
The Potential of Bioactive Fish Collagen Oligopeptides against Hydrogen Peroxide-Induced NIH/3T3 and HUVEC Damage: The Involvement of the Mitochondria



Abstract
:1. Introduction
2. Materials and Methods
2.1. FCOPs
2.2. Study Design
2.3. Cell Viability
2.4. Flow Cytometry
2.5. Biochemical Analysis
2.6. Western Blot Analysis
2.7. Statistical Analysis
3. Results
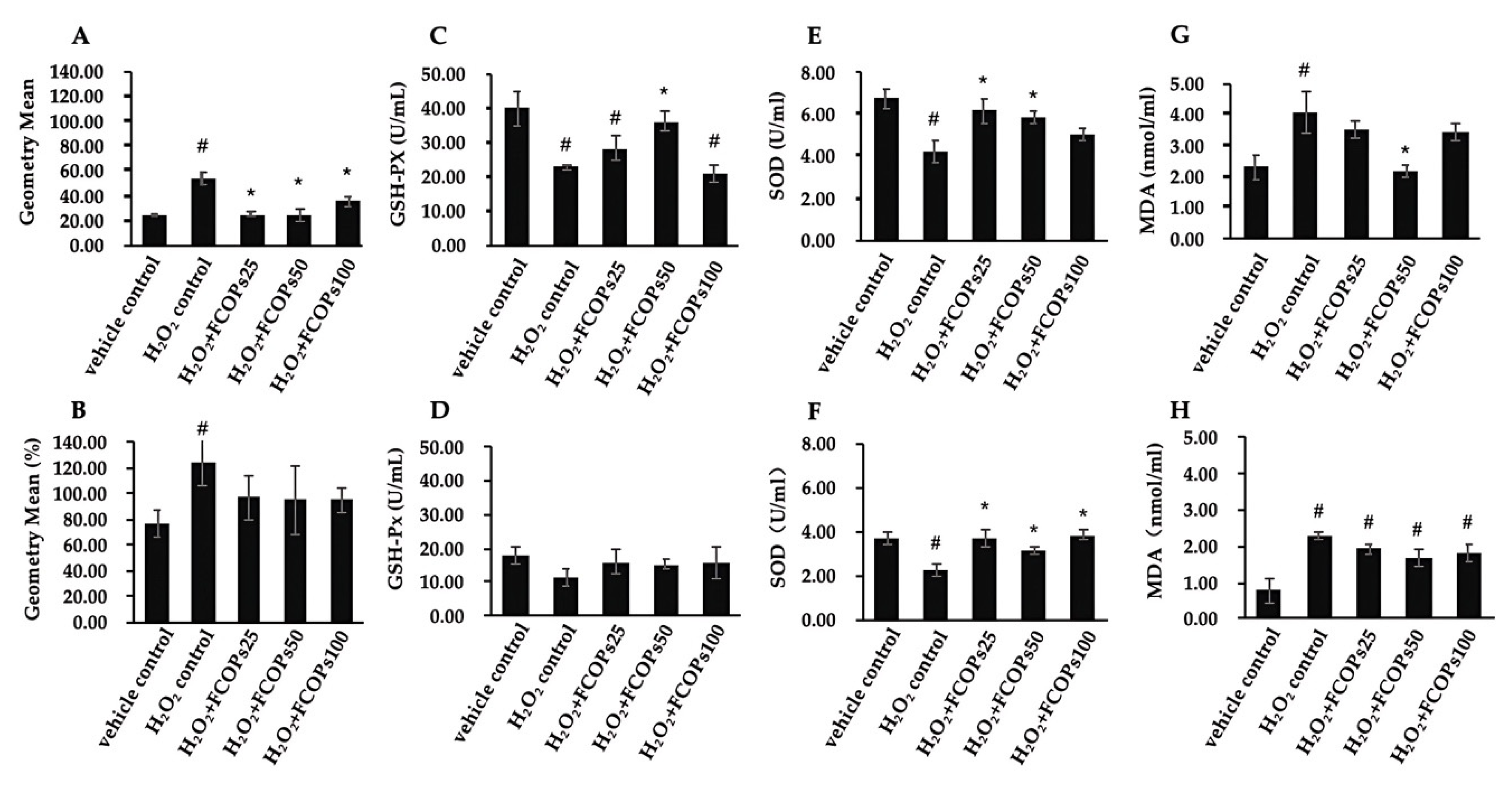
3.1. FCOP Enhanced Antioxidant Activities in H2O2-Treated NIH/3T3 and HUVECs
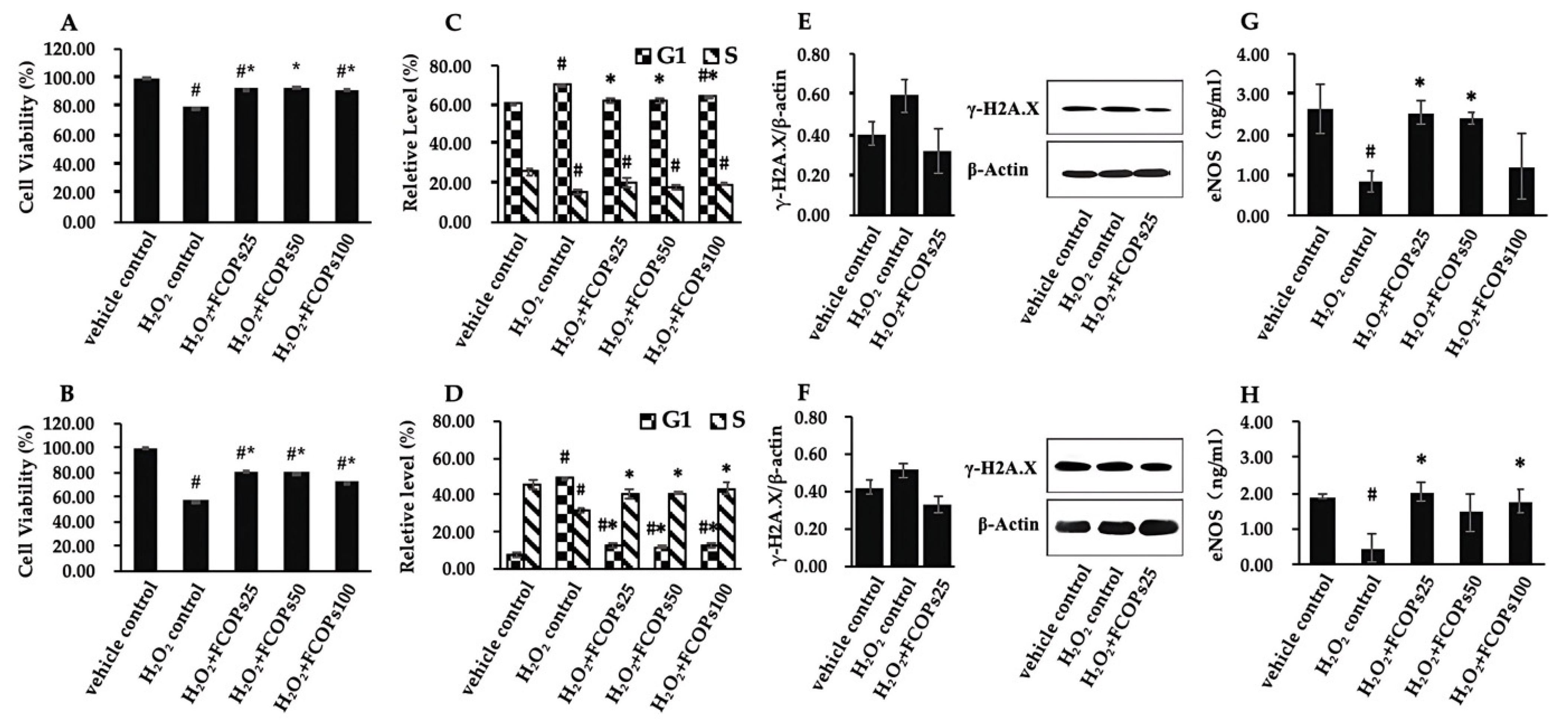
3.2. FCOPs Promote Homeostasis in H2O2-Treated NIH/3T3 and HUVECs
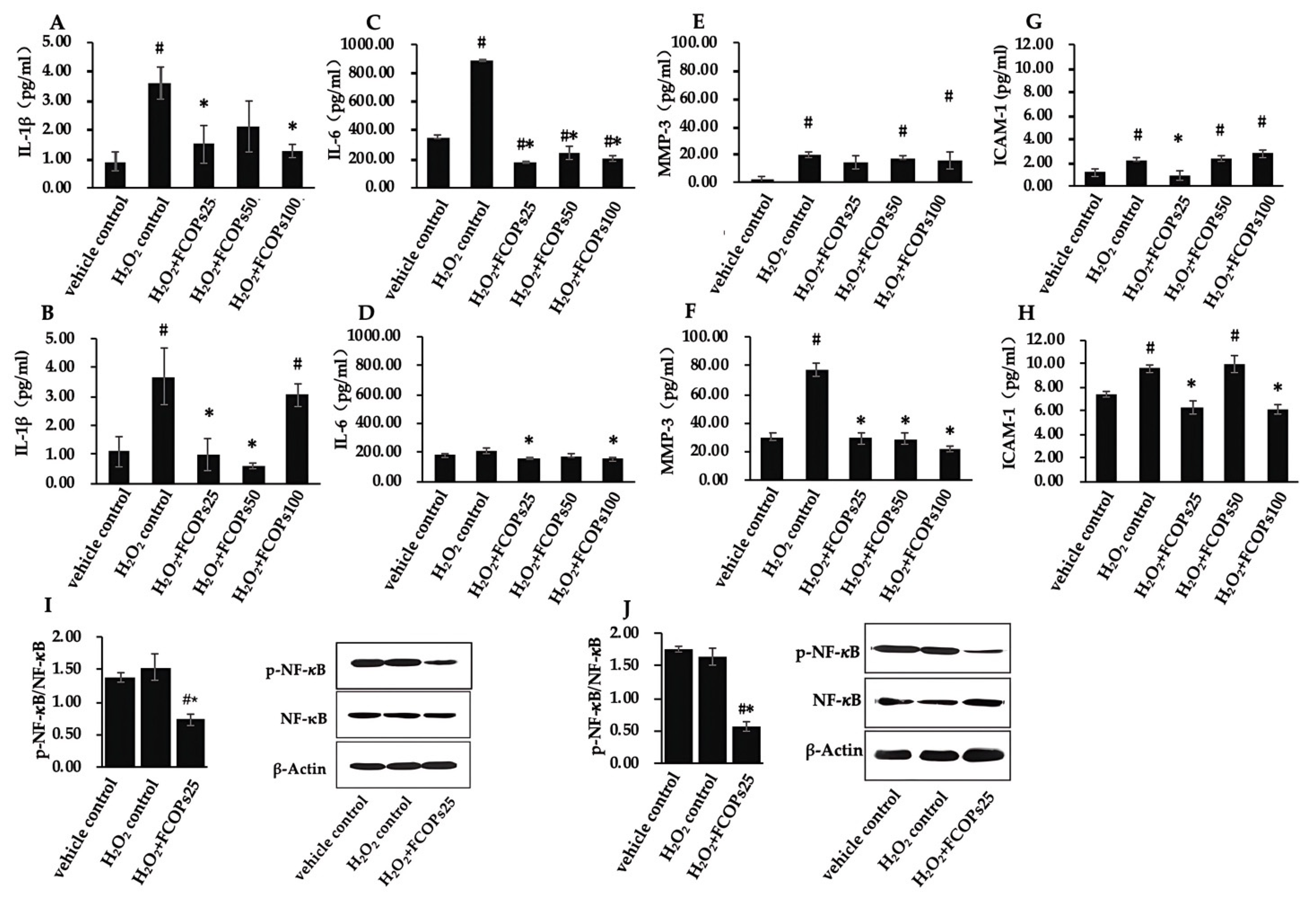
3.3. FCOPs Suppressed Inflammation in H2O2-Treated NIH/3T3 and HUVECs
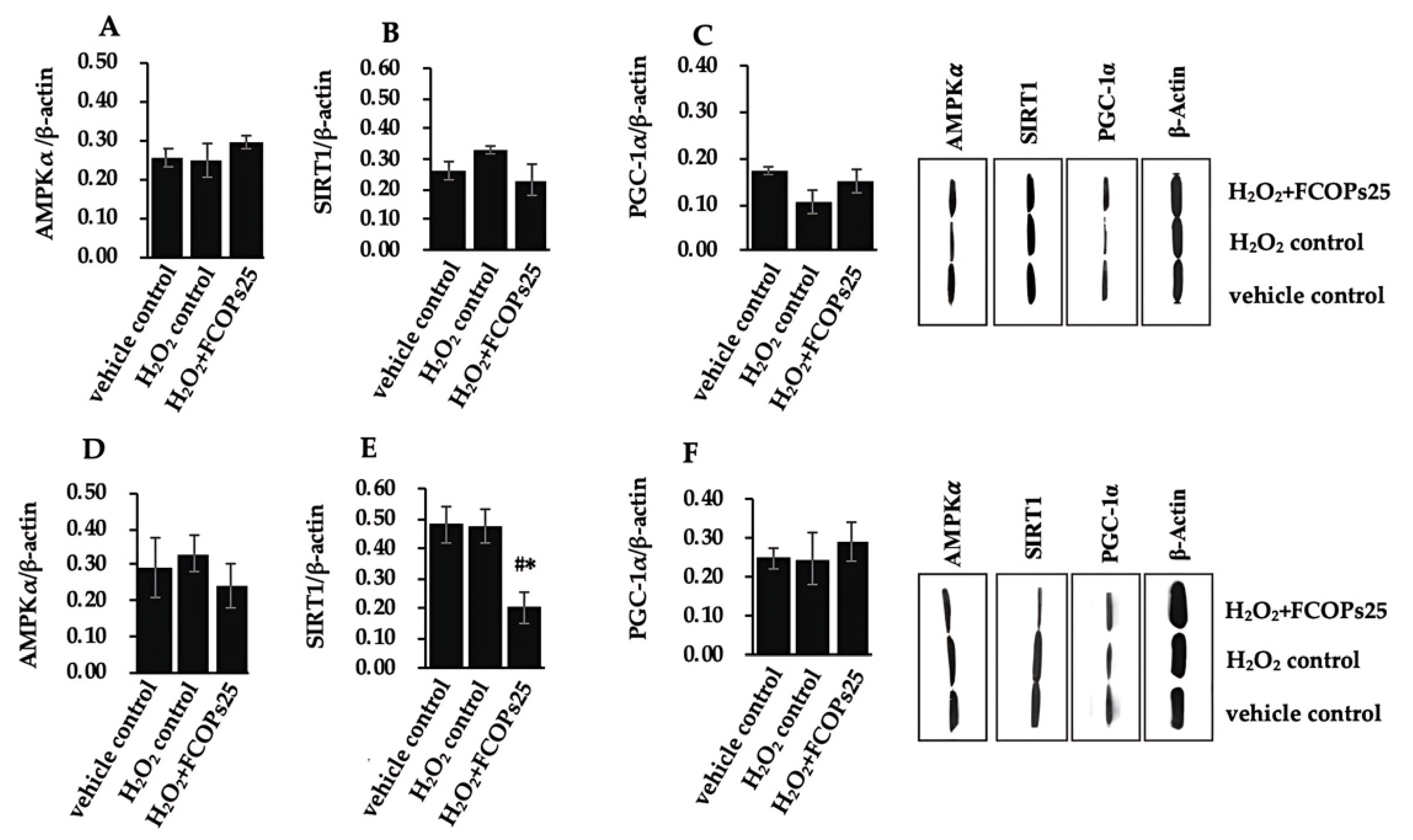
3.4. FCOPs Influenced Mitochondrial Function and Biogenesis in H2O2-Treated NIH/3T3 and HUVECs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sies, H.; Berndt, C.; Jones, D.P. Oxidative Stress. Annu. Rev. Biochem. 2017, 86, 715–748. [Google Scholar] [CrossRef] [PubMed]
- Sies, H. Oxidative stress: A concept in redox biology and medicine. Redox Biol. 2015, 4, 180–183. [Google Scholar] [CrossRef] [PubMed]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative Stress: Harms and Benefits for Human Health. Oxidative Med. Cell. Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef]
- Freudenthal, B.D.; Beard, W.A.; Perera, L.; Shock, D.D.; Kim, T.; Schlick, T.; Wilson, S.H. Uncovering the polymerase-induced cytotoxicity of an oxidized nucleotide. Nature 2015, 517, 635–639. [Google Scholar] [CrossRef] [PubMed]
- Crisóstomo, L.; Oliveira, P.F.; Alves, M.G. Antioxidants, Oxidative Stress, and Non-Communicable Diseases. Antioxidants 2022, 11, 1080. [Google Scholar] [CrossRef]
- Teleanu, D.M.; Niculescu, A.G.; Lungu, I.I.; Radu, C.I.; Vladâcenco, O.; Roza, E.; Costăchescu, B.; Grumezescu, A.M.; Teleanu, R.I. An Overview of Oxidative Stress, Neuroinflammation, and Neurodegenerative Diseases. Int. J. Mol. Sci. 2022, 23, 5938. [Google Scholar] [CrossRef] [PubMed]
- Sharifi-Rad, M.; Anil Kumar, N.V.; Zucca, P.; Varoni, E.M.; Dini, L.; Panzarini, E.; Rajkovic, J.; Tsouh Fokou, P.V.; Azzini, E.; Peluso, I.; et al. Lifestyle, Oxidative Stress, and Antioxidants: Back and Forth in the Pathophysiology of Chronic Diseases. Front. Physiol. 2020, 11, 694. [Google Scholar] [CrossRef] [PubMed]
- Senoner, T.; Dichtl, W. Oxidative Stress in Cardiovascular Diseases: Still a Therapeutic Target? Nutrients 2019, 11, 2090. [Google Scholar] [CrossRef] [PubMed]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. Hallmarks of aging: An expanding universe. Cell 2023, 186, 243–278. [Google Scholar] [CrossRef] [PubMed]
- Liguori, I.; Russo, G.; Curcio, F.; Bulli, G.; Aran, L.; Della-Morte, D.; Gargiulo, G.; Testa, G.; Cacciatore, F.; Bonaduce, D.; et al. Oxidative stress, aging, and diseases. Clin. Interv. Aging 2018, 13, 757–772. [Google Scholar] [CrossRef] [PubMed]
- Ursini, F.; Maiorino, M.; Forman, H.J. Redox homeostasis: The Golden Mean of healthy living. Redox Biol. 2016, 8, 205–215. [Google Scholar] [CrossRef]
- Griendling, K.K.; Camargo, L.L.; Rios, F.J.; Alves-Lopes, R.; Montezano, A.C.; Touyz, R.M. Oxidative Stress and Hypertension. Circ. Res. 2021, 128, 993–1020. [Google Scholar] [CrossRef] [PubMed]
- Harrington, J.S.; Ryter, S.W.; Plataki, M.; Price, D.R.; Choi, A.M.K. Mitochondria in health, disease, and aging. Physiol. Rev. 2023, 103, 2349–2422. [Google Scholar] [CrossRef] [PubMed]
- Downer, S.; Berkowitz, S.A.; Harlan, T.S.; Olstad, D.L.; Mozaffarian, D. Food is medicine: Actions to integrate food and nutrition into healthcare. BMJ 2020, 369, m2482. [Google Scholar] [CrossRef] [PubMed]
- Bhat, S.; Coyle, D.H.; Trieu, K.; Neal, B.; Mozaffarian, D.; Marklund, M.; Wu, J.H.Y. Healthy Food Prescription Programs and their Impact on Dietary Behavior and Cardiometabolic Risk Factors: A Systematic Review and Meta-Analysis. Adv. Nutr. 2021, 12, 1944–1956. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Dai, Z. Physicochemical, Structural and Antioxidant Properties of Collagens from the Swim Bladder of Four Fish Species. Mar. Drugs 2022, 20, 550. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Wang, J.; Liu, Z.; Pei, X.; Han, X.; Li, Y. Antioxidant effect of a marine oligopeptide preparation from chum salmon (Oncorhynchus keta) by enzymatic hydrolysis in radiation injured mice. Mar. Drugs 2011, 9, 2304–2315. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Li, Q.; Lin, B.; Yu, Y.; Ding, Y.; Dai, X.; Li, Y. Comparative studies of oral administration of marine collagen peptides from Chum Salmon (Oncorhynchus keta) pre- and post-acute ethanol intoxication in female Sprague-Dawley rats. Food Funct. 2014, 5, 2078–2085. [Google Scholar] [CrossRef]
- Yang, R.; Pei, X.; Wang, J.; Zhang, Z.; Zhao, H.; Li, Q.; Zhao, M.; Li, Y. Protective effect of a marine oligopeptide preparation from chum salmon (Oncorhynchus keta) on radiation-induced immune suppression in mice. J. Sci. Food Agric. 2010, 90, 2241–2248. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.F.; Li, G.Z.; Peng, H.B.; Zhang, F.; Chen, Y.; Li, Y. Treatment with marine collagen peptides modulates glucose and lipid metabolism in Chinese patients with type 2 diabetes mellitus. Appl. Physiol. Nutr. Metab. Physiol. Appl. Nutr. Metab. 2010, 35, 797–804. [Google Scholar] [CrossRef] [PubMed]
- Lin, B.; Zhang, F.; Yu, Y.; Jiang, Q.; Zhang, Z.; Wang, J.; Li, Y. Marine collagen peptides protect against early alcoholic liver injury in rats. Br. J. Nutr. 2012, 107, 1160–1166. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Pei, X.R.; Wang, N.; Zhang, Z.F.; Wang, J.B.; Li, Y. Marine collagen peptides prepared from chum salmon (Oncorhynchus keta) skin extend the life span and inhibit spontaneous tumor incidence in Sprague-Dawley Rats. J. Med. Food 2010, 13, 757–770. [Google Scholar] [CrossRef] [PubMed]
- Zhai, L.; Xu, X.; Liu, J.; Jing, C.; Yang, X.; Zhao, D.; Jiang, R.; Sun, L.W. A Novel Biochemical Study of Anti-Dermal Fibroblast Replicative Senescence Potential of Panax Notoginseng Oligosaccharides. Front. Pharmacol. 2021, 12, 690538. [Google Scholar] [CrossRef] [PubMed]
- Schneider, V.S.; Bark, J.M.; Winnischofer, S.M.B.; Dos Santos, E.F.; Iacomini, M.; Cordeiro, L.M.C. Dietary fibres from guavira pomace, a co-product from fruit pulp industry: Characterization and cellular antioxidant activity. Food Res. Int. 2020, 132, 109065. [Google Scholar] [CrossRef] [PubMed]
- Liang, B.; Xiang, Y.; Zhang, X.; Wang, C.; Jin, B.; Zhao, Y.; Zheng, F. Systematic Pharmacology and GEO Database Mining Revealed the Therapeutic Mechanism of Xuefu Zhuyu Decoration for Atherosclerosis Cardiovascular Disease. Front. Cardiovasc. Med. 2020, 7, 592201. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Bi, R.; Hao, J.; Wang, S.; Huo, Y.; Demoz, R.M.; Banda, R.; Tian, S.; Xin, C.; Fu, M.; et al. A study on the protective effects of taxifolin on human umbilical vein endothelial cells and THP-1 cells damaged by hexavalent chromium: A probable mechanism for preventing cardiovascular disease induced by heavy metals. Food Funct. 2020, 11, 3851–3859. [Google Scholar] [CrossRef] [PubMed]
- Singh, A. Quantifying radiation damage. Nat. Methods 2022, 19, 520. [Google Scholar] [CrossRef] [PubMed]
- Sheng, Y.; Qiu, Y.T.; Wang, Y.M.; Chi, C.F.; Wang, B. Novel Antioxidant Collagen Peptides of Siberian Sturgeon (Acipenserbaerii) Cartilages: The Preparation, Characterization, and Cytoprotection of H2O2-Damaged Human Umbilical Vein Endothelial Cells (HUVECs). Mar. Drugs 2022, 20, 325. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Wu, H.; Chi, Y.; Deng, R.; He, Q. Structural characterization, erythrocyte protection, and antifatigue effect of antioxidant collagen peptides from tilapia (Oreochromis nilotica L.) skin. Food Funct. 2020, 11, 10149–10160. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.P.; Liang, C.H.; Wu, H.T.; Pang, H.Y.; Chen, C.; Wang, G.H.; Chan, L.P. Antioxidant and anti-inflammatory capacities of collagen peptides from milkfish (Chanos chanos) scales. J. Food Sci. Technol. 2018, 55, 2310–2317. [Google Scholar] [CrossRef] [PubMed]
- Xia, E.; Zhu, X.; Gao, X.; Ni, J.; Guo, H. Antiaging Potential of Peptides from Underused Marine Bioresources. Mar. Drugs 2021, 19, 513. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Yang, Q.; Chen, X.; Tian, Y.; Liu, Z.; Wang, S. Bioactive peptides derived from crimson snapper and in vivo anti-aging effects on fat diet-induced high fat Drosophila melanogaster. Food Funct. 2020, 11, 524–533. [Google Scholar] [CrossRef] [PubMed]
- Sies, H.; Belousov, V.V.; Chandel, N.S.; Davies, M.J.; Jones, D.P.; Mann, G.E.; Murphy, M.P.; Yamamoto, M.; Winterbourn, C. Defining roles of specific reactive oxygen species (ROS) in cell biology and physiology. Nat. Rev. Mol. Cell Biol. 2022, 23, 499–515. [Google Scholar] [CrossRef] [PubMed]
- Heo, A.J.; Kim, S.B.; Ji, C.H.; Han, D.; Lee, S.J.; Lee, S.H.; Lee, M.J.; Lee, J.S.; Ciechanover, A.; Kim, B.Y.; et al. The N-terminal cysteine is a dual sensor of oxygen and oxidative stress. Proc. Natl. Acad. Sci. USA 2021, 118, e2107993118. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.H.; Gu, Z.W.; Li, J.; Yang, W.Q.; Li, Y.L.; Qi, D.M.; Wang, D.Y.; Jiang, H.Q. Isorhynchophylline inhibits inflammatory responses in endothelial cells and macrophages through the NF-κB/NLRP3 signaling pathway. BMC Complement. Med. Ther. 2023, 23, 80. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Im, E. Regulation of miRNAs by Natural Antioxidants in Cardiovascular Diseases: Focus on SIRT1 and eNOS. Antioxidants 2021, 10, 377. [Google Scholar] [CrossRef]
- Inbaraj, B.S.; Lai, Y.W.; Chen, B.H. A comparative study on inhibition of lung cancer cells by nanoemulsion, nanoliposome, nanogold and their folic acid conjugates prepared with collagen peptides from Taiwan tilapia skin. Int. J. Biol. Macromol. 2024, 261, 129722. [Google Scholar] [CrossRef]
- Gaspardi, A.L.A.; da Silva, D.C.; Ponte, L.G.S.; Galland, F.; da Silva, V.S.N.; Simabuco, F.M.; Bezerra, R.M.N.; Pacheco, M.T.B. In vitro inhibition of glucose gastro-intestinal enzymes and antioxidant activity of hydrolyzed collagen peptides from different species. J. Food Biochem. 2022, 46, e14383. [Google Scholar] [CrossRef] [PubMed]
- Haghi Aminjan, H.; Abtahi, S.R.; Hazrati, E.; Chamanara, M.; Jalili, M.; Paknejad, B. Targeting of oxidative stress and inflammation through ROS/NF-kappaB pathway in phosphine-induced hepatotoxicity mitigation. Life Sci. 2019, 232, 116607. [Google Scholar] [CrossRef] [PubMed]
- Rahabi, M.; Salon, M.; Bruno-Bonnet, C.; Prat, M.; Jacquemin, G.; Benmoussa, K.; Alaeddine, M.; Parny, M.; Bernad, J.; Bertrand, B.; et al. Bioactive fish collagen peptides weaken intestinal inflammation by orienting colonic macrophages phenotype through mannose receptor activation. Eur. J. Nutr. 2022, 61, 2051–2066. [Google Scholar] [CrossRef] [PubMed]
- Woo, M.; Seol, B.G.; Kang, K.H.; Choi, Y.H.; Cho, E.J.; Noh, J.S. Effects of collagen peptides from skate (Raja kenojei) skin on improvements of the insulin signaling pathway via attenuation of oxidative stress and inflammation. Food Funct. 2020, 11, 2017–2025. [Google Scholar] [CrossRef] [PubMed]
- Subhan, F.; Kang, H.Y.; Lim, Y.; Ikram, M.; Baek, S.Y.; Jin, S.; Jeong, Y.H.; Kwak, J.Y.; Yoon, S. Fish Scale Collagen Peptides Protect against CoCl2/TNF-α-Induced Cytotoxicity and Inflammation via Inhibition of ROS, MAPK, and NF-κB Pathways in HaCaT Cells. Oxidative Med. Cell. Longev. 2017, 2017, 9703609. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Li, Y.; Wang, W.; Bai, Y.; Jia, H.; Yuan, Z.; Yang, Z. Role and mechanisms of the NF-κB signaling pathway in various developmental processes. Biomed. Pharmacother. 2022, 153, 113513. [Google Scholar] [CrossRef] [PubMed]
- Chook, C.Y.B.; Cheung, Y.M.; Ma, K.Y.; Leung, F.P.; Zhu, H.; Niu, Q.J.; Wong, W.T.; Chen, Z.Y. Physiological concentration of protocatechuic acid directly protects vascular endothelial function against inflammation in diabetes through Akt/eNOS pathway. Front. Nutr. 2023, 10, 1060226. [Google Scholar] [CrossRef] [PubMed]
- Ku, C.W.; Ho, T.J.; Huang, C.Y.; Chu, P.M.; Ou, H.C.; Hsieh, P.L. Cordycepin Attenuates Palmitic Acid-Induced Inflammation and Apoptosis of Vascular Endothelial Cells through Mediating PI3K/Akt/eNOS Signaling Pathway. Am. J. Chin. Med. 2021, 49, 1703–1722. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Kim, H. Inhibitory Effect of Astaxanthin on Oxidative Stress-Induced Mitochondrial Dysfunction-A Mini-Review. Nutrients 2018, 10, 1137. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhou, Z.; Min, W. Mitochondria, Oxidative Stress and Innate Immunity. Front. Physiol. 2018, 9, 1487. [Google Scholar] [CrossRef] [PubMed]
- Cantó, C.; Menzies, K.J.; Auwerx, J. NAD+ Metabolism and the Control of Energy Homeostasis: A Balancing Act between Mitochondria and the Nucleus. Cell Metab. 2015, 22, 31–53. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Li, R.; Ye, M.; Zhang, L.; Zheng, J.; Yang, Y.; Wei, X.; Zhao, Q. The circadian clock has roles in mesenchymal stem cell fate decision. Stem Cell Res. Ther. 2022, 13, 200. [Google Scholar] [CrossRef] [PubMed]
- Abu Shelbayeh, O.; Arroum, T.; Morris, S.; Busch, K.B. PGC-1α Is a Master Regulator of Mitochondrial Lifecycle and ROS Stress Response. Antioxidants 2023, 12, 1075. [Google Scholar] [CrossRef] [PubMed]
- Jannig, P.R.; Dumesic, P.A.; Spiegelman, B.M.; Ruas, J.L. SnapShot: Regulation and biology of PGC-1α. Cell 2022, 185, 1444–1444.e1. [Google Scholar] [CrossRef] [PubMed]
- Rius-Pérez, S.; Torres-Cuevas, I.; Millán, I.; Ortega, Á.L.; Pérez, S. PGC-1α, Inflammation, and Oxidative Stress: An Integrative View in Metabolism. Oxidative Med. Cell. Longev. 2020, 2020, 1452696. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino Acid | Ratio (g/100 g) | Amino Acid | Ratio (g/100 g) |
|---|---|---|---|
| Glycine | 23.67 | Threonine | 2.77 |
| Glutamic Acid | 11.22 | Leucine | 2.62 |
| Proline | 10.42 | Valine | 2.34 |
| Alanine | 10.24 | Phenylalanine | 1.90 |
| Hydroxyproline | 8.72 | Isoleucine | 1.30 |
| Arginine | 8.34 | Methionine | 1.06 |
| Aspartic Acid | 5.44 | Histidine | 0.75 |
| Lysine | 3.40 | Tyrosine | 0.32 |
| Serine | 2.83 | Cystine | 0.04 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, N.; Liu, R.; Xu, M.; Li, Y. The Potential of Bioactive Fish Collagen Oligopeptides against Hydrogen Peroxide-Induced NIH/3T3 and HUVEC Damage: The Involvement of the Mitochondria. Nutrients 2024, 16, 1004. https://doi.org/10.3390/nu16071004
Zhu N, Liu R, Xu M, Li Y. The Potential of Bioactive Fish Collagen Oligopeptides against Hydrogen Peroxide-Induced NIH/3T3 and HUVEC Damage: The Involvement of the Mitochondria. Nutrients. 2024; 16(7):1004. https://doi.org/10.3390/nu16071004
Chicago/Turabian StyleZhu, Na, Rui Liu, Meihong Xu, and Yong Li. 2024. "The Potential of Bioactive Fish Collagen Oligopeptides against Hydrogen Peroxide-Induced NIH/3T3 and HUVEC Damage: The Involvement of the Mitochondria" Nutrients 16, no. 7: 1004. https://doi.org/10.3390/nu16071004
APA StyleZhu, N., Liu, R., Xu, M., & Li, Y. (2024). The Potential of Bioactive Fish Collagen Oligopeptides against Hydrogen Peroxide-Induced NIH/3T3 and HUVEC Damage: The Involvement of the Mitochondria. Nutrients, 16(7), 1004. https://doi.org/10.3390/nu16071004