients with Hepatitis B-Related Liver Diseases. World J. Clin. Cases 2018, 6, 600–610. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Dong, A.; Liu, G.; Anderson, G.J.; Hu, T.Y.; Shi, J.; Hu, Y.; Nie, G. Correlation of Serum Hepcidin Levels with Disease Progression in Hepatitis B Virus-Related Disease Assessed by Nanopore Film Based Assay. Sci. Rep. 2016, 6, 34252. [Google Scholar] [CrossRef]


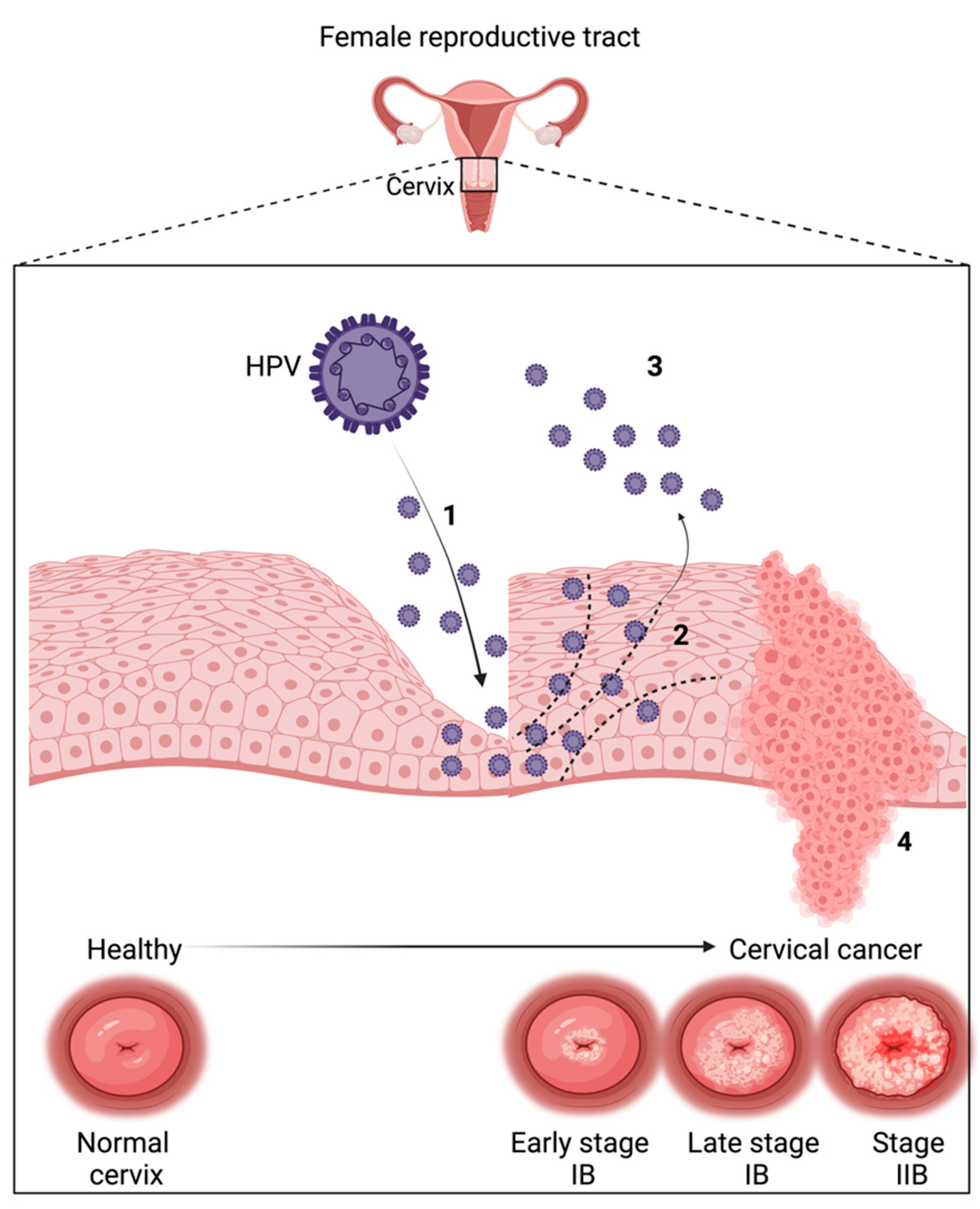

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Effect | Dairy Species | Study | Reference |
|---|---|---|---|
| Binding to HPV-16 virus-like particles and inhibits their internalization to HaCaT cells | Bovine | In vitro | [59] |
| Bovine lactoferrin derivative lactoferricin inhibits the infection of HPV-16 and HPV-5 in HaCaT and C33A cells | Bovine | In vitro | [88] |
| Improvement of genital warts caused by HPV via the system of transdermal delivery | Bovine | In vitro | [69] |
| Viral replication and infection inhibition in HeLa cells | Goat | In vitro | [87] |
| Stimulation of apoptosis in HeLa cells by triggering oxygen radicals and glutathione level | Bovine | In vitro | [89] |
| Anticancer activity and cytotoxicity effect on AMN3 and REF cells | Goat | In vitro | [70] |
| Triggering to increase the level of antiviral cytokines and chemokines in the vaginal tissue | Bovine | In vitro | [96] |
| The prevention of HSV-2 infection in AgNP and AuNP | Bovine | In vitro, in vivo | [97] |
| The prevention of SARS-CoV-2 infection by combining RBD and inhibiting coronaviruses’ RdRp activity | Bovine | In vitro | [98] |
| Increase in the antiviral response associated with the double-stranded RNA-stimulated signaling pathway | Bovine | In vitro | [99] |
| The neutralization of rotavirus infection in Caco-2/TV7 cells differentiated as human enterocytes | Bovine, camel | In vitro | [100] |
| Inhibition of hepatitis B virus DNA in HepG2 cells | Bovine | In vitro | [101] |
| Lessened symptoms in individuals with mild or moderate COVID-19 | Hen egg white bovine colostrum mixture | In vivo | [102] |
| CoV-2-Wtpv entry suppression into ACE2-expressing cells | Bovine | In vitro | [103] |
| Inhibition of activity and secretion of 110-Mh metalloprotease from M. haemolytica A2 | Bovine | In vitro | [104] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaplan, M.; Baktıroğlu, M.; Kalkan, A.E.; Canbolat, A.A.; Lombardo, M.; Raposo, A.; de Brito Alves, J.L.; Witkowska, A.M.; Karav, S. Lactoferrin: A Promising Therapeutic Molecule against Human Papillomavirus. Nutrients 2024, 16, 3073. https://doi.org/10.3390/nu16183073
Kaplan M, Baktıroğlu M, Kalkan AE, Canbolat AA, Lombardo M, Raposo A, de Brito Alves JL, Witkowska AM, Karav S. Lactoferrin: A Promising Therapeutic Molecule against Human Papillomavirus. Nutrients. 2024; 16(18):3073. https://doi.org/10.3390/nu16183073
Chicago/Turabian StyleKaplan, Merve, Merve Baktıroğlu, Arda Erkan Kalkan, Ahmet Alperen Canbolat, Mauro Lombardo, António Raposo, José Luiz de Brito Alves, Anna Maria Witkowska, and Sercan Karav. 2024. "Lactoferrin: A Promising Therapeutic Molecule against Human Papillomavirus" Nutrients 16, no. 18: 3073. https://doi.org/10.3390/nu16183073
APA StyleKaplan, M., Baktıroğlu, M., Kalkan, A. E., Canbolat, A. A., Lombardo, M., Raposo, A., de Brito Alves, J. L., Witkowska, A. M., & Karav, S. (2024). Lactoferrin: A Promising Therapeutic Molecule against Human Papillomavirus. Nutrients, 16(18), 3073. https://doi.org/10.3390/nu16183073