
Blood Phytosterol Concentration and Genetic Variant Associations in a Sample Population
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. The Study Population
2.2. Data Collection
2.3. Analytical Determinations
- -
- Phytosterol analysis
- -
- α-tocopherol analysis
- -
- DNA isolation and genotyping
2.4. Statistical Analysis
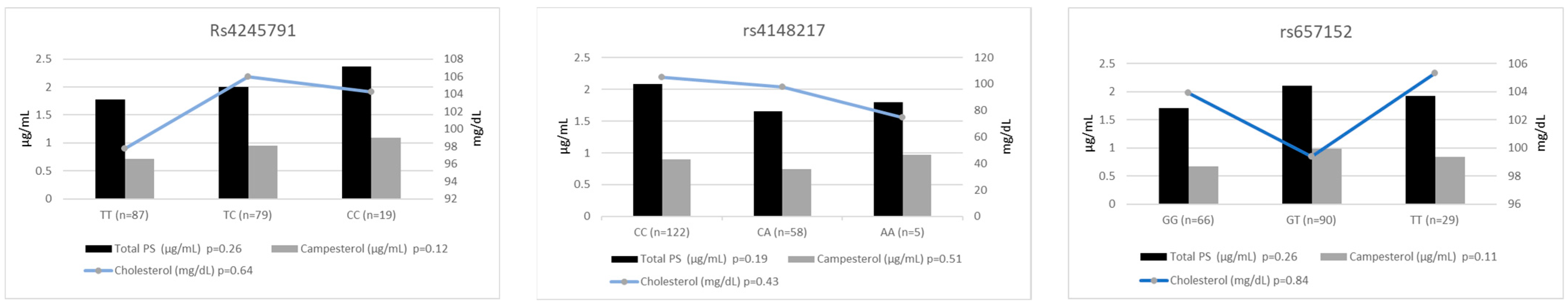
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
| 1. Are you vegetarian/vegan? | □ Yes □ No |
| 2. What types of oil do you usually consume? | □ None □ Sesame □ Olive □ Rice □ Sunflower □ Peanut □ Others. Which? |
| 3. How many days a week do you eat nuts? | □ None □ 1–3 □ More than 3 |
| 4. How many days a week do you consume butter or margarine? | □ None □ 1–3 □ More than 3 |
| 5. How many days a week do you consume pastries? | □ None □ 1–3 □ More than 3 |
| 6. How many days a week do you consume sausages? | □ None □ 1–3 □ More than 3 |
| 7. Do you take supplements rich in vitamin E, α-tocopherol (Auxina®), or a multivitamin complex (Supradyn®, Pharmaton®, Multicentrum®…)? | □ Yes □ No |
| 8. Do you take Danacol® or some product to lower cholesterol? | □ Yes □ No |
| 9. Do you take a cholesterol-lowering drug? | □ Yes □ No |
References
- Bhattacharyya, A.K.; Connor, W. Beta-Sitosterolemia and xanthomatosis. J. Clin. Investig. 1974, 53, 1033–1043. [Google Scholar] [CrossRef] [PubMed]
- Lu, K.; Lee, M.-H.; Hazard, S.; Brooks-Wilson, A.; Hidaka, H.; Kojima, H.; Ose, L.; Stalenhoef, A.F.; Mietinnen, T.; Bjorkhem, I.; et al. Two Genes That Map to the STSL Locus Cause Sitosterolemia: Genomic Structure and Spectrum of Mutations Involving Sterolin-1 and Sterolin-2, Encoded by ABCG5 and ABCG8, Respectively. Am. J. Hum. Genet. 2001, 69, 278–290. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.-H.; Lu, K.; Hazard, S.; Yu, H.; Shulenin, S.; Hidaka, H.; Kojima, H.; Allikmets, R.; Sakuma, N.; Pegoraro, R.; et al. Identification of a gene, ABCG5, important in the regulation of dietary cholesterol absorption. Nat. Genet. 2001, 27, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.B.; Salen, G.; Hidaka, H.; Kwiterovich, P.O.; Stalenhoef, A.F.; Miettinen, T.A.; Grundy, S.M.; Lee, M.H.; Rubenstein, J.S.; Polymeropoulos, M.H.; et al. Mapping a gene involved in regulating dietary cholesterol absorption. The sitosterolemia locus is found at chromosome 2p21. J. Clin. Investig. 1998, 102, 1041–1044. [Google Scholar] [CrossRef] [PubMed]
- Berge, K.E.; Tian, H.; Graf, G.A.; Yu, L.; Grishin, N.V.; Schultz, J.; Kwiterovich, P.; Shan, B.; Barnes, R.; Hobbs, H.H. Accumulation of dietary cholesterol in sitosterolemia caused by mutations in adjacent ABC transporters. Science 2000, 290, 1771–1775. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.H.; Liu, M.; Portincasa, P.; Wang, D.Q.H. Recent advances in the critical role of the sterol efflux transporters ABCG5/G8 in health and disease. Adv. Exp. Med. Biol. 2020, 1276, 105–136. [Google Scholar] [PubMed]
- Teupser, D.; Baber, R.; Ceglarek, U.; Scholz, M.; Illig, T.; Gieger, C.; Holdt, L.M.; Leichtle, A.; Greiser, K.H.; Huster, D.; et al. Genetic Regulation of Serum Phytosterol Levels and Risk of Coronary Artery Disease. Circ. Cardiovasc. Genet. 2010, 3, 331–339. [Google Scholar] [CrossRef] [PubMed]
- Berge, K.E.; von Bergmann, K.; Lutjohann, D.; Guerra, R.; Grundy, S.M.; Hobbs, H.H.; Cohen, J.C. Heritability of plasma noncholesterol sterols and relationship to DNA sequence polymorphism in ABCG5 and ABCG8. J. Lipid Res. 2002, 43, 486–494. [Google Scholar] [CrossRef] [PubMed]
- Plat, J.; Bragt, M.C.; Mensink, R.P. Common sequence variations in ABCG8 are related to plant sterol metabolism in healthy volunteers. J. Lipid Res. 2005, 46, 68–75. [Google Scholar] [CrossRef]
- Gylling, H.; Hallikainen, M.; Pihlajamäki, J.; Ågren, J.; Laakso, M.; Rajaratnam, R.A.; Rauramaa, R.; Miettinen, T.A. Polymorphisms in the ABCG5 and ABCG8 genes associate with cholesterol absorption and insulin sensitivity. J. Lipid Res. 2004, 45, 1660–1665. [Google Scholar] [CrossRef]
- gNomad. Available online: https://ldlink.nih.gov/…ldpair (accessed on 12 June 2023).
- Helgadottir, A.; Thorleifsson, G.; Alexandersson, K.F.; Tragante, V.; Thorsteinsdottir, M.; Eiriksson, F.F.; Gretarsdottir, S.; Björnsson, E.; Magnusson, O.; Sveinbjornsson, G.; et al. Genetic variability in the absorption of dietary sterols affects the risk of coronary artery disease. Eur. Heart J. 2020, 41, 2618–2628. [Google Scholar] [CrossRef] [PubMed]
- Kurvinen, A.; Nissinen, M.J.; Gylling, H.; Miettinen, T.A.; Lampela, H.; Koivusalo, A.I.; Rintala, R.J.; Pakarinen, M.P. Effects of long-term parenteral nutrition on serum lipids, plant sterols, cholesterol metabolism, and liver histology in pediatric intestinal failure. J. Pediatr. Gastroenterol. Nutr. 2011, 53, 440–446. [Google Scholar] [CrossRef] [PubMed]
- Pianese, P.; Salvia, G.; Campanozzi, A.; D’Apolito, O.; Russo, A.D.; Pettoello-Mantovani, M.; Corso, G. Sterol profiling in red blood cell membranes and plasma of newborns receiving total parenteral nutrition. J. Pediatr. Gastroenterol. Nutr. 2008, 47, 645–651. [Google Scholar] [CrossRef] [PubMed]
- Clayton, P.T.; Bowron, A.; Mills, K.A.; Massoud, A.; Casteels, M.; Milla, P.J. Phytosterolemia in children with parenteral nutrition—Associated cholestatic liver disease. Gastroenterology 1993, 105, 1806–1813. [Google Scholar] [CrossRef] [PubMed]
- Kurvinen, A.; Nissinen, M.J.; Andersson, S.; Korhonen, P.; Ruuska, T.; Taimisto, M.; Kalliomäki, M.; Lehtonen, L.; Sankilampi, U.; Arikoski, P.; et al. Parenteral plant sterols and intestinal failure–associated liver disease in neonates. J. Pediatr. Gastroenterol. Nutr. 2012, 54, 803–811. [Google Scholar] [CrossRef] [PubMed]
- Llop-Talaveron, J.; Badía-Tahull, M.; Lozano-Andreu, T.; Rigo-Bonnin, R.; Virgili-Casas, N.; Farran-Teixidó, L.; Miró-Martín, M.; Suárez-Lledó, A.; Leiva-Badosa, E. Phytosterolemia and γ-glutamyl transferase in adults with parenteral nutrition: Fish versus vegetal lipids: A randomized clinical trial. Nutrition 2019, 70, 110587. [Google Scholar] [CrossRef] [PubMed]
- Llop-Talaveron, J.; Leiva-Badosa, E.; Alia-Ramos, P.; Rigo-Bonnin, R.; Virgili-Casas, N.; Farran-Teixidor, L.; Miró-Martín, M.; Garrido-Sanchez, L.; Suárez-Lledó, A.; Badía-Tahull, M.B. Genetic factors associated with alterations in liver function test results in adult hospitalized patients treated with parenteral nutrition: A substudy of a clinical trial. Nutrition 2022, 93, 111507. [Google Scholar] [CrossRef] [PubMed]
- Merino, J.; Masana, L.; Guijarro, C.; Ascaso, J.; Lagares, M.; Civeira, F. Recomendaciones para la utilización clínica de los alimentos enriquecidos con fitosteroles/fitostanoles en el manejo de la hipercolesterolemia. Clin. Investig. Arterioscler. 2014, 26, 147–158. [Google Scholar] [PubMed]
- Schroor, M.M.; Mokhtar, F.B.A.; Plat, J.; Mensink, R.P. Associations between SNPs in intestinal cholesterol absorption and endogenous cholesterol synthesis genes with cholesterol metabolism. Biomedicines 2021, 9, 1475. [Google Scholar] [CrossRef]
- Silbernagel, G.; Chapman, M.J.; Genser, B.; Kleber, M.E.; Fauler, G.; Scharnagl, H.; Grammer, T.B.; Boehm, B.O.; Mäkelä, K.-M.; Kähönen, M.; et al. High intestinal cholesterol absorption is associated with cardiovascular disease and risk alleles in ABCG8 and ABO: Evidence from the LURIC and YFS cohorts and from a meta-analysis. J. Am. Coll. Cardiol. 2013, 62, 291–299. [Google Scholar] [CrossRef]
- Jakulj, L.; Vissers, M.N.; Tanck, M.W.; Hutten, B.A.; Stellaard, F.; Kastelein, J.J.; Dallinga-Thie, G.M. ABCG5/G8 polymorphisms and markers of cholesterol metabolism: Systematic review and meta-analysis. J. Lipid Res. 2010, 51, 3016–3023. [Google Scholar] [CrossRef] [PubMed]
- Assmann, G.; Cullen, P.; Erbey, J.; Ramey, D.R.; Kannenberg, F.; Schulte, H. Plasma sitosterol elevations are associated with an increased incidence of coronary events in men: Results of a nested case-control analysis of the Prospective Cardiovascular Münster (PROCAM) study. Nutr. Metab. Cardiovasc. Dis. 2006, 16, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Windler, E.; Zyriax, B.-C.; Kuipers, F.; Linseisen, J.; Boeing, H. Association of plasma phytosterol concentrations with incident coronary heart disease. Data from the CORA study, a case-control study of coronary artery disease in women. Atherosclerosis 2009, 203, 284–290. [Google Scholar] [CrossRef]
- Pinedo, S.; Vissers, M.N.; von Bergmann, K.; Elharchaoui, K.; Lütjohann, D.; Luben, R.; Wareham, N.J.; Kastelein, J.J.P.; Khaw, K.-T.; Boekholdt, S.M. Plasma levels of plant sterols and the risk of coronary artery disease: The prospective EPIC-Norfolk Population Study. J. Lipid Res. 2007, 48, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Weingärtner, O.; Bogeski, I.; Kummerow, C.; Schirmer, S.H.; Husche, C.; Vanmierlo, T.; Wagenpfeil, G.; Hoth, M.; Böhm, M.; Lütjohann, D.; et al. Plant sterol ester diet supplementation increases serum plant sterols and markers of cholesterol synthesis, but has no effect on total cholesterol levels. J. Steroid Biochem. Mol. Biol. 2017, 169, 219–225. [Google Scholar] [CrossRef]
- Fassbender, K.; Lütjohann, D.; Dik, M.G.; Bremmer, M.; König, J.; Walter, S.; Liu, Y.; Letièmbre, M.; von Bergmann, K.; Jonker, C. Moderately elevated plant sterol levels are associated with reduced cardiovascular risk—The LASA study. Atherosclerosis 2008, 196, 283–288. [Google Scholar] [CrossRef]
- Silbernagel, G.; Fauler, G.; Renner, W.; Landl, E.M.; Hoffmann, M.M.; Winkelmann, B.R.; Boehm, B.O.; März, W. The relationships of cholesterol metabolism and plasma plant sterols with the severity of coronary artery disease. J. Lipid Res. 2009, 50, 334–341. [Google Scholar] [CrossRef] [PubMed]
- Ellegård, L.; Sunesson, Å.; Bosaeus, I. High serum phytosterol levels in short bowel patients on parenteral nutrition support. Clin. Nutr. 2005, 24, 415–420. [Google Scholar] [CrossRef] [PubMed]
- Llop, J.M.; Virgili, N.; Moreno-Villares, J.M.; García-Peris, P.; Serrano, T.; Forga, M.; Solanich, J.; Pita, A.M. Phytosterolemia in parenteral nutrition patients: Implications for liver disease development. Nutrition 2008, 24, 1145–1152. [Google Scholar] [CrossRef]
- Gylling, H.; Puska, P.; Vartiainen, E.; Miettinen, T. Retinol, vitamin D, carotenes and α-tocopherol in serum of a moderately hypercholesterolemic population consuming sitostanol ester margarine. Atherosclerosis 1999, 145, 279–285. [Google Scholar] [CrossRef]
- Chalasani, N.; Younossi, Z.; LaVine, J.E.; Diehl, A.M.; Brunt, E.M.; Cusi, K.; Charlton, M.; Sanyal, A.J. The diagnosis and management of non-alcoholic fatty liver disease: Practice Guideline by the American Association for the Study of Liver Diseases, American College of Gastroenterology, and the American Gastroenterological Association. Hepatology 2012, 55, 2005–2023. [Google Scholar] [CrossRef]
- Wanten, G.; Beunk, J.; Naber, A.; Swinkels, D. Tocopherol isoforms in parenteral lipid emulsions and neutrophil activation. Clin. Nutr. 2002, 21, 417–422. [Google Scholar] [CrossRef]
- Pironi, L.; Guidetti, M.; Zolezzi, C.; Fasano, M.C.; Paganelli, F.; Merli, C.; Bersani, G.; Pizzoferrato, A.; Miglioli, M. Peroxidation potential of lipid emulsions after compounding in all-in-one solutions. Nutrition 2003, 19, 784–788. [Google Scholar] [CrossRef]
- Miettinen, T.; Hallikainen, M.; Raitakari, O.; Viikari, J.; Gylling, H. Twenty-one year tracking of serum non-cholesterol sterols. The Cardiovascular Risk in Young Finns study. Nutr. Metab. Cardiovasc. Dis. 2009, 19, 525–531. [Google Scholar] [CrossRef]
- Racette, S.B.; Lin, X.; Ma, L.; Ostlund, J.R.E. Natural Dietary Phytosterols. J. AOAC Int. 2015, 98, 679–684. [Google Scholar] [CrossRef] [PubMed]
- Escurriol, V.; Cofán, M.; Moreno-Iribas, C.; Larrañaga, N.; Martínez, C.; Navarro, C.; Rodríguez, L.; González, C.A.; Corella, D.; Ros, E. Phytosterol plasma concentrations and coronary heart disease in the prospective Spanish EPIC cohort. J. Lipid Res. 2010, 51, 618–624. [Google Scholar] [CrossRef] [PubMed]
- Ras, R.T.; Geleijnse, J.M.; Trautwein, E.A. LDL-cholesterol-lowering effect of plant sterols and stanols across different dose ranges: A meta-analysis of randomised controlled studies. Br. J. Nutr. 2014, 112, 214–219. [Google Scholar] [CrossRef]
- Andersson, S.W.; Skinner, J.; Ellegård, L.; Welch, A.A.; Bingham, S.; Mulligan, A.; Andersson, H.; Shaw, K.-T. Intake of dietary plant sterols is inversely related to serum cholesterol concentration in men and women in the EPIC Norfolk population: A cross-sectional study. Eur. J. Clin. Nutr. 2004, 58, 1378–1385. [Google Scholar] [CrossRef] [PubMed]
- Klingberg, S.; Ellegård, L.; Johansson, I.; Hallmans, G.; Weinehall, L.; Andersson, H.; Winkvist, A. Inverse relation between dietary intake of naturally occurring plant sterols and serum cholesterol in northern Sweden. Am. J. Clin. Nutr. 2008, 87, 993–1001. [Google Scholar] [CrossRef]
- Gylling, H.; Plat, J.; Turley, S.; Ginsberg, H.N.; Ellegård, L.; Jessup, W.; Jones, P.J.H.; Lütjohann, D.; Maerz, W.; Masana, L.; et al. Plant sterols and plant stanols in the management of dyslipidaemia and prevention of cardiovascular disease. Atherosclerosis 2014, 232, 346–360. [Google Scholar] [CrossRef]
- Heinemann, T.; Axtmann, G.; Von Bergmann, K. Comparison of intestinal absorption of cholesterol with different plant sterols in man. Eur. J. Clin. Investig. 1993, 23, 827–831. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, H.; Tada, H.; Ito, K.; Kishimoto, Y.; Yanai, H.; Okamura, T.; Ikewaki, K.; Inagaki, K.; Shoji, T.; Bujo, H.; et al. Reference intervals of serum non-cholesterol sterols by gender in healthy Japanese individuals. J. Atheroscler. Thromb. 2020, 27, 409–417. [Google Scholar] [CrossRef] [PubMed]
- Wu, A.H.; Ruan, W.; Todd, J.; Lynch, K.L. Biological variation of β-sitosterol, campesterol, and lathosterol as cholesterol absorption and synthesis biomarkers. Clin. Chim. Acta 2014, 430, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Salen, G.; Ahrens, E.H.; Grundy, S.M. Metabolism of beta-sitosterol in man. J. Clin. Investig. 1970, 49, 952–967. [Google Scholar] [CrossRef]
- Miettinen, T.A.; Tilvis, R.S.; Kesäniemi, Y.A. Serum plant sterols and cholesterol precursors reflect cholesterol absorption and synthesis in volunteers of a randomly selected male population. Am. J. Epidemiol. 1990, 131, 20–31. [Google Scholar] [CrossRef]

| Variants (Gene) | Ref Allele > Alt allele (Allele Frequency) [11] * | Effect of Alt Allele on Plasma PS Levels | References |
|---|---|---|---|
| rs41360247 (ABCG8) | T > C (C = 0.0604) | Lower levels | [7] |
| rs4245791 (ABCG8) | C > T (C = 0.3195) | Higher levels | [7] |
| rs4148217 (ABCG8) | C > A, T (A = 0.1901) | Lower levels | [8,9,12] |
| rs11887534 (ABCG8) | G > C (C = 0.0651) | Lower levels | [8,10,12] |
| rs657152 (ABO) | A > C, T (A = 0.3915) | Higher levels | [7] |
| Variable | Total (n = 185) Mean ± SD/N (%) | PS ≤ P50 (n = 93) Mean ± SD/N (%) | PS > P50 (n = 92) Mean ± SD/N (%) | p * |
|---|---|---|---|---|
| Demographic and anthropometric data | ||||
| Age (years) | 41.49 ± 18.57 | 42.82 ± 19.07 | 40.14 ± 18.07 | 0.164 |
| BMI (kg/m2) | 24.47 ± 4.28 | 25.23 ± 4.83 | 23.71 ± 3.51 | 0.008 |
| Sex (women) | 115 (62.20%) | 56 (60.20%) | 59 (64.10%) | 0.583 |
| Dietary habits | ||||
| Vegetarian-vegan (yes) | 9 (4.90%) | 4 (4.30%) | 5 (5.40%) | 0.720 |
| Nuts | 0.125 ** | |||
| No consumption | 58 (31.40%) | 32 (34.4%) | 26 (28.30%) | |
| 1–3 times a week | 99 (53.50%) | 51 (54.80%) | 48 (52.20%) | |
| >3 times a week | 28 (15.10%) | 10 (10.80%) | 18 (19.60%) | |
| Pastries | 0.069 ** | |||
| No consumption | 94 (50.80%) | 43 (46.20%) | 51 (55.40%) | |
| 1–3 times a week | 78 (42.20%) | 40 (43.00%) | 38 (41.30%) | |
| >3 times a week | 13 (7.00%) | 10 (10.80%) | 3 (3.30%) | |
| Sausages | 0.632 ** | |||
| No consumption | 24 (13.00%) | 13 (14.00%) | 11 (12.00%) | |
| 1–3 times a week | 103 (55.70%) | 52 (55.90%) | 51 (55.40%) | |
| >3 times a week | 58 (31.40%) | 28 (30.10%) | 30 (32.60%) | |
| FI (0–6) | 2.05 ± 1.16 | 2.06 ± 1.24 | 2.04 ± 1.07 | 0.451 |
| VII (0–6) | 1.22 ± 1.33 | 0.98 ± 1.11 | 1.34 ± 1.46 | 0.031 |
| Cholesterol treatment and dietary supplementation | ||||
| Cholesterol-lowering treatment (Yes) | 16 (8.6%) | 9 (9.7%) | 7 (7.6%) | 0.617 |
| PS-enriched supplements (Yes) | 8 (4.30%) | 2 (2.2%) | 6 (6.5%) | 0.144 |
| Vitamins (Yes) | 12 (6.50%) | 6 (6.50%) | 6 (6.50%) | 0.985 |
| Plasma values | ||||
| Total cholesterol (mg/dL) | 101.92 ± 57.65 | 88.18 ± 54.63 | 115.81 ± 57.57 | <0.001 |
| Alanine aminotransferase (µKat/L) | 0.29 ± 0.13 | 0.29 ± 0.14 | 0.28 ± 0.12 | 0.323 |
| Alkaline phosphatase (µKat/L) | 0.98 ± 0.28 | 1.00 ± 0.30 | 0.96 ± 0.25 | 0.184 |
| α-Tocopherol (µmol/L) | 30.25 ± 6.20 | 30.12 ± 5.99 | 30.39 ± 6.42 | 0.380 |
| Creatinine (µmol/L) | 76.82 ± 16.83 | 78.24 ± 17.67 | 75.38 ± 15.89 | 0.125 |
| Albumin (g/L) | 47.74 ± 3.29 | 47.82 ± 3.34 | 47.66 ± 3.26 | 0.375 |
| Polymorphism | Genotypes (N/%) | ||
|---|---|---|---|
| rs41360247 | CC | TC | TT |
| 0 (0%) | 26 (14.1%) | 159 (85.9%) | |
| rs4245791 | CC | TC | TT |
| 19 (10.3%) | 79 (42.7%) | 87 (47.0%) | |
| rs4148217 | AA | CA | CC |
| 5 (2.7%) | 58 (31.4%) | 122 (65.9%) | |
| rs11887534 | CC | GC | GG |
| 0 (0%) | 27 (14.6%) | 158 (85.4%) | |
| rs657152 | TT | GT | GG |
| 29 (15.7%) | 90 (48.6%) | 66 (35.7%) | |
| Variable | Total PS | Sitosterol | Campesterol | |||
|---|---|---|---|---|---|---|
| B [95% CI] | p | B [95% CI] | p | B [95% CI] | p | |
| Cholesterol (mg/dL) (×104) | 10.00 [0.00–10.00] | <0.001 | 0.00 [0.00–10.00] | <0.001 | 0.00 [−0.18–10] | 0.065 |
| rs4245791_TT vs. rs4245791_CC | −0.57 [−1.28 –0.15] | 0.119 | −0.16 [−0.47–0.15] | 0.311 | −0.48 [−0.93–(−0.03)] | 0.037 |
| rs4245791_TC vs. rs4245791_CC | −0.41 [−1.10–0.28] | 0.244 | −0.23 [−0.52–0.07] | 0.137 | −0.20 [−0.63–0.22] | 0.344 |
| rs4148217_AA vs. rs4148217_CC | 0.10 [−1.16–1.36] | 0.880 | −0.16 [−0.70–0.39] | 0.577 | 0.40 [−0.33–1.13] | 0.282 |
| rs4148217_AC vs. rs4148217_CC | −0.38 [−0.83–0.06] | 0.093 | −0.19 [−0.38–0.01] | 0.059 | −0.12 [−0.42–0.18] | 0.443 |
| rs6577152_TT vs. rs6577152_GG | 0.29 [−0.34–0.92] | 0.366 | 0.08 [−0.20–0.35] | 0.584 | 0.26 [0.02–0.51] | 0.035 |
| rs6577152_GT vs. rs6577152_GG | 0.52 [0.07–0.96] | 0.022 | 0.17 [−0.02–0.36] | 0.074 | 0.36 [0.07–0.65] | 0.016 |
| Sex (women) | 0.20 [−0.34–0.74] | 0.477 | 0.19 [−0.04–0.42] | 0.109 | −0.02 [−0.43–0.38] | 0.907 |
| Age (years) (×10) | 0.23 [0.09–0.37] | 0.002 | 0.02 [−0.04–0.08] | 0.491 | 0.17 [0.09–0.25] | <0.001 |
| BMI (kg/m2) (×10) | −0.57 [−1.10–(−0.04)] | 0.034 | −0.25 [−0.47–(−0.02)] | 0.033 | −0.37 [−0.69–(−0.04)] | 0.029 |
| Cholesterol-lowering treatment (Yes) | −0.10 [−0.94–0.74] | 0.813 | 0.05 [−0.31–0.41] | 0.791 | 0.04 [−0.43–0.51] | 0.870 |
| Vitamins (Yes) | −0.17 [−1.00–0.65] | 0.685 | 0.04 [−0.32–0.40] | 0.827 | −0.19 [−0.58–0.20] | 0.349 |
| Alanine aminotransferase (µKat/L) | 0.22 [−1.45–1.90] | 0.794 | 0.27 [−0.46–1.00] | 0.468 | −0.20 [−0.89–0.49] | 0.566 |
| Alkaline phosphatase (µKat/L) | 0.58 [−0.22–1.38] | 0.152 | 0.12 [−0.23–0.46] | 0.512 | 0.50 [−0.34–1.33] | 0.247 |
| Creatinine (µmol/L) (×104) | −80.00 [−220.00–70.00] | 0.303 | 0.01 [−60.00–60.00] | 1.000 | −80.00 [−180.00–20.00] | 0.127 |
| FI | −0.06 [−0.24–0.12] | 0.494 | −0.04 [−0.12–0.04] | 0.300 | −0.04 [−0.16–0.09] | 0.551 |
| VII | 0.03 [−0.13–0.18] | 0.755 | 0.03 [−0.03–0.10] | 0.334 | −0.02 [−0.10–0.07] | 0.723 |
| α-Tocopherol (µmol/L) (×10) | −0.21 [−0.55–0.13] | 0.222 | −0.02 [−0.17–0.12] | 0.757 | −0.19 [−0.39–0.01] | 0.057 |
| Omnibus χ2 | 41.38 | <0.001 | 54.50 | <0.001 | 43.95 | <0.001 |
| R2 | 0.20 | 0.21 | 0.25 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garrido-Sanchez, L.; Leiva-Badosa, E.; Llop-Talaveron, J.; Pintó-Sala, X.; Lozano-Andreu, T.; Corbella-Inglés, E.; Alia-Ramos, P.; Arias-Barquet, L.; Ramon-Torrel, J.M.; Badía-Tahull, M.B. Blood Phytosterol Concentration and Genetic Variant Associations in a Sample Population. Nutrients 2024, 16, 1067. https://doi.org/10.3390/nu16071067
Garrido-Sanchez L, Leiva-Badosa E, Llop-Talaveron J, Pintó-Sala X, Lozano-Andreu T, Corbella-Inglés E, Alia-Ramos P, Arias-Barquet L, Ramon-Torrel JM, Badía-Tahull MB. Blood Phytosterol Concentration and Genetic Variant Associations in a Sample Population. Nutrients. 2024; 16(7):1067. https://doi.org/10.3390/nu16071067
Chicago/Turabian StyleGarrido-Sanchez, Leticia, Elisabet Leiva-Badosa, Josep Llop-Talaveron, Xavier Pintó-Sala, Toni Lozano-Andreu, Emili Corbella-Inglés, Pedro Alia-Ramos, Lluis Arias-Barquet, Josep Maria Ramon-Torrel, and Maria B. Badía-Tahull. 2024. "Blood Phytosterol Concentration and Genetic Variant Associations in a Sample Population" Nutrients 16, no. 7: 1067. https://doi.org/10.3390/nu16071067