
Metabolomic Biomarkers of Dietary Approaches to Stop Hypertension (DASH) Dietary Patterns in Pregnant Women

 , , , ,
, , , ,  , ,
, , 

Abstract
1. Introduction
2. Methods
2.1. Study Population and Design
2.2. Assessment of Dietary Intakes and DASH Score
2.3. Biospecimen Collection, Metabolomic Profiling, and Data Pre-Treatment
2.4. Covariates
2.5. Statistical Methods
3. Results
3.1. Women’s Baseline Characteristics
3.2. Associations of Dietary DASH Score with Plasma Metabolites
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Appel, L.J.; Moore, T.J.; Obarzanek, E.; Vollmer, W.M.; Svetkey, L.P.; Sacks, F.M.; Bray, G.A.; Vogt, T.M.; Cutler, J.A.; Windhauser, M.M.; et al. A clinical trial of the effects of dietary patterns on blood pressure. DASH Collaborative Research Group. N. Engl. J. Med. 1997, 336, 1117–1124. [Google Scholar] [CrossRef]
- Nilsson, A.; Halvardsson, P.; Kadi, F. Adherence to DASH-Style Dietary Pattern Impacts on Adiponectin and Clustered Metabolic Risk in Older Women. Nutrients 2019, 11, 805. [Google Scholar] [CrossRef]
- Pickworth, C.K.; Deichert, D.A.; Corroon, J.; Bradley, R.D. Randomized controlled trials investigating the relationship between dietary pattern and high-sensitivity C-reactive protein: A systematic review. Nutr. Rev. 2019, 77, 363–375. [Google Scholar] [CrossRef] [PubMed]
- Samadi, M.; Moradi, S.; Moradinazar, M.; Mostafai, R.; Pasdar, Y. Dietary pattern in relation to the risk of Alzheimer’s disease: A systematic review. Neurol. Sci. Off. J. Ital. Neurol. Soc. Ital. Soc. Clin. Neurophysiol. 2019, 40, 2031–2043. [Google Scholar] [CrossRef] [PubMed]
- Asemi, Z.; Samimi, M.; Tabassi, Z.; Sabihi, S.S.; Esmaillzadeh, A. A randomized controlled clinical trial investigating the effect of DASH diet on insulin resistance, inflammation, and oxidative stress in gestational diabetes. Nutrition 2013, 29, 619–624. [Google Scholar] [CrossRef] [PubMed]
- Saneei, P.; Hashemipour, M.; Kelishadi, R.; Esmaillzadeh, A. The Dietary Approaches to Stop Hypertension (DASH) diet affects inflammation in childhood metabolic syndrome: A randomized cross-over clinical trial. Ann. Nutr. Metab. 2014, 64, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Martin, C.L.; Sotres-Alvarez, D.; Siega-Riz, A.M. Maternal Dietary Patterns during the Second Trimester Are Associated with Preterm Birth. J. Nutr. 2015, 145, 1857–1864. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Grewal, J.; Hinkle, S.N.; Yisahak, S.F.; Grobman, W.A.; Newman, R.B.; Skupski, D.W.; Chien, E.K.; Wing, D.A.; Grantz, K.L.; et al. Healthy dietary patterns and common pregnancy complications: A prospective and longitudinal study. Am. J. Clin. Nutr. 2021, 114, 1229–1237. [Google Scholar] [CrossRef] [PubMed]
- Brennan, L.; Hu, F.B. Metabolomics-Based Dietary Biomarkers in Nutritional Epidemiology-Current Status and Future Opportunities. Mol. Nutr. Food Res. 2019, 63, e1701064. [Google Scholar] [CrossRef]
- Guasch-Ferré, M.; Bhupathiraju, S.N.; Hu, F.B. Use of Metabolomics in Improving Assessment of Dietary Intake. Clin. Chem. 2018, 64, 82–98. [Google Scholar] [CrossRef] [PubMed]
- Jin, Q.; Black, A.; Kales, S.N.; Vattem, D.; Ruiz-Canela, M.; Sotos-Prieto, M. Metabolomics and Microbiomes as Potential Tools to Evaluate the Effects of the Mediterranean Diet. Nutrients 2019, 11, 207. [Google Scholar] [CrossRef] [PubMed]
- Rebholz, C.M.; Lichtenstein, A.H.; Zheng, Z.; Appel, L.J.; Coresh, J. Serum untargeted metabolomic profile of the Dietary Approaches to Stop Hypertension (DASH) dietary pattern. Am. J. Clin. Nutr. 2018, 108, 243–255. [Google Scholar] [CrossRef] [PubMed]
- Playdon, M.C.; Moore, S.C.; Derkach, A.; Reedy, J.; Subar, A.F.; Sampson, J.N.; Albanes, D.; Gu, F.; Kontto, J.; Lassale, C.; et al. Identifying biomarkers of dietary patterns by using metabolomics. Am. J. Clin. Nutr. 2017, 105, 450–465. [Google Scholar] [CrossRef] [PubMed]
- Grewal, J.; Grantz, K.L.; Zhang, C.; Sciscione, A.; Wing, D.A.; Grobman, W.A.; Newman, R.B.; Wapner, R.; D’Alton, M.E.; Skupski, D.; et al. Cohort Profile: NICHD Fetal Growth Studies-Singletons and Twins. Int. J. Epidemiol. 2018, 47, 25–25l. [Google Scholar] [CrossRef] [PubMed]
- Subar, A.F.; Thompson, F.E.; Kipnis, V.; Midthune, D.; Hurwitz, P.; McNutt, S.; McIntosh, A.; Rosenfeld, S. Comparative validation of the Block, Willett, and National Cancer Institute food frequency questionnaires: The Eating at America’s Table Study. Am. J. Epidemiol. 2001, 154, 1089–1099. [Google Scholar] [CrossRef]
- Subar, A.F.; Kipnis, V.; Troiano, R.P.; Midthune, D.; Schoeller, D.A.; Bingham, S.; Sharbaugh, C.O.; Trabulsi, J.; Runswick, S.; Ballard-Barbash, R.; et al. Using intake biomarkers to evaluate the extent of dietary misreporting in a large sample of adults: The OPEN study. Am. J. Epidemiol. 2003, 158, 1–13. [Google Scholar] [CrossRef]
- Moshfegh, A.J.; Rhodes, D.G.; Baer, D.J.; Murayi, T.; Clemens, J.C.; Rumpler, W.V.; Paul, D.R.; Sebastian, R.S.; Kuczynski, K.J.; Ingwersen, L.A.; et al. The US Department of Agriculture Automated Multiple-Pass Method reduces bias in the collection of energy intakes. Am. J. Clin. Nutr. 2008, 88, 324–332. [Google Scholar] [CrossRef]
- Fung, T.T.; Chiuve, S.E.; McCullough, M.L.; Rexrode, K.M.; Logroscino, G.; Hu, F.B. Adherence to a DASH-style diet and risk of coronary heart disease and stroke in women. Arch. Intern. Med. 2008, 168, 713–720. [Google Scholar] [CrossRef]
- Cajka, T.; Smilowitz, J.T.; Fiehn, O. Validating Quantitative Untargeted Lipidomics Across Nine Liquid Chromatography-High-Resolution Mass Spectrometry Platforms. Anal. Chem. 2017, 89, 12360–12368. [Google Scholar] [CrossRef]
- Fiehn, O. Metabolomics by Gas Chromatography-Mass Spectrometry: Combined Targeted and Untargeted Profiling. Curr. Protoc. Mol. Biol. 2016, 114, 30.34.31–30.34.32. [Google Scholar] [CrossRef]
- Wei, R.; Wang, J.; Su, M.; Jia, E.; Chen, S.; Chen, T.; Ni, Y. Missing Value Imputation Approach for Mass Spectrometry-based Metabolomics Data. Sci. Rep. 2018, 8, 663. [Google Scholar] [CrossRef]
- Chasan-Taber, L.; Schmidt, M.D.; Roberts, D.E.; Hosmer, D.; Markenson, G.; Freedson, P.S. Development and validation of a Pregnancy Physical Activity Questionnaire. Med. Sci. Sports Exerc. 2004, 36, 1750–1760. [Google Scholar] [CrossRef]
- van de Geer, S.; Bühlmann, P.; Ritov, Y.A.; Dezeure, R. On asymptotically optimal confidence regions and tests for high-dimensional models. Ann. Stat. 2014, 42, 1166–1202. [Google Scholar] [CrossRef]
- Fei, Z.; Li, Y. Estimation and Inference for High Dimensional Generalized Linear Models: A Splitting and Smoothing Approach. arXiv 2019, arXiv:1903.04408. [Google Scholar]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Pourafshar, S.; Nicchitta, M.; Tyson, C.C.; Svetkey, L.P.; Corcoran, D.L.; Bain, J.R.; Muehlbauer, M.J.; Ilkayeva, O.; O’Connell, T.M.; Lin, P.H.; et al. Urine and Plasma Metabolome of Healthy Adults Consuming the DASH (Dietary Approaches to Stop Hypertension) Diet: A Randomized Pilot Feeding Study. Nutrients 2021, 13, 1768. [Google Scholar] [CrossRef] [PubMed]
- Stevens, V.L.; Carter, B.D.; McCullough, M.L.; Campbell, P.T.; Wang, Y. Metabolomic Profiles Associated with BMI, Waist Circumference, and Diabetes and Inflammation Biomarkers in Women. Obesity 2020, 28, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Walker, M.E.; Song, R.J.; Xu, X.; Gerszten, R.E.; Ngo, D.; Clish, C.B.; Corlin, L.; Ma, J.; Xanthakis, V.; Jacques, P.F.; et al. Proteomic and Metabolomic Correlates of Healthy Dietary Patterns: The Framingham Heart Study. Nutrients 2020, 12, 1476. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Hu, E.A.; Wong, K.E.; Yu, B.; Steffen, L.M.; Seidelmann, S.B.; Boerwinkle, E.; Coresh, J.; Rebholz, C.M. Serum Metabolites Associated with Healthy Diets in African Americans and European Americans. J. Nutr. 2021, 151, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Anderson, C.A.; Hu, E.A.; Zheng, Z.; Appel, L.J.; He, J.; Feldman, H.I.; Anderson, A.H.; Ricardo, A.C.; Bhat, Z.; et al. Plasma Metabolomic Signatures of Healthy Dietary Patterns in the Chronic Renal Insufficiency Cohort (CRIC) Study. J. Nutr. 2021, 151, 2894–2907. [Google Scholar] [CrossRef] [PubMed]
- Yashpal, S.; Liese, A.D.; Boucher, B.A.; Wagenknecht, L.E.; Haffner, S.M.; Johnston, L.W.; Bazinet, R.P.; Rewers, M.; Rotter, J.I.; Watkins, S.M.; et al. Metabolomic profiling of the Dietary Approaches to Stop Hypertension diet provides novel insights for the nutritional epidemiology of type 2 diabetes mellitus. Br. J. Nutr. 2022, 128, 487–497. [Google Scholar] [CrossRef]
- McCullough, M.L.; Maliniak, M.L.; Stevens, V.L.; Carter, B.D.; Hodge, R.A.; Wang, Y. Metabolomic markers of healthy dietary patterns in US postmenopausal women. Am. J. Clin. Nutr. 2019, 109, 1439–1451. [Google Scholar] [CrossRef]
- Ford, E.S.; Schleicher, R.L.; Mokdad, A.H.; Ajani, U.A.; Liu, S. Distribution of serum concentrations of alpha-tocopherol and gamma-tocopherol in the US population. Am. J. Clin. Nutr. 2006, 84, 375–383. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q. Natural forms of vitamin E: Metabolism, antioxidant, and anti-inflammatory activities and their role in disease prevention and therapy. Free Radic. Biol. Med. 2014, 72, 76–90. [Google Scholar] [CrossRef] [PubMed]
- Cook-Mills, J.M.; McCary, C.A. Isoforms of vitamin E differentially regulate inflammation. Endocr. Metab. Immune Disord. Drug Targets 2010, 10, 348–366. [Google Scholar] [CrossRef] [PubMed]
- Berdnikovs, S.; Abdala-Valencia, H.; McCary, C.; Somand, M.; Cole, R.; Garcia, A.; Bryce, P.; Cook-Mills, J.M. Isoforms of vitamin E have opposing immunoregulatory functions during inflammation by regulating leukocyte recruitment. J. Immunol. 2009, 182, 4395–4405. [Google Scholar] [CrossRef] [PubMed]
- Marchese, M.E.; Kumar, R.; Colangelo, L.A.; Avila, P.C.; Jacobs, D.R., Jr.; Gross, M.; Sood, A.; Liu, K.; Cook-Mills, J.M. The vitamin E isoforms α-tocopherol and γ-tocopherol have opposite associations with spirometric parameters: The CARDIA study. Respir. Res. 2014, 15, 31. [Google Scholar] [CrossRef]
- Ford, E.S.; Mokdad, A.H.; Ajani, U.A.; Liu, S. Associations between concentrations of alpha- and gamma-tocopherol and concentrations of glucose, glycosylated haemoglobin, insulin and C-peptide among US adults. Br. J. Nutr. 2005, 93, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Nagao, M.; Moriyama, Y.; Yamagishi, K.; Iso, H.; Tamakoshi, A. Relation of serum α- and γ-tocopherol levels to cardiovascular disease-related mortality among Japanese men and women. J. Epidemiol. 2012, 22, 402–410. [Google Scholar] [CrossRef]
- Krall, A.S.; Xu, S.; Graeber, T.G.; Braas, D.; Christofk, H.R. Asparagine promotes cancer cell proliferation through use as an amino acid exchange factor. Nat. Commun. 2016, 7, 11457. [Google Scholar] [CrossRef]
- Huang, H.; Vandekeere, S.; Kalucka, J.; Bierhansl, L.; Zecchin, A.; Brüning, U.; Visnagri, A.; Yuldasheva, N.; Goveia, J.; Cruys, B.; et al. Role of glutamine and interlinked asparagine metabolism in vessel formation. EMBO J. 2017, 36, 2334–2352. [Google Scholar] [CrossRef]
- Ross, A.B.; Svelander, C.; Undeland, I.; Pinto, R.; Sandberg, A.S. Herring and Beef Meals Lead to Differences in Plasma 2-Aminoadipic Acid, β-Alanine, 4-Hydroxyproline, Cetoleic Acid, and Docosahexaenoic Acid Concentrations in Overweight Men. J. Nutr. 2015, 145, 2456–2463. [Google Scholar] [CrossRef]
- Hobson, R.M.; Saunders, B.; Ball, G.; Harris, R.C.; Sale, C. Effects of β-alanine supplementation on exercise performance: A meta-analysis. Amino Acids 2012, 43, 25–37. [Google Scholar] [CrossRef]
- Creighton, J.V.; de Souza Gonçalves, L.; Artioli, G.G.; Tan, D.; Elliott-Sale, K.J.; Turner, M.D.; Doig, C.L.; Sale, C. Physiological Roles of Carnosine in Myocardial Function and Health. Adv. Nutr. 2022, 13, 1914–1929. [Google Scholar] [CrossRef] [PubMed]
- Razak, M.A.; Begum, P.S.; Viswanath, B.; Rajagopal, S. Multifarious Beneficial Effect of Nonessential Amino Acid, Glycine: A Review. Oxidative Med. Cell. Longev. 2017, 2017, 1716701. [Google Scholar] [CrossRef] [PubMed]
- Adeva-Andany, M.; Souto-Adeva, G.; Ameneiros-Rodríguez, E.; Fernández-Fernández, C.; Donapetry-García, C.; Domínguez-Montero, A. Insulin resistance and glycine metabolism in humans. Amino Acids 2018, 50, 11–27. [Google Scholar] [CrossRef] [PubMed]
- Stapleton, P.P.; Charles, R.P.; Redmond, H.P.; Bouchier-Hayes, D.J. Taurine and human nutrition. Clin. Nutr. 1997, 16, 103–108. [Google Scholar] [CrossRef]
- Wójcik, O.P.; Koenig, K.L.; Zeleniuch-Jacquotte, A.; Costa, M.; Chen, Y. The potential protective effects of taurine on coronary heart disease. Atherosclerosis 2010, 208, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Barupal, D.K.; Fiehn, O. Generating the Blood Exposome Database Using a Comprehensive Text Mining and Database Fusion Approach. Environ. Health Perspect. 2019, 127, 97008. [Google Scholar] [CrossRef]
- Grembecka, M. Natural sweeteners in a human diet. Rocz. Panstw. Zakl. Hig. 2015, 66, 195–202. [Google Scholar]
- Rhee, E.P.; Cheng, S.; Larson, M.G.; Walford, G.A.; Lewis, G.D.; McCabe, E.; Yang, E.; Farrell, L.; Fox, C.S.; O’Donnell, C.J.; et al. Lipid profiling identifies a triacylglycerol signature of insulin resistance and improves diabetes prediction in humans. J. Clin. Investig. 2011, 121, 1402–1411. [Google Scholar] [CrossRef] [PubMed]
- Stegemann, C.; Pechlaner, R.; Willeit, P.; Langley, S.R.; Mangino, M.; Mayr, U.; Menni, C.; Moayyeri, A.; Santer, P.; Rungger, G.; et al. Lipidomics profiling and risk of cardiovascular disease in the prospective population-based Bruneck study. Circulation 2014, 129, 1821–1831. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Cheong, L.Z.; Zhang, X.; Ali, A.H.; Jin, Q.; Wei, W.; Wang, X. Dietary Sphingomyelin Metabolism and Roles in Gut Health and Cognitive Development. Adv. Nutr. 2022, 13, 474–491. [Google Scholar] [CrossRef] [PubMed]


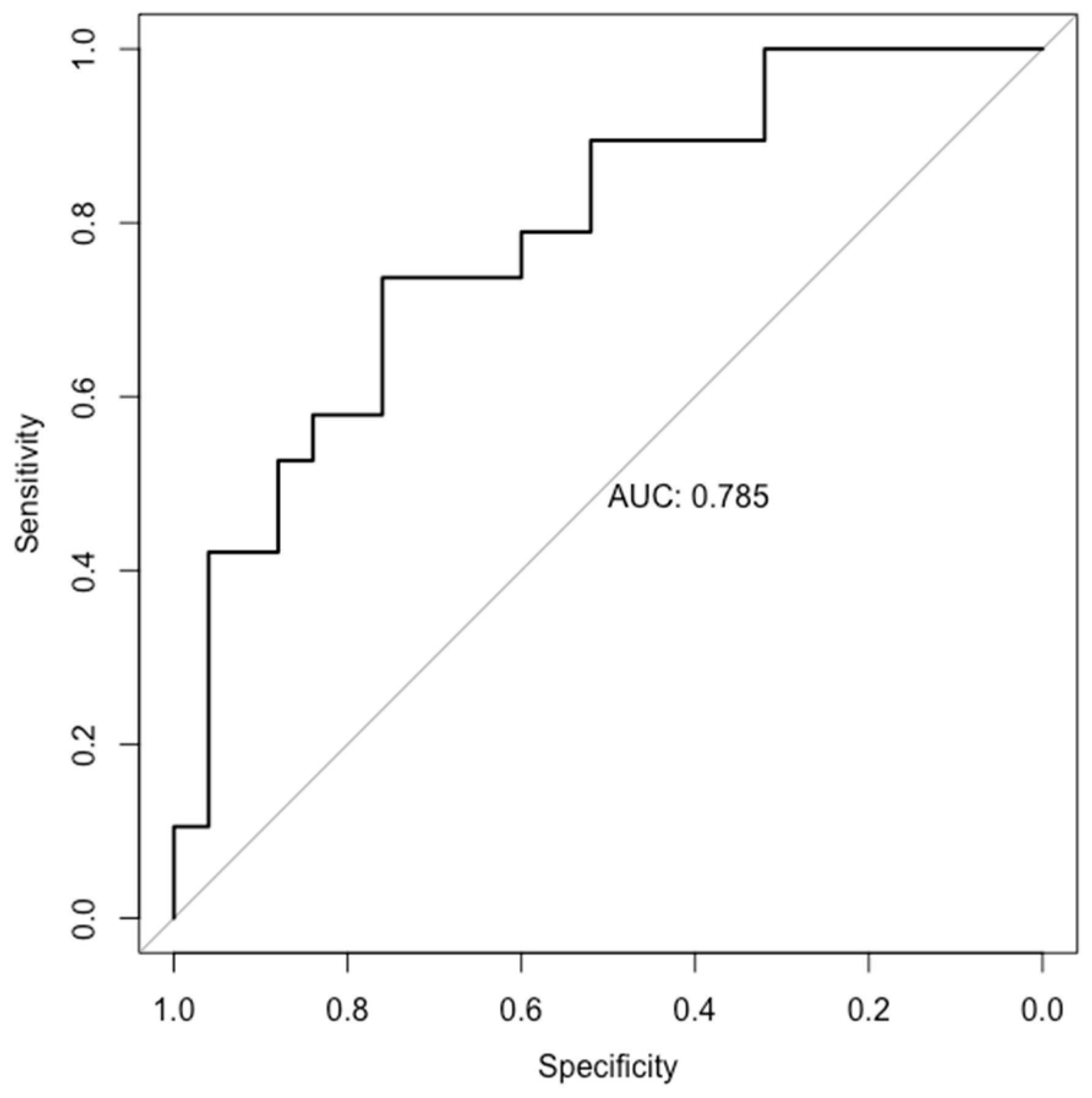{kind=link}
| Characteristics | All (N = 186) | DASH Score Tertiles | |||
|---|---|---|---|---|---|
| T1 (N = 70) | T2 (N = 54) | T3 (N = 62) | p | ||
| DASH score (median, IQR) | 25 (22–28) | 21 (19–22) | 25 (24–26) | 30 (28–31) | |
| Age, years | 28.00 (0.40) | 26.95 (0.63) | 28.72 (0.79) | 28.46 (0.68) | 0.15 |
| Race/ethnicity, % (N) | <0.001 | ||||
| Non-Hispanic Whites | 27.18 (37) | 20.18 (10) | 30.03 (12) | 31.75 (15) | |
| Non-Hispanic Blacks | 25.70 (23) | 31.60 (12) | 17.99 (5) | 25.85 (6) | |
| Hispanics | 24.39 (70) | 26.84 (28) | 27.28 (20) | 19.86 (22) | |
| Asian/Pacific Islanders | 22.73 (56) | 21.38 (20) | 24.70 (17) | 22.54 (19) | |
| Education, N (%) | 0.001 | ||||
| High-school or less | 47.15 (84) | 52.34 (38) | 40.74 (20) | 46.99 (26) | |
| Associates or higher | 52.85 (102) | 47.66 (32) | 59.26 (34) | 53.01 (36) | |
| Married/living with a partner, % (N) | 70.14 (147) | 76.41 (56) | 64.79 (40) | 68.17 (51) | <0.001 |
| Born in the United States, % (N) | 66.99 (99) | 73.72 (39) | 71.93 (29) | 56.83 (31) | <0.001 |
| Health insurance with private or managed care, % (N) | 63.71 (123) | 55.06 (41) | 70.70 (36) | 66.75 (46) | <0.001 |
| Nulliparous, % (N) | 50.54 (81) | 35.79 (23) | 59.37 (28) | 58.03 (30) | <0.001 |
| Consumed alcoholic beverage 3 months before pregnancy, % (N) | 61.52 (113) | 55.06 (42) | 75.68 (35) | 57.08 (36) | <0.001 |
| Pre-pregnancy BMI, kg/m2 | 25.51 (0.38) | 26.64 (0.68) | 26.54 (0.77) | 23.67 (0.43) | 0.001 |
| Pre-pregnancy BMI status, % (N) | <0.001 | ||||
| Normal (BMI < 25.0, kg/m2) | 50.38 (84) | 49.79 (29) | 43.72 (21) | 55.95 (34) | |
| Overweight (BMI: 25.0–29.9 kg/m2) | 34.80 (63) | 26.28 (23) | 38.86 (21) | 39.91 (19) | |
| Obese (BMI > 30, kg/m2) | 14.82 (39) | 23.93 (18) | 17.42 (12) | 4.14 (9) | |
| Total physical activity, metabolic equivalent hours/week | 307.61 (10.59) | 339.12 (20.03) | 303.48 (19.88) | 280.56 (14.18) | 0.06 |
| Dietary intakes §, | |||||
| Total energy, kcal/day | 2191.16 (71.19) | 1876.97 (81.76) | 2150.24 (135.83) | 2522.49 (137.75) | <0.001 |
| Total carbohydrate, g/day | 299.60 (10.82) | 244.91 (12.90) | 275.49 (17.25) | 367.32 (21.40) | <0.001 |
| Total protein, g/day | 86.15 (2.98) | 73.58 (3.29) | 82.96 (5.86) | 100.58 (5.73) | <0.001 |
| Total fatty acids, g/day | 77.71 (2.81) | 70.39 (2.94) | 84.56 (6.23) | 79.56 (5.36) | 0.12 |
| Saturated fatty acids (SAT), g/day | 25.51 (1.03) | 24.52 (1.16) | 26.51 (2.29) | 25.71 (1.98) | 0.75 |
| Monounsaturated fatty acids (MUFA), g/day | 29.65 (1.13) | 26.42 (1.13) | 32.77 (2.49) | 30.40 (2.17) | 0.07 |
| Polyunsaturated fatty acids (PUFAs), g/day | 16.40 (0.60) | 13.81 (0.57) | 18.97 (1.29) | 16.94 (0.97) | <0.001 |
| Total dietary fiber, g/day | 22.37 (0.84) | 15.88 (0.81) | 22.22 (1.46) | 28.70 (1.56) | <0.001 |
| Cholesterol, mg/day | 286.53 (11.88) | 312.13 (18.10) | 278.15 (22.24) | 268.33 (21.57) | 0.27 |
| Vegetables, serving/day | 3.66 (0.19) | 2.67 (0.23) | 3.48 (0.37) | 4.73 (0.36) | <0.001 |
| Whole fruit, serving/day | 6.39 (0.45) | 3.93 (0.44) | 4.63 (0.59) | 10.06 (0.94) | <0.001 |
| Whole grain, g/day | 27.65 (1.42) | 18.53 (1.80) | 31.09 (2.92) | 33.79 (2.35) | <0.001 |
| Nuts and legumes, serving/day | 0.67 (0.05) | 0.32 (0.04) | 0.78 (0.11) | 0.93 (0.08) | <0.001 |
| Red/processed meat, serving/day | 0.47 (0.03) | 0.55 (0.04) | 0.45 (0.05) | 0.42 (0.06) | 0.14 |
| Low-fat diary, serving/day | 1.52 (0.14) | 0.65 (0.16) | 0.99 (0.14) | 2.75 (0.30) | <0.001 |
| Sodium, mg/d ‡ | 3598.52 (60.64) | 3129.12 (122.93) | 3573.03 (271.13) | 3730.03 (229.77) | 0.09 |
| Sugar-sweetened beverages, serving/day | 0.69 (0.09) | 1.07 (0.16) | 0.73 (0.21) | 0.29 (0.08) | <0.001 |
| Superclass 1 | Class/ Subclass | Metabolites | Direction of the Association in LASSO 2 | Association in Linear Regression 3 | Associated Food Groups (Regression Coefficients) | |
|---|---|---|---|---|---|---|
| Coefficients | Benjamini–Hochberg-adjusted p-values 3 | |||||
| Organic acids and derivatives | Amino acids | Asparagine | Positive | 0.13 | 0.03 | Sodium (5 10−5) |
| Beta-alanine | Positive | 0.18 | 0.03 | Low-fat dairy (0.04) | ||
| Glycine | Negative | −0.18 | 0.003 | Low-fat dairy (−0.05) | ||
| Taurine | Negative | −0.49 | 0.003 | / 4 | ||
| Amino acid derivatives | Hydroxycarbamate | Positive | 0.32 | <0.001 | Low-fat dairy (0.04) | |
| Organoheterocyclic compounds | Pyrrolidines | Maleimide | Positive | 0.25 | 0.04 | / |
| Organic oxygen compounds | Carbohydrate/Monosaccharides | Xylitol | Positive | 0.16 | 0.01 | / |
| Lipids and lipids-like molecules | Fatty acyls | Dodecanoic acid | Positive | 0.22 | 0.02 | Fruit (0.02) and low-fat dairy (0.05) |
| Prenol lipids/vitamin E | Gamma-tocopherol | Negative | −0.19 | 0.03 | / | |
| Glycerolipids | TG (49:1) | Positive | 0.38 | <0.001 | Low-fat dairy (0.07) | |
| TG (52:2) | Negative | −0.08 | 0.01 | / | ||
| Glycerophospholipids | PC (31:0) | Positive | 0.25 | 0.002 | Whole grains (0.01), low-fat dairy (0.07), and sodium (6 10−5) | |
| PC (35:3) | Positive | 0.23 | 0.01 | Low-fat dairy (0.07) and sodium ( 10−5) | ||
| PC (36:4) C | Positive | 0.07 | 0.001 | Fruits (0.01) | ||
| PC (36:5) B | Positive | 0.43 | 0.002 | / | ||
| PC (38:4) B | Positive | 0.16 | <0.001 | Low-fat dairy (0.04) and sodium (3 10−5) | ||
| PC (42:6) | Positive | 0.60 | <0.001 | Low-fat dairy (0.09) | ||
| Sphingolipids | SM (d32:0) | Positive | 0.29 | 0.003 | Low-fat dairy (0.10) and sodium (8 10−5) | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, L.; Dai, J.; Yu, G.; Pang, W.W.; Rahman, M.L.; Liu, X.; Fiehn, O.; Guivarch, C.; Chen, Z.; Zhang, C. Metabolomic Biomarkers of Dietary Approaches to Stop Hypertension (DASH) Dietary Patterns in Pregnant Women. Nutrients 2024, 16, 492. https://doi.org/10.3390/nu16040492
Chen L, Dai J, Yu G, Pang WW, Rahman ML, Liu X, Fiehn O, Guivarch C, Chen Z, Zhang C. Metabolomic Biomarkers of Dietary Approaches to Stop Hypertension (DASH) Dietary Patterns in Pregnant Women. Nutrients. 2024; 16(4):492. https://doi.org/10.3390/nu16040492
Chicago/Turabian StyleChen, Liwei, Jin Dai, Guoqi Yu, Wei Wei Pang, Mohammad L. Rahman, Xinyue Liu, Oliver Fiehn, Claire Guivarch, Zhen Chen, and Cuilin Zhang. 2024. "Metabolomic Biomarkers of Dietary Approaches to Stop Hypertension (DASH) Dietary Patterns in Pregnant Women" Nutrients 16, no. 4: 492. https://doi.org/10.3390/nu16040492
APA StyleChen, L., Dai, J., Yu, G., Pang, W. W., Rahman, M. L., Liu, X., Fiehn, O., Guivarch, C., Chen, Z., & Zhang, C. (2024). Metabolomic Biomarkers of Dietary Approaches to Stop Hypertension (DASH) Dietary Patterns in Pregnant Women. Nutrients, 16(4), 492. https://doi.org/10.3390/nu16040492