1?,25-dihydroxyvitamin D3 Attenuates TGF-?-Induced Pro-Fibrotic Effects in Human Lung Epithelial Cells through Inhibition of Epithelial–Mesenchymal Transition


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Treatment
2.2. Western Blot
2.3. Immunofluorescence Staining
2.4. Scratch Wound Healing Assay
2.5. Invasion Assay
2.6. RNA Isolation and Real-Time Reverse Transcription Quantitative PCR
2.7. Statistical Analysis
3. Results
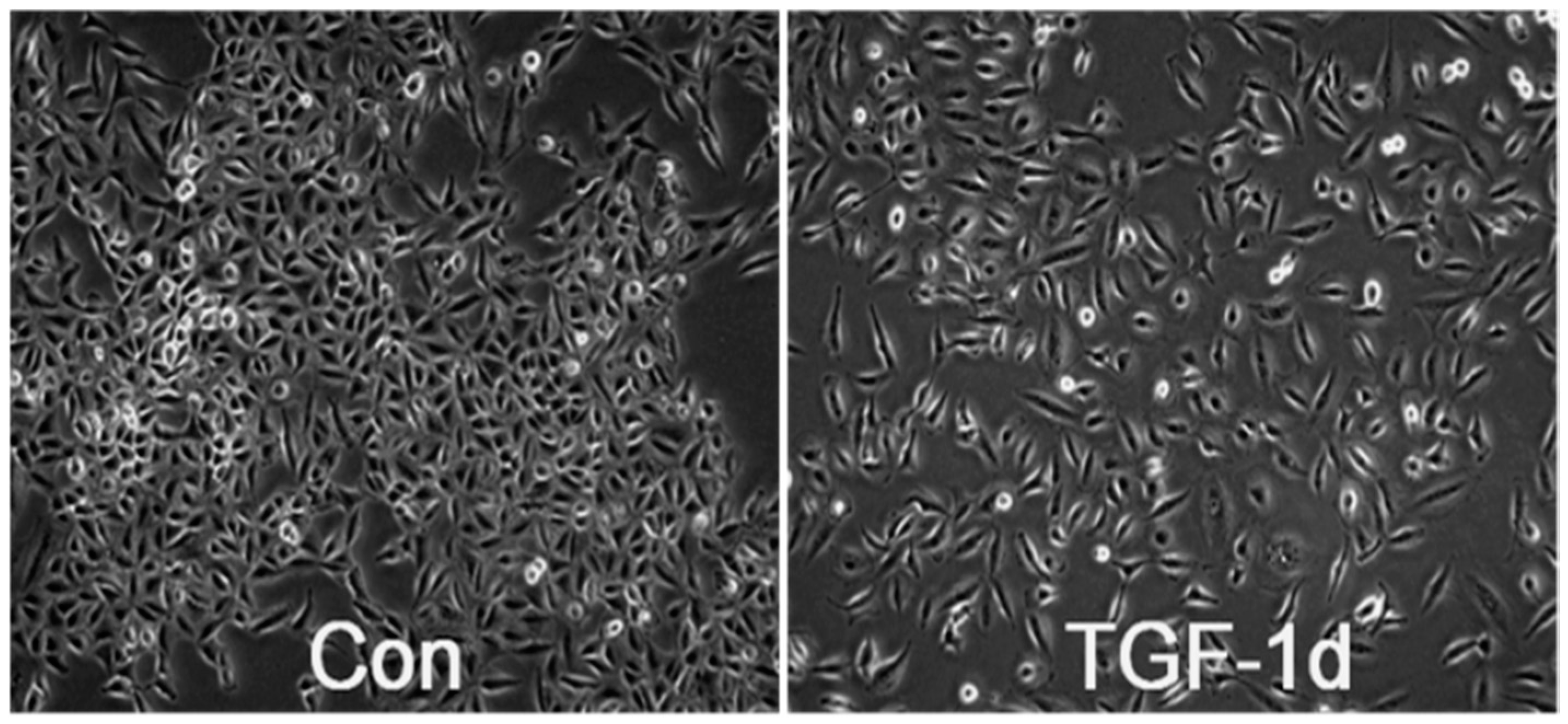
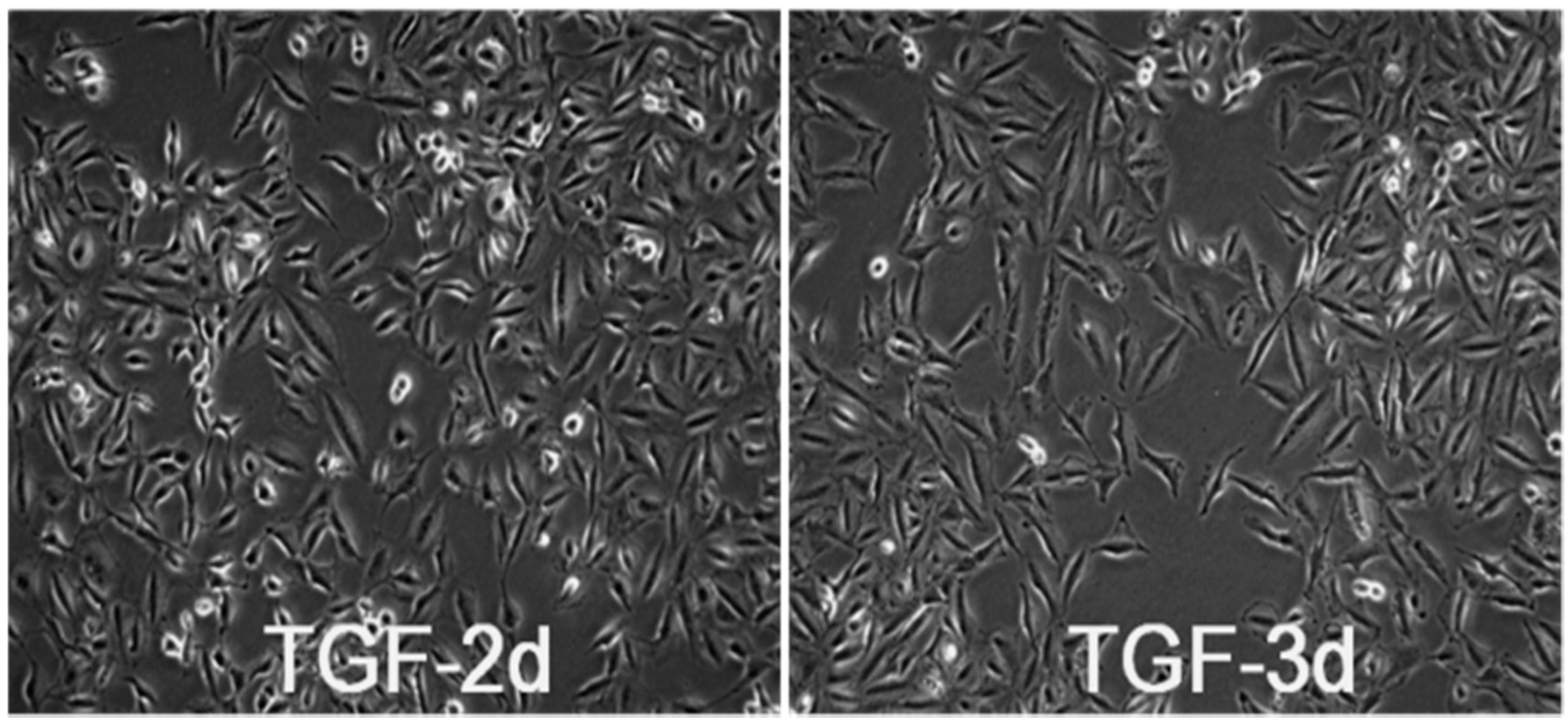
3.1. TGF-β Induces Morphological Alteration in A549 Cells
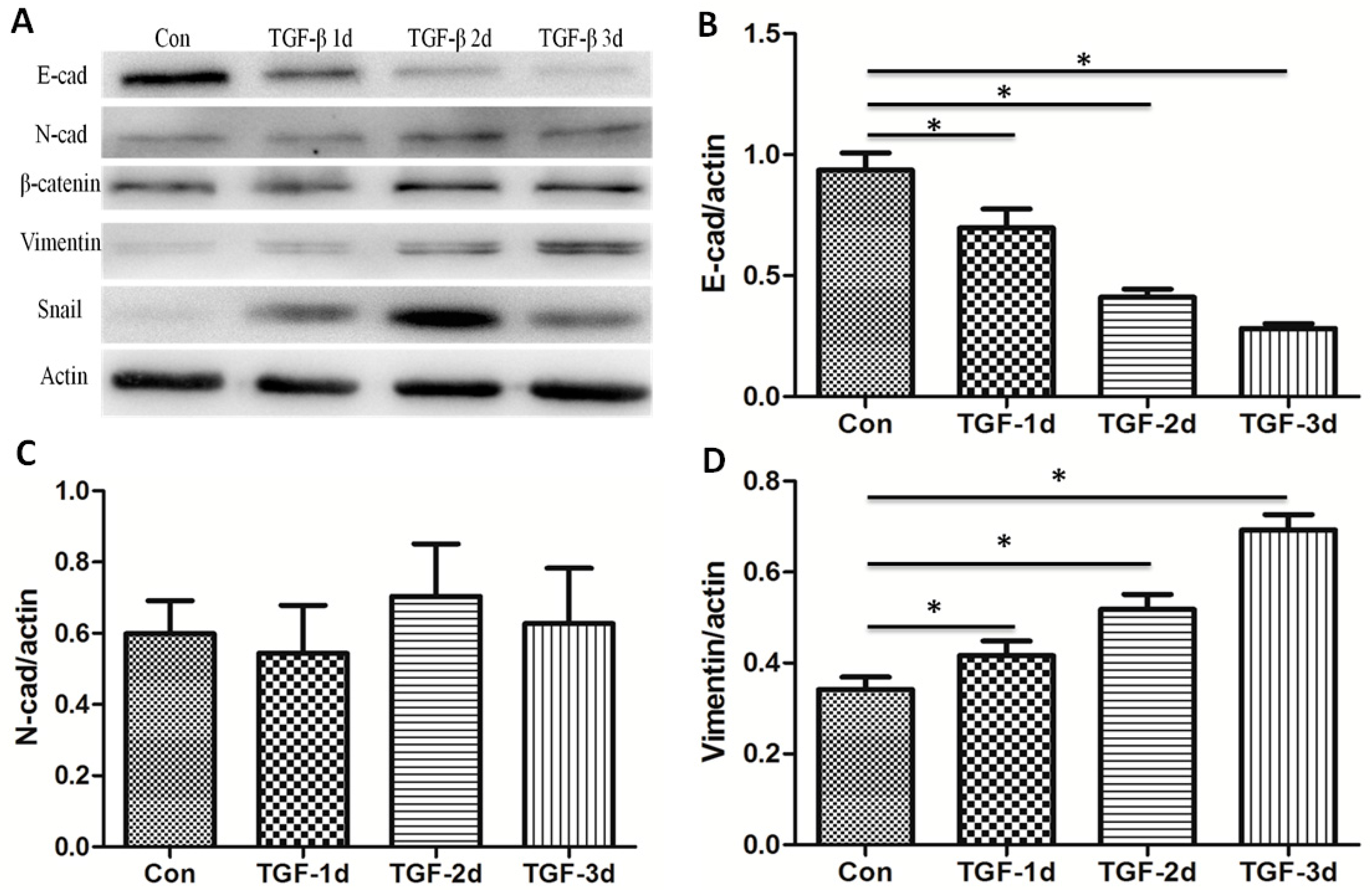
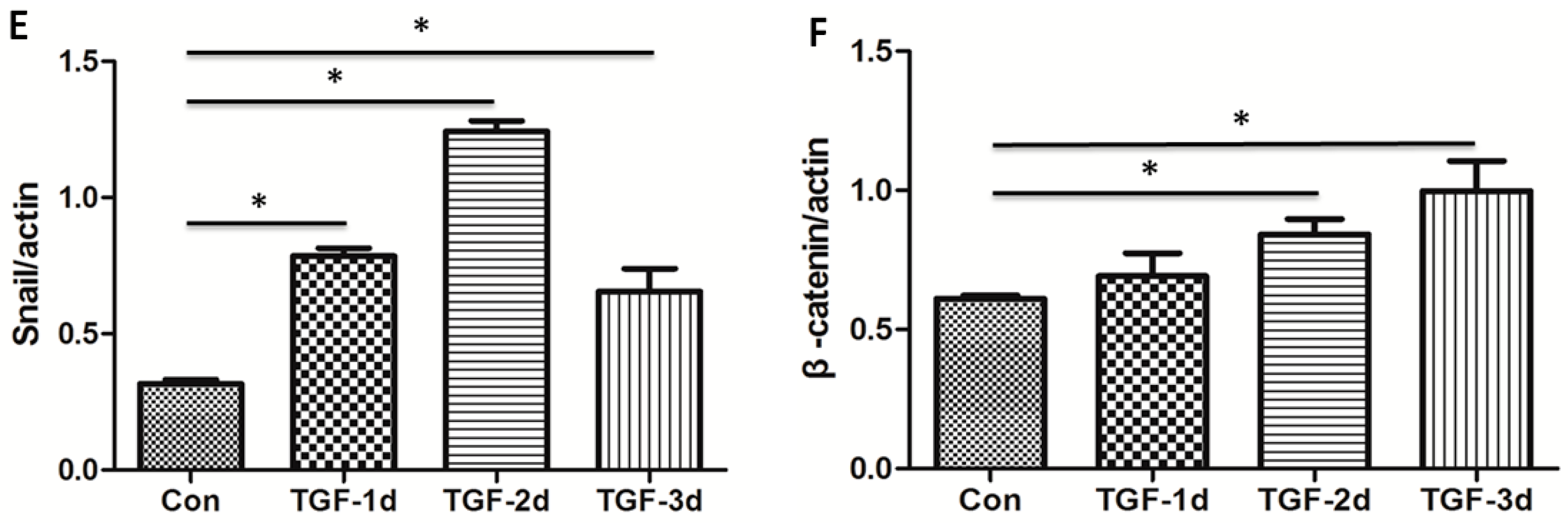
3.2. TGF-β Induces the Alteration of EMT Markers in A549 Cells
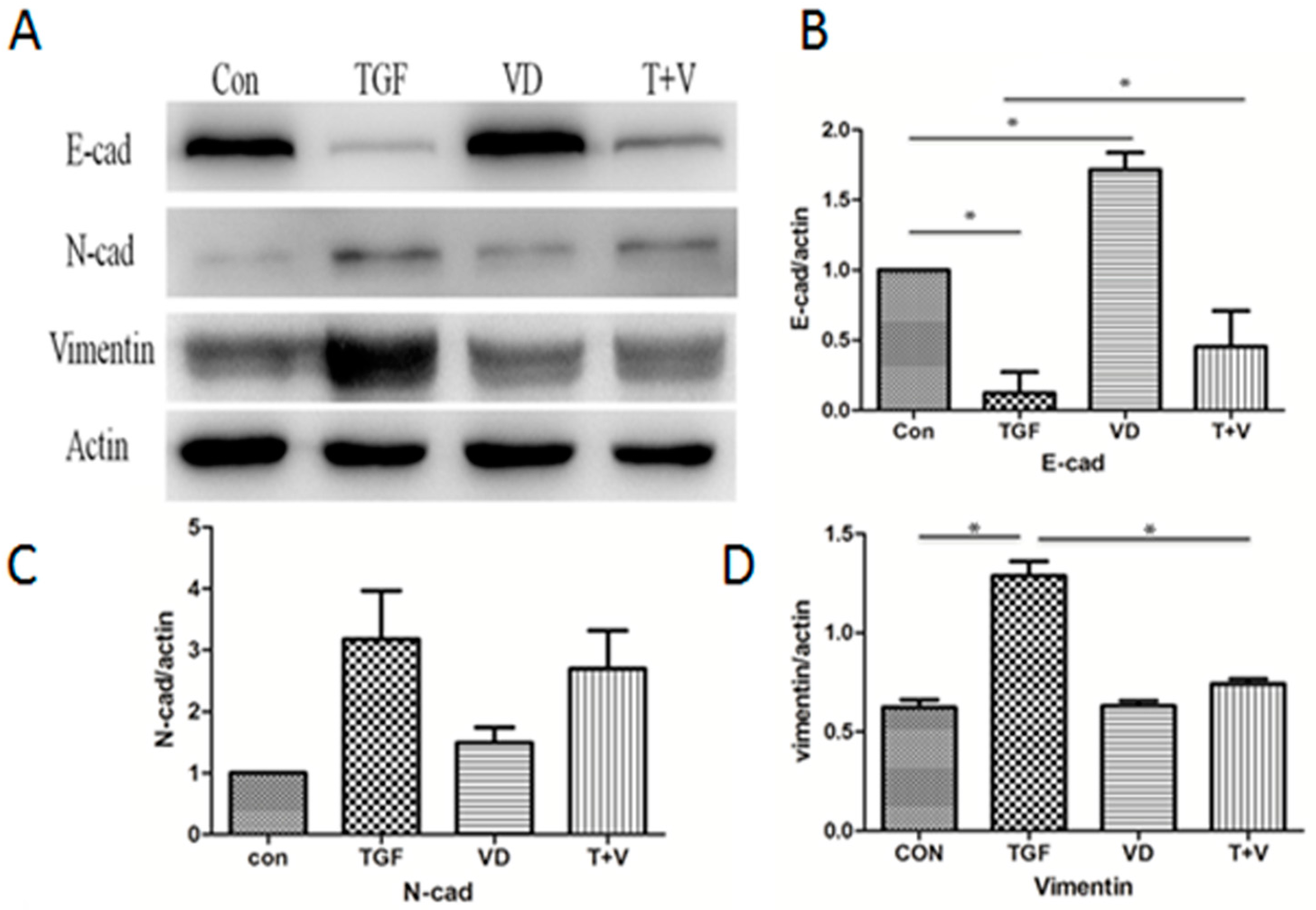
3.3. 1α,25(OH)2D3 Opposes the Expression of EMT Markers and Extracellular Matrix Components Induced by TGF-β
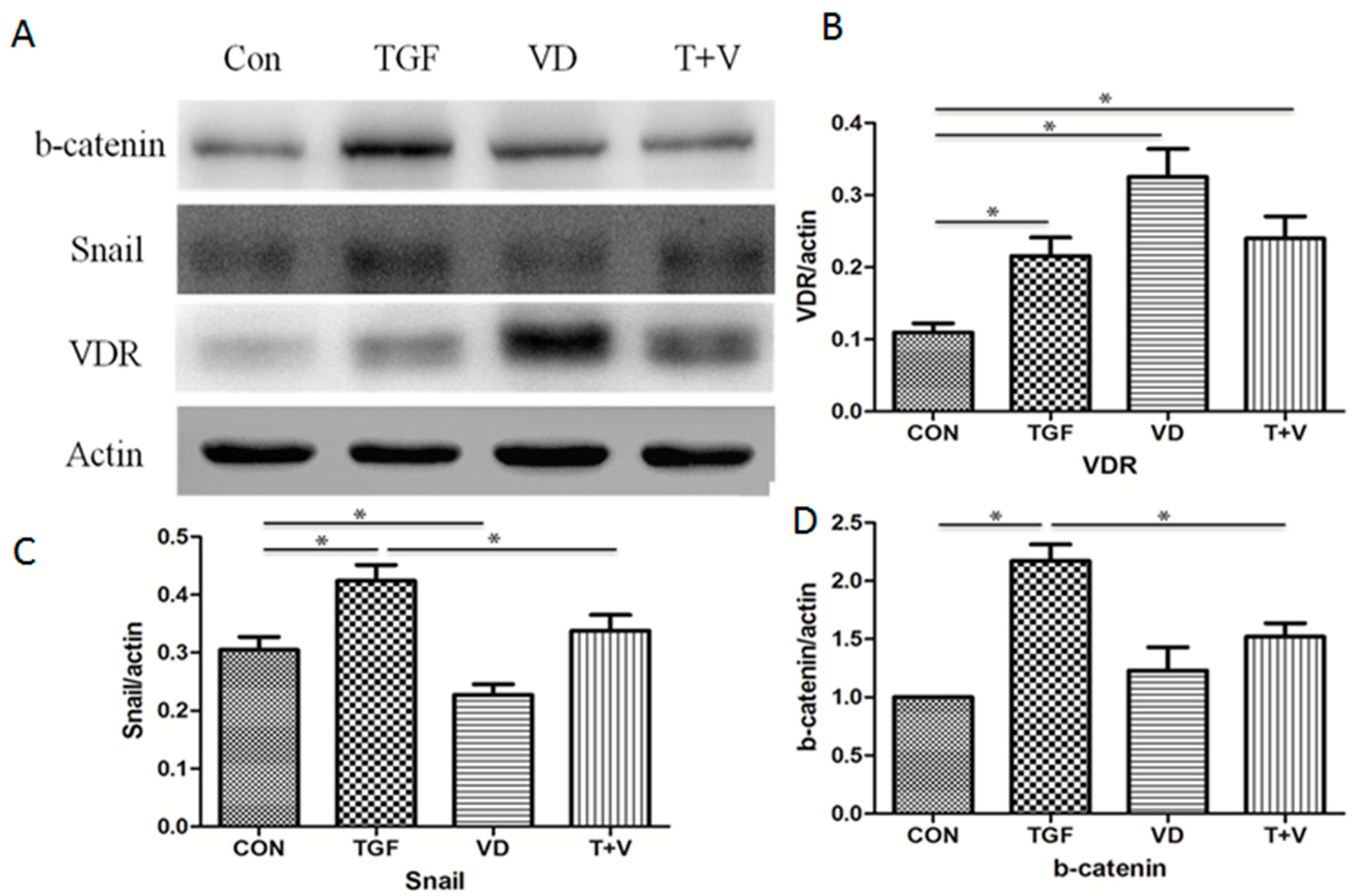
3.4. 1α,25(OH)2D3 Represses EMT-Related Transcription Factors by TGF-β and Increases VDR Expression
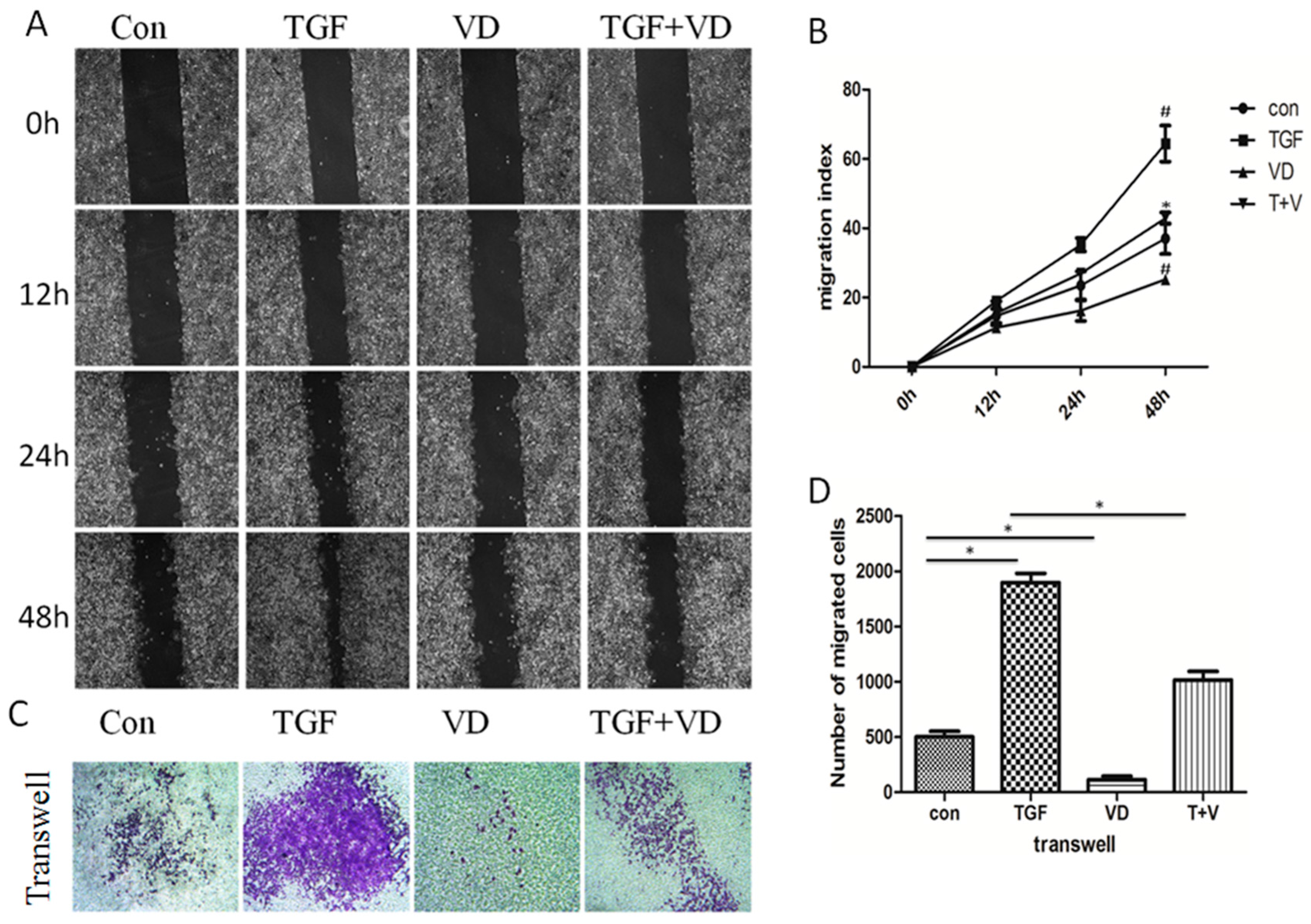
3.5. 1α,25(OH)2D3 Prevents TGF-β-Induced Invasion and Metastasis in A549 Cells
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wynn, T.A. Integrating mechanisms of pulmonary fibrosis. J. Exp. Med. 2011, 208, 1339–1350. [Google Scholar] [CrossRef] [PubMed]
- Camelo, A.; Dunmore, R.; Sleeman, M.A.; Clarke, D.L. The epithelium in idiopathic pulmonary fibrosis: Breaking the barrier. Front. Pharmacol. 2014, 4, 173. [Google Scholar] [CrossRef] [PubMed]
- Kalluri, R.; Neilson, E.G. Epithelial-mesenchymal transition and its implications for fibrosis. J. Clin. Investig. 2003, 112, 1776–1784. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.K.; Kugler, M.C.; Wolters, P.J.; Robillard, L.; Galvez, M.G.; Brumwell, A.N.; Sheppard, D.; Chapman, H.A. Alveolar epithelial cell mesenchymal transition develops in vivo during pulmonary fibrosis and is regulated by the extracellular matrix. Proc. Natl. Acad. Sci. USA 2006, 103, 13180–13185. [Google Scholar] [CrossRef] [PubMed]
- Zeisberg, E.M.; Tarnavski, O.; Zeisberg, M.; Dorfman, A.L.; McMullen, J.R.; Gustafsson, E.; Chandraker, A.; Yuan, X.; Pu, W.T.; Roberts, A.B.; et al. Endothelial-to-mesenchymal transition contributes to cardiac fibrosis. Nat. Med. 2007, 13, 952–961. [Google Scholar] [CrossRef] [PubMed]
- Zeisberg, M.; Yang, C.; Martino, M.; Duncan, M.B.; Rieder, F.; Tanjore, H.; Kalluri, R. Fibroblasts derive from hepatocytes in liver fibrosis via epithelial to mesenchymal transition. J. Biol. Chem. 2007, 282, 23337–23347. [Google Scholar] [CrossRef] [PubMed]
- Potenta, S.; Zeisberg, E.; Kalluri, R. The role of endothelial-to-mesenchymal transition in cancer progression. Br. J. Cancer 2008, 99, 1375–1379. [Google Scholar] [CrossRef] [PubMed]
- Moustakas, A.; Heldin, P. TGF-beta and matrix-regulated epithelial to mesenchymal transition. Biochim. Biophys. Acta 2014, 1840, 2621–2634. [Google Scholar] [CrossRef] [PubMed]
- Cantelli, G.; Crosas-Molist, E.; Georgouli, M.; Sanz-Moreno, V. TGF-beta-induced transcription in cancer. Semin. Cancer Biol. 2017, 42, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Akhurst, R.J. Targeting TGF-beta signaling for therapeutic gain. Cold Spring Harb. Perspect. Biol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Coker, R.K.; Laurent, G.J. Anticytokine approaches in pulmonary fibrosis: Bringing factors into focus. Thorax 1997, 52, 294–296. [Google Scholar] [CrossRef] [PubMed]
- Coker, R.K.; Laurent, G.J.; Jeffery, P.K.; du Bois, R.M.; Black, C.M.; McAnulty, R.J. Localisation of transforming growth factor beta1 and beta3 mRNA transcripts in normal and fibrotic human lung. Thorax 2001, 56, 549–556. [Google Scholar] [CrossRef] [PubMed]
- Dusso, A.S.; Brown, A.J. Mechanism of vitamin D action and its regulation. Am. J. Kidney Dis. Off. J. Natl. Kidney Found. 1998, 32, S13–S24. [Google Scholar] [CrossRef]
- Brown, A.J. Vitamin D analogues. Am. J. Kidney Dis. Off. J. Natl. Kidney Found. 1998, 32, S25–S39. [Google Scholar] [CrossRef]
- Melamed, M.L.; Michos, E.D.; Post, W.; Astor, B. 25-hydroxy vitamin D levels and the risk of mortality in the general population. Arch. Intern. Med. 2008, 168, 1629–1637. [Google Scholar] [CrossRef] [PubMed]
- Petta, S.; Camma, C.; Scazzone, C.; Tripodo, C.; Di Marco, V.; Bono, A.; Cabibi, D.; Licata, G.; Porcasi, R.; Marchesini, G.; et al. Low vitamin D serum level is related to severe fibrosis and low responsiveness to interferon-based therapy in genotype 1 chronic hepatitis C. Hepatology 2010, 51, 1158–1167. [Google Scholar] [CrossRef] [PubMed]
- Arteh, J.; Narra, S.; Nair, S. Prevalence of vitamin D deficiency in chronic liver disease. Dig. Dis. Sci. 2010, 55, 2624–2628. [Google Scholar] [CrossRef] [PubMed]
- Abramovitch, S.; Dahan-Bachar, L.; Sharvit, E.; Weisman, Y.; Ben Tov, A.; Brazowski, E.; Reif, S. Vitamin D inhibits proliferation and profibrotic marker expression in hepatic stellate cells and decreases thioacetamide-induced liver fibrosis in rats. Gut 2011, 60, 1728–1737. [Google Scholar] [CrossRef] [PubMed]
- Tan, X.; Li, Y.; Liu, Y. Therapeutic role and potential mechanisms of active vitamin D in renal interstitial fibrosis. J. Steroid Biochem. Mol. Biol. 2007, 103, 491–496. [Google Scholar] [CrossRef] [PubMed]
- Artaza, J.N.; Norris, K.C. Vitamin D reduces the expression of collagen and key profibrotic factors by inducing an antifibrotic phenotype in mesenchymal multipotent cells. J. Endocrinol. 2009, 200, 207–221. [Google Scholar] [CrossRef] [PubMed]
- Fischer, K.D.; Agrawal, D.K. Vitamin D regulating TGF-beta induced epithelial-mesenchymal transition. Respir. Res. 2014, 15, 146. [Google Scholar] [CrossRef] [PubMed]
- Fischer, K.D.; Agrawal, D.K. Erratum to: Vitamin D regulating TGF-β induced epithelial-mesenchymal transition. Respir. Res. 2015, 16, 139. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Yuan, T.; Du, G.; Zhao, Q.; Ma, L.; Zhu, J. The impact of 1,25-dihydroxyvitamin D3 on the expression of connective tissue growth factor and transforming growth factor-beta1 in the myocardium of rats with diabetes. Diabetes Res. Clin. Pract. 2014, 104, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Ren, Z.; Li, W.; Zhao, Q.; Ma, L.; Zhu, J. The impact of 1,25-dihydroxyvitamin D3 on the expressions of vascular endothelial growth factor and transforming growth factor-beta(1) in the retinas of rats with diabetes. Diabetes Res. Clin. Pract. 2012, 98, 474–480. [Google Scholar] [CrossRef] [PubMed]
- Kabel, A.M.; Abd Elmaaboud, M.A.; Atef, A.; Baali, M.H. Ameliorative potential of linagliptin and/or calcipotriol on bleomycin-induced lung fibrosis: In vivo and in vitro study. Environ. Toxicol. Pharmacol. 2017, 50, 216–226. [Google Scholar] [CrossRef] [PubMed]
- Foong, R.E.; Shaw, N.C.; Berry, L.J.; Hart, P.H.; Gorman, S.; Zosky, G.R. Vitamin D deficiency causes airway hyperresponsiveness, increases airway smooth muscle mass, and reduces TGF-beta expression in the lungs of female BALB/c mice. Physiol. Rep. 2014, 2, e00276. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.Y.; Zhou, X.J.; Li, X.; Li, Z.; Hong, J.G. Effect of 1,25-(OH)2D3 supplementation during gestation and lactation on TGF-beta1 and Smad3 expression in lungs of rat offspring with asthma. Chin. J. Contemp. Pediatr. 2012, 14, 366–370. [Google Scholar]
- Taher, Y.A.; van Esch, B.C.; Hofman, G.A.; Henricks, P.A.; van Oosterhout, A.J. 1alpha,25-dihydroxyvitamin D3 potentiates the beneficial effects of allergen immunotherapy in a mouse model of allergic asthma: Role for IL-10 and TGF-beta. J. Immunol. 2008, 180, 5211–5221. [Google Scholar] [CrossRef] [PubMed]
- Weinreich, T.; Landolt, M.; Booy, C.; Wuthrich, R.; Binswanger, U. 1,25-dihydroxyvitamin D3 stimulates transforming growth factor-beta1 synthesis by mouse renal proximal tubular cells. Kidney Blood Press. Res. 1999, 22, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Boyan, B.D.; Schwartz, Z.; Park-Snyder, S.; Dean, D.D.; Yang, F.; Twardzik, D.; Bonewald, L.F. Latent transforming growth factor-beta is produced by chondrocytes and activated by extracellular matrix vesicles upon exposure to 1,25-(OH)2D3. J. Biol. Chem. 1994, 269, 28374–28381. [Google Scholar] [PubMed]
- Ding, N.; Yu, R.T.; Subramaniam, N.; Sherman, M.H.; Wilson, C.; Rao, R.; Leblanc, M.; Coulter, S.; He, M.; Scott, C.; et al. A vitamin D receptor/SMAD genomic circuit gates hepatic fibrotic response. Cell 2013, 153, 601–613. [Google Scholar] [CrossRef] [PubMed]
- Larriba, M.J.; Gonzalez-Sancho, J.M.; Bonilla, F.; Munoz, A. Interaction of vitamin D with membrane-based signaling pathways. Front. Physiol. 2014, 5, 60. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, A.M.; Wongtrakool, C.; Welch, T.; Steinmeyer, A.; Zugel, U.; Roman, J. Vitamin D inhibition of pro-fibrotic effects of transforming growth factor beta1 in lung fibroblasts and epithelial cells. J. Steroid Biochem. Mol. Biol. 2010, 118, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Lamouille, S.; Xu, J.; Derynck, R. Molecular mechanisms of epithelial-mesenchymal transition. Nat. Rev. Mol. Cell Biol. 2014, 15, 178–196. [Google Scholar] [CrossRef] [PubMed]
- Donovan, D.S., Jr.; Papadopoulos, A.; Staron, R.B.; Addesso, V.; Schulman, L.; McGregor, C.; Cosman, F.; Lindsay, R.L.; Shane, E. Bone mass and vitamin D deficiency in adults with advanced cystic fibrosis lung disease. Am. J. Respir. Crit. Care Med. 1998, 157, 1892–1899. [Google Scholar] [CrossRef] [PubMed]
- Ito, I.; Waku, T.; Aoki, M.; Abe, R.; Nagai, Y.; Watanabe, T.; Nakajima, Y.; Ohkido, I.; Yokoyama, K.; Miyachi, H.; et al. A nonclassical vitamin D receptor pathway suppresses renal fibrosis. J. Clin. Investig. 2013, 123, 4579–4594. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Spataro, B.C.; Yang, J.; Dai, C.; Liu, Y. 1,25-dihydroxyvitamin D inhibits renal interstitial myofibroblast activation by inducing hepatocyte growth factor expression. Kidney Int. 2005, 68, 1500–1510. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Yu, X.; Fang, X.; Liang, A.; Yu, Z.; Gu, P.; Zeng, Y.; He, J.; Zhu, H.; Li, S.; et al. Preventive effects of vitamin D treatment on bleomycin-induced pulmonary fibrosis. Sci. Rep. 2015, 5, 17638. [Google Scholar] [CrossRef] [PubMed]
- Tan, Z.X.; Chen, Y.H.; Xu, S.; Qin, H.Y.; Zhang, C.; Zhao, H.; Xu, D.X. Calcitriol inhibits bleomycin-induced early pulmonary inflammatory response and epithelial-mesenchymal transition in mice. Toxicol. Lett. 2016, 240, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Batlle, E.; Sancho, E.; Franci, C.; Dominguez, D.; Monfar, M.; Baulida, J.; Garcia De Herreros, A. The transcription factor snail is a repressor of E-cadherin gene expression in epithelial tumour cells. Nat. Cell Biol. 2000, 2, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Boutet, A.; De Frutos, C.A.; Maxwell, P.H.; Mayol, M.J.; Romero, J.; Nieto, M.A. Snail activation disrupts tissue homeostasis and induces fibrosis in the adult kidney. EMBO J. 2006, 25, 5603–5613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menezes, R.J.; Cheney, R.T.; Husain, A.; Tretiakova, M.; Loewen, G.; Johnson, C.S.; Jayaprakash, V.; Moysich, K.B.; Salgia, R.; Reid, M.E. Vitamin D receptor expression in normal, premalignant, and malignant human lung tissue. Cancer Epidemiol. Biomark. Prev. 2008, 17, 1104–1110. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.L.; Zhang, H.M.; Hu, Z.Y.; Wang, P.; Wan, J.M.; Li, B.Y. Synergy of 1,25-dihydroxyvitamin D3 and carboplatin in growth suppression of SKOV-3 cells. Oncol. Lett. 2014, 8, 1348–1354. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Forward Sequence (5′-3′) | Reverse Sequence (5′-3′) |
|---|---|---|
| Collagen I | ACGTCCTGGTGAAGTTGGTC | ACCAGGGAAGCCTCTCTCTC |
| Fibronectin | GAGCTATTCCCTGCACCTGA | CGTGCAAGGCAACCACACT |
| GAPDH | CGTGCAAGGCAACCACACT | TGGCAGGTTTTTCTAGACGG |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, F.; Yang, Y.; Xue, L.; Li, B.; Zhang, Z. 1?,25-dihydroxyvitamin D3 Attenuates TGF-?-Induced Pro-Fibrotic Effects in Human Lung Epithelial Cells through Inhibition of Epithelial–Mesenchymal Transition. Nutrients 2017, 9, 980. https://doi.org/10.3390/nu9090980
Jiang F, Yang Y, Xue L, Li B, Zhang Z. 1?,25-dihydroxyvitamin D3 Attenuates TGF-?-Induced Pro-Fibrotic Effects in Human Lung Epithelial Cells through Inhibition of Epithelial–Mesenchymal Transition. Nutrients. 2017; 9(9):980. https://doi.org/10.3390/nu9090980
Chicago/Turabian StyleJiang, Fei, Yong Yang, Lian Xue, Bingyan Li, and Zengli Zhang. 2017. "1?,25-dihydroxyvitamin D3 Attenuates TGF-?-Induced Pro-Fibrotic Effects in Human Lung Epithelial Cells through Inhibition of Epithelial–Mesenchymal Transition" Nutrients 9, no. 9: 980. https://doi.org/10.3390/nu9090980
APA StyleJiang, F., Yang, Y., Xue, L., Li, B., & Zhang, Z. (2017). 1?,25-dihydroxyvitamin D3 Attenuates TGF-?-Induced Pro-Fibrotic Effects in Human Lung Epithelial Cells through Inhibition of Epithelial–Mesenchymal Transition. Nutrients, 9(9), 980. https://doi.org/10.3390/nu9090980