Echium Oil Reduces Plasma Triglycerides by Increasing Intravascular Lipolysis in apoB100-Only Low Density Lipoprotein (LDL) Receptor Knockout Mice

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Experimental Section
2.1. Animals and Diets
2.2. Plasma Lipid Analysis
2.3. RNA Analysis
2.4. VLDL Characterization
2.5. Hepatic Lipid Analysis
2.6. Determination of Plasma Post-Heparin Lipase Activity
2.7. VLDL Lipolysis by LPL
2.8. TG Synthesis Rate Analysis
2.9. Turnover Studies
2.10. Statistical Analysis
3. Results
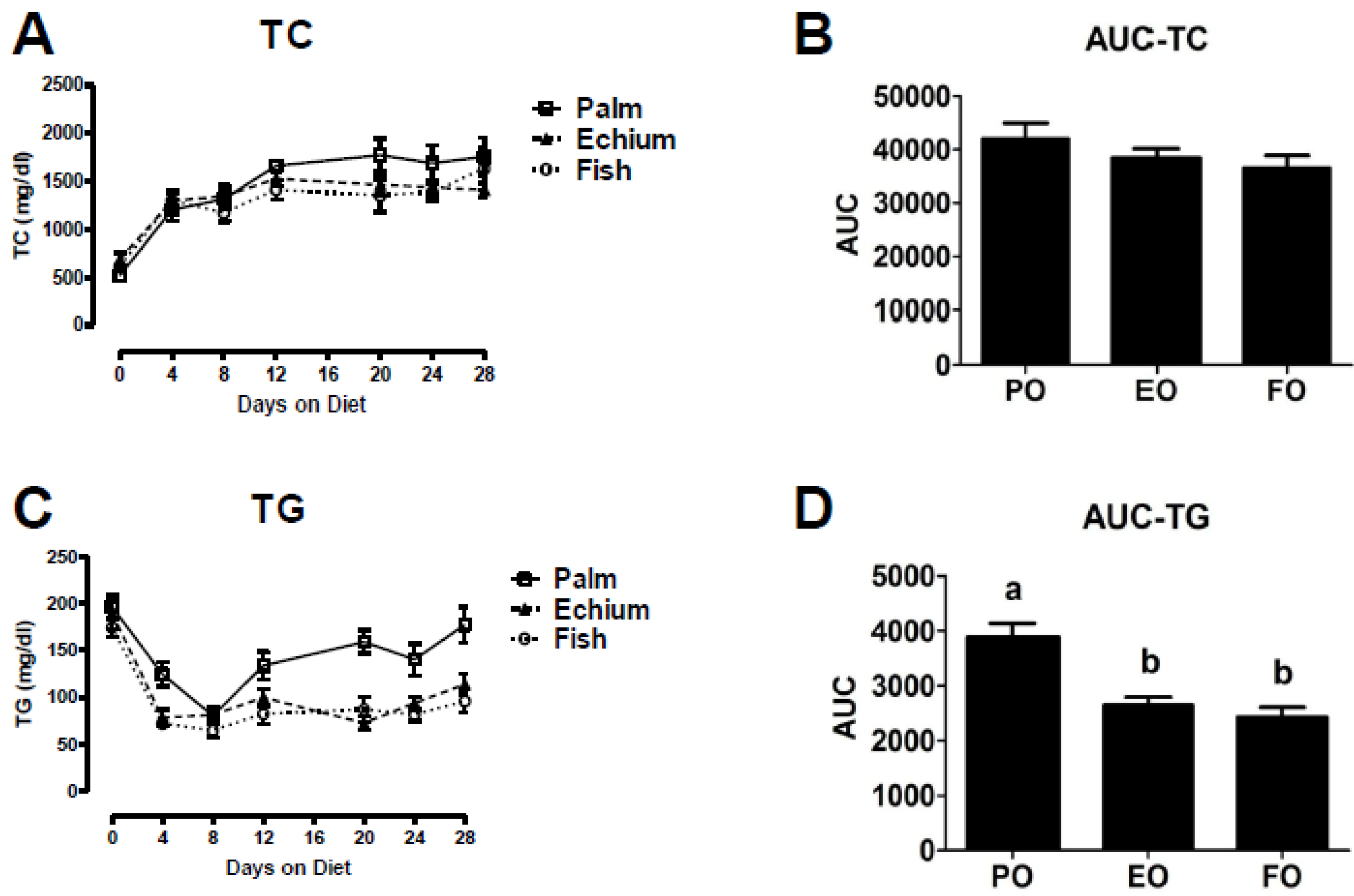
3.1. Echium Oil Reduces Plasma Triglyceride Concentrations

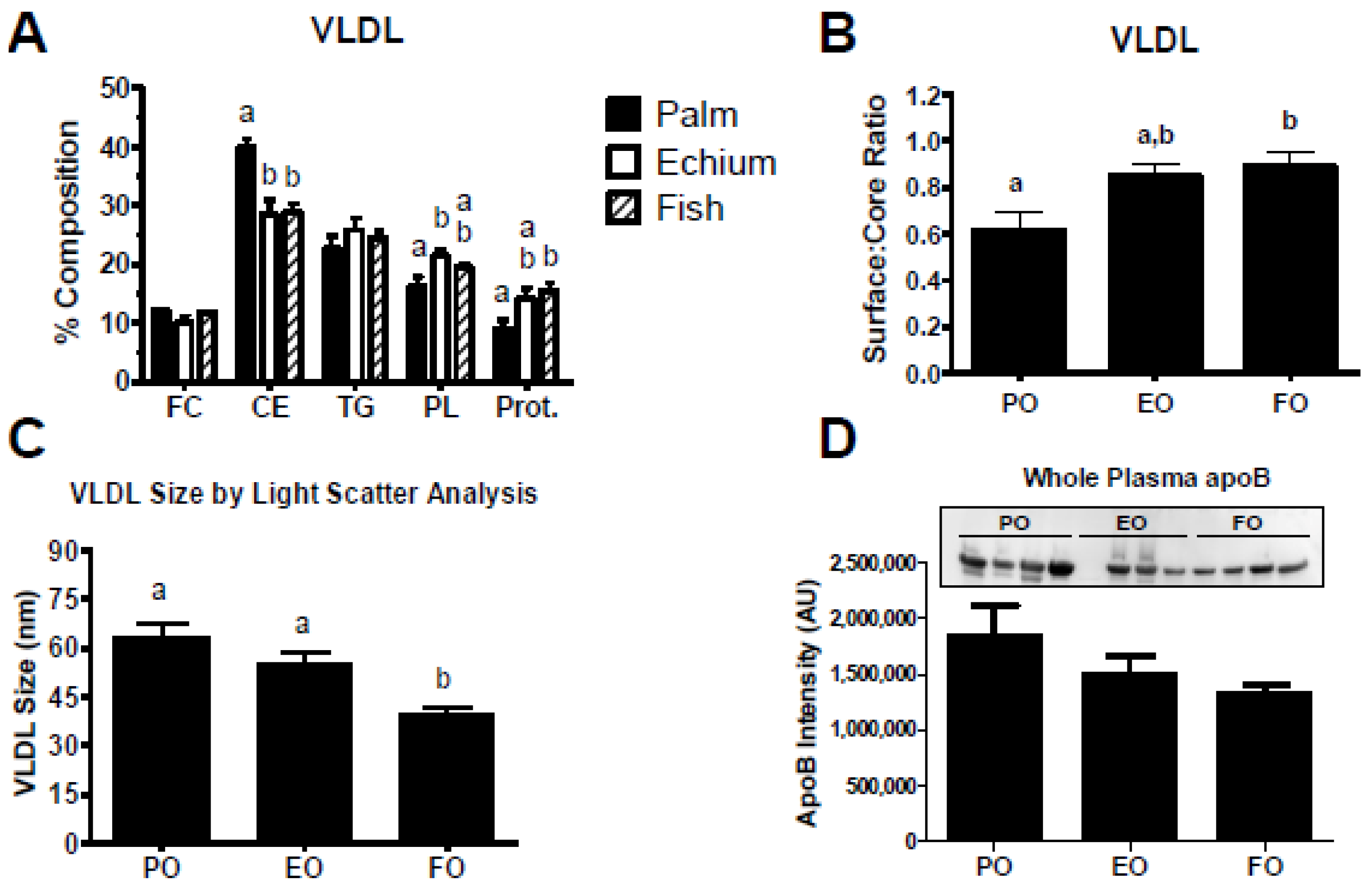
3.2. Echium Oil Alters VLDL Composition

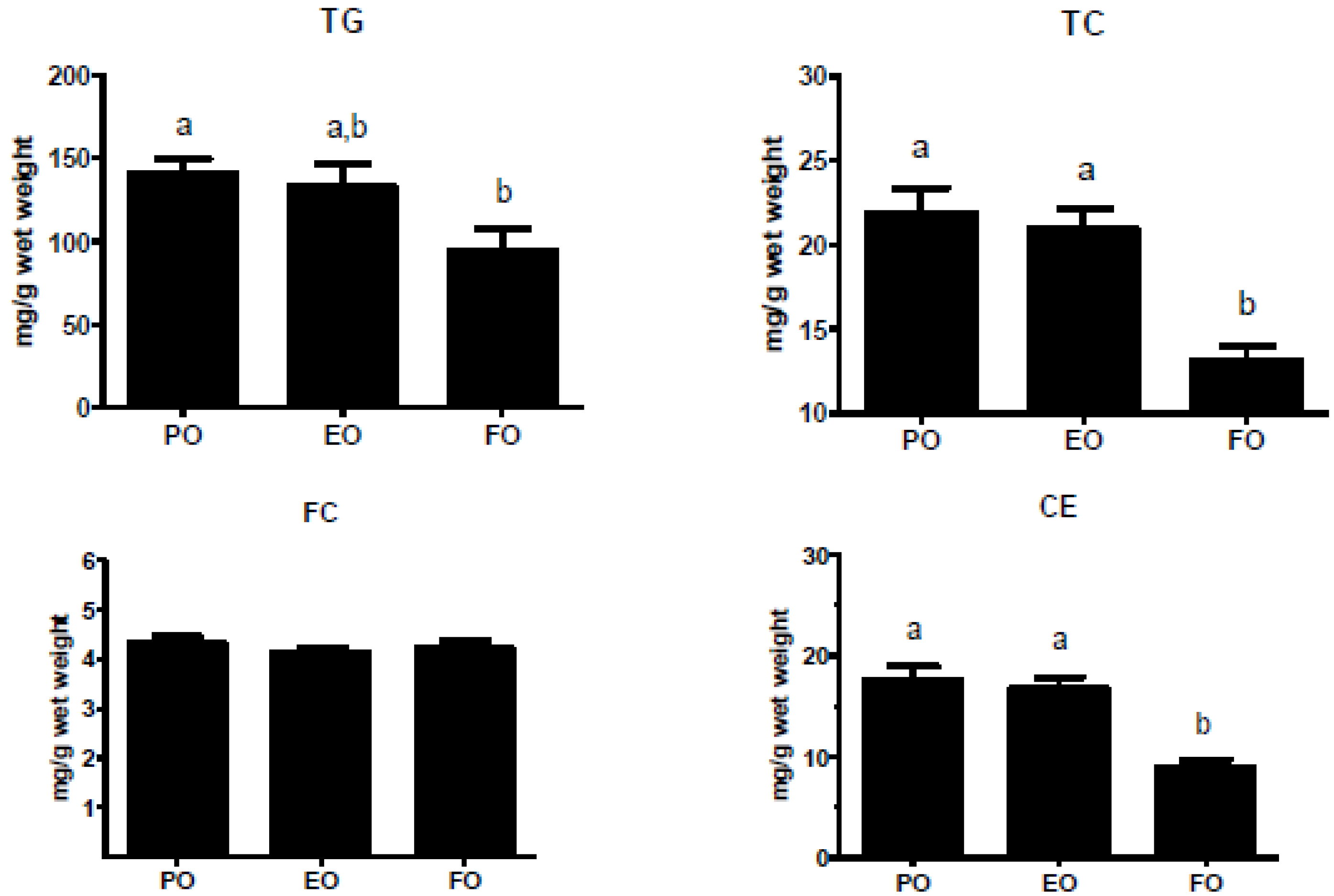
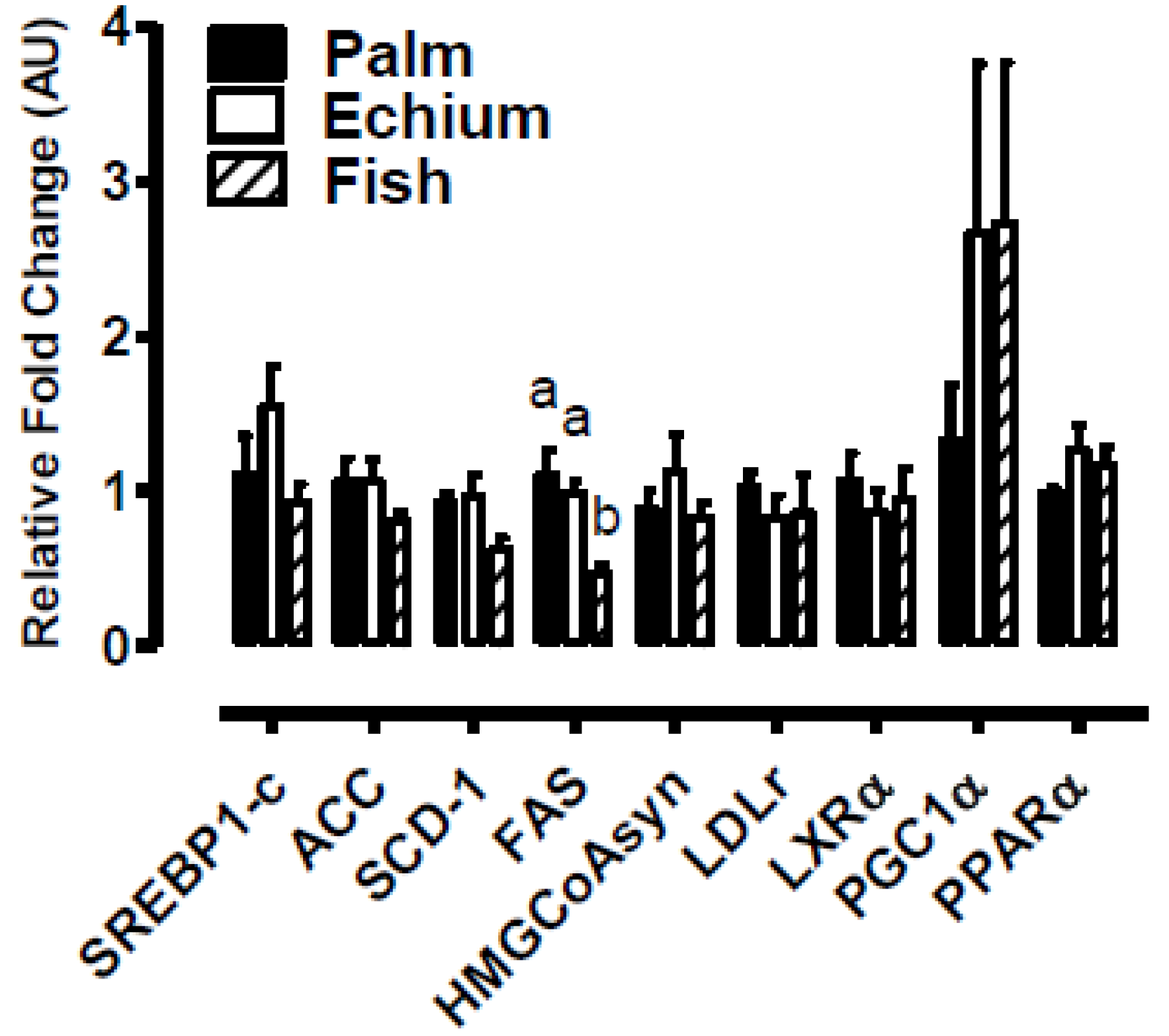
3.3. Echium Oil does not Affect Liver Lipid Content and Gene Expression


3.4. Echium Oil does not Affect liver TG Secretion Rate

3.5. Echium Oil-Derived VLDL Particles Are More Susceptible to Hydrolysis

3.6. Echium Oil Has Minimal Impact on Plasma VLDL Particle Turnover

4. Discussion
5. Conclusions
Acknowledgements
Conflict of Interest
References
- Defilippis, A.P.; Blaha, M.J.; Jacobson, T.A. Omega-3 fatty acids for cardiovascular disease prevention. Curr. Treat. Options Cardiovasc. Med. 2010, 12, 365–380. [Google Scholar] [CrossRef]
- Kromhout, D.; Giltay, E.J.; Geleijnse, J.M. N-3 fatty acids and cardiovascular events after myocardial infarction. N. Engl. J. Med. 2010, 363, 2015–2026. [Google Scholar] [CrossRef]
- Roth, E.M.; Harris, W.S. Fish oil for primary and secondary prevention of coronary heart disease. Curr. Atheroscler. Rep. 2010, 12, 66–72. [Google Scholar] [CrossRef]
- Dyerberg, J.; Bang, H.O. A hypothesis on the development of acute myocardial infarction in Greenlanders. Scand. J. Clin. Lab. Investig. 1982, 161, 7–13. [Google Scholar]
- Dyerberg, J.; Bang, H.O. Lipid metabolism, atherogenesis, and haemostasis in Eskimos: The role of the prostaglandin-3 family. Haemostasis 1979, 8, 227–233. [Google Scholar]
- Bang, H.O.; Dyerberg, J.; Hjoorne, N. The composition of food consumed by Greenland Eskimos. Acta Med. Scand. 1976, 200, 69–73. [Google Scholar]
- Davis, H.R.; Bridenstine, R.T.; Vesselinovitch, D.; Wissler, R.W. Fish oil inhibits development of atherosclerosis in rhesus monkeys. Arteriosclerosis 1987, 7, 441–449. [Google Scholar] [CrossRef]
- Rudel, L.L.; Kelley, K.; Sawyer, J.K.; Shah, R.; Wilson, M.D. Dietary monounsaturated fatty acids promote aortic atherosclerosis in LDL receptor-null, human ApoB100-overexpressing transgenic mice. Arterioscler. Thromb. Vasc. Biol. 1998, 18, 1818–1827. [Google Scholar] [CrossRef]
- Parks, J.S.; Kaduck-Sawyer, J.; Bullock, B.C.; Rudel, L.L. Effect of dietary fish oil on coronary artery and aortic atherosclerosis in African green monkeys. Arteriosclerosis 1990, 10, 1102–1112. [Google Scholar] [CrossRef]
- Albert, C.M.; Campos, H.; Stampfer, M.J.; Ridker, P.M.; Manson, J.E.; Willett, W.C.; Ma, J. Blood levels of long-chain n-3 fatty acids and the risk of sudden death. N. Engl. J. Med. 2002, 346, 1113–1118. [Google Scholar] [CrossRef]
- Burr, M.L.; Fehily, A.M.; Gilbert, J.F.; Rogers, S.; Holliday, R.M.; Sweetnam, P.M.; Elwood, P.C.; Deadman, N.M. Effects of changes in fat, fish, and fibre intakes on death and myocardial reinfarction: Diet and reinfarction trial (DART). Lancet 1989, 2, 757–761. [Google Scholar]
- Sanders, T.A.; Sullivan, D.R.; Reeve, J.; Thompson, G.R. Triglyceride-lowering effect of marine polyunsaturates in patients with hypertriglyceridemia. Arteriosclerosis 1985, 5, 459–465. [Google Scholar] [CrossRef]
- Harris, W.S.; Connor, W.E.; Alam, N.; Illingworth, D.R. Reduction of postprandial triglyceridemia in humans by dietary n-3 fatty acids. J. Lipid Res. 1988, 29, 1451–1460. [Google Scholar]
- Norling, L.V.; Serhan, C.N. Profiling in resolving inflammatory exudates identifies novel anti-inflammatory and pro-resolving mediators and signals for termination. J. Intern. Med. 2010, 268, 15–24. [Google Scholar]
- Harris, W.S. Fish oils and plasma lipid and lipoprotein metabolism in humans: A critical review. J. Lipid Res. 1989, 30, 785–807. [Google Scholar]
- Harris, W.S. N-3 fatty acids and lipoproteins: Comparison of results from human and animal studies. Lipids 1996, 31, 243–252. [Google Scholar] [CrossRef]
- Kris-Etherton, P.M.; Yu, S. Individual fatty acid effects on plasma lipids and lipoproteins: Human studies. Am. J. Clin. Nutr. 1997, 65, 1628S–1644S. [Google Scholar]
- Cullen, P. Evidence that triglycerides are an independent coronary heart disease risk factor. Am. J. Cardiol. 2000, 86, 943–949. [Google Scholar] [CrossRef]
- Oosterveer, M.H.; van Dijk, T.H.; Tietge, U.J.; Boer, T.; Havinga, R.; Stellaard, F.; Groen, A.K.; Kuipers, F.; Reijngoud, D.J. High fat feeding induces hepatic fatty acid elongation in mice. PLoSOne 2009, 4, e6066. [Google Scholar]
- Larter, C.Z.; Yeh, M.M.; Cheng, J.; Williams, J.; Brown, S.; dela Pena, A.; Bell-Anderson, K.S.; Farrell, G.C. Activation of peroxisome proliferator-activated receptor alpha by dietary fish oil attenuates steatosis, but does not prevent experimental steatohepatitis because of hepatic lipoperoxide accumulation. J. Gastroenterol. Hepatol. 2008, 23, 267–275. [Google Scholar]
- Jump, D.B.; Clarke, S.D. Regulation of gene expression by dietary fat. Annu. Rev. Nutr. 1999, 19, 63–90. [Google Scholar] [CrossRef]
- Vasandani, C.; Kafrouni, A.I.; Caronna, A.; Bashmakov, Y.; Gotthardt, M.; Horton, J.D.; Spady, D.K. Upregulation of hepatic LDL transport by n-3 fatty acids in LDL receptor knockout mice. J. Lipid Res. 2002, 43, 772–784. [Google Scholar]
- Qi, K.; Fan, C.; Jiang, J.; Zhu, H.; Jiao, H.; Meng, Q.; Deckelbaum, R.J. Omega-3 fatty acid containing diets decrease plasma triglyceride concentrations in mice by reducing endogenous triglyceride synthesis and enhancing the blood clearance of triglyceride-rich particles. Clin. Nutr. 2008, 27, 424–430. [Google Scholar] [CrossRef]
- Wang, H.; Chen, X.; Fisher, E.A. N-3 fatty acids stimulate intracellular degradation of apoprotein B in rat hepatocytes. J. Clin. Investig. 1993, 91, 1380–1389. [Google Scholar] [CrossRef]
- Halvorsen, B.; Rustan, A.C.; Madsen, L.; Reseland, J.; Berge, R.K.; Sletnes, P.; Christiansen, E.N. Effects of long-chain monounsaturated and n-3 fatty acids on fatty acid oxidation and lipid composition in rats. Ann. Nutr. Metab. 2001, 45, 30–37. [Google Scholar] [CrossRef]
- Kris-Etherton, P.M.; Taylor, D.S.; Yu-Poth, S.; Huth, P.; Moriarty, K.; Fishell, V.; Hargrove, R.L.; Zhao, G.; Etherton, T.D. Polyunsaturated fatty acids in the food chain in the United States. Am. J. Clin. Nutr. 2000, 71, 179S–188S. [Google Scholar]
- Burdge, G. α-Linolenic acid metabolism in men and women: Nutritional and biological implications. Curr. Opin. Clin. Nutr. Metab. Care 2004, 7, 137–144. [Google Scholar] [CrossRef]
- Nakamura, M.T.; Nara, T.Y. Structure, function, and dietary regulation of delta6, delta5, and delta9 desaturases. Annu. Rev. Nutr. 2004, 24, 345–376. [Google Scholar] [CrossRef]
- Guil-Guerrero, J.L.; Gomez-Mercado, F.; Garcia-Maroto, F.; Campra-Madrid, P. Occurrence and characterization of oils rich in gamma-linolenic acid Part I: Echium seeds from Macaronesia. Phytochemistry 2000, 53, 451–456. [Google Scholar] [CrossRef]
- Zhang, P.; Boudyguina, E.; Wilson, M.D.; Gebre, A.K.; Parks, J.S. Echium oil reduces plasma lipids and hepatic lipogenic gene expression in apoB100-only LDL receptor knockout mice. J. Nutr. Biochem. 2008, 19, 655–663. [Google Scholar] [CrossRef]
- Powell-Braxton, L.; Veniant, M.; Latvala, R.D.; Hirano, K.I.; Won, W.B.; Ross, J.; Dybdal, N.; Zlot, C.H.; Young, S.G.; Davidson, N.O. A mouse model of human familial hypercholesterolemia: Markedly elevated low density lipoprotein cholesterol levels and severe atherosclerosis on a low-fat chow diet. Nat. Med. 1998, 4, 934–938. [Google Scholar] [CrossRef]
- Farese, R.V., Jr.; Veniant, M.M.; Cham, C.M.; Flynn, L.M.; Pierotti, V.; Loring, J.F.; Traber, M.; Ruland, S.; Stokowski, R.S.; Huszar, D.; et al. Phenotypic analysis of mice expressing exclusively apolipoprotein B48 or apolipoprotein B100. Proc. Natl. Acad. Sci. USA 1996, 93, 6393–6398. [Google Scholar] [CrossRef]
- Bell, T.A., III; Kelley, K.; Wilson, M.D.; Sawyer, J.K.; Rudel, L.L. Dietary fat-induced alterations in atherosclerosis are abolished by ACAT2-deficiency in apoB100 only, LDLr−/− mice. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 1396–1402. [Google Scholar] [CrossRef]
- Furbee, J.W., Jr.; Francone, O.; Parks, J.S. Alteration of plasma HDL cholesteryl ester composition with transgenic expression of a point mutation (E149A) of human LCAT. J. Lipid Res. 2001, 42, 1626–1635. [Google Scholar]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Rouser, G.; Fkeischer, S.; Yamamoto, A. Two dimensional then layer chromatographic separation of polar lipids and determination of phospholipids by phosphorus analysis of spots. Lipids 1970, 5, 494–496. [Google Scholar] [CrossRef]
- Lee, J.Y.; Timmins, J.M.; Mulya, A.; Smith, T.L.; Zhu, Y.; Rubin, E.M.; Chisholm, J.W.; Colvin, P.L.; Parks, J.S. HDLs in apoA-I transgenic Abca1 knockout mice are remodeled normally in plasma but are hypercatabolized by the kidney. J. Lipid Res. 2005, 46, 2233–2245. [Google Scholar] [CrossRef]
- Wilcox, R.W.; Thuren, T.; Sisson, P.; Kucera, G.L.; Waite, M. Hydrolysis of neutral lipid substrates by rat hepatic lipase. Lipids 1991, 26, 283–288. [Google Scholar] [CrossRef]
- Otway, S.; Robinson, D.S. The use of a non-ionic detergent (Triton WR 1339) to determine rates of triglyceride entry into the circulation of the rat under different physiological conditions. J. Physiol. 1967, 190, 321–332. [Google Scholar]
- McFarlane, A.S. Efficient trace-labelling of proteins with iodine. Nature 1958, 182, 53. [Google Scholar] [CrossRef]
- Lee, J.Y.; Lanningham-Foster, L.; Boudyguina, E.Y.; Smith, T.L.; Young, E.R.; Colvin, P.L.; Thomas, M.J.; Parks, J.S. Preβ high density lipoprotein has two metabolic fates in human apolipoprotein A-I transgenic mice. J. Lipid Res. 2004, 45, 716–728. [Google Scholar]
- Davidson, M.H. Mechanisms for the hypotriglyceridemic effect of marine omega-3 fatty acids. Am. J. Cardiol. 2006, 98, 27i–33i. [Google Scholar] [CrossRef]
- Qi, K.; Al-Haideri, M.; Seo, T.; Carpentier, Y.A.; Deckelbaum, R.J. Effects of particle size on blood clearance and tissue uptake of lipid emulsions with different triglyceride compositions. J. Parenter. Enter. Nutr. 2003, 27, 58–64. [Google Scholar] [CrossRef]
- Nozaki, S.; Garg, A.; Vega, G.L.; Grundy, S.M. Postheparin lipolytic activity and plasma lipoprotein response to omega-3 polyunsaturated fatty acids in patients with primary hypertriglyceridemia. Am. J. Clin. Nutr. 1991, 53, 638–642. [Google Scholar]
- Egusa, G.; Brady, D.W.; Grundy, S.M.; Howard, B.V. Isopropanol precipitation method for the determination of apolipoprotein B specific activity and plasma concentrations during metabolic studies of very low density lipoprotein and low density lipoprotein apolipoprotein B. J. Lipid Res. 1983, 24, 1261–1267. [Google Scholar]
- Ishibashi, S.; Brown, M.S.; Goldstein, J.L.; Gerard, R.D.; Hammer, R.E.; Herz, J. Hypercholesterolemia in low density lipoprotein receptor knockout mice and its reversal by adenovirus-mediated gene delivery. J. Clin. Investig. 1993, 92, 883–893. [Google Scholar] [CrossRef]
- Surette, M.E.; Edens, M.; Chilton, F.H.; Tramposch, K.M. Dietary echium oil increases plasma and neutrophil long-chain (n-3) fatty acids and lowers serum triacylglycerols in hypertriglyceridemic humans. J. Nutr. 2004, 134, 1406–1411. [Google Scholar]
- Bordin, P.; Bodamer, O.A.; Venkatesan, S.; Gray, R.M.; Bannister, P.A.; Halliday, D. Effects of fish oil supplementation on apolipoprotein B100 production and lipoprotein metabolism in normolipidaemic males. Eur. J. Clin. Nutr. 1998, 52, 104–109. [Google Scholar]
- Harris, W.S.; Connor, W.E.; Illingworth, D.R.; Rothrock, D.W.; Foster, D.M. Effects of fish oil on VLDL triglyceride kinetics in humans. J. Lipid Res. 1990, 31, 1549–1558. [Google Scholar]
- Nestel, P.J.; Connor, W.E.; Reardon, M.F.; Connor, S.; Wong, S.; Boston, R. Suppression by diets rich in fish oil of very low density lipoprotein production in man. J. Clin. Investig. 1984, 74, 82–89. [Google Scholar] [CrossRef]
- Parks, J.S.; Johnson, F.L.; Wilson, M.D.; Rudel, L.L. Effect of fish oil diet on hepatic lipid metabolism in nonhuman primates: Lowering of secretion of hepatic triglyceride but not apoB. J. Lipid Res. 1990, 31, 455–466. [Google Scholar]
- Nestel, P.J. Fish oil and cardiovascular disease: Lipids and arterial function. Am. J. Clin. Nutr. 2000, 71, 228S–231S. [Google Scholar]
- Jump, D.B. Fatty acid regulation of gene transcription. Crit. Rev. Clin. Lab. Sci. 2004, 41, 41–78. [Google Scholar]
- Xu, J.; Nakamura, M.T.; Cho, H.P.; Clarke, S.D. Sterol regulatory element binding protein-1 expression is suppressed by dietary polyunsaturated fatty acids. A mechanism for the coordinate suppression of lipogenic genes by polyunsaturated fats. J. Biol. Chem. 1999, 274, 23577–23583. [Google Scholar]
- Kim, H.J.; Takahashi, M.; Ezaki, O. Fish oil feeding decreases mature sterol regulatory element-binding protein 1 (SREBP-1) by down-regulation of SREBP-1c mRNA in mouse liver. A possible mechanism for down-regulation of lipogenic enzyme mRNAs. J. Biol. Chem. 1999, 274, 25892–25898. [Google Scholar]
- Yahagi, N.; Shimano, H.; Hasty, A.H.; Amemiya-Kudo, M.; Okazaki, H.; Tamura, Y.; Iizuka, Y.; Shionoiri, F.; Ohashi, K.; Osuga, J.; et al. A crucial role of sterol regulatory element-binding protein-1 in the regulation of lipogenic gene expression by polyunsaturated fatty acids. J. Biol. Chem. 1999, 274, 35840–35844. [Google Scholar]
- Pan, M.; Cederbaum, A.I.; Zhang, Y.L.; Ginsberg, H.N.; Williams, K.J.; Fisher, E.A. Lipid peroxidation and oxidant stress regulate hepatic apolipoprotein B degradation and VLDL production. J. Clin. Investig. 2004, 113, 1277–1287. [Google Scholar]
- Wong, S.H.; Nestel, P.J.; Trimble, R.P.; Stoker, G.B.; Illman, R.J.; Topping, D.L. The adaptive effects of dietary fish and safflower oil on lipid and lipoprotein metabolism in perfused rat liver. Biochim. Biophys. Acta 1984, 792, 103–109. [Google Scholar] [CrossRef]
- Parks, J.S.; Wilson, M.D.; Johnson, F.L.; Rudel, L.L. Fish oil decreases hepatic cholesteryl ester secretion but not apoB secretion in African green monkeys. J. Lipid Res. 1989, 30, 1535–1544. [Google Scholar]
- Lang, C.A.; Davis, R.A. Fish oil fatty acids impair VLDL assembly and/or secretion by cultured rat hepatocytes. J. Lipid Res. 1990, 31, 2079–2086. [Google Scholar]
- Hulsmann, W.C.; Oerlemans, M.C.; Jansen, H. Activity of heparin-releasable liver lipase. Dependence on the degree of saturation of the fatty acids in the acylglycerol substrates. Biochim. Biophys. Acta 1980, 618, 364–369. [Google Scholar] [CrossRef]
- Weintraub, M.S.; Zechner, R.; Brown, A.; Eisenberg, S.; Breslow, J.L. Dietary polyunsaturated fats of the W-6 and W-3 series reduce postprandial lipoprotein levels. Chronic and acute effects of fat saturation on postprandial lipoprotein metabolism. J. Clin. Investig. 1988, 82, 1884–1893. [Google Scholar]
- Forrest, L.M.; Boudyguina, E.; Wilson, M.D.; Parks, J.S. Echium oil reduces atherosclerosis in apoB100-only LDLrKO mice. Atherosclerosis 2012, 220, 118–121. [Google Scholar] [CrossRef]
- Brown, A.L.; Zhu, X.; Rong, S.; Shewale, S.; Seo, J.; Boudyguina, E.; Gebre, A.K.; Alexander-Miller, M.A.; Parks, J.S. Omega-3 fatty acids ameliorate atherosclerosis by favorably altering monocyte subsets and limiting monocyte recruitment to aortic lesions. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 2122–2130. [Google Scholar] [CrossRef]
Supplementary Files
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Forrest, L.M.; Lough, C.M.; Chung, S.; Boudyguina, E.Y.; Gebre, A.K.; Smith, T.L.; Colvin, P.L.; Parks, J.S. Echium Oil Reduces Plasma Triglycerides by Increasing Intravascular Lipolysis in apoB100-Only Low Density Lipoprotein (LDL) Receptor Knockout Mice. Nutrients 2013, 5, 2629-2645. https://doi.org/10.3390/nu5072629
Forrest LM, Lough CM, Chung S, Boudyguina EY, Gebre AK, Smith TL, Colvin PL, Parks JS. Echium Oil Reduces Plasma Triglycerides by Increasing Intravascular Lipolysis in apoB100-Only Low Density Lipoprotein (LDL) Receptor Knockout Mice. Nutrients. 2013; 5(7):2629-2645. https://doi.org/10.3390/nu5072629
Chicago/Turabian StyleForrest, Lolita M., Christopher M. Lough, Soonkyu Chung, Elena Y. Boudyguina, Abraham K. Gebre, Thomas L. Smith, Perry L. Colvin, and John S. Parks. 2013. "Echium Oil Reduces Plasma Triglycerides by Increasing Intravascular Lipolysis in apoB100-Only Low Density Lipoprotein (LDL) Receptor Knockout Mice" Nutrients 5, no. 7: 2629-2645. https://doi.org/10.3390/nu5072629
APA StyleForrest, L. M., Lough, C. M., Chung, S., Boudyguina, E. Y., Gebre, A. K., Smith, T. L., Colvin, P. L., & Parks, J. S. (2013). Echium Oil Reduces Plasma Triglycerides by Increasing Intravascular Lipolysis in apoB100-Only Low Density Lipoprotein (LDL) Receptor Knockout Mice. Nutrients, 5(7), 2629-2645. https://doi.org/10.3390/nu5072629