Bioavailability of Bergamot (Citrus bergamia) Flavanones and Biological Activity of Their Circulating Metabolites in Human Pro-Angiogenic Cells


 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Ethics Statement
2.3. Bioavailability Study
2.4. Extraction and UHPLC-MS Analysis of Phenolic Compounds from Bergamot Juice
2.5. Extraction and UHPLC-MS Analysis of Phenolic Compounds in Urine and Plasma Samples
2.6. Cell Culture and Conditions
2.7. Pro-Inflammatory Marker Gene Expression
2.8. Apoptosis Assessment
2.9. Statistical Analysis
3. Results
3.1. Flavanone Composition of the Bergamot Juice
3.2. Flavanone Metabolites in Plasma and Urine Samples
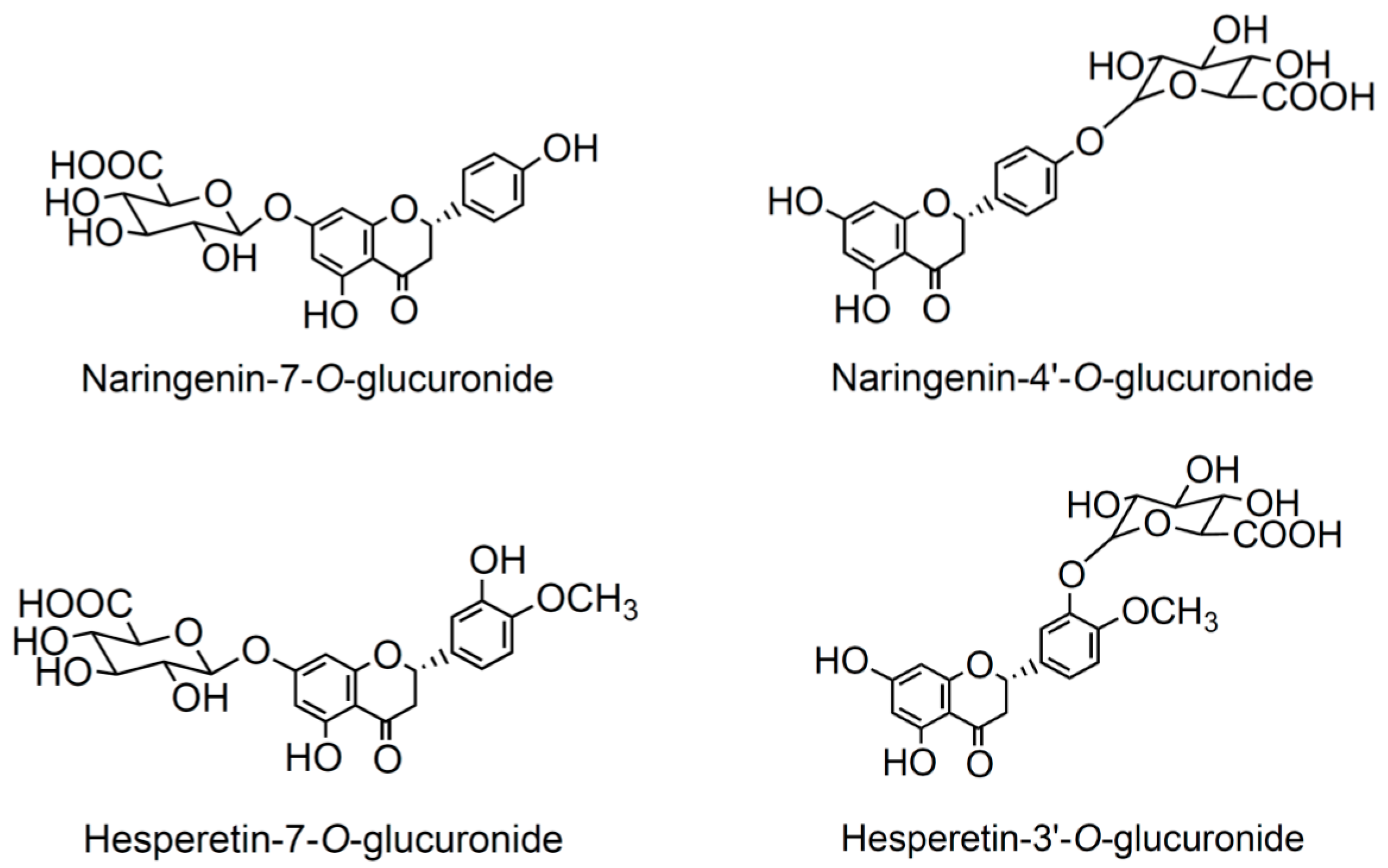
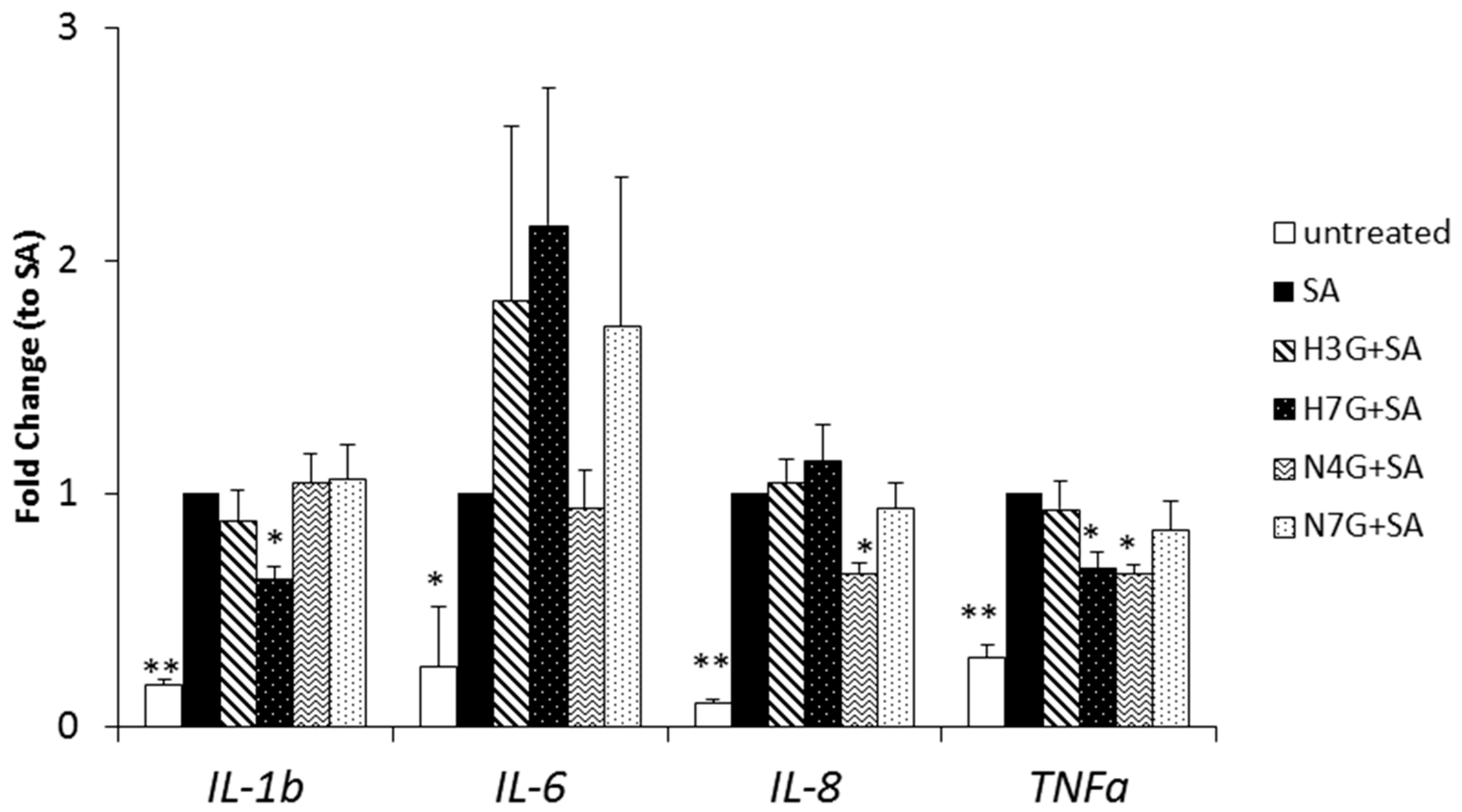
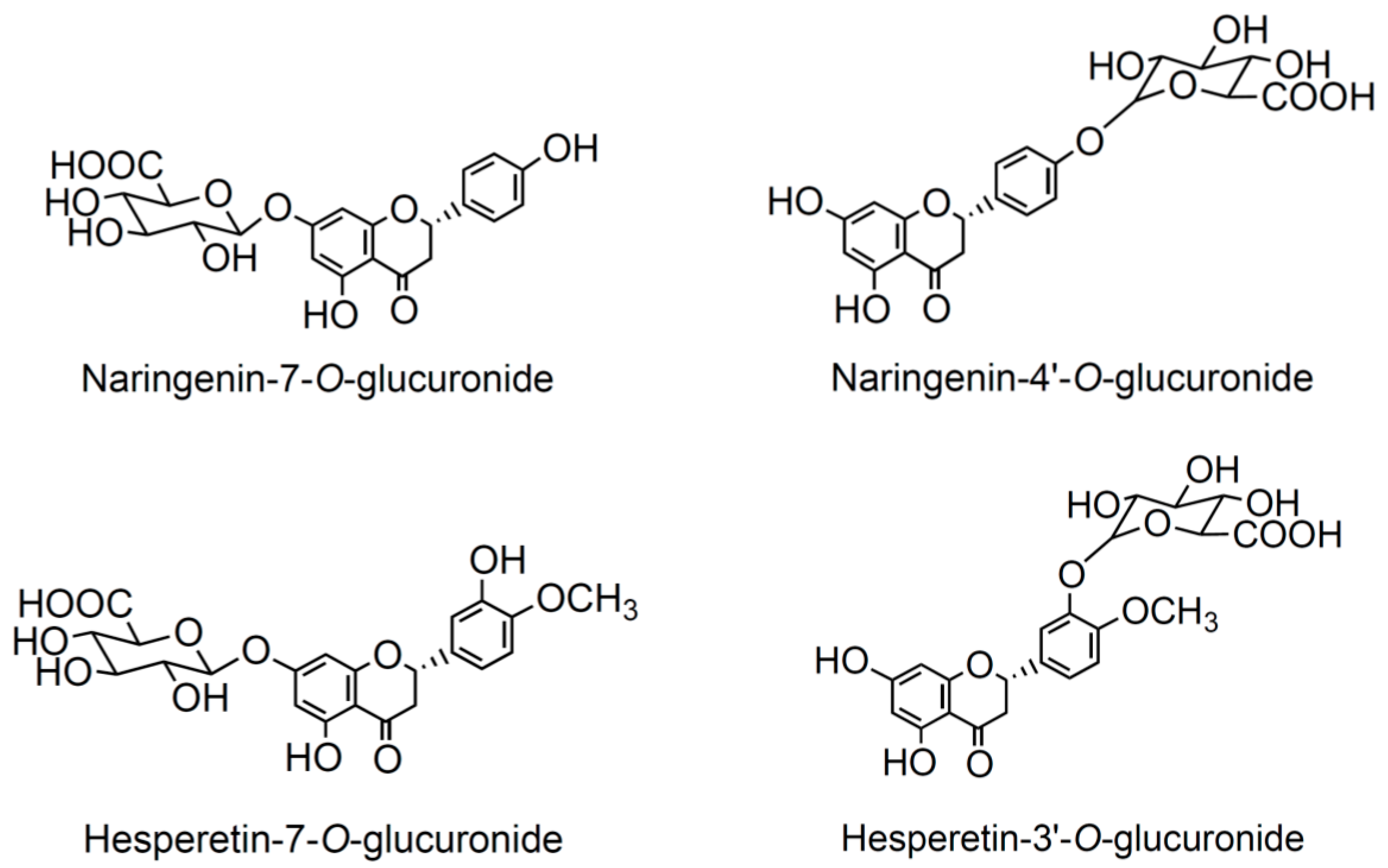
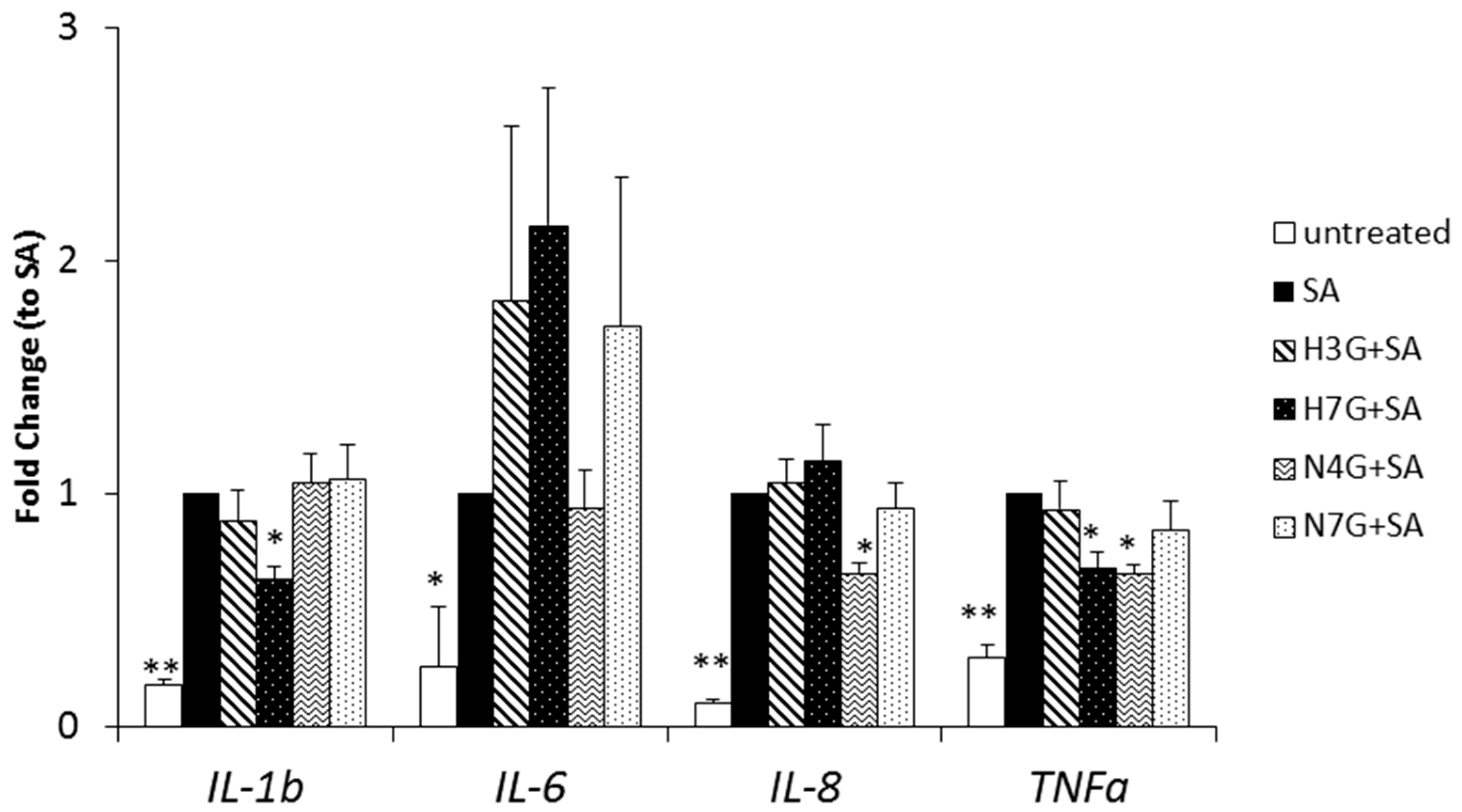
3.3. Effects of Selected Flavanone Metabolites on the Gene Expression of Inflammation Markers
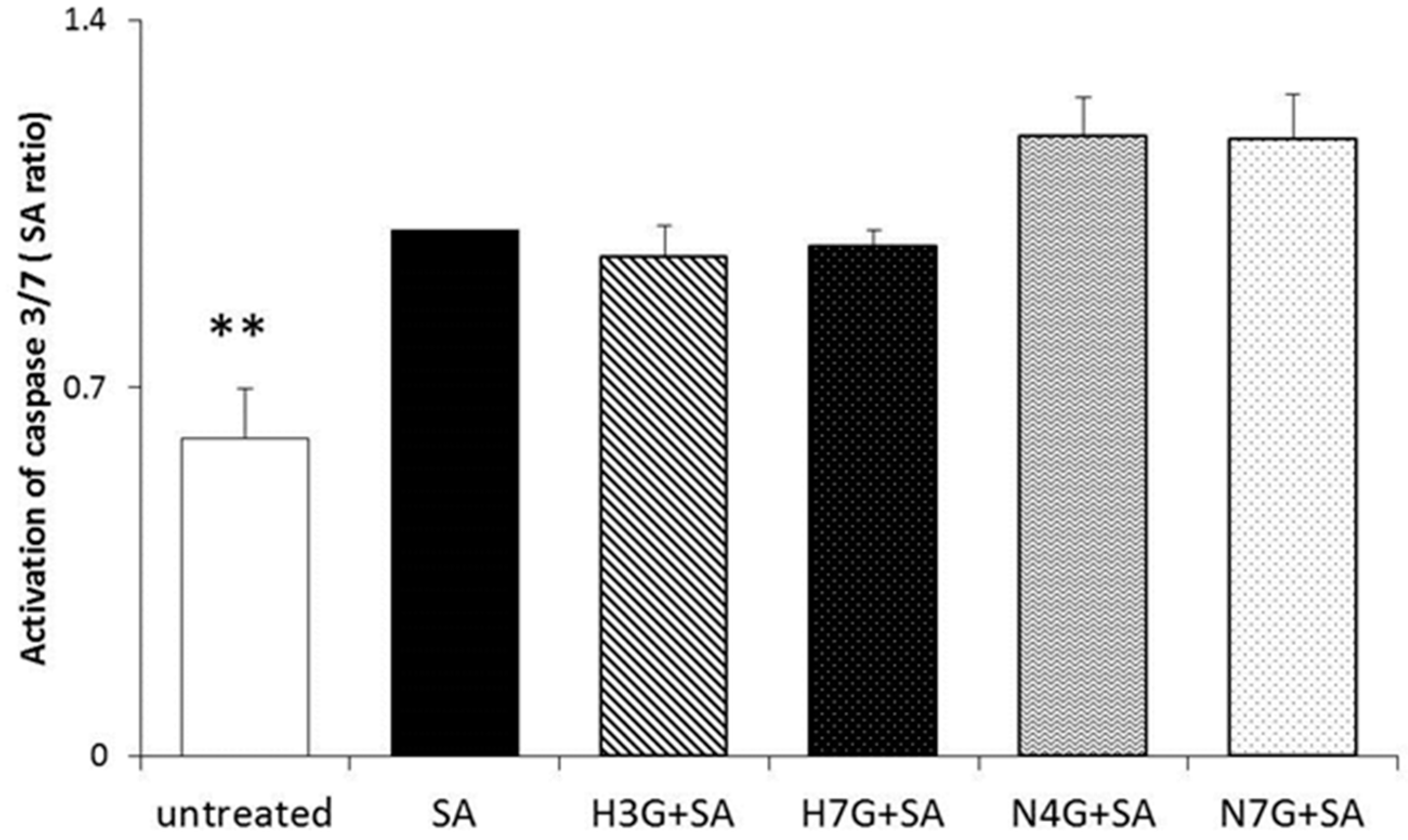
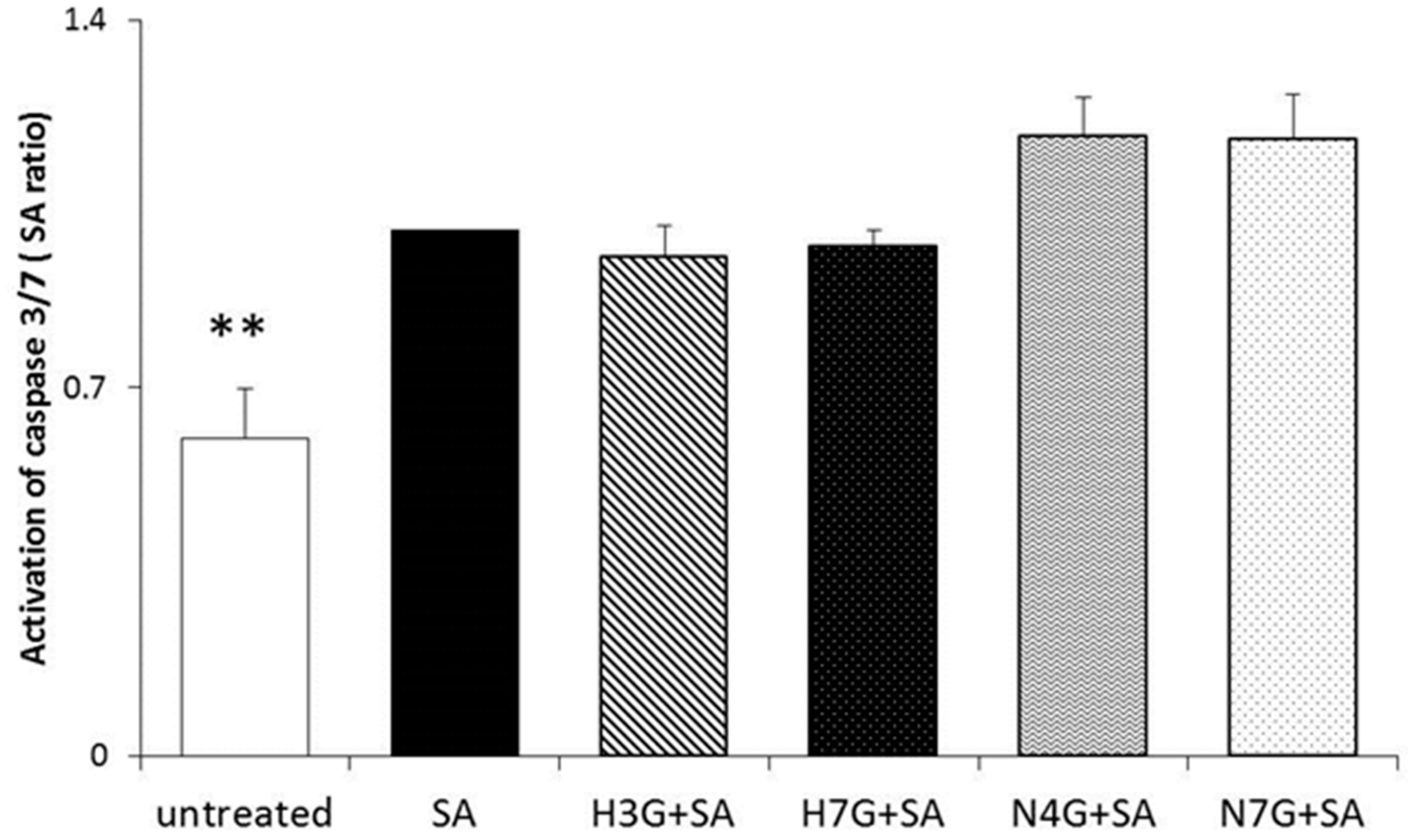
3.4. Effects of Flavanone Metabolites on Cell Apoptosis
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Sandoo, A.; Veldhuijzen van Zanten, J.J.C.S.; Metsios, G.S.; Carroll, D.; Kitas, G.D. The Endothelium and Its Role in Regulating Vascular Tone. Open Cardiovasc. Med. J. 2010, 4, 302–312. [Google Scholar] [CrossRef] [PubMed]
- Fadini, G.P.; Losordo, D.; Dimmeler, S. Critical reevaluation of endothelial progenitor cell phenotypes for therapeutic and diagnostic use. Circ. Res. 2012, 110, 624–637. [Google Scholar] [CrossRef] [PubMed]
- Werner, N.; Kosiol, S.; Schiegl, T.; Ahlers, P.; Walenta, K.; Link, A.; Böhm, M.; Nickenig, G. Circulating Endothelial Progenitor Cells and Cardiovascular Outcomes. N. Engl. J. Med. 2005, 353, 999–1007. [Google Scholar] [CrossRef] [PubMed]
- Zampetaki, A.; Kirton, J.P.; Xu, Q. Vascular repair by endothelial progenitor cells. Cardiovasc. Res. 2008, 78, 413–421. [Google Scholar] [CrossRef] [PubMed]
- DeFronzo, R.A. Insulin resistance, lipotoxicity, type 2 diabetes and atherosclerosis: The missing links. The Claude Bernard Lecture 2009. Diabetologia 2010, 53, 1270–1287. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.A.; Montagnani, M.; Koh, K.K.; Quon, M.J. Reciprocal relationships between insulin resistance and endothelial dysfunction: Molecular and pathophysiological mechanisms. Circulation 2006, 113, 1888–1904. [Google Scholar] [CrossRef] [PubMed]
- Krogmann, A.; Staiger, K.; Haas, C.; Gommer, N.; Peter, A.; Heni, M.; Machicao, F.; Haring, H.U.; Staiger, H. Inflammatory response of human coronary artery endothelial cells to saturated long-chain fatty acids. Microvasc. Res. 2011, 81, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Stentz, F.B.; Kitabchi, A.E. Palmitic acid-induced activation of human T-lymphocytes and aortic endothelial cells with production of insulin receptors, reactive oxygen species, cytokines, and lipid peroxidation. Biochem. Biophys. Res. Commun. 2006, 346, 721–726. [Google Scholar] [CrossRef] [PubMed]
- Staiger, K.; Staiger, H.; Weigert, C.; Haas, C.; Haring, H.U.; Kellerer, M. Saturated, but not unsaturated, fatty acids induce apoptosis of human coronary artery endothelial cells via nuclear factor-kappaB activation. Diabetes 2006, 55, 3121–3126. [Google Scholar] [CrossRef] [PubMed]
- Artwohl, M.; Lindenmair, A.; Sexl, V.; Maier, C.; Rainer, G.; Freudenthaler, A.; Huttary, N.; Wolzt, M.; Nowotny, P.; Luger, A.; et al. Different mechanisms of saturated versus polyunsaturated FFA-induced apoptosis in human endothelial cells. J. Lipid Res. 2008, 49, 2627–2640. [Google Scholar] [CrossRef] [PubMed]
- Spigoni, V.; Fantuzzi, F.; Fontana, A.; Cito, M.; Derlindati, E.; Zavaroni, I.; Cnop, M.; Bonadonna, R.; Dei Cas, A. Stearic Acid at Physiologic Concentrations Induces In Vitro Lipotoxicity In Circulating Angiogenic Cells. Atherosclerosis 2017, 265, 162–171. [Google Scholar] [CrossRef] [PubMed]
- Bose, M.; Lambert, J.D.; Ju, J.; Reuhl, K.R.; Shapses, S.A.; Yang, C.S. The Major Green Tea Polyphenol, (−)-Epigallocatechin-3-Gallate, Inhibits Obesity, Metabolic Syndrome, and Fatty Liver Disease in High-Fat–Fed Mice. J. Nutr. 2008, 138, 1677–1683. [Google Scholar] [PubMed]
- Wang, S.; Moustaid-Moussa, N.; Chen, L.; Mo, H.; Shastri, A.; Su, R.; Bapat, P.; Kwun, I.; Shen, C.L. Novel insights of dietary polyphenols and obesity. J. Nutr. Biochem. 2014, 25, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Huxley, R.R.; Neil, H.A. The relation between dietary flavonol intake and coronary heart disease mortality: A meta-analysis of prospective cohort studies. Eur. J. Clin. Nutr. 2003, 57, 904–908. [Google Scholar] [CrossRef] [PubMed]
- Del Rio, D.; Rodriguez-Mateos, A.; Spencer, J.P.E.; Tognolini, M.; Borges, G.; Crozier, A. Dietary (poly)phenolics in human health: Structures, bioavailability, and evidence of protective effects against chronic diseases. Antioxid. Redox Signal. 2013, 18, 1818–1892. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Mateos, A.; Vauzour, D.; Krueger, C.G.; Shanmuganayagam, D.; Reed, J.; Calani, L.; Mena, P.; Del Rio, D.; Crozier, A. Bioavailability, bioactivity and impact on health of dietary flavonoids and related compounds: An update. Arch. Toxicol. 2014, 88, 1803–1853. [Google Scholar] [CrossRef] [PubMed]
- Zanotti, I.; Dall’Asta, M.; Mena, P.; Mele, L.; Bruni, R.; Ray, S.; Del Rio, D. Atheroprotective effects of (poly)phenols: A focus on cell cholesterol metabolism. Food Funct. 2015, 6, 13–31. [Google Scholar] [CrossRef] [PubMed]
- Gattuso, G.; Barreca, D.; Gargiulli, C.; Leuzzi, U.; Caristi, C. Flavonoid composition of citrus juices. Molecules 2007, 12, 1641–1673. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Lafuente, A.; Guillamon, E.; Villares, A.; Rostagno, M.A.; Martinez, J.A. Flavonoids as anti-inflammatory agents: Implications in cancer and cardiovascular disease. Inflamm. Res. 2009, 58, 537–552. [Google Scholar] [CrossRef] [PubMed]
- Testai, L.; Calderone, V. Nutraceutical value of citrus flavanones and their implications in cardiovascular disease. Nutrients 2017, 9, 502. [Google Scholar] [CrossRef] [PubMed]
- Ferlazzo, N.; Cirmi, S.; Calapai, G.; Ventura-Spagnolo, E.; Gangemi, S.; Navarra, M. Anti-Inflammatory Activity of Citrus bergamia Derivatives: Where Do We Stand? Molecules 2016, 21, 1273. [Google Scholar] [CrossRef] [PubMed]
- Currò, M.; Risitano, R.; Ferlazzo, N.; Cirmi, S.; Gangemi, C.; Caccamo, D.; Ientile, R.; Navarra, M. Citrus bergamia Juice Extract Attenuates beta-Amyloid-Induced Pro-Inflammatory Activation of THP-1 Cells Through MAPK and AP-1 Pathways. Sci. Rep. 2016, 6, 20809. [Google Scholar] [CrossRef] [PubMed]
- Risitano, R.; Currò, M.; Cirmi, S.; Ferlazzo, N.; Campiglia, P.; Caccamo, D.; Ientile, R.; Navarra, M. Flavonoid fraction of Bergamot juice reduces LPS-induced inflammatory response through SIRT1-mediated NF-kappaB inhibition in THP-1 monocytes. PLoS ONE 2014, 9, e107431. [Google Scholar] [CrossRef] [PubMed]
- Navarra, M.; Ursino, M.R.; Ferlazzo, N.; Russo, M.; Schumacher, U.; Valentiner, U. Effect of Citrus bergamia juice on human neuroblastoma cells in vitro and in metastatic xenograft models. Fitoterapia 2014, 95, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Toth, P.P.; Patti, A.M.; Nikolic, D.; Giglio, R.V.; Castellino, G.; Biancucci, T.; Geraci, F.; David, S.; Montalto, G.; Rizvi, A.; et al. Bergamot Reduces Plasma Lipids, Atherogenic Small Dense LDL, and Subclinical Atherosclerosis in Subjects with Moderate Hypercholesterolemia: A 6 Months Prospective Study. Front. Pharmacol. 2016, 6, 299. [Google Scholar] [CrossRef] [PubMed]
- Vallejo, F.; Larrosa, M.; Escudero, E.; Zafrilla, M.P.; Cerdá, B.; Boza, J.; García-Conesa, M.T.; Espín, J.C.; Tomás-Barberán, F.A. Concentration and solubility of flavanones in orange beverages affect their bioavailability in humans. J. Agric. Food Chem. 2010, 58, 6516–6524. [Google Scholar] [CrossRef] [PubMed]
- Mena, P.; Sánchez-Salcedo, E.M.; Tassotti, M.; Martínez, J.J.; Hernández, F.; Del Rio, D. Phytochemical evaluation of eight white (Morus alba L.) and black (Morus nigra L.) mulberry clones grown in Spain based on UHPLC-ESI-MSn metabolomic profiles. Food Res. Int. 2016, 89, 1116–1122. [Google Scholar] [CrossRef]
- Ludwig, I.A.; Mena, P.; Calani, L.; Borges, G.; Pereira-Caro, G.; Bresciani, L.; Del Rio, D.; Lean, M.E.J.; Crozier, A. New insights into the bioavailability of red raspberry anthocyanins and ellagitannins. Free Radic. Biol. Med. 2015, 89, 758–769. [Google Scholar] [CrossRef] [PubMed]
- Spigoni, V.; Picconi, A.; Cito, M.; Ridolfi, V.; Bonomini, S.; Casali, C.; Zavaroni, I.; Gnudi, L.; Metra, M.; Dei Cas, A. Pioglitazone Improves In Vitro Viability and Function of Endothelial Progenitor Cells from Individuals with Impaired Glucose Tolerance. PLoS ONE 2012, 7, e48283. [Google Scholar] [CrossRef] [PubMed]
- Spigoni, V.; Cito, M.; Alinovi, R.; Pinelli, S.; Passeri, G.; Zavaroni, I.; Goldoni, M.; Campanini, M.; Aliatis, I.; Mutti, A.; et al. Effects of TiO2 and Co3O4 Nanoparticles on Circulating Angiogenic Cells. PLoS ONE 2015, 10, e0119310. [Google Scholar] [CrossRef] [PubMed]
- Spigoni, V.; Lombardi, C.; Cito, M.; Picconi, A.; Ridolfi, V.; Andreoli, R.; Anelli, N.; Gnudi, L.; Goldoni, M.; Zavaroni, I.; et al. n-3 PUFA increase bioavailability and function of endothelial progenitor cells. Food Funct. 2014, 5, 1881–1890. [Google Scholar] [CrossRef] [PubMed]
- Cousin, S.P.; Hügl, S.; Wrede, C.; Kajio, H.; Myers, M.G.; Rhodes, C.J. Free fatty acid-induced inhibition of glucose and insulin-like growth factor I-induced deoxyribonucleic acid synthesis in the pancreatic beta-cell line INS-1. Endocrinology 2001, 142, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Mullen, W.; Archeveque, M.A.; Edwards, C.A.; Matsumoto, H.; Crozier, A. Bioavailability and metabolism of orange juice flavanones in humans: Impact of a full-fat yogurt. J. Agric. Food Chem. 2008, 56, 11157–11164. [Google Scholar] [CrossRef] [PubMed]
- Sommella, E.; Pepe, G.; Pagano, F.; Tenore, G.C.; Marzocco, S.; Manfra, M.; Calabrese, G.; Aquino, R.P.; Campiglia, P. UHPLC profiling and effects on LPS-stimulated J774A.1 macrophages of flavonoids from bergamot (Citrus bergamia) juice, an underestimated waste product with high anti-inflammatory potential. J. Funct. Foods 2014, 7, 641–649. [Google Scholar] [CrossRef]
- Salerno, R.; Casale, F.; Calandruccio, C.; Procopio, A. Characterization of flavonoids in Citrus bergamia (Bergamot) polyphenolic fraction by liquid chromatography–high resolution mass spectrometry (LC/HRMS). PharmaNutrition 2016, 4, S1–S7. [Google Scholar] [CrossRef]
- Di Donna, L.; De Luca, G.; Mazzotti, F.; Napoli, A.; Salerno, R.; Taverna, D.; Sindona, G. Statin-like principles of bergamot fruit (Citrus bergamia): Isolation of 3-hydroxymethylglutaryl flavonoid glycosides. J. Nat. Prod. 2009, 72, 1352–1354. [Google Scholar] [CrossRef] [PubMed]
- Gattuso, G.; Caristi, C.; Gargiulli, C.; Bellocco, E.; Toscano, G.; Leuzzi, U. Flavonoid glycosides in bergamot juice (Citrus bergamia Risso). J. Agric. Food Chem. 2006, 54, 3929–3935. [Google Scholar] [CrossRef] [PubMed]
- Shi, P.; He, Q.; Song, Y.; Qu, H.; Cheng, Y. Characterization and identification of isomeric flavonoid O-diglycosides from genus Citrus in negative electrospray ionization by ion trap mass spectrometry and time-of-flight mass spectrometry. Anal. Chim. Acta 2007, 598, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Escudero-López, B.; Calani, L.; Fernández-Pachón, M.S.; Ortega, A.; Brighenti, F.; Crozier, A.; Del Rio, D. Absorption, metabolism, and excretion of fermented orange juice (poly)phenols in rats. BioFactors 2014, 40, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Schär, M.Y.; Curtis, P.J.; Hazim, S.; Ostertag, L.M.; Kay, C.D.; Potter, J.F.; Cassidy, A. Orange juice–derived flavanone and phenolic metabolites do not acutely affect cardiovascular risk biomarkers: A randomized, placebo-controlled, crossover trial in men at moderate risk of cardiovascular disease. Am. J. Clin. Nutr. 2015, 101, 931–938. [Google Scholar] [CrossRef] [PubMed]
- Pereira-Caro, G.; Ludwig, I.A.; Polyviou, T.; Malkova, D.; García, A.; Moreno-Rojas, J.M.; Crozier, A. Identification of Plasma and Urinary Metabolites and Catabolites Derived from Orange Juice (Poly)phenols: Analysis by High-Performance Liquid Chromatography-High-Resolution Mass Spectrometry. J. Agric. Food Chem. 2016, 64, 5724–5735. [Google Scholar] [CrossRef] [PubMed]
- Di Donna, L.; Iacopetta, D.; Cappello, A.R.; Gallucci, G.; Martello, E.; Fiorillo, M.; Dolce, V.; Sindona, G. Hypocholesterolaemic activity of 3-hydroxy-3-methyl-glutaryl flavanones enriched fraction from bergamot fruit (Citrus bergamia): “In vivo” studies. J. Funct. Foods 2014, 7, 558–568. [Google Scholar] [CrossRef]
- Sommella, E.; Pepe, G.; Pagano, F.; Tenore, G.C.; Dugo, P.; Manfra, M.; Campiglia, P. Ultra high performance liquid chromatography with ion-trap TOF-MS for the fast characterization of flavonoids in Citrus bergamia juice. J. Sep. Sci. 2013, 36, 3351–3355. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.K.; Zill, E.H.; Dangles, O. A comprehensive review on flavanones, the major citrus polyphenols. J. Food Compost. Anal. 2014, 33, 85–104. [Google Scholar] [CrossRef]
- Kay, C.D.; Pereira-Caro, G.; Ludwig, I.A.; Clifford, M.N.; Crozier, A. Anthocyanins and Flavanones Are More Bioavailable than Previously Perceived: A Review of Recent Evidence. Annu. Rev. Food Sci. Technol. 2017, 8, 155–180. [Google Scholar] [CrossRef] [PubMed]
- Orrego-Lagarón, N.; Martínez-Huélamo, M.; Vallverdú-Queralt, A.; Lamuela-Raventos, R.M.; Escribano-Ferrer, E. High gastrointestinal permeability and local metabolism of naringenin: Influence of antibiotic treatment on absorption and metabolism. Br. J. Nutr. 2015, 114, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Pereira-Caro, G.; Borges, G.; Van Der Hooft, J.; Clifford, M.N.; Del Rio, D.; Lean, M.E.J.; Roberts, S.A.; Kellerhals, M.B.; Crozier, A. Orange juice (poly)phenols are highly bioavailable in humans. Am. J. Clin. Nutr. 2014, 100, 1378–1384. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, I.L.F.; Chee, W.S.S.; Poulsen, L.; Offord-Cavin, E.; Rasmussen, S.E.; Frederiksen, H.; Enslen, M.; Barron, D.; Horcajada, M.N.; Williamson, G. Bioavailability is improved by enzymatic modification of the citrus flavonoid hesperidin in humans: A randomized, double-blind, crossover trial. J. Nutr. 2006, 136, 404–408. [Google Scholar] [PubMed]
- Bredsdorff, L.; Nielsen, I.L.F.; Rasmussen, S.E.; Cornett, C.; Barron, D.; Bouisset, F.; Offord, E.; Williamson, G. Absorption, conjugation and excretion of the flavanones, naringenin and hesperetin from-rhamnosidase-treated orange juice in human subjects. Br. J. Nutr. 2010, 103, 1602–1609. [Google Scholar] [CrossRef] [PubMed]
- Pereira-Caro, G.; Polyviou, T.; Ludwig, I.A.; Nastase, A.-M.; Moreno-Rojas, J.M.; Garcia, A.L.; Malkova, D.; Crozier, A. Bioavailability of orange juice (poly)phenols: The impact of short-term cessation of training by male endurance athletes. Am. J. Clin. Nutr. 2017. [Google Scholar] [CrossRef] [PubMed]
- Rizza, S.; Muniyappa, R.; Iantorno, M.; Kim, J.A.; Chen, H.; Pullikotil, P.; Senese, N.; Tesauro, M.; Lauro, D.; Cardillo, C.; et al. Citrus polyphenol hesperidin stimulates production of nitric oxide in endothelial cells while improving endothelial function and reducing inflammatory markers in patients with metabolic syndrome. J. Clin. Endocrinol. Metab. 2011, 96, E782–E792. [Google Scholar] [CrossRef] [PubMed]
- Rendeiro, C.; Dong, H.; Saunders, C.; Harkness, L.; Blaze, M.; Hou, Y.; Belanger, R.L.; Corona, G.; Lovegrove, J.A.; Spencer, J.P. Flavanone-rich citrus beverages counteract the transient decline in postprandial endothelial function in humans: A randomised, controlled, double-masked, cross-over intervention study. Br. J. Nutr. 2016, 116, 1999–2010. [Google Scholar] [CrossRef] [PubMed]
- Morand, C.; Dubray, C.; Milenkovic, D.; Lioger, D.; Martin, J.F.; Scalbert, A.; Mazur, A. Hesperidin contributes to the vascular protective effects of orange juice: A randomized crossover study in healthy volunteers. Am. J. Clin. Nutr. 2011, 93, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Janda, E.; Lascala, A.; Martino, C.; Ragusa, S.; Nucera, S.; Walker, R.; Gratteri, S.; Mollace, V. Molecular mechanisms of lipid- and glucose-lowering activities of bergamot flavonoids. PharmaNutrition 2016, 4, S8–S18. [Google Scholar] [CrossRef]
- Impellizzeri, D.; Cordaro, M.; Campolo, M.; Gugliandolo, E.; Esposito, E.; Benedetto, F.; Cuzzocrea, S.; Navarra, M. Anti-inflammatory and Antioxidant Effects of Flavonoid-Rich Fraction of Bergamot Juice (BJe) in a Mouse Model of Intestinal Ischemia/Reperfusion Injury. Front. Pharmacol 2016, 7, 203. [Google Scholar] [CrossRef] [PubMed]
- Gimenez-Bastida, J.A.; Gonzalez-Sarrias, A.; Vallejo, F.; Espin, J.C.; Tomas-Barberan, F.A. Hesperetin and its sulfate and glucuronide metabolites inhibit TNF-alpha induced human aortic endothelial cell migration and decrease plasminogen activator inhibitor-1 (PAI-1) levels. Food Funct. 2016, 7, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Chanet, A.; Milenkovic, D.; Claude, S.; Maier, J.A.; Kamran Khan, M.; Rakotomanomana, N.; Shinkaruk, S.; Berard, A.M.; Bennetau-Pelissero, C.; Mazur, A.; et al. Flavanone metabolites decrease monocyte adhesion to TNF-alpha-activated endothelial cells by modulating expression of atherosclerosis-related genes. Br. J. Nutr. 2013, 110, 587–598. [Google Scholar] [CrossRef] [PubMed]
- Dall’Asta, M.; Derlindati, E.; Curella, V.; Mena, P.; Calani, L.; Ray, S.; Zavaroni, I.; Brighenti, F.; Del Rio, D. Effects of naringenin and its phase II metabolites on in vitro human macrophage gene expression. Int. J. Food Sci. Nutr. 2013, 64, 843–849. [Google Scholar] [CrossRef] [PubMed]
- Galleano, M.; Calabro, V.; Prince, P.D.; Litterio, M.C.; Piotrkowski, B.; Vazquez-Prieto, M.A.; Miatello, R.M.; Oteiza, P.I.; Fraga, C.G. Flavonoids and metabolic syndrome. Ann. N. Y. Acad. Sci. 2012, 1259, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, M.; Jokura, H.; Hashizume, K.; Ominami, H.; Shibuya, Y.; Suzuki, A.; Hase, T.; Shimotoyodome, A. Hesperidin metabolite hesperetin-7-O-glucuronide, but not hesperetin-3’-O-glucuronide, exerts hypotensive, vasodilatory, and anti-inflammatory activities. Food Funct. 2013, 4, 1346–1351. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Id. | Compounds | RT (min) | [M − H]− (m/z) | MS2 Ion Fragments (m/z) a | MS3 Ion Fragments (m/z) a | Ident. c | Concentration (μmol/g dw) |
|---|---|---|---|---|---|---|---|
| 1 | Eriodictyol-7-O-rutinoside (eriocitrin) | 9.5 | 595 | 287 b | 151 | [34] | 2.30 ± 0.22 |
| 2 | Eriodictyol-7-O-neohesperidoside (neoeriocitrin) | 9.71 | 595 | 459, 287, 235 | 235, 357, 441, 271, 151 | [34] | 7.83 ± 0.85 |
| 3 | Naringenin-7-O-rutinoside (narirutin) | 10.19 | 579 | 271 | 151, 177 | Std | 1.06 ± 0.05 |
| 4 | Hesperetin-7-O-rutinoside (hesperidin) | 10.2 | 609 | 301 | 286, 242, 283, 257, 125 | Std | 2.10 ± 0.22 |
| 5 | Naringenin-7-O-neohesperidoside (naringin) | 10.4 | 579 | 459, 271, 235 | 357, 235, 271, 441, 339 | [38] | 12.46 ± 1.61 |
| 6 | Eriodictyol-7-O-neohesperidoside-6″-O-HMG | 10.57 | 739 | 595, 637, 677 | 459, 287 | [35] | 7.53 ± 0.76 |
| 7 | Hesperetin-7-O-neohesperidoside (neohesperidin) | 10.79 | 609 | 301, 343, 489, 447 | 286, 242, 283, 257, 125 | [34] | 10.31 ± 0.61 |
| 8 | Naringenin-7-O-neohesperidoside-6″-O-HMG (melitidin) | 11.13 | 723 | 579, 621, 661 | 459, 271, 313 | [34] | 8.25 ± 0.93 |
| 9 | Naringenin-7-O-glucoside-6″-O-HMG | 11.39 | 577 | 271, 433, 475 | 151, 177 | [35] | 0.27 ± 0.03 |
| 10 | Hesperetin-7-O-neohesperidoside-6″-O-HMG (brutieridin) | 11.43 | 753 | 609, 651, 691 | 301, 343, 489 | [34] | 19.70 ± 2.04 |
| 11 | Isosakuranetin-7-O-rutinoside (didymin) | 12.25 | 593 | 285, 327, 473 | 270, 243, 164, 241, 151 | Std | 0.26 ± 0.02 |
| 12 | Isosakuranetin-7-O-neohesperidoside-6″-O-HMG (parmigin) | 12.75 | 737 | 593, 635, 675 | 285 (MS4: 270, 243), 327, 473 | - | 0.99 ± 0.03 |
| 13 | Naringenin | 13.32 | 271 | 151, 177 | Std | traces | |
| 14 | Isosakuranetin | 13.57 | 285 | 270, 243 | Std | traces | |
| 15 | Hesperetin | 13.78 | 301 | 286, 242 | Std | traces | |
| 16 | Eriodyctiol | 13.82 | 287 | 151 | [38] | traces |
| Id. | Compounds | RT (min) | [M − H]− (m/z) | MS2 Ion Fragments (m/z) a | MS3 Ion Fragments (m/z) a | MS4 Ion Fragments (m/z) a | Location c |
|---|---|---|---|---|---|---|---|
| M1 | Hesperitin-O-glucuronide-sulfate | 3.32 | 557 | 381 b, 447 | 301 | 286, 242, 199, 283 | P |
| M2 | Hesperitin-O-glucuronide-sulfate | 3.51 | 557 | 381, 447 | 301, 229 | 286, 242, 199, 283 | P, U |
| M3 | Eriodictyol-O-glucuronide | 4.04 | 463 | 287, 175 | 151 | P, U | |
| M4 | Naringenin-7-O-glucuronide | 4.09 | 447 | 271, 175 | 151, 177 | P, U | |
| M5 | Naringenin-4′-O-glucuronide | 4.23 | 447 | 271, 175 | 151, 177 | P, U | |
| M6 | Eriodictyol-O-glucuronide | 4.23 | 463 | 287, 175 | 151, 269 | P, U | |
| M7 | Hesperitin-7-O-glucuronide | 4.45 | 477 | 301, 175 | 286, 243, 283 | P, U | |
| M8 | Hesperitin-3′-O-glucuronide | 4.63 | 477 | 301, 175 | 286, 243, 283 | P, U | |
| M9 | Naringenin-sulfate | 4.68 | 351 | 271 | 151, 177 | P | |
| M10 | Eriodictyol-sulfate | 4.78 | 367 | 287 | 151 | P, U | |
| M11 | Naringenin-sulfate | 4.82 | 351 | 271 | 151, 177, 165 | P | |
| M12 | Hesperetin-sulfate | 4.9 | 381 | 301 | 286, 243, 283, 199 | P, U |
| Id. | Compounds | 1 h | 4 h | ||
|---|---|---|---|---|---|
| Mean ± SEM | CV | Mean ± SEM | CV | ||
| M1 | Hesperitin-O-glucuronide-sulfate | 0.021 ± 0.004 | 19% | 0.019 ± 0.006 | 31% |
| M2 | Hesperitin-O-glucuronide-sulfate | 0.033 ± 0.008 | 24% | 0.094 ± 0.093 | 98% |
| M3 | Eriodictyol-O-glucuronide | 0.096 ± 0.010 | 10% | 0.085 ± 0.022 | 26% |
| M4 | Naringenin-7-O-glucuronide | 0.169 ± 0.032 | 19% | 0.163 ± 0.042 | 26% |
| M5 | Naringenin-4′-O-glucuronide | 0.183 ± 0.017 | 9% | 0.194 ± 0.050 | 26% |
| M6 | Eriodictyol-O-glucuronide | 0.095 ± 0.016 | 16% | 0.087 ± 0.045 | 52% |
| M7 | Hesperitin-7-O-glucuronide | 0.112 ± 0.011 | 10% | 0.162 ± 0.091 | 56% |
| M8 | Hesperitin-3′-O-glucuronide | 0.177 ± 0.020 | 11% | 0.411 ± 0.355 | 86% |
| M9 | Naringenin-sulfate | 0.085 ± 0.050 | 59% | 0.100 ± 0.091 | 91% |
| M10 | Eriodictyol-sulfate | 0.620 ± 0.195 | 31% | 0.960 ± 0.935 | 97% |
| M11 | Naringenin-sulfate | 0.059 ± 0.016 | 28% | 0.058 ± 0.059 | 102% |
| M12 | Hesperetin-sulfate | 0.451 ± 0.163 | 36% | 3.542 ± 4.562 | 129% |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Spigoni, V.; Mena, P.; Fantuzzi, F.; Tassotti, M.; Brighenti, F.; Bonadonna, R.C.; Del Rio, D.; Dei Cas, A. Bioavailability of Bergamot (Citrus bergamia) Flavanones and Biological Activity of Their Circulating Metabolites in Human Pro-Angiogenic Cells. Nutrients 2017, 9, 1328. https://doi.org/10.3390/nu9121328
Spigoni V, Mena P, Fantuzzi F, Tassotti M, Brighenti F, Bonadonna RC, Del Rio D, Dei Cas A. Bioavailability of Bergamot (Citrus bergamia) Flavanones and Biological Activity of Their Circulating Metabolites in Human Pro-Angiogenic Cells. Nutrients. 2017; 9(12):1328. https://doi.org/10.3390/nu9121328
Chicago/Turabian StyleSpigoni, Valentina, Pedro Mena, Federica Fantuzzi, Michele Tassotti, Furio Brighenti, Riccardo C. Bonadonna, Daniele Del Rio, and Alessandra Dei Cas. 2017. "Bioavailability of Bergamot (Citrus bergamia) Flavanones and Biological Activity of Their Circulating Metabolites in Human Pro-Angiogenic Cells" Nutrients 9, no. 12: 1328. https://doi.org/10.3390/nu9121328
APA StyleSpigoni, V., Mena, P., Fantuzzi, F., Tassotti, M., Brighenti, F., Bonadonna, R. C., Del Rio, D., & Dei Cas, A. (2017). Bioavailability of Bergamot (Citrus bergamia) Flavanones and Biological Activity of Their Circulating Metabolites in Human Pro-Angiogenic Cells. Nutrients, 9(12), 1328. https://doi.org/10.3390/nu9121328