
The Effect of an Elevated Dietary Copper Level on the Vascular Contractility and Oxidative Stress in Middle-Aged Rats

 ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Drugs and Chemicals
2.2. Animals and Experimental Treatments
2.3. Vascular Reactivity Studies
2.4. The Langendorff Heart
2.5. Atomic Absorption Spectroscopy (AAS) for Cu, Zn, Fe, Se in Rat Serum and Liver
2.6. Total Antioxidant Status (TAS)
2.7. ELISA Protocol
2.8. Cu and Zn Determination in Rats’ Feed
2.9. Body Composition Analysis
2.10. NO, O2•− and H2O2 Detection
2.11. Data Analysis and Statistics
3. Results
3.1. Copper and Zinc Concentration in Rats’ Feed (Atomic Absorption Spectroscopy—AAS)
3.2. Animal Weight Gain and Body Composition (Time-Domain NMR)
3.3. The Langendorff Heart
3.4. Vascular Contraction
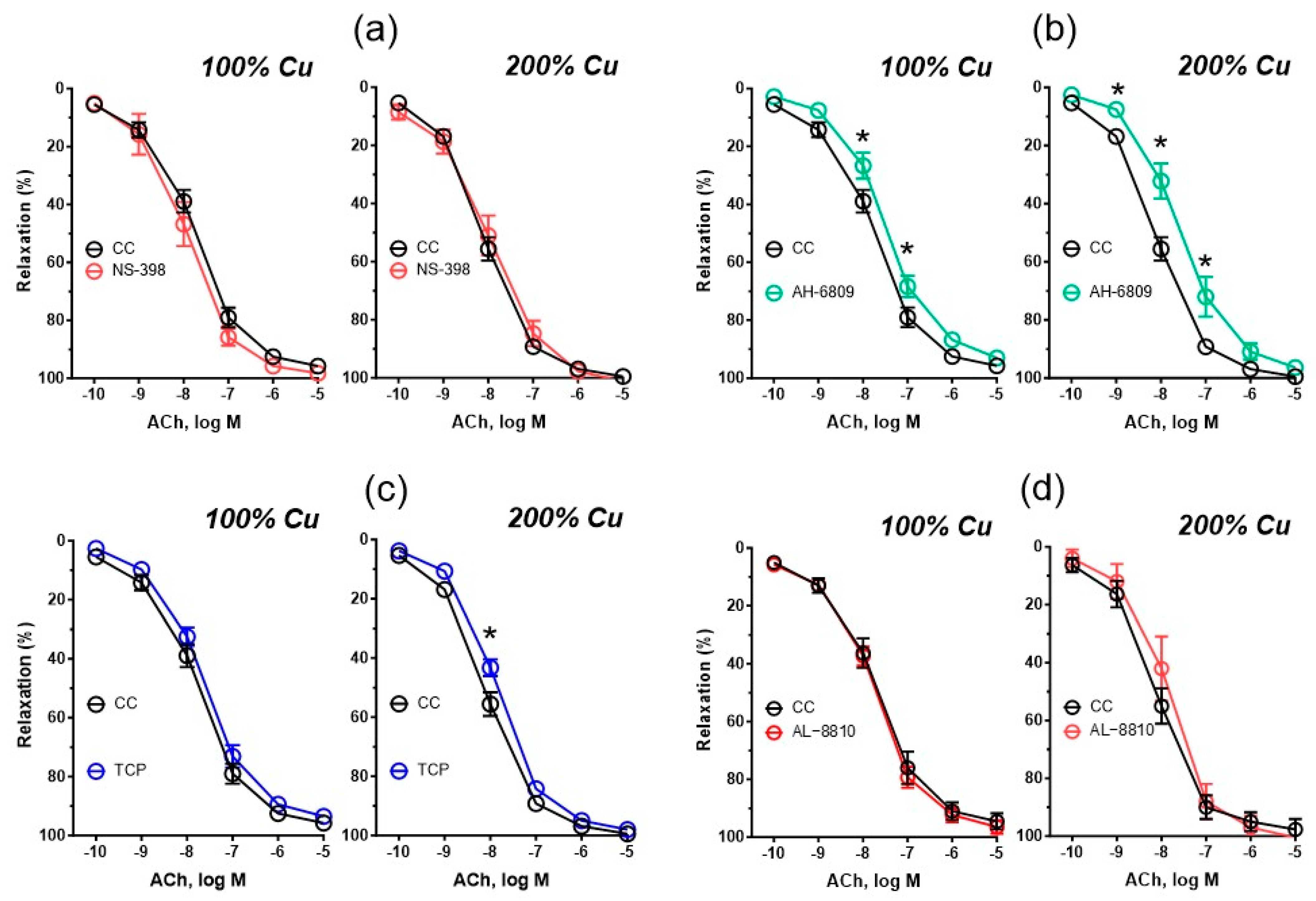
3.5. Vascular Relaxation
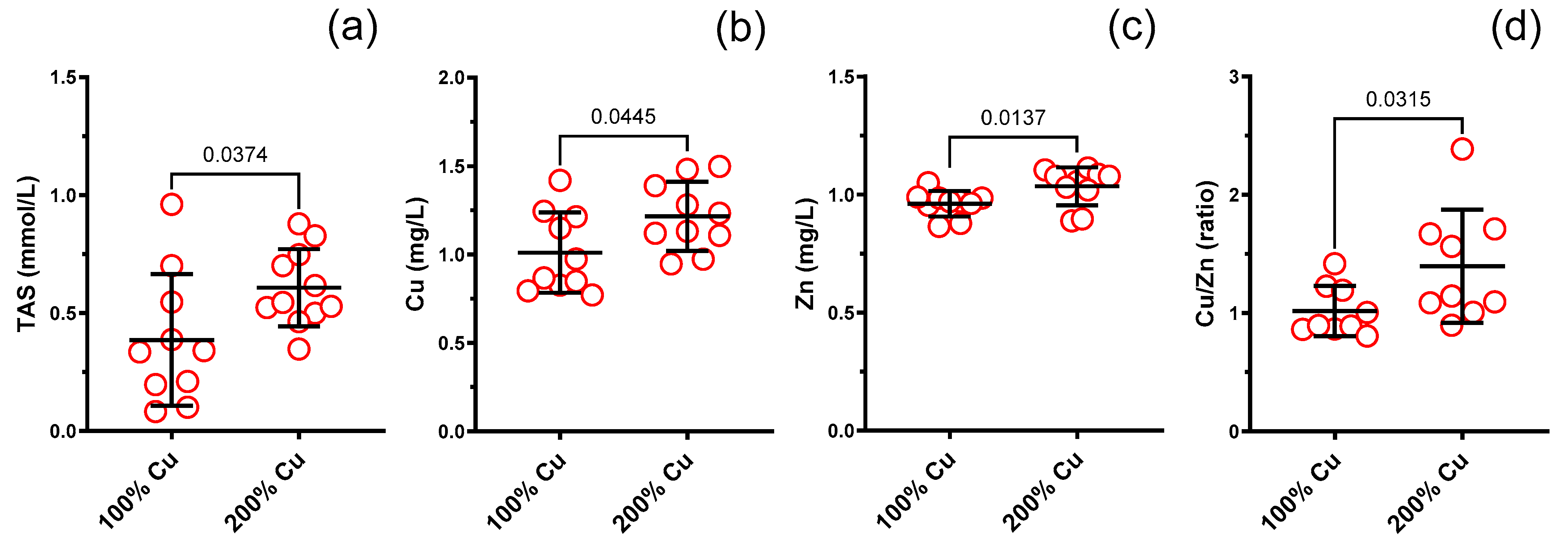
3.6. Total Antioxidant Status (TAS) in the Blood Serum
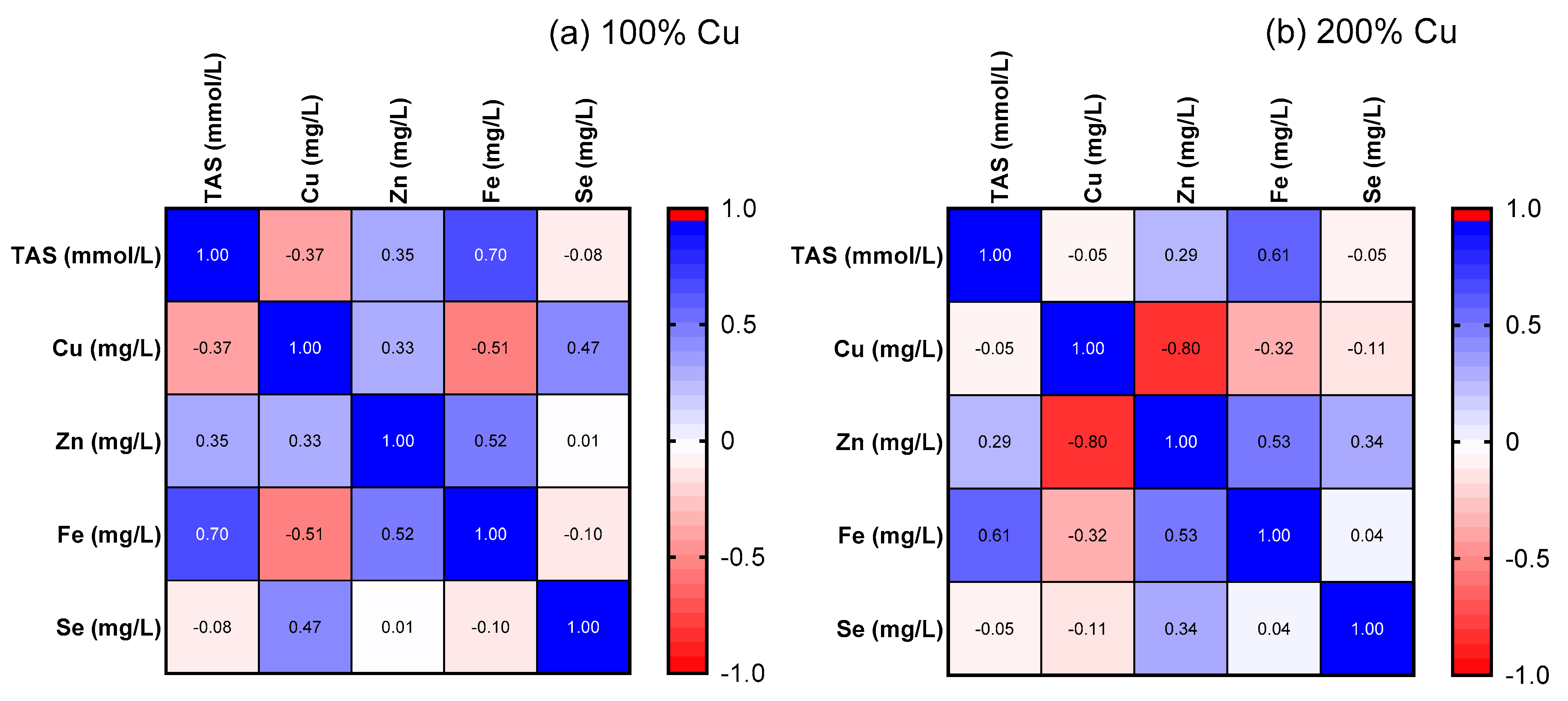
3.7. Cu, Zn, Fe, Se in the Rat Serum (AAS)
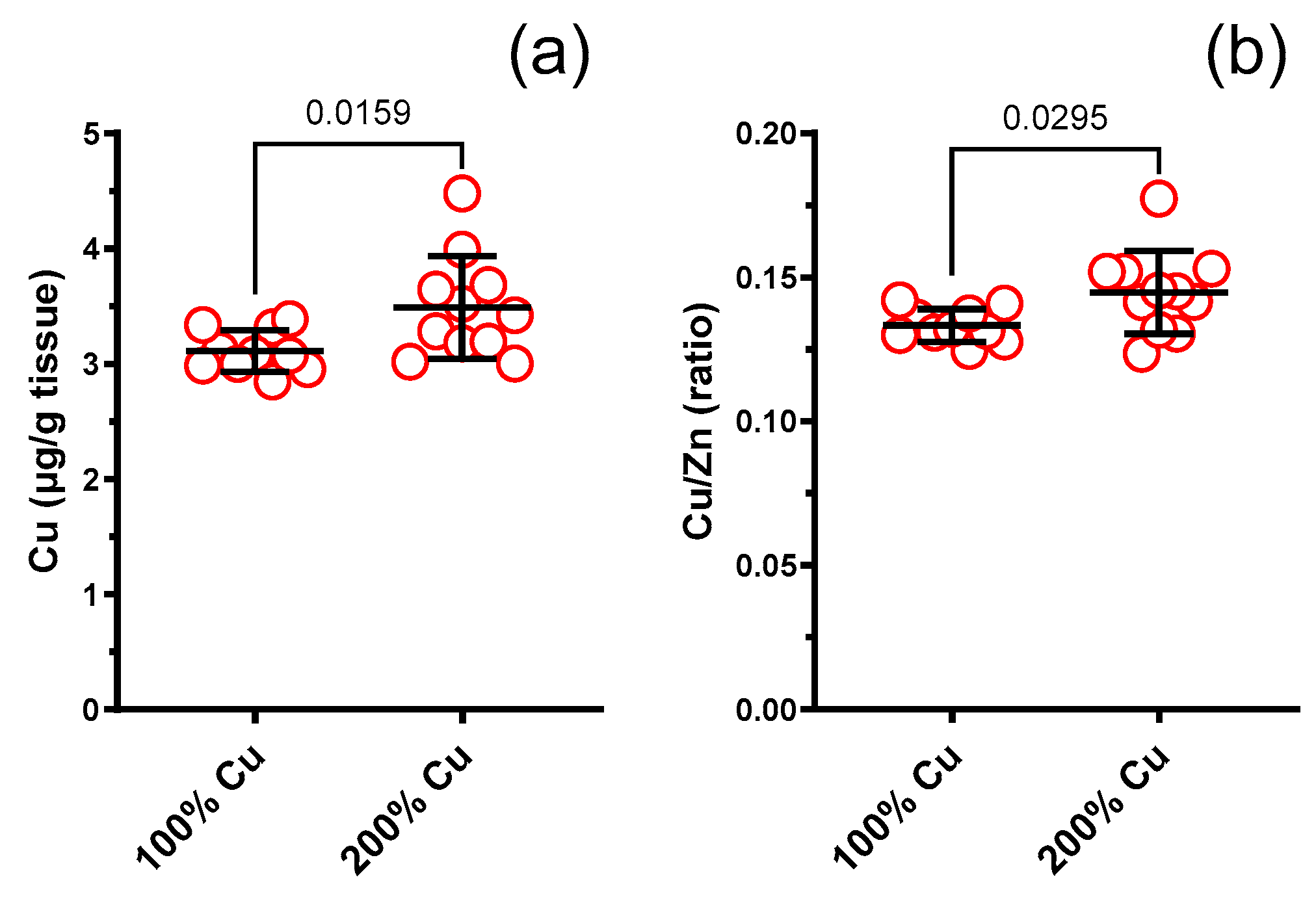
3.8. Cu, Zn, Fe, Se in Rat Liver (AAS)
3.9. ELISA Studies
3.10. NO, O2•− and H2O2 Detection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Prohaska, J.R. Copper. In Present Knowledge in Nutrition, 10th ed.; Erdman, J.W., Macdonald, I.A., Zeisel, S.H., Eds.; Wiley−Blackwell: Washington, DC, USA, 2012. [Google Scholar]
- Collins, J.F. Copper. In Modern Nutrition in Health and Disease, 11th ed.; Ross, A.C., Caballero, B., Cousins, R.J., Tucker, K.L., Ziegler, T.R., Eds.; Lippincott Williams & Wilkins: Baltimore, MD, USA, 2014. [Google Scholar]
- Fukai, T.; Ushio−Fukai, M.; Kaplan, J.H. Copper transporters and copper chaperones: Roles in cardiovascular physiology and disease. Am. J. Physiol. Cell Physiol. 2018, 315, 186–201. [Google Scholar] [CrossRef]
- Jomova, K.; Valko, M. Advances in metal−induced oxidative stress and human disease. Toxicology 2011, 283, 65–87. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, R.; Ramond, A.; O’Keeffe, L.M.; Shahzad, S.; Kunutsor, S.K.; Muka, T.; Gregson, J.; Willeit, P.; Warnakula, S.; Khan, H.; et al. Environmental toxic metal contaminants and risk of cardiovascular disease: Systematic review and meta−analysis. BMJ 2018, 362, k3310. [Google Scholar] [CrossRef]
- Fry, R.S.; Ashwell, M.S.; Lloyd, K.E.; O’Nan, A.T.; Flowers, W.L.; Stewart, K.R.; Spears, J.W. Amount and source of dietary copper affects small intestine morphology, duodenal lipid peroxidation, hepatic oxidative stress, and mRNA expression of hepatic copper regulatory proteins in weanling pigs. J. Anim. Sci. 2012, 90, 3112–3119. [Google Scholar] [CrossRef]
- Kang, Y.J. Copper and homocysteine in cardiovascular diseases. Pharmacol. Ther. 2011, 129, 321–331. [Google Scholar] [CrossRef]
- Konukoğlu, D.; Serin, O.; Ercan, M.; Turhan, M.S. Plasma homocysteine levels in obese and non−obese subjects with or without hypertension; its relationship with oxidative stress and copper. Clin. Biochem. 2003, 36, 405–408. [Google Scholar] [CrossRef]
- Nunes, K.Z.; Fioresi, M.; Marques, V.B.; Vassallo, D.V. Acute copper overload induces vascular dysfunction in aortic rings due to endothelial oxidative stress and increased nitric oxide production. J. Toxicol. Environ. Health A 2018, 81, 218–228. [Google Scholar] [CrossRef]
- Steven, S.; Frenis, K.; Oelze, M.; Kalinovic, S.; Kuntic, M.; Bayo Jimenez, M.T.; Vujacic−Mirski, K.; Helmstädter, J.; Kröller−Schön, S.; Münzel, T.; et al. Vascular Inflammation and Oxidative Stress: Major Triggers for Cardiovascular Disease. Oxid. Med. Cell Longev. 2019, 2019, 7092151. [Google Scholar] [CrossRef]
- Guzik, T.J.; Touyz, R.M. Oxidative Stress, Inflammation, and Vascular Aging in Hypertension. Hypertension 2017, 70, 660–667. [Google Scholar] [CrossRef]
- DiSilvestro, R.A.; Joseph, E.L.; Zhang, W.; Raimo, A.E.; Kim, Y.M. A randomized trial of copper supplementation effects on blood copper enzyme activities and parameters related to cardiovascular health. Metabolism 2012, 61, 1242–1246. [Google Scholar] [CrossRef]
- Majewski, M.; Gromadziński, L.; Cholewińska, E.; Ognik, K.; Fotschki, B.; Juśkiewicz, J. The Interaction of Dietary Pectin, Inulin, and Psyllium with Copper Nanoparticle Induced Changes to the Cardiovascular System. Nutrients 2023, 15, 3557. [Google Scholar] [CrossRef]
- Majewski, M.; Ognik, K.; Juśkiewicz, J. Copper nanoparticles modify the blood plasma antioxidant status and modulate the vascular mechanisms with nitric oxide and prostanoids involved in Wistar rats. Pharmacol. Rep. 2019, 71, 509–516. [Google Scholar] [CrossRef] [PubMed]
- Majewski, M.; Ognik, K.; Juśkiewicz, J. The interaction between resveratrol and two forms of copper as carbonate and nanoparticles on antioxidant mechanisms and vascular function in Wistar rats. Pharmacol. Rep. 2019, 71, 862–869. [Google Scholar] [CrossRef]
- National Research Council (US) Subcommittee on Laboratory Animal Nutrition. 2, Nutrient Requirements of the Laboratory Rat. In Nutrient Requirements of Laboratory Animals, 4th ed.; National Academies Press (US): Washington, DC, USA, 1995. Available online: https://www.ncbi.nlm.nih.gov/books/NBK231925 (accessed on 14 April 2024).
- Majewski, M.; Ognik, K.; Juśkiewicz, J. The antioxidant status, lipid profile, and modulation of vascular function by fish oil supplementation in nano-copper and copper carbonate fed Wistar rats. J. Funct. Foods 2020, 64, 103595. [Google Scholar] [CrossRef]
- Majewski, M.; Klett-Mingo, M.; Verdasco-Martín, C.M.; Otero, C.; Ferrer, M. Spirulina extract improves age-induced vascular dysfunction. Pharm. Biol. 2022, 60, 627–637. [Google Scholar] [CrossRef]
- Yan, M.; Liu, D.L.; Chua, Y.L.; Chen, C.; Lim, Y.L. Effects of micromolar concentrations of manganese, copper, and zinc on alpha1−adrenoceptor−mediating contraction in rat aorta. Biol. Trace Elem. Res. 2001, 82, 159–166. [Google Scholar] [CrossRef]
- Wang, Y.C.; Hu, C.W.; Liu, M.Y.; Jiang, H.C.; Huo, R.; Dong, D.L. Copper induces vasorelaxation and antagonizes noradrenaline−induced vasoconstriction in rat mesenteric artery. Cell Physiol. Biochem. 2013, 32, 1247–1254. [Google Scholar] [CrossRef]
- Afrin, F.; Basir, S.F.; Khan, L.A. Copper−promoted hypercontraction of rat aortic rings and its mitigation by natural molecules. 2022; Preprint. [Google Scholar] [CrossRef]
- Tian, J.; Yan, Z.; Wu, Y.; Zhang, S.L.; Wang, K.; Ma, X.R.; Guo, L.; Wang, J.; Zuo, L.; Liu, J.Y.; et al. Inhibition of iNOS protects endothelial−dependent vasodilation in aged rats. Acta Pharmacol. Sin. 2010, 31, 1324–1328. [Google Scholar] [CrossRef]
- Holowatz, L.A.; Thompson, C.S.; Minson, C.T.; Kenney, W.L. Mechanisms of acetylcholine−mediated vasodilatation in young and aged human skin. J. Physiol, 2005; 563, Pt 3, 965–973. [Google Scholar] [CrossRef]
- Lamb, D.J.; Tickner, M.L.; Hourani, S.M.; Ferns, G.A. Dietary copper supplements modulate aortic superoxide dismutase, nitric oxide and atherosclerosis. Int. J. Exp. Pathol. 2005, 86, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Chiarugi, A.; Pitari, G.M.; Costa, R.; Ferrante, M.; Villari, L.; Amico−Roxas, M.; Godfraind, T.; Bianchi, A.; Salomone, S. Effect of prolonged incubation with copper on endothelium−dependent relaxation in rat isolated aorta. Br. J. Pharmacol. 2002, 136, 1185–1193. [Google Scholar] [CrossRef] [PubMed]
- Emsley, A.M.; Jeremy, J.Y.; Gomes, G.N.; Angelini, G.D.; Plane, F. Investigation of the inhibitory effects of homocysteine and copper on nitric oxide−mediated relaxation of rat isolated aorta. Br. J. Pharmacol. 1999, 126, 1034–1040. [Google Scholar] [CrossRef]
- Nelson, S.K.; Huang, C.J.; Mathias, M.M.; Allen, K.G. Copper−marginal and copper−deficient diets decrease aortic prostacyclin production and copper−dependent superoxide dismutase activity, and increase aortic lipid peroxidation in rats. J. Nutr. 1992, 122, 2101–2108. [Google Scholar] [CrossRef]
- Majewski, M.; Juśkiewicz, J.; Krajewska−Włodarczyk, M.; Gromadziński, L.; Socha, K.; Cholewińska, E.; Ognik, K. The Role of 20−HETE, COX, Thromboxane Receptors, and Blood Plasma Antioxidant Status in Vascular Relaxation of Copper−Nanoparticle−Fed WKY Rats. Nutrients 2021, 13, 3793. [Google Scholar] [CrossRef]
- Mitchell, J.A.; Kirkby, N.S.; Ahmetaj−Shala, B.; Armstrong, P.C.; Crescente, M.; Ferreira, P.; Lopes Pires, M.E.; Vaja, R.; Warner, T.D. Cyclooxygenases and the cardiovascular system. Pharmacol. Ther. 2021, 217, 107624. [Google Scholar] [CrossRef]
- Schuschke, D.A.; Adeagbo, A.S.; Patibandla, P.K.; Egbuhuzo, U.; Fernandez−Botran, R.; Johnson, W.T. Cyclooxygenase−2 is upregulated in copper−deficient rats. Inflammation 2009, 32, 333–339. [Google Scholar] [CrossRef]
- Nicholls, C.; Li, H.; Liu, J.P. GAPDH: A common enzyme with uncommon functions. Clin. Exp. Pharmacol. Physiol. 2012, 39, 674–679. [Google Scholar] [CrossRef]
- Khan, F.; Choong, W.L.; Du, Q.; Jovanovi’c, A. Real−time RT−PCR Ct values for blood GAPDH correlate with measures of vascular endothelial function in humans. Clin. Transl. Sci. 2013, 6, 481–484. [Google Scholar] [CrossRef]
- Young, I.S. Measurement of total antioxidant capacity. J. Clin. Pathol. 2001, 54, 339. [Google Scholar] [CrossRef]
- Bo, S.; Durazzo, M.; Gambino, R.; Berutti, C.; Milanesio, N.; Caropreso, A.; Gentile, L.; Cassader, M.; Cavallo−Perin, P.; Pagano, G. Associations of dietary and serum copper with inflammation, oxidative stress, and metabolic variables in adults. J. Nutr. 2008, 138, 305–310. [Google Scholar] [CrossRef]
- Osredkar, J.; Sustar, N. Copper and zinc, biological role and significance of copper/zinc imbalance. J. Clinic Toxicol. 2011, S3, 0495. [Google Scholar] [CrossRef]
- McClain, C.J.; McClain, M.; Barve, S.; Boosalis, M.G. Trace metals and the elderly. Clin. Geriatr. Med. 2002, 18, 801–818. [Google Scholar] [CrossRef] [PubMed]
- Agunbiade, J.A.; Babatunde, G.M. Copper and iron supplementation in the tropical environment effects on haematological measurements, organ weights, tissue copper and iron. Niger. J. Anim. Prod. 1995, 22, 49–55. [Google Scholar] [CrossRef]
- Filetti, F.M.; Schereider, I.R.G.; Wiggers, G.A.; Miguel, M.; Vassallo, D.V.; Simões, M.R. Cardiovascular Harmful Effects of Recommended Daily Doses (13 µg/kg/day), Tolerable Upper Intake Doses (0.14 mg/kg/day) and Twice the Tolerable Doses (0.28 mg/kg/day) of Copper. Cardiovasc. Toxicol. 2023, 23, 218–229. [Google Scholar] [CrossRef] [PubMed]
- Banach, W.; Nitschke, K.; Krajewska, N.; Mongiałło, W.; Matuszak, O.; Muszyński, J.; Skrypnik, D. The Association between Excess Body Mass and Disturbances in Somatic Mineral Levels. Int. J. Mol. Sci. 2020, 21, 7306. [Google Scholar] [CrossRef]
- Cunnane, S.C.; McAdoo, K.R.; Prohaska, J.R. Lipid and Fatty Acid Composition of Organs from Copper−Deficient Mice. J. Nutr. 1986, 116, 1248–1256. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (%) | g/kg of Diet | |||
|---|---|---|---|---|
| Casein | 20 | 200 | ||
| DL-methionine | 0.3 | 3.0 | ||
| Cellulose | 5.0 | 50 | ||
| Sucrose | 10 | 100 | ||
| Rapeseed oil | 2.0 | 20 | ||
| Lard | 6.0 | 60 | ||
| Vitamin mixture | 1.0 | 10 | ||
| Mineral mixture * | 3.5 | 35 † | ||
| Choline chloride | 0.2 | 2.0 | ||
| Corn starch | 52 (to 100%) | 520 | ||
| * Mineral mixture composition | % of mineral mixture | g/kg of diet | ||
| Calcium carbonate (CaCO3) | 35.7 | 12.495 | ||
| Single-basic potassium phosphate (K2HPO4) | 19.6 | 6.86 | ||
| Potassium citrate (K3C6H5O7) | 7.078 | 2.4773 | ||
| Sodium chloride (NaCl) | 7.4 | 2.59 | ||
| Potassium sulfate (K2SO4) | 4.66 | 1.631 | ||
| Magnesium oxide (MgO) | 2.4 | 0.84 | ||
| Micro mix ** | 1.8 | 0.63 ‡ | ||
| Corn starch | 21.362 (to 100%) | 7.4767 (to 35.0 †) | ||
| ** Micro mineral mixture composition | % of Diet A | % of Diet B | mg/kg of Diet A | mg/kg of Diet B |
| Iron citrate [16.7% Fe] | 31.0 | 31.0 | 195.3 | 195.3 |
| ZnCO3 [56% Zn] | 4.5 | 4.5 | 28.35 | 28.35 |
| MnCO3 [44.4% Mn] | 23.4 | 23.4 | 147.42 | 147.42 |
| CuCO3 [55.5% Cu] | 1.85 | 3.7 | 11.655 | 23.31 |
| KJ | 0.04 | 0.04 | 0.252 | 0.252 |
| Citric acid | 39.21 (to 100%) | 37.36 (to 100%) | 247.023 (to 630 ‡) | 235.368 (to 630 ‡) |
| 100%—Group A *† | 200%—Group B † | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| n | Mean | “Std. Deviation” | n | Mean | “Std. Deviation” | t Test | p Value | x-Fold | |
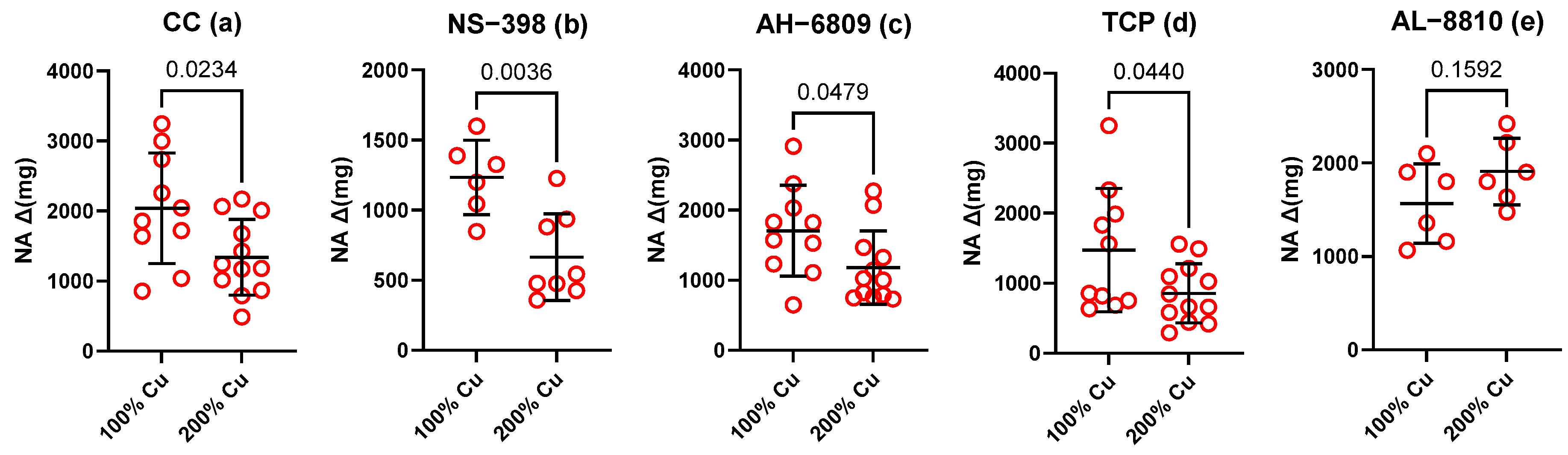
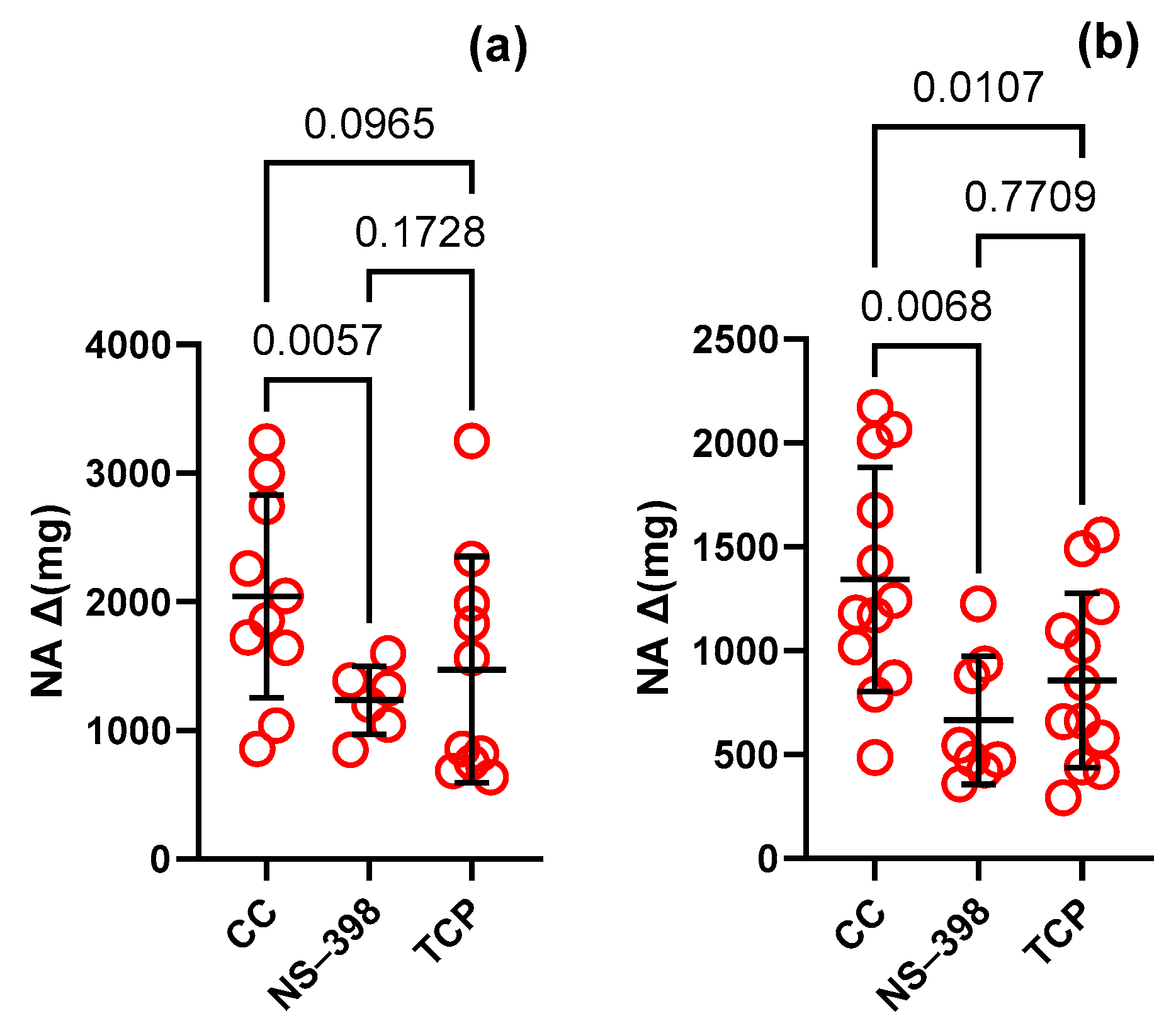
| NA induced contraction Δ(mg) | 9 | 2040 | 789.3 | 9 | 1342 | 539.6 | Unpaired t test | 0.0303 | 0.7 |
| NS-398 Δ(mg) | 6 | 1234 | 265.9 | 8 | 665.7 | 309.3 | Unpaired t test | 0.0036 | 0.5 |
| AH-6809 Δ(mg) | 10 | 1706 | 648.9 | 12 | 1181 | 521.3 | Unpaired t test | 0.0479 | 0.7 |
| TCP Δ(mg) | 10 | 1471 | 879.9 | 12 | 856.5 | 420.2 | Unpaired t test | 0.0440 | 0.6 |
| AL-8810 Δ(mg) | 6 | 1422 | 467.1 | 6 | 1938 | 453.1 | Unpaired t test | 0.1592 | 1.4 |
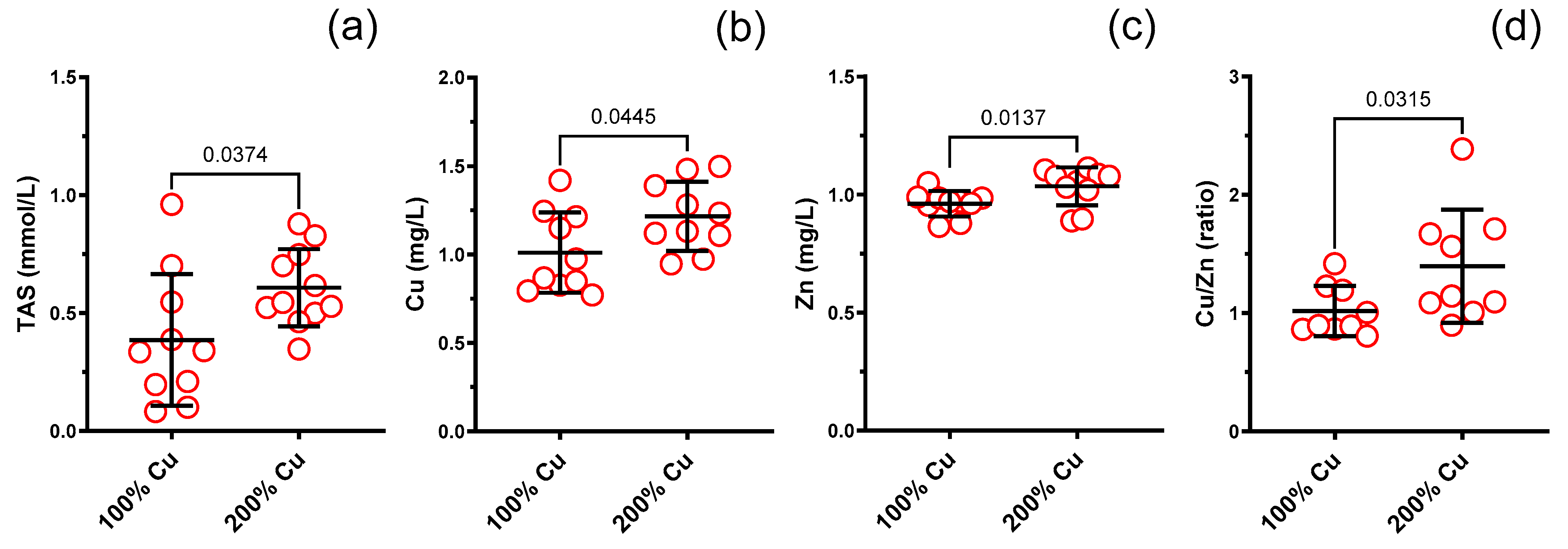
| TAS (mmol/L) in rat serum | 10 | 0.386 | 0.279 | 11 | 0.607 | 0.164 | Unpaired t test | 0.0374 | 1.6 |
| Cu (mg/L) in rat serum | 10 | 1.011 | 0.227 | 10 | 1.216 | 0.195 | Unpaired t test | 0.0445 | 1.2 |
| Zn (mg/L) in rat serum | 10 | 0.961 | 0.054 | 10 | 1.020 | 0.080 | Mann–Whitney test | 0.0137 | 1.1 |
| Cu/Zn molar ratio in rat serum | 9 | 1.018 | 0.211 | 9 | 1.396 | 0.479 | Mann–Whitney test | 0.0315 | 1.4 |
| Cu (µg/g) in the liver | 10 | 3.110 | 0.180 | 11 | 3.492 | 0.446 | Mann–Whitney test | 0.0159 | 1.12 |
| Cu/Zn molar ratio in the liver | 10 | 0.133 | 0.006 | 11 | 0.145 | 0.014 | Mann–Whitney test | 0.0295 | 1.09 |
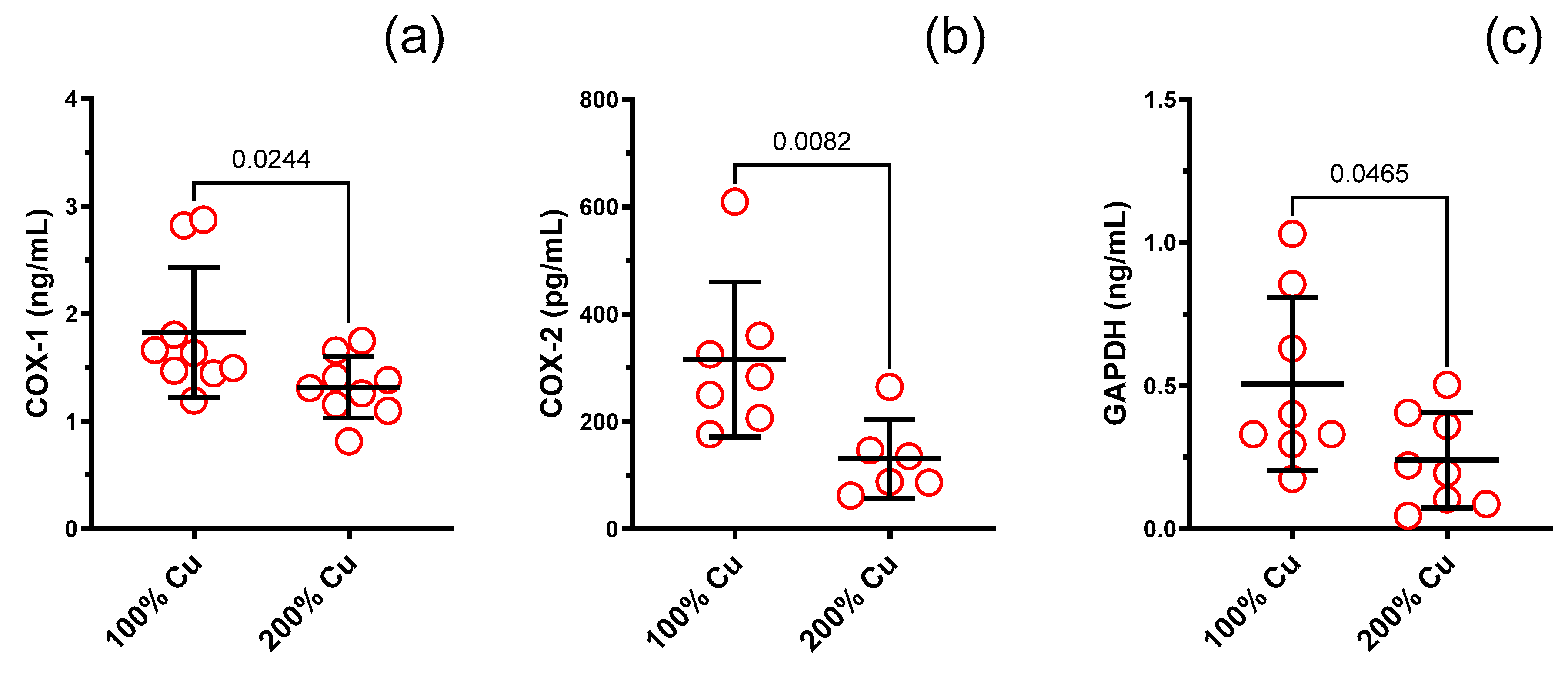
| COX-1 (ng/mL) in rat serum | 9 | 1.823 | 0.607 | 9 | 1.315 | 0.285 | Mann–Whitney test | 0.0244 | 0.7 |
| COX-2 (pg/mL) in rat serum | 7 | 315.9 | 144.3 | 6 | 130.6 | 73.11 | Mann–Whitney test | 0.0082 | 0.4 |
| GAPDH (ng/mL) in rat serum | 8 | 0.506 | 0.302 | 8 | 0.240 | 0.166 | Unpaired t test | 0.0465 | 0.5 |
| NO detection (DAF) in aortic rings | 5 | 6212 | 1012 | 5 | 12,600 | 1209 | Unpaired t test | 0.0254 | 2.08 |
| O2•− detection (DHE) in aortic rings | 6 | 4026 | 358.8 | 5 | 6064 | 1789 | Unpaired t test | 0.0238 | 1.5 |
| H2O2 detection (DCF) in aortic rings | 5 | 6023 | 897.0 | 5 | 14,104 | 1187 | Unpaired t test | 0.0187 | 2.33 |
| 100% Cu—Group A | 200% Cu—Group B | |||||||
|---|---|---|---|---|---|---|---|---|
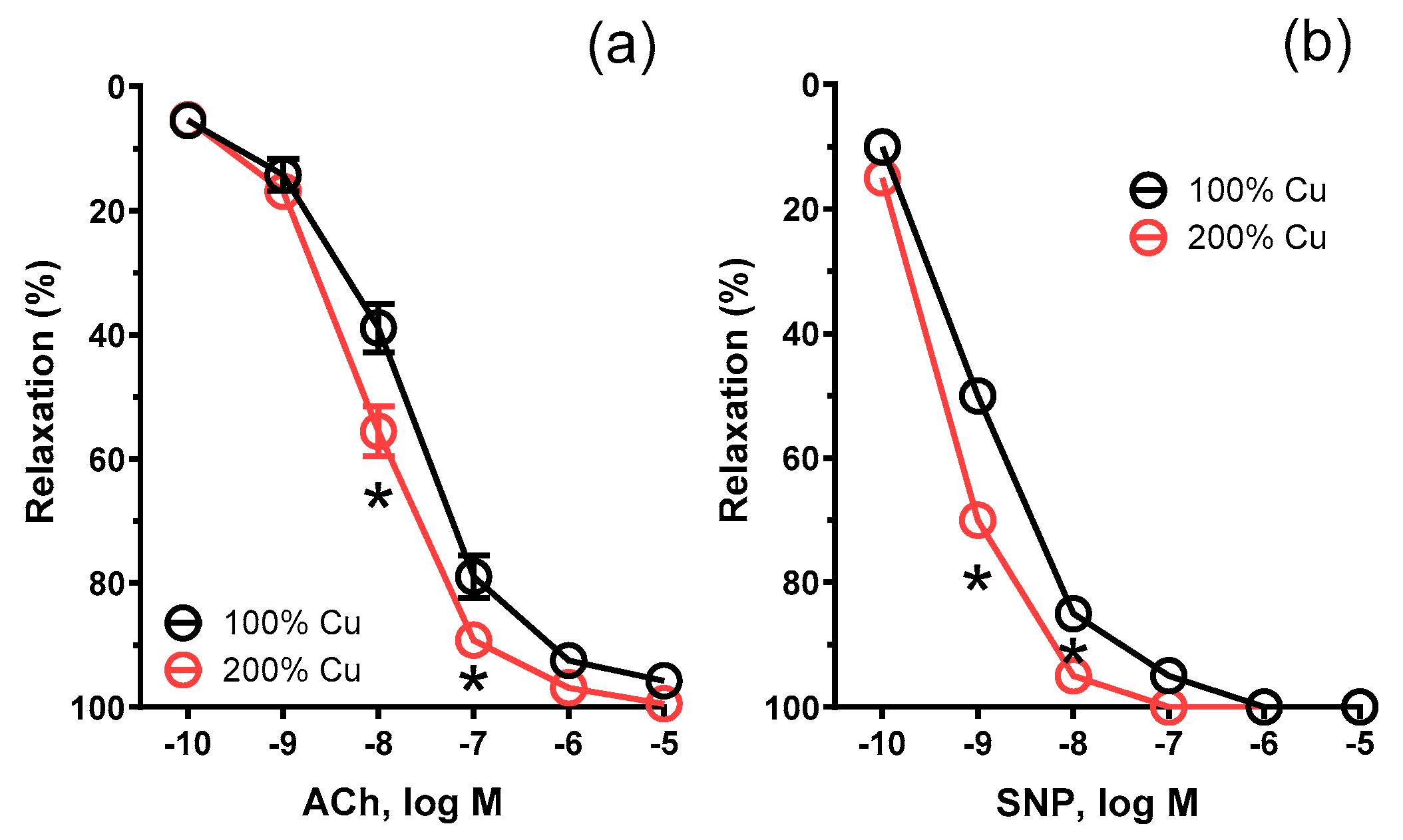
| n | AUC | Emax (%) | pEC50 | n | AUC | Emax (%) | pEC50 | |
| Control Conditions | 8 | 275.2 ± 33.58 | 94.33 ± 1.722 | 7.740 ± 0.062 | 9 | 310.1 # ± 23.95 | 97.94 ± 1.213 | 8.074 # ± 0.043 |
| NS-398 | 6 | 295.6 ± 29.72 | 97.09 ± 2.412 | 7.906 ± 0.087 | 7 | 306.8 ± 40.23 | 98.44 ± 2.213 | 7.917 ± 0.080 |
| AH-6809 | 10 | 244.7 * ± 37.1 | 90.98 * ± 2.835 | 7.497 * ± 0.104 | 12 | 241.2 * ± 44.0 | 93.81 ± 2.311 | 7.597 * ± 0.079 |
| Tranylcypromine | 9 | 251.8 ± 46.85 | 91.79 ± 1.906 | 7.641 ± 0.069 | 10 | 287.4 *# ± 22.91 | 96.76 ± 1.021 | 7.859 ± 0.035 |
| AL-8810 | 6 | 272.6 ± 20.18 | 94.91 ± 1.743 | 7.709 ± 0.063 | 6 | 247.4 # ± 56.66 | 89.81 ± 3.853 | 7.685 ± 0.140 |
| 1400W | 6 | 267.7 ± 42.35 | 96.36 ± 1.693 | 7.772 ± 0.055 | 6 | 253.3 ± 40.27 | 95.83 ± 1.385 | 8.126 # ± 0.053 |
| SNP | 6 | 385 ± 83.93 | 98.88 ± 1.417 | 8.661 ± 0.234 | 6 | 422.5 # ± 67.84 | 99.92 ± 3.626 | 9.397 # ± 0.263 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kitala-Tańska, K.; Socha, K.; Juśkiewicz, J.; Krajewska-Włodarczyk, M.; Majewski, M. The Effect of an Elevated Dietary Copper Level on the Vascular Contractility and Oxidative Stress in Middle-Aged Rats. Nutrients 2024, 16, 1172. https://doi.org/10.3390/nu16081172
Kitala-Tańska K, Socha K, Juśkiewicz J, Krajewska-Włodarczyk M, Majewski M. The Effect of an Elevated Dietary Copper Level on the Vascular Contractility and Oxidative Stress in Middle-Aged Rats. Nutrients. 2024; 16(8):1172. https://doi.org/10.3390/nu16081172
Chicago/Turabian StyleKitala-Tańska, Klaudia, Katarzyna Socha, Jerzy Juśkiewicz, Magdalena Krajewska-Włodarczyk, and Michał Majewski. 2024. "The Effect of an Elevated Dietary Copper Level on the Vascular Contractility and Oxidative Stress in Middle-Aged Rats" Nutrients 16, no. 8: 1172. https://doi.org/10.3390/nu16081172
APA StyleKitala-Tańska, K., Socha, K., Juśkiewicz, J., Krajewska-Włodarczyk, M., & Majewski, M. (2024). The Effect of an Elevated Dietary Copper Level on the Vascular Contractility and Oxidative Stress in Middle-Aged Rats. Nutrients, 16(8), 1172. https://doi.org/10.3390/nu16081172