Dietary Intake of Meat Cooking-Related Mutagens (HCAs) and Risk of Colorectal Adenoma and Cancer: A Systematic Review and Meta-Analysis




Abstract
:1. Introduction
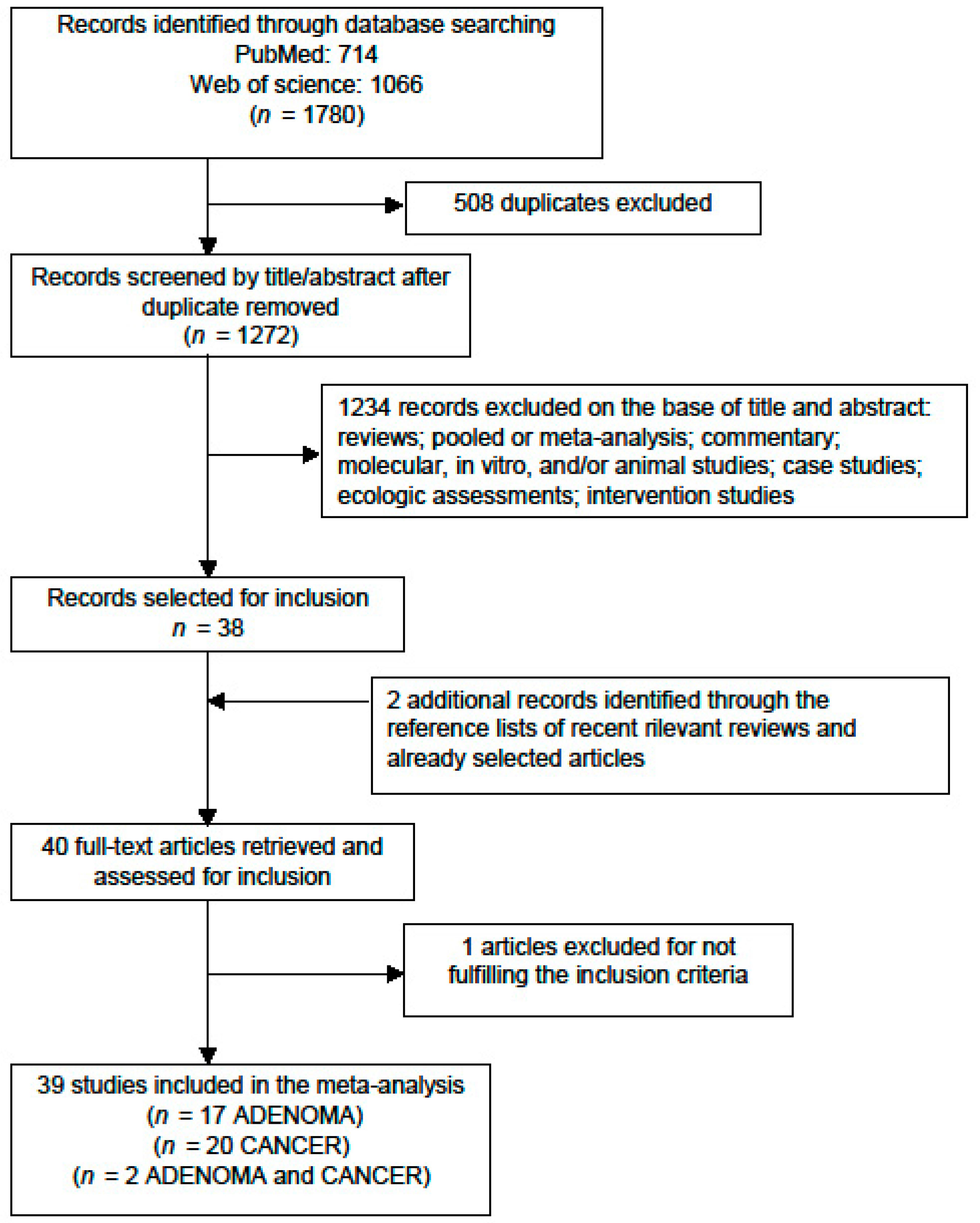
2. Materials and Methods
2.1. Search Strategy and Data Source
2.2. Inclusion Criteria
2.3. Study Selection
2.4. Data Extraction and Quality Assessment
2.5. Statistical Analysis
3. Results
3.1. Colorectal Adenoma (CRA)
3.1.1. Study Characteristics and Quality Assessment
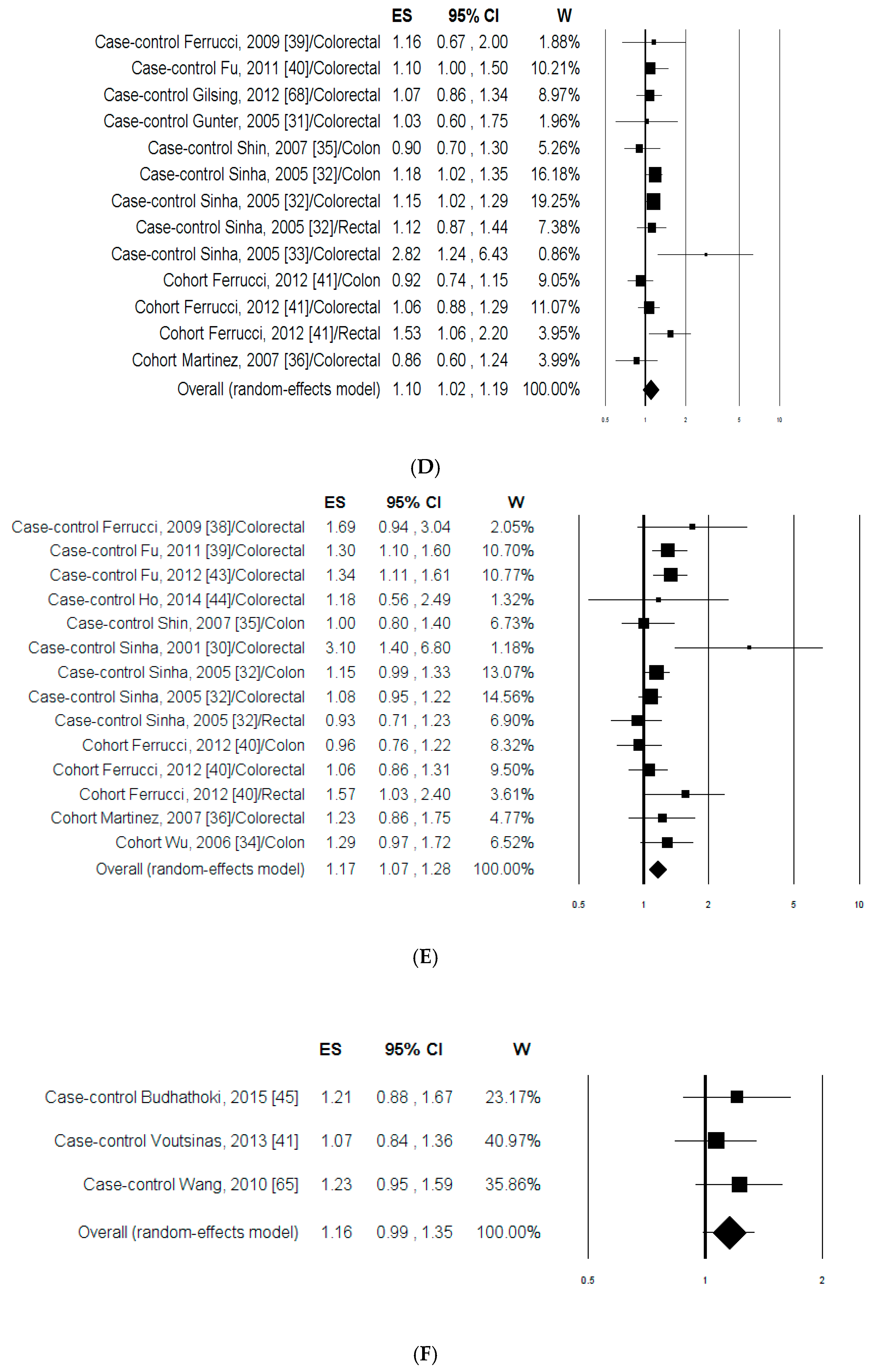
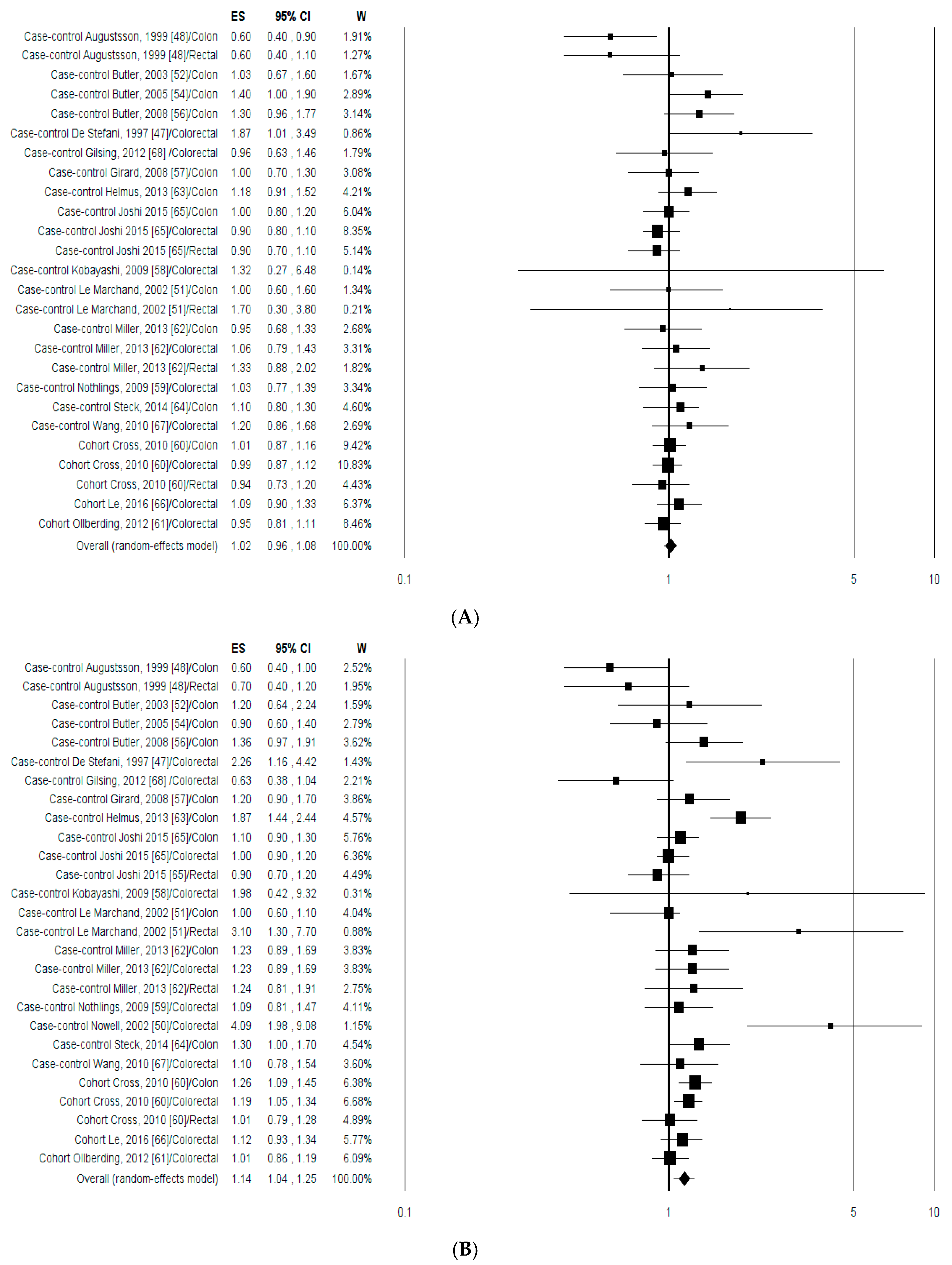
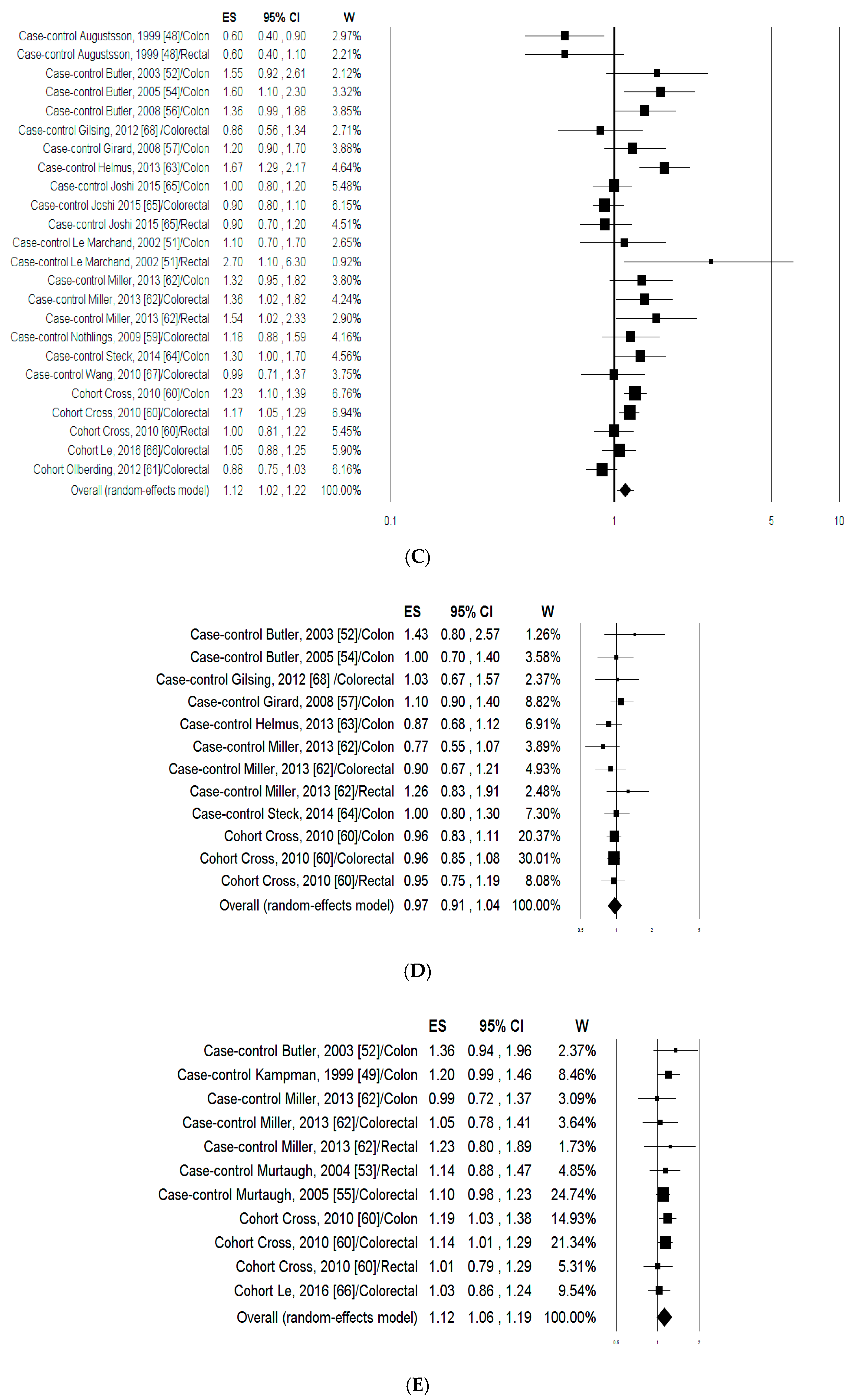
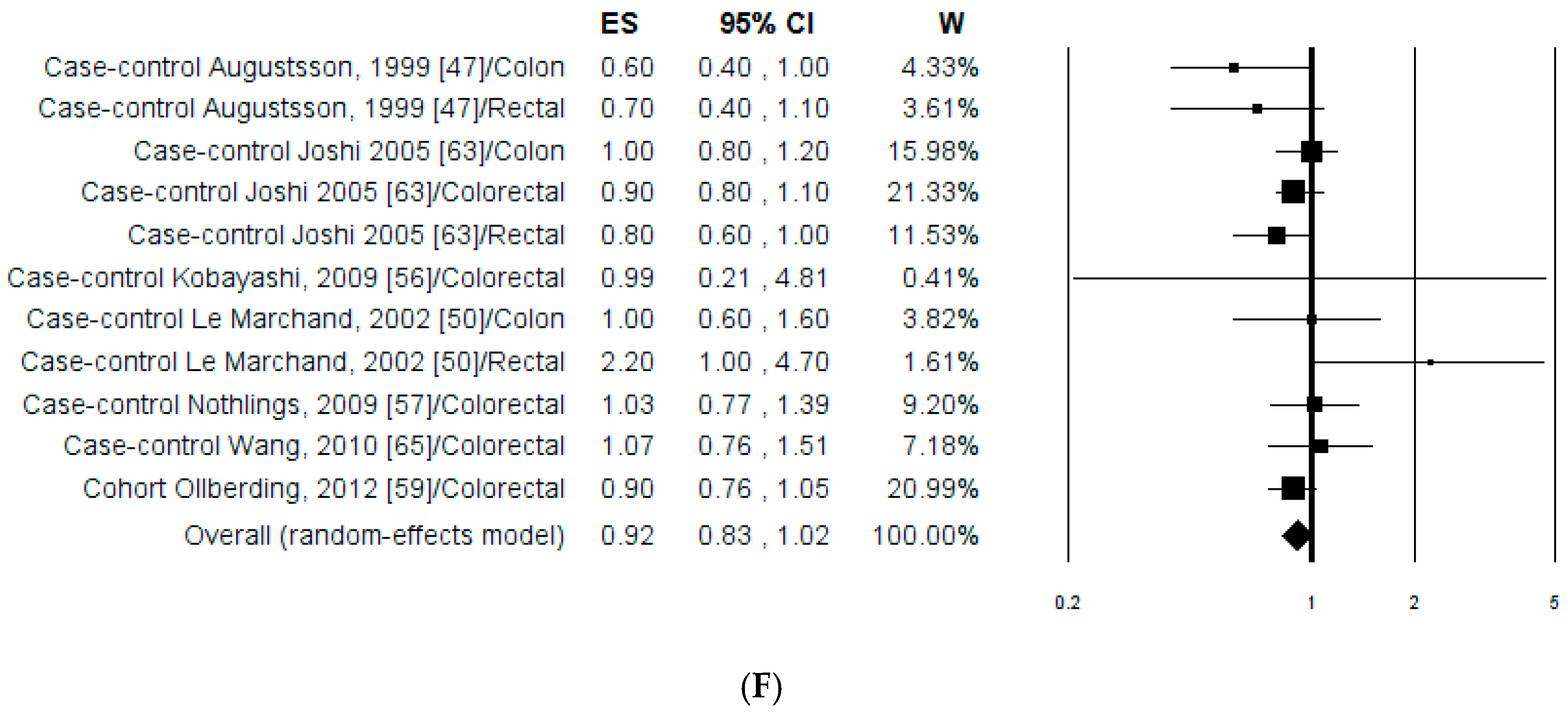
3.1.2. Meta-Analysis
3.1.3. Publication Bias and Sensitivity Analysis
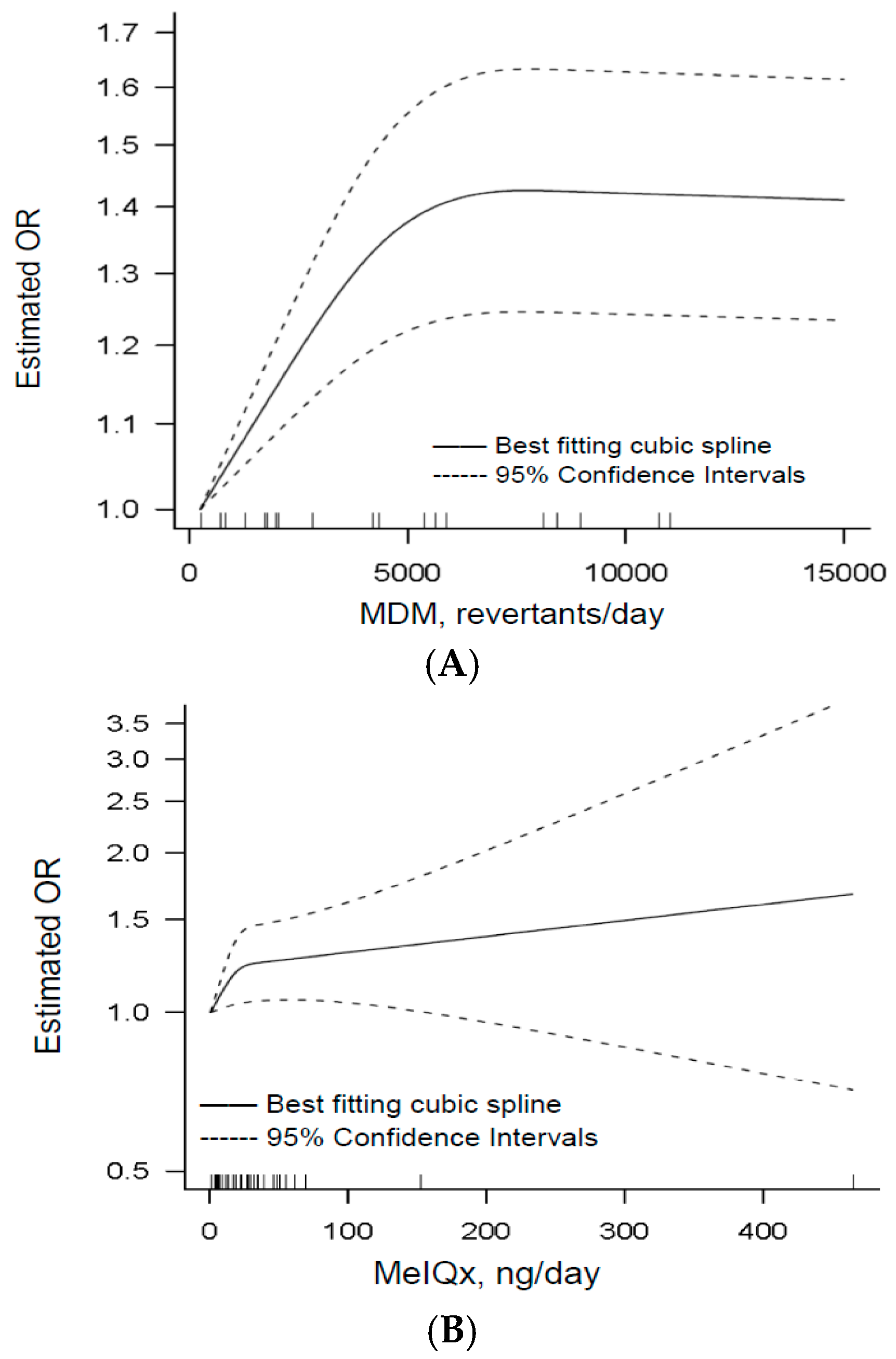
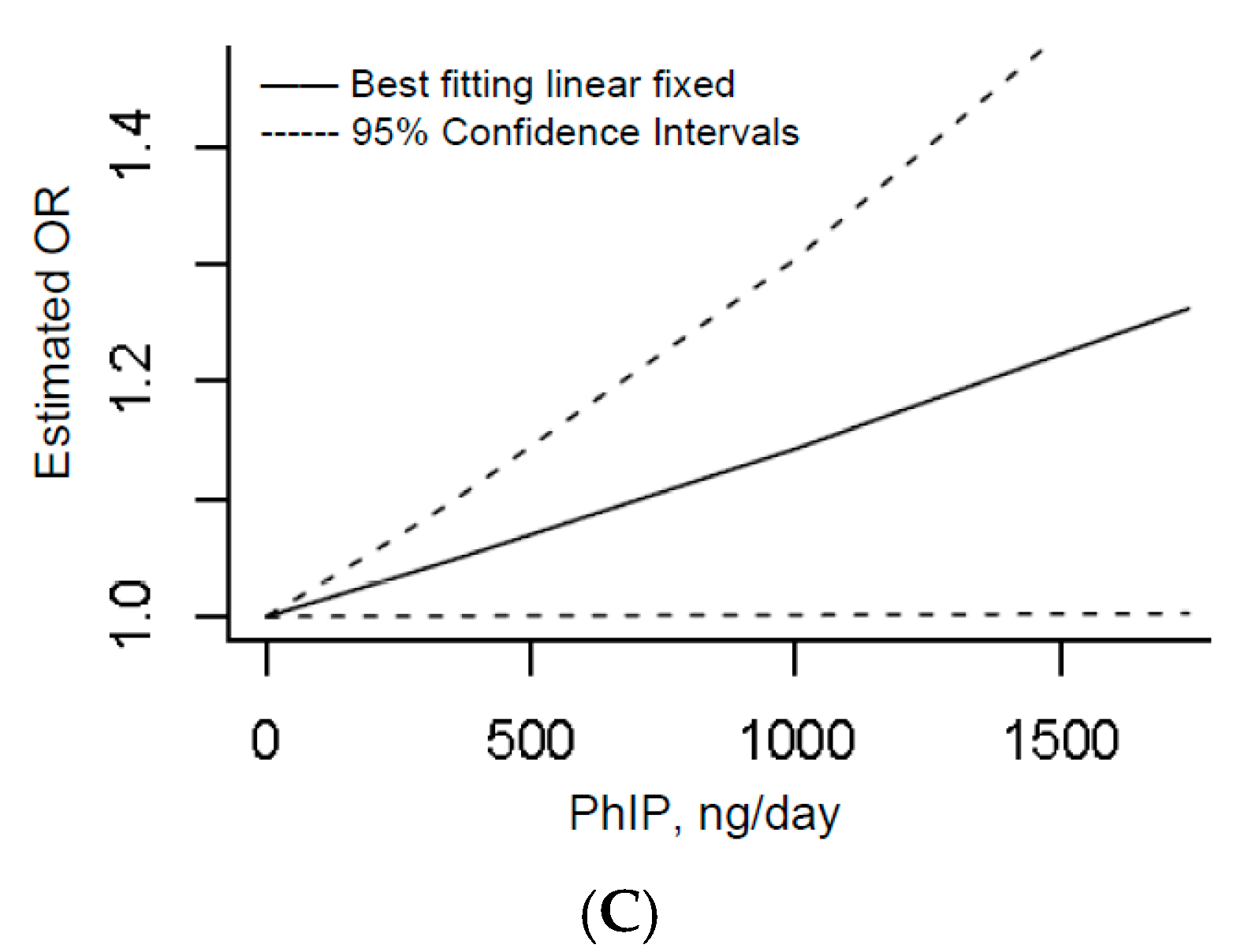
3.1.4. Dose–Response
3.2. Colorectal Cancer (CRC)
3.2.1. Study Characteristics and Quality Assessment
3.2.2. Meta-Analysis
3.2.3. Publication Bias and Sensitivity Analysis
3.2.4. Dose–Response
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in GLOBOCAN 2012. Int. J. Cancer 2015, 136, E359–E386. [Google Scholar] [CrossRef] [PubMed]
- Chan, D.S.; Lau, R.; Aune, D.; Vieira, R.; Greenwood, D.C.; Kampman, E.; Norat, T. Red and processed meat and colorectal cancer incidence: Meta-analysis of prospective studies. PLoS ONE 2011, 6, e20456. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Yu, E.; Gao, X.; Song, N.; Liu, L.; Wei, X.; Zhang, W.; Fu, C. Red and processed meat intake and risk of colorectal adenomas: A meta-analysis of observational studies. Int. J. Cancer 2013, 132, 437–448. [Google Scholar] [CrossRef] [PubMed]
- Alexander, D.D.; Weed, D.L.; Miller, P.E.; Mohamed, M.A. Red Meat and Colorectal Cancer: A Quantitative Update on the State of the Epidemiologic Science. J. Am. Coll. Nutr. 2015, 34, 521–543. [Google Scholar] [CrossRef] [PubMed]
- Bouvard, V.; Loomis, D.; Guyton, K.Z.; Grosse, Y.; Ghissassi, F.E.; Benbrahim-Tallaa, L.; Guha, N.; Mattock, H.; Straif, K. International Agency for Research on Cancer Monograph Working Group. Carcinogenicity of consumption of red and processed meat. Lancet Oncol. 2015, 16, 1599–1600. [Google Scholar] [CrossRef]
- Abid, Z.; Cross, A.J.; Sinha, R. Meat, dairy, and cancer. Am. J. Clin. Nutr. 2014, 100, 386S–393S. [Google Scholar] [CrossRef] [PubMed]
- Bastide, N.M.; Pierre, F.H.; Corpet, D.E. Heme iron from meat and risk of colorectal cancer: A meta-analysis and a review of the mechanisms involved. Cancer Prev. Res. 2011, 4, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Cross, A.J.; Sinha, R. Meat-related mutagens/carcinogens in the etiology of colorectal cancer. Environ. Mol. Mutagen. 2004, 44, 44–55. [Google Scholar] [CrossRef] [PubMed]
- Turesky, R.J. Formation and biochemistry of carcinogenic heterocyclic aromatic amines in cooked meats. Toxicol. Lett. 2007, 168, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Sugimura, T.; Wakabayashi, K.; Nakagama, H.; Nagao, M. Heterocyclic amines: Mutagens/carcinogens produced during cooking of meat and fish. Cancer Sci. 2004, 95, 290–299. [Google Scholar] [CrossRef] [PubMed]
- International Agency for Research on Cancer. Some naturally occurring substances: Food items and constituents, heterocyclic aromatic amines and mycotoxins. IARC Monogr. Eval. Carcinog. Risks Hum. 1993, 56, 211–242. [Google Scholar]
- Zheng, W.; Lee, S.A. Well-done meat intake, heterocyclic amine exposure, and cancer risk. Nutr. Cancer 2009, 61, 437–446. [Google Scholar] [CrossRef] [PubMed]
- Sinha, R.; Chow, W.H.; Kulldorff, M.; Denobile, J.; Butler, J.; Garcia-Closas, M.; Weil, R.; Hoover, R.N.; Rothman, N. Well-done, grilled red meat increases the risk of colorectal adenomas. Cancer Res. 1999, 59, 4320–4324. [Google Scholar] [PubMed]
- Augustsson, K.; Skog, K.; Jägerstad, M.; Steineck, G. Assessment of the human exposure to heterocyclic amines. Carcinogenesis 1997, 18, 1931–1935. [Google Scholar] [CrossRef] [PubMed]
- Sinha, R.; Rothman, N. Exposure assessment of heterocyclic amines (HCAs) in epidemiologic studies. Mutat. Res. 1997, 376, 195–202. [Google Scholar] [CrossRef]
- Voskuil, D.W.; Augustsson, K.; Dickman, P.W.; van’t Veer, P.; Steineck, G. Assessing the human intake of heterocyclic amines: Limited loss of information using reduced sets of questions. Cancer Epidemiol. Biomark. Prev. 1999, 8, 809–814. [Google Scholar]
- Cantwell, M.; Mittl, B.; Curtin, J.; Carroll, R.; Potischman, N.; Caporaso, N.; Sinha, R. Relative validity of a food frequency questionnaire with a meat-cooking and heterocyclic amine module. Cancer Epidemiol. Biomark. Prev. 2004, 13, 293–298. [Google Scholar] [CrossRef]
- Sinha, R.; Cross, A.; Curtin, J.; Zimmerman, T.; McNutt, S.; Risch, A.; Holden, J. Development of a food frequency questionnaire module and databases for compounds in cooked and processed meats. Mol. Nutr. Food Res. 2005, 49, 648–655. [Google Scholar] [CrossRef] [PubMed]
- Deziel, N.C.; Buckley, T.J.; Sinha, R.; Abubaker, S.; Platz, E.A.; Strickland, P.T. Comparability and repeatability of methods for estimating the dietary intake of the heterocyclic amine contaminant 2-amino-1-methyl-6-phenylimidazo[4,5b]pyridine (PhIP). Food Addit. Contam. Part A 2012, 29, 1202–1211. [Google Scholar] [CrossRef] [PubMed]
- Stroup, D.F.; Berlin, J.A.; Morton, S.C.; Olkin, I.; Williamson, G.D.; Rennie, D.; Moher, D.; Becker, B.J.; Sipe, T.A.; Thacker, S.B. Meta-analysis of observational studies in epidemiology: A proposal for reporting. Meta-analysis Of Observational Studies in Epidemiology (MOOSE) group. JAMA 2000, 283, 2008–2012. [Google Scholar] [CrossRef] [PubMed]
- Liberati, A.; Altman, D.G.; Tetzlaff, J.; Mulrow, C.; Gøtzsche, P.C.; Ioannidis, J.P.; Clarke, M.; Devereaux, P.J.; Kleijnen, J.; Moher, D. The PRISMA statement for reporting systematic reviews and meta-analyses of studies that evaluate healthcare interventions: Explanation and elaboration. BMJ 2009, 339, b2700. [Google Scholar] [CrossRef] [PubMed]
- Wells, G.A.; Shea, B.; O’Connell, D.; Peterson, J.; Welch, V.; Losos, M.; Tugwell, P. The Newcastle-Ottawa Scale (NOS) for Assessing the Quality of Non-Randomised Studies in Meta-Analyses. 2015. Available online: http://www.ohri.ca/programs/clinical_epidemiology/oxford.asp (accessed on 1 March 2017).
- Greenland, S.; Longnecker, M.P. Methods for trend estimation from summarized dose-response data, with applications to meta-analysis. Am. J. Epidemiol. 1992, 135, 1301–1309. [Google Scholar] [CrossRef] [PubMed]
- Orsini, N.; Li, R.; Wolk, A.; Khudyakov, P.; Spiegelman, D. Meta-analysis for linear and nonlinear dose-response relations: Examples, an evaluation of approximations, and software. Am. J. Epidemiol. 2012, 175, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Cook, N.R.; Bergström, A. A two-stage hierarchical regression model for meta-analysis of epidemiologic nonlinear dose–response data. Comp. Stat. Data Anal. 2009, 53, 4157–4167. [Google Scholar] [CrossRef]
- Higgins, J.P.; Thompson, S.G. Quantifying heterogeneity in a meta-analysis. Stat. Med. 2002, 21, 1539–1558. [Google Scholar] [CrossRef] [PubMed]
- Higgins, J.P.; Thompson, S.G.; Deeks, J.J. Measuring inconsistency in meta-analyses. BMJ 2003, 327, 557–560. [Google Scholar] [CrossRef] [PubMed]
- Egger, M.; Davey Smith, G.; Schneider, M. Bias in meta-analysis detected by a simple, graphical test. BMJ 1997, 315, 629–634. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014. Available online: http://www.R-project.org/ (accessed on 31 January 2017).
- Sinha, R.; Kulldorff, M.; Chow, W.H.; Denobile, J.; Rothman, N. Dietary intake of heterocyclic amines, meat-derived mutagenic activity, and risk of colorectal adenomas. Cancer Epidemiol. Biomark. Prev. 2001, 10, 559–562. [Google Scholar]
- Gunter, M.J.; Probst-Hensch, N.M.; Cortessis, V.K.; Kulldorff, M.; Haile, R.W.; Sinha, R. Meat intake, cooking-related mutagens and risk of colorectal adenoma in a sigmoidoscopy-based case-control study. Carcinogenesis 2005, 26, 637–642. [Google Scholar] [CrossRef] [PubMed]
- Sinha, R.; Peters, U.; Cross, A.J.; Kulldorff, M.; Weissfeld, J.L.; Pinsky, P.F.; Rothman, N.; Hayes, R.B. Meat, meat cooking methods and preservation, and risk for colorectal adenoma. Cancer Res. 2005, 65, 8034–8041. [Google Scholar] [PubMed]
- Sinha, R.; Kulldorff, M.; Gunter, M.J.; Strickland, P.; Rothman, N. Dietary benzo[a]pyrene intake and risk of colorectal adenoma. Cancer Epidemiol. Biomark. Prev. 2005, 14, 2030–2034. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; Giovannucci, E.; Byrne, C.; Platz, E.A.; Fuchs, C.; Willett, W.C.; Sinha, R. Meat mutagens and risk of distal colon adenoma in a cohort of U.S. men. Cancer Epidemiol. Biomark. Prev. 2006, 15, 1120–1125. [Google Scholar] [CrossRef] [PubMed]
- Shin, A.; Shrubsole, M.J.; Ness, R.M.; Wu, H.; Sinha, R.; Smalley, W.E.; Shyr, Y.; Zheng, W. Meat and meat-mutagen intake, doneness preference and the risk of colorectal polyps: The Tennessee Colorectal Polyp Study. Int. J. Cancer 2007, 121, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Martínez, M.E.; Jacobs, E.T.; Ashbeck, E.L.; Sinha, R.; Lance, P.; Alberts, D.S.; Thompson, P.A. Meat intake, preparation methods, mutagens and colorectal adenoma recurrence. Carcinogenesis 2007, 28, 2019–2027. [Google Scholar] [CrossRef] [PubMed]
- Shin, A.; Shrubsole, M.J.; Rice, J.M.; Cai, Q.; Doll, M.A.; Long, J.; Smalley, W.E.; Shyr, Y.; Sinha, R.; Ness, R.M.; et al. Meat intake, heterocyclic amine exposure, and metabolizing enzyme polymorphisms in relation to colorectal polyp risk. Cancer Epidemiol. Biomark. Prev. 2008, 17, 320–329. [Google Scholar] [CrossRef] [PubMed]
- Rohrmann, S.; Hermann, S.; Linseisen, J. Heterocyclic aromatic amine intake increases colorectal adenoma risk: Findings from a prospective European cohort study. Am. J. Clin. Nutr. 2009, 89, 1418–1424. [Google Scholar] [CrossRef] [PubMed]
- Ferrucci, L.M.; Sinha, R.; Graubard, B.I.; Mayne, S.T.; Ma, X.; Schatzkin, A.; Schoenfeld, P.S.; Cash, B.D.; Flood, A.; Cross, A.J. Dietary meat intake in relation to colorectal adenoma in asymptomatic women. Am. J. Gastroenterol. 2009, 104, 1231–1240. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.; Shrubsole, M.J.; Smalley, W.E.; Wu, H.; Chen, Z.; Shyr, Y.; Ness, R.M.; Zheng, W. Association of meat intake and meat-derived mutagen exposure with the risk of colorectal polyps by histologic type. Cancer Prev. Res. 2011, 4, 1686–1697. [Google Scholar] [CrossRef] [PubMed]
- Ferrucci, L.M.; Sinha, R.; Huang, W.Y.; Berndt, S.I.; Katki, H.A.; Schoen, R.E.; Hayes, R.B.; Cross, A.J. Meat consumption and the risk of incident distal colon and rectal adenoma. Br. J. Cancer 2012, 106, 608–616. [Google Scholar] [CrossRef] [PubMed]
- Voutsinas, J.; Wilkens, L.R.; Franke, A.; Vogt, T.M.; Yokochi, L.A.; Decker, R.; Le Marchand, L. Heterocyclic amine intake, smoking, cytochrome P450 1A2 and N-acetylation phenotypes, and risk of colorectal adenoma in a multiethnic population. Gut 2013, 62, 416–422. [Google Scholar] [CrossRef] [PubMed]
- Barbir, A.; Linseisen, J.; Hermann, S.; Kaaks, R.; Teucher, B.; Eichholzer, M.; Rohrmann, S. Effects of phenotypes in heterocyclic aromatic amine (HCA) metabolism-related genes on the association of HCA intake with the risk of colorectal adenomas. Cancer Causes Control 2012, 23, 1429–1442. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.; Shrubsole, M.; Li, G.; Smalley, W.E.; Hein, D.W.; Chen, Z.; Shyr, Y.; Cai, Q.; Ness, R.M.; Zheng, W. Using gene-environment interaction analyses to clarify the role of well-done meat and heterocyclic amine exposure in the etiology of colorectal polyps. Am. J. Clin. Nutr. 2012, 96, 1119–1128. [Google Scholar] [CrossRef] [PubMed]
- Ho, V.; Peacock, S.; Massey, T.E.; Ashbury, J.E.; Vanner, S.J.; King, W.D. Meat-derived carcinogens, genetic susceptibility and colorectal adenoma risk. Genes Nutr. 2014, 9, 430. [Google Scholar] [CrossRef] [PubMed]
- Budhathoki, S.; Iwasaki, M.; Yamaji, T.; Sasazuki, S.; Takachi, R.; Sakamoto, H.; Yoshida, T.; Tsugane, S. Dietary heterocyclic amine intake, NAT2 genetic polymorphism, and colorectal adenoma risk: The colorectal adenoma study in Tokyo. Cancer Epidemiol. Biomark. Prev. 2015, 24, 613–620. [Google Scholar] [CrossRef] [PubMed]
- De Stefani, E.; Deneo-Pellegrini, H.; Mendilaharsu, M.; Ronco, A. Meat intake, heterocyclic amines and risk of colorectal cancer. Int. J. Oncol. 1997, 10, 573–580. [Google Scholar]
- Augustsson, K.; Skog, K.; Jägerstad, M.; Dickman, P.W.; Steineck, G. Dietary heterocyclic amines and cancer of the colon, rectum, bladder, and kidney: A population-based study. Lancet 1999, 353, 703–707. [Google Scholar] [CrossRef]
- Kampman, E.; Slattery, M.L.; Bigler, J.; Leppert, M.; Samowitz, W.; Caan, B.J.; Potter, J.D. Meat consumption, genetic susceptibility, and colon cancer risk: A United States multicenter case-control study. Cancer Epidemiol. Biomark. Prev. 1999, 8, 15–24. [Google Scholar]
- Nowell, S.; Coles, B.; Sinha, R.; MacLeod, S.; Luke Ratnasinghe, D.; Stotts, C.; Kadlubar, F.F.; Ambrosone, C.B.; Lang, N.P. Analysis of total meat intake and exposure to individual heterocyclic amines in a case-control study of colorectal cancer: Contribution of metabolic variation to risk. Mutat. Res. 2002, 506, 175–185. [Google Scholar] [CrossRef]
- Le Marchand, L.; Hankin, J.H.; Pierce, L.M.; Sinha, R.; Nerurkar, P.V.; Franke, A.A.; Wilkens, L.R.; Kolonel, L.N.; Donlon, T.; Seifried, A.; et al. Well-done red meat, metabolic phenotypes and colorectal cancer in Hawaii. Mutat. Res. 2002, 506, 205–214. [Google Scholar] [CrossRef]
- Butler, L.M.; Sinha, R.; Millikan, R.C.; Martin, C.F.; Newman, B.; Gammon, M.D.; Ammerman, A.S.; Sandler, R.S. Heterocyclic amines, meat intake, and association with colon cancer in a population-based study. Am. J. Epidemiol. 2003, 157, 434–445. [Google Scholar] [CrossRef] [PubMed]
- Murtaugh, M.A.; Ma, K.N.; Sweeney, C.; Caan, B.J.; Slattery, M.L. Meat consumption patterns and preparation, genetic variants of metabolic enzymes, and their association with rectal cancer in men and women. J. Nutr. 2004, 134, 776–784. [Google Scholar] [PubMed]
- Butler, L.M.; Duguay, Y.; Millikan, R.C.; Sinha, R.; Gagné, J.F.; Sandler, R.S.; Guillemette, C. Joint effects between UDP-glucuronosyltransferase 1A7 genotype and dietary carcinogen exposure on risk of colon cancer. Cancer Epidemiol. Biomark. Prev. 2005, 14, 1626–1632. [Google Scholar] [CrossRef] [PubMed]
- Murtaugh, M.A.; Sweeney, C.; Ma, K.N.; Caan, B.J.; Slattery, M.L. The CYP1A1 genotype may alter the association of meat consumption patterns and preparation with the risk of colorectal cancer in men and women. J. Nutr. 2005, 135, 179–186. [Google Scholar] [PubMed]
- Butler, L.M.; Millikan, R.C.; Sinha, R.; Keku, T.O.; Winkel, S.; Harlan, B.; Eaton, A.; Gammon, M.D.; Sandler, R.S. Modification by N-acetyltransferase 1 genotype on the association between dietary heterocyclic amines and colon cancer in a multiethnic study. Mutat. Res. 2008, 638, 162–174. [Google Scholar] [CrossRef] [PubMed]
- Girard, H.; Butler, L.M.; Villeneuve, L.; Millikan, R.C.; Sinha, R.; Sandler, R.S.; Guillemette, C. UGT1A1 and UGT1A9 functional variants, meat intake, and colon cancer, among Caucasians and African-Americans. Mutat. Res. 2008, 644, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, M.; Otani, T.; Iwasaki, M.; Natsukawa, S.; Shaura, K.; Koizumi, Y.; Kasuga, Y.; Sakamoto, H.; Yoshida, T.; Tsugane, S. Association between dietary heterocyclic amine levels, genetic polymorphisms of NAT2, CYP1A1, and CYP1A2 and risk of colorectal cancer: A hospital-based case-control study in Japan. Scand. J. Gastroenterol. 2009, 44, 952–959. [Google Scholar] [CrossRef] [PubMed]
- Nöthlings, U.; Yamamoto, J.F.; Wilkens, L.R.; Murphy, S.P.; Park, S.Y.; Henderson, B.E.; Kolonel, L.N.; Le Marchand, L. Meat and heterocyclic amine intake, smoking, NAT1 and NAT2 polymorphisms, and colorectal cancer risk in the multiethnic cohort study. Cancer Epidemiol. Biomark. Prev. 2009, 18, 2098–2106. [Google Scholar] [CrossRef] [PubMed]
- Cross, A.J.; Ferrucci, L.M.; Risch, A.; Graubard, B.I.; Ward, M.H.; Park, Y.; Hollenbeck, A.R.; Schatzkin, A.; Sinha, R. A large prospective study of meat consumption and colorectal cancer risk: An investigation of potential mechanisms underlying this association. Cancer Res. 2010, 70, 2406–2414. [Google Scholar] [CrossRef] [PubMed]
- Ollberding, N.J.; Wilkens, L.R.; Henderson, B.E.; Kolonel, L.N.; Le Marchand, L. Meat consumption, heterocyclic amines and colorectal cancer risk: The Multiethnic Cohort Study. Int. J. Cancer 2012, 131, E1125–E1133. [Google Scholar] [CrossRef] [PubMed]
- Miller, P.E.; Lazarus, P.; Lesko, S.M.; Cross, A.J.; Sinha, R.; Laio, J.; Zhu, J.; Harper, G.; Muscat, J.E.; Hartman, T.J. Meat-related compounds and colorectal cancer risk by anatomical subsite. Nutr. Cancer 2013, 65, 202–226. [Google Scholar] [CrossRef] [PubMed]
- Helmus, D.S.; Thompson, C.L.; Zelenskiy, S.; Tucker, T.C.; Li, L. Red meat-derived heterocyclic amines increase risk of colon cancer: A population-based case-control study. Nutr. Cancer 2013, 65, 1141–1150. [Google Scholar] [CrossRef] [PubMed]
- Steck, S.E.; Butler, L.M.; Keku, T.; Antwi, S.; Galanko, J.; Sandler, R.S.; Hu, J.J. Nucleotide excision repair gene polymorphisms, meat intake and colon cancer risk. Mutat. Res. 2014, 762, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Joshi, A.D.; Kim, A.; Lewinger, J.P.; Ulrich, C.M.; Potter, J.D.; Cotterchio, M.; Le Marchand, L.; Stern, M.C. Meat intake, cooking methods, dietary carcinogens, and colorectal cancer risk: Findings from the Colorectal Cancer Family Registry. Cancer Med. 2015, 4, 936–952. [Google Scholar] [CrossRef] [PubMed]
- Le, N.T.; Michels, F.A.; Song, M.; Zhang, X.; Bernstein, A.M.; Giovannucci, E.L.; Fuchs, C.S.; Ogino, S.; Chan, A.T.; Sinha, R.; et al. A Prospective Analysis of Meat Mutagens and Colorectal Cancer in the Nurses’ Health Study and Health Professional Follow-up Study. Environ. Health Perspect. 2016, 124, 1529–1536. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yamamoto, J.F.; Caberto, C.; Saltzman, B.; Decker, R.; Vogt, T.M.; Yokochi, L.; Chanock, S.; Wilkens, L.R.; Le Marchand, L. Genetic variation in the bioactivation pathway for polycyclic hydrocarbons and heterocyclic amines in relation to risk of colorectal neoplasia. Carcinogenesis 2011, 32, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Gilsing, A.M.; Berndt, S.I.; Ruder, E.H.; Graubard, B.I.; Ferrucci, L.M.; Burdett, L.; Weissfeld, J.L.; Cross, A.J.; Sinha, R. Meat-related mutagen exposure, xenobiotic metabolizing gene polymorphisms and the risk of advanced colorectal adenoma and cancer. Carcinogenesis 2012, 33, 1332–1339. [Google Scholar] [CrossRef] [PubMed]
- National Cancer Institute: Division of Cancer Epidemiology & Genetics. CHARRED: Computerized Heterocyclic Amines Resource for Research in Epidemiology of Disease. 2006. Available online: http://dceg.cancer.gov/neb/tools/charred (accessed on 31 January 2017).
- Leslie, A.; Carey, F.A.; Pratt, N.R.; Steele, R.J. The colorectal adenoma-carcinoma sequence. Br. J. Surg. 2002, 89, 845–860. [Google Scholar] [CrossRef] [PubMed]
- Schatzkin, A.; Gail, M. The promise and peril of surrogate end points in cancer research. Nat. Rev. Cancer 2002, 2, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Chen, Y.; Wang, X.; Wang, J.; Yan, Z.; Gong, G.; Li, G.; Li, C. Meta-analysis of prospective cohort studies of cigarette smoking and the incidence of colon and rectal cancers. Eur. J. Cancer Prev. 2015, 24, 6–15. [Google Scholar] [CrossRef] [PubMed]
- Bastide, N.; Morois, S.; Cadeau, C.; Kangas, S.; Serafini, M.; Gusto, G.; Dossus, L.; Pierre, F.H.; Clavel-Chapelon, F.; Boutron-Ruault, M.C. Heme Iron Intake, Dietary Antioxidant Capacity, and Risk of Colorectal Adenomas in a Large Cohort Study of French Women. Cancer Epidemiol. Biomark. Prev. 2016, 25, 640–647. [Google Scholar] [CrossRef] [PubMed]
- Iacopetta, B. Are there two sides to colorectal cancer? Int. J. Cancer 2002, 101, 403–408. [Google Scholar] [CrossRef] [PubMed]
- Le Marchand, L.; Donlon, T.; Seifried, A.; Wilkens, L.R. Red meat intake, CYP2E1 genetic polymorphisms, and colorectal cancer risk. Cancer Epidemiol. Biomark. Prev. 2002, 11, 1019–1024. [Google Scholar]
- Povey, A.C.; Hall, C.N.; Badawi, A.F.; Cooper, D.P.; O’Connor, P.J. Elevated levels of the pro-carcinogenic adduct, O(6)-methylguanine, in normal DNA from the cancer prone regions of the large bowel. Gut 2000, 47, 362–365. [Google Scholar] [CrossRef] [PubMed]
- Bleday, R.; Braidt, J.; Ruoff, K.; Shellito, P.C.; Ackroyd, F.W. Quantitative cultures of the mucosal-associated bacteria in the mechanically prepared colon and rectum. Dis. Colon Rectum 1993, 36, 844–849. [Google Scholar] [CrossRef] [PubMed]
- Bultman, S.J. The microbiome and its potential as a cancer preventive intervention. Semin. Oncol. 2016, 43, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, L.R. Meat and cancer. Meat Sci. 2010, 84, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Magalhães, B.; Peleteiro, B.; Lunet, N. Dietary patterns and colorectal cancer: Systematic review and meta-analysis. Eur. J. Cancer Prev. 2012, 21, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.L.; Shu, L.; Zheng, P.F.; Zhang, X.Y.; Si, C.J.; Yu, X.L.; Gao, W.; Zhang, L. Dietary patterns and colorectal cancer risk: A meta-analysis. Eur. J. Cancer Prev. 2017, 26, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Mehta, R.S.; Song, M.; Nishihara, R.; Drew, D.A.; Wu, K.; Qian, Z.R.; Fung, T.T.; Hamada, T.; Masugi, Y.; da Silva, A.; et al. Dietary Patterns and Risk of Colorectal Cancer: Analysis by Tumor Location and Molecular Subtypes. Gastroenterology 2017. [Google Scholar] [CrossRef] [PubMed]
- Wada, K.; Oba, S.; Tsuji, M.; Tamura, T.; Konishi, K.; Goto, Y.; Mizuta, F.; Koda, S.; Hori, A.; Tanabashi, S.; et al. Meat consumption and colorectal cancer risk in Japan: The Takayama study. Cancer Sci. 2017. [Google Scholar] [CrossRef] [PubMed]
- Felton, J.S.; Knize, M.G.; Hatch, F.T.; Tanga, M.J.; Colvin, M.E. Heterocyclic amine formation and the impact of structure on their mutagenicity. Cancer Lett. 1999, 143, 127–134. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Combined Risk Estimate | Test of Heterogeneity | Publication Bias | |||||
|---|---|---|---|---|---|---|---|
| Mutagen | Value (95% CI) | p | Q | I2 % | p | p (Egger Test) | p (Begg Test) |
| PhIP | |||||||
| Case-Control (n = 15) 2 | 1.19 (1.11–1.28) | <0.001 | 17.80 | 21.34 | 0.216 | 0.374 | 0.520 |
| Cohort (n = 8 ) | 1.23 (1.08–1.41) | 0.002 | 10.48 | 33.21 | 0.163 | 0.576 | 0.621 |
| Polled (n = 23 ) | 1.20 (1.13–1.28) | <0.001 | 28.55 | 22.94 | 0.158 | 0.242 | 0.279 |
| Colon (n = 5) | 1.16 (1.05–1.29) | 0.003 | 3.70 | 0.00 | 0.448 | 0.656 | 0.624 |
| Rectal (n = 3) | 1.23 (0.86–1.76) | 0.248 | 4.91 | 59.23 | 0.086 | 0.695 | 0.602 |
| Colorectal (n = 15) | 1.22 (1.13–1.32) | <0.001 | 19.48 | 28.14 | 0.147 | 0.363 | 0.586 |
| MeIQx | |||||||
| Case-Control (n = 14) | 1.14 (1.03–1.27) | 0.009 | 29.49 | 52.52 | 0.009 | 0.716 | 0.882 |
| Cohort (n = 6) | 1.11 (0.99–1.25) | 0.068 | 5.01 | 0.21 | 0.415 | 0.194 | 0.573 |
| Polled (n = 21) | 1.14 (1.05–1.23) | 0.001 | 34.70 | 42.37 | 0.022 | 0.559 | 0.365 |
| Colon (n = 4) | 1.08 (0.93–1.26) | 0.303 | 4.94 | 39.28 | 0.176 | 0.584 | 0.497 |
| Rectal (n = 2) | – | ||||||
| Colorectal (n = 15) | 1.19 (1.09–1.30) | <0.001 | 21.49 | 34.85 | 0.090 | 0.279 | 0.125 |
| DiMeIQx | |||||||
| Case-Control (n = 13) | 1.16 (1.06–1.27) | 0.001 | 20.47 | 41.38 | 0.059 | 0.266 | 0.393 |
| Cohort (n = 6) | 1.06 (0.95–1.19) | 0.308 | 6.15 | 18.73 | 0.292 | 0.068 | 0.091 |
| Polled (n = 19) | 1.13 (1.05–1.21) | 0.001 | 28.39 | 36.59 | 0.056 | 0.190 | 0.278 |
| Colon (n = 4) | 1.06 (0.96–1.16) | 0.275 | 3.04 | 1.39 | 0.385 | 0.634 | 1.000 |
| Rectal (n = 2) | – | ||||||
| Colorectal (n = 13) | 1.19 (1.09–1.33) | <0.001 | 18.96 | 36.71 | 0.090 | 0.111 | 0.714 |
| B(a)P | |||||||
| Case-Control (n = 9) | 1.13 (1.05–1.21) | 0.001 | 7.70 | 0.00 | 0.463 | 0.770 | 1.000 |
| Cohort (n = 4) | 1.05 (0.86–1.28) | 0.663 | 6.56 | 54.24 | 0.087 | 0.709 | 0.497 |
| Polled (n = 13) | 1.10 (1.02–1.19) | 0.017 | 15.86 | 24.35 | 0.198 | 0.855 | 0.542 |
| Colon (n = 3) | 1.02 (0.84–1.24) | 0.832 | 4.88 | 58.99 | 0.087 | 0.254 | 0.602 |
| Rectal (n = 2) | – | ||||||
| Colorectal (n = 8) | 1.10 (1.01–1.20) | 0.026 | 7.61 | 8.05 | 0.368 | 0.739 | 0.458 |
| MDM | |||||||
| Case-Control (n = 9) | 1.18 (1.05–1.33) | 0.005 | 16.09 | 50.28 | 0.041 | 0.256 | 0.297 |
| Cohort (n = 5) | 1.14 (0.99–1.33) | 0.077 | 5.54 | 27.77 | 0.236 | 0.071 | 0.142 |
| Polled (n = 14) | 1.17 (1.07–1.28) | 0.001 | 21.81 | 40.41 | 0.058 | 0.128 | 0.208 |
| Colon (n = 4) | 1.10 (0.98–1.23) | 0.092 | 3.25 | 7.68 | 0.355 | 0.757 | 0.497 |
| Rectal (n = 2) | – | ||||||
| Colorectal (n = 8) | 1.22 (1.09–1.40) | 0.001 | 12.94 | 45.90 | 0.074 | 0.089 | 0.322 |
| Total HCAs | |||||||
| Case-Control (n = 3) | |||||||
| Colorectal (n = 3) | 1.16 (0.99–1.35) | 0.063 | 0.70 | 0.00 | 0.706 | 0.607 | 0.602 |
| Combined Risk Estimate | Test of Heterogeneity | Publication Bias | |||||
|---|---|---|---|---|---|---|---|
| Mutagen | Value (95% CI) | p | Q | I2 % | p | p (Egger Test) | p (Begg Test) |
| PhIP | |||||||
| Case-Control (n = 21) 2 | 1.03 (0.95–1.13) | 0.445 | 29.37 | 31.91 | 0.081 | 0.312 | 0.506 |
| Cohort (n = 5) | 1.00 (0.93–1.07) | 0.893 | 1.42 | 0.00 | 0.841 | 0.973 | 1.000 |
| Polled (n = 26) | 1.02 (0.96–1.08) | 0.605 | 31.06 | 19.52 | 0.187 | 0.251 | 0.209 |
| Colon (n = 11) | 1.05 (0.95–1.16) | 0.348 | 14.19 | 29.54 | 0.164 | 0.877 | 0.938 |
| Rectal (n = 5) | 0.94 (0.76–1.17) | 0.593 | 6.68 | 40.13 | 0.154 | 0.719 | 0.624 |
| Colorectal (n = 10) | 1.00 (0.93–1.07) | 0.951 | 8.25 | 0.00 | 0.509 | 0.055 | 0.128 |
| MeIQx | |||||||
| Case-Control (n = 22) | 1.16 (1.02–1.32) | 0.024 | 60.49 | 65.28 | 0.0001 | 0.249 | 0.284 |
| Cohort (n = 5) | 1.14 (1.04–1.24) | 0.003 | 5.50 | 27.24 | 0.240 | 0.223 | 0.327 |
| Polled (n = 27) | 1.14 (1.04–1.25) | 0.004 | 66.09 | 60.66 | 0.0001 | 0.471 | 0.428 |
| Colon (n = 11) | 1.18 (1.03–1.36) | 0.017 | 24.96 | 59.93 | 0.005 | 0.395 | 0.186 |
| Rectal (n = 5) | 1.05 (0.80–1.38) | 0.732 | 9.39 | 57.42 | 0.052 | 0.363 | 0.624 |
| Colorectal (n = 11) | 1.13 (0.99–1.29) | 0.062 | 25.98 | 61.51 | 0.004 | 0.243 | 0.186 |
| DiMeIQx | |||||||
| Case-Control (n = 19) | 1.15 (1.01–1.30) | 0.036 | 53.82 | 66.56 | 0.0001 | 0.267 | 0.600 |
| Cohort (n = 5) | 1.07 (0.95–1.21) | 0.255 | 13.56 | 70.50 | 0.009 | 0.165 | 0.327 |
| Polled (n = 24) | 1.12 (1.02–1.22) | 0.014 | 67.40 | 65.88 | 0.0001 | 0.682 | 0.457 |
| Colon (n = 11) | 1.23 (1.07–1.41) | 0.003 | 25.06 | 60.09 | 0.005 | 0.964 | 0.815 |
| Rectal (n = 5) | 1.06 (0.78–1.45) | 0.693 | 13.67 | 70.74 | 0.008 | 0.534 | 0.624 |
| Colorectal (n = 8) | 1.04 (0.93–1.16) | 0.534 | 17.20 | 59.31 | 0.016 | 0.726 | 0.805 |
| B(a)P | |||||||
| Case-Control (n = 9) | 0.99 (0.89–1.09) | 0.794 | 7.35 | 0.00 | 0.500 | 0.408 | 0.297 |
| Cohort (n = 3) | 0.96 (0.88–1.04) | 0.334 | 0.01 | 0.00 | 0.997 | 0.206 | 0.117 |
| Polled (n = 12) | 0.97 (0.91–1.04) | 0.365 | 7.53 | 0.00 | 0.755 | 0.369 | 0.217 |
| Colon (n = 7) | 0.97 (0.89–1.06) | 0.530 | 5.63 | 0.00 | 0.465 | 0.698 | 0.881 |
| Rectal (n = 2) | 1.04 (0.80–1.33) | 0.791 | 1.35 | 25.93 | 0.245 | – | – |
| Colorectal (n = 3) | 0.96 (0.86–1.06) | 0.413 | 0.28 | 0.00 | 0.868 | 0.989 | 0.602 |
| MDM | |||||||
| Case-Control (n = 7) | 1.13 (1.04–1.22) | 0.004 | 2.59 | 0.00 | 0.858 | 0.536 | 0.652 |
| Cohort (n = 4) | 1.12 (1.03–1.21) | 0.005 | 2.3 | 0.00 | 0.527 | 0.214 | 0.497 |
| Polled (n = 11) | 1.12 (1.06–1.19) | 0.0001 | 4.83 | 0.00 | 0.902 | 0.997 | 0.697 |
| Colon (n = 4) | 1.18 (1.06–1.31) | 0.002 | 1.76 | 0.00 | 0.624 | 0.886 | 0.497 |
| Rectal (n = 3) | 1.09 (0.93–1.29) | 0.293 | 0.79 | 0.00 | 0.673 | 0.489 | 0.117 |
| Colorectal (n = 4) | 1.10 (1.02–1.18) | 0.011 | 0.92 | 0.00 | 0.821 | 0.329 | 0.497 |
| Total HCAs | |||||||
| Case-Control (n = 10) | 0.92 (0.82–1.05) | 0.213 | 12.64 | 28.78 | 0.180 | 0.740 | 0.788 |
| Cohort (n = 1) | – | ||||||
| Polled (n = 11) | 0.92 (0.83–1.02) | 0.096 | 12.70 | 21.29 | 0.241 | 0.687 | 0.697 |
| Colon (n = 3) | 0.87 (0.64–1.19) | 0.391 | 4.09 | 51.12 | 0.129 | 0.572 | 0.602 |
| Rectal (n = 3) | 0.97 (0.58–1.60) | 0.893 | 6.58 | 69.62 | 0.037 | 0.530 | 0.117 |
| Colorectal (n = 5) | 0.93 (0.84–1.03) | 0.150 | 1.43 | 0.00 | 0.840 | 0.249 | 0.142 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiavarini, M.; Bertarelli, G.; Minelli, L.; Fabiani, R. Dietary Intake of Meat Cooking-Related Mutagens (HCAs) and Risk of Colorectal Adenoma and Cancer: A Systematic Review and Meta-Analysis. Nutrients 2017, 9, 514. https://doi.org/10.3390/nu9050514
Chiavarini M, Bertarelli G, Minelli L, Fabiani R. Dietary Intake of Meat Cooking-Related Mutagens (HCAs) and Risk of Colorectal Adenoma and Cancer: A Systematic Review and Meta-Analysis. Nutrients. 2017; 9(5):514. https://doi.org/10.3390/nu9050514
Chicago/Turabian StyleChiavarini, Manuela, Gaia Bertarelli, Liliana Minelli, and Roberto Fabiani. 2017. "Dietary Intake of Meat Cooking-Related Mutagens (HCAs) and Risk of Colorectal Adenoma and Cancer: A Systematic Review and Meta-Analysis" Nutrients 9, no. 5: 514. https://doi.org/10.3390/nu9050514
APA StyleChiavarini, M., Bertarelli, G., Minelli, L., & Fabiani, R. (2017). Dietary Intake of Meat Cooking-Related Mutagens (HCAs) and Risk of Colorectal Adenoma and Cancer: A Systematic Review and Meta-Analysis. Nutrients, 9(5), 514. https://doi.org/10.3390/nu9050514