Phytotoxic Metabolites Produced by Legume-Associated Ascochyta and Its Related Genera in the Dothideomycetes


Abstract
:1. Introduction
2. Biological Activities and the Modes of Action
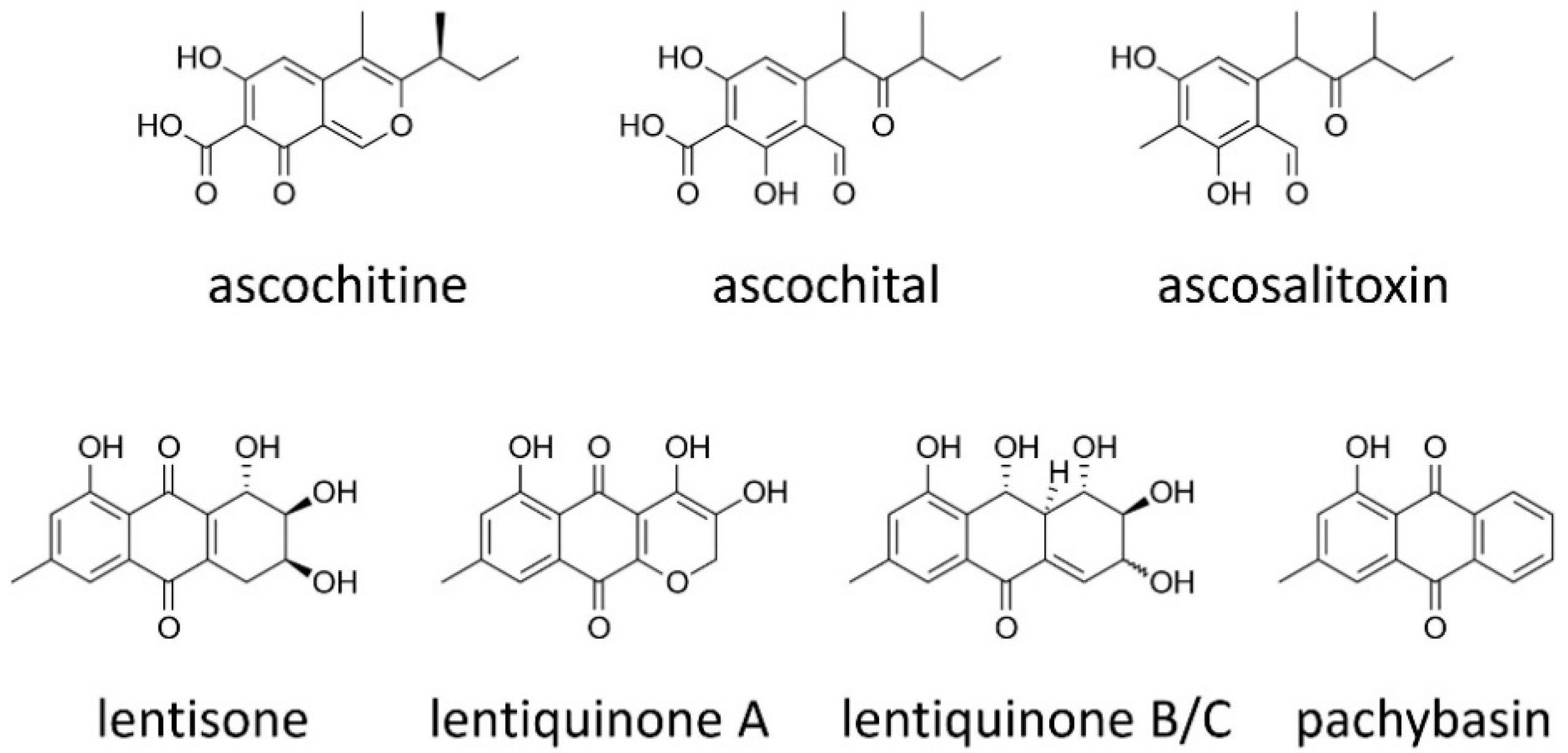
2.1. Ascochitine and Its Derivatives
2.2. Anthraquinones
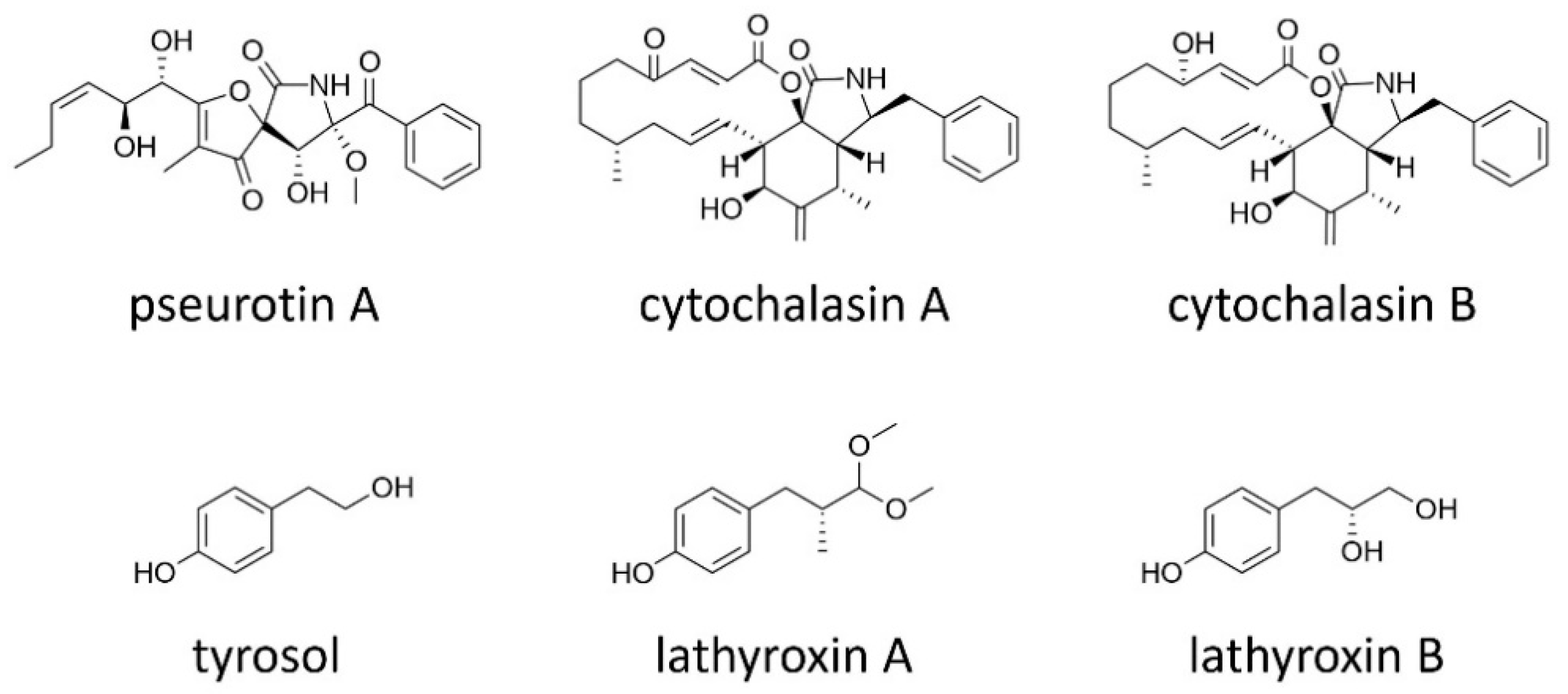
2.3. Macrolides
2.4. Meroterpenoids
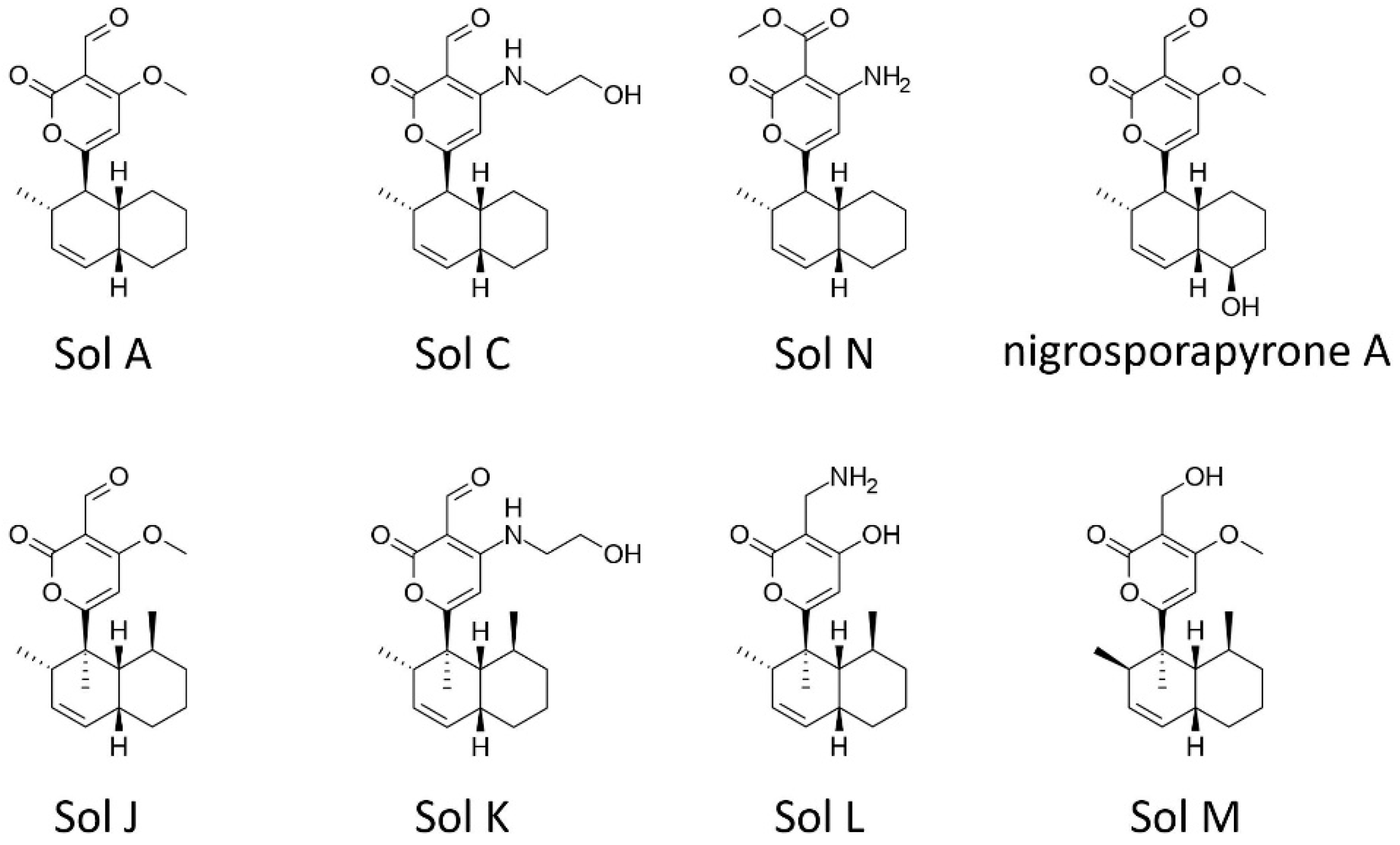
2.5. Solanapyrones
2.6. Other Secondary Metabolites
3. Biosynthesis Gene Cluster and Ecological Roles
3.1. Solanapyrone Biosynthesis Gene Cluster
3.2. Ascochitine Biosynthesis Gene Cluster
3.3. Ascofuranone Biosynthesis Gene Cluster
3.4. Identification of Other SM Gene Clustesrs in the Legume-Associated Ascochyta Species
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Ohm, R.A.; Feau, N.; Henrissat, B.; Schoch, C.L.; Horwitz, B.A.; Barry, K.W.; Condon, B.J.; Copeland, A.C.; Dhillon, B.; Glaser, F.; et al. Diverse lifestyles and strategies of plant pathogenesis encoded in the genomes of eighteen Dothideomycetes fungi. PLoS Pathog. 2012, 8, e1003037. [Google Scholar] [CrossRef] [PubMed]
- Solomon, P.S. Assessing the mycotoxigenic threat of necrotrophic pathogens of wheat. Mycotoxin Res. 2011, 27, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Boerema, G.H.; Bollen, G.J. Conidiogenesis and conidial septation as differentiating criteria between Phoma and Ascochyta. Persoonia 1975, 8, 111–114. [Google Scholar]
- Aveskamp, M.M.; de Gruyter, J.; Woudenberg, J.H.C.; Verkley, G.J.M.; Crous, P.W. Highlights of the Didymellaceae: A polyphasic approach to characterise Phoma and related pleosporalean genera. Stud. Mycol. 2010, 65, 1–60. [Google Scholar] [CrossRef] [PubMed]
- Rubiales, D.; Fondevilla, S. Future prospects for Ascochyta blight resistance breeding in cool season food legumes. Front. Plant Sci. 2012, 3, 27. [Google Scholar] [CrossRef]
- Nene, Y.L. A review of Ascochyta blight of chickpea. Trop. Pest Manag. 1982, 28, 61–70. [Google Scholar] [CrossRef]
- Höhl, B.; Pfautsch, M.; Barz, W. Histology of disease development in resistant and susceptible cultivars of chickpea (Cicer arietinum L.) inoculated with spores of Ascochyta rabiei. J. Phytopathol. 1990, 129, 31–45. [Google Scholar] [CrossRef]
- Kaiser, W.J. Inter- and intranational spread of ascochyta pathogens of chickpea, faba bean, and lentil. Can. J. Plant Pathol. 1997, 19, 215–224. [Google Scholar] [CrossRef]
- Peever, T.L.; Barve, M.P.; Stone, L.J. Evolutionary relationships among Ascochyta species infecting wild and cultivated hosts in the legume tribes Cicereae and Vicieae. Mycologia 2007, 99, 59–77. [Google Scholar] [CrossRef]
- Hernandez-Bello, M.A.; Chilvers, M.I.; Akamatsu, H.; Peever, T.L. Host specificity of Ascochyta spp. infecting legumes of the Viciae and Cicerae tribes and pathogenicity of an interspecific hybrid. Phytopathology 2006, 96, 1148–1156. [Google Scholar] [CrossRef]
- Kaiser, W.J.; Wang, B.-C.; Rogers, J.D. Ascochyta fabae and A. lentis: Host specificity, teleomorphs (Didymella), hybrid analysis, and taxonomic status. Plant Dis. 1997, 81, 809–816. [Google Scholar] [CrossRef] [PubMed]
- Infantino, A.; Zaccardelli, M.; Costa, C.; Ozkilinc, H.; Habibi, A.; Peever, T. A new disease of grasspea (Lathyrus sativus) caused by Ascochyta lentis var. lathyri. J. Plant Pathol. 2016, 98, 541–548. [Google Scholar]
- Chilvers, M.I.; Rogers, J.D.; Dugan, F.M.; Stewart, J.E.; Chen, W.; Peever, T.L. Didymella pisi sp. nov., the teleomorph of Ascochyta pisi. Mycol. Res. 2009, 113, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Barilli, E.; Cobos, M.J.; Rubiales, D. Clarification on host range of Didymella pinodes the causal agent of pea Ascochyta blight. Front. Plant Sci. 2016, 7, 592. [Google Scholar] [CrossRef] [PubMed]
- Djebali, N. Aggressiveness and host range of Phoma medicaginis isolated from Medicago species growing in Tunisia. Phytopathol. Mediterr. 2013, 52, 3–15. [Google Scholar]
- Kim, W.; Peever, T.L.; Park, J.-J.; Park, C.-M.; Gang, D.R.; Xian, M.; Davidson, J.A.; Infantino, A.; Kaiser, W.J.; Chen, W. Use of metabolomics for the chemotaxonomy of legume-associated Ascochyta and allied genera. Sci. Rep. 2016, 6, 20192. [Google Scholar] [CrossRef]
- Stergiopoulos, I.; Collemare, J.; Mehrabi, R.; De Wit, P.J.G.M. Phytotoxic secondary metabolites and peptides produced by plant pathogenic Dothideomycete fungi. FEMS Microbiol. Rev. 2013, 37, 67–93. [Google Scholar] [CrossRef]
- Muria-Gonzalez, M.J.; Chooi, Y.-H.; Breen, S.; Solomon, P.S. The past, present and future of secondary metabolite research in the Dothideomycetes: Secondary metabolites in the Dothideomycetes. Mol. Plant Pathol. 2015, 16, 92–107. [Google Scholar] [CrossRef]
- Akimitsu, K.; Tsuge, T.; Kodama, M.; Yamamoto, M.; Otani, H. Alternaria host-selective toxins: Determinant factors of plant disease. J. Gen. Plant Pathol. 2014, 80, 109–122. [Google Scholar] [CrossRef]
- Rai, M.; Deshmukh, P.; Gade, A.; Ingle, A.; Kövics, G.J.; Irinyi, L. Phoma Saccardo: Distribution, secondary metabolite production and biotechnological applications. Crit. Rev. Microbiol. 2009, 35, 182–196. [Google Scholar] [CrossRef]
- Tsuge, T.; Harimoto, Y.; Akimitsu, K.; Ohtani, K.; Kodama, M.; Akagi, Y.; Egusa, M.; Yamamoto, M.; Otani, H. Host-selective toxins produced by the plant pathogenic fungus Alternaria alternata. FEMS Microbiol. Rev. 2013, 37, 44–66. [Google Scholar] [CrossRef] [PubMed]
- Bertini, S. Su di un composto ad antibiotica prodotto da Ascochyta pisi Lib. Annali Sperimentaz Agraria (Roma) 1956, 11, 545–556. [Google Scholar]
- Oku, H.; Nakanishi, T. A toxic metabolite from Ascochyta fabae having antibiotic activity. Phytopathology 1963, 53, 1321–1325. [Google Scholar]
- Smith, G.R.; Munro, M.H.G.; Fineran, B.A.; Cole, A.L.J. Evidence for the involvement of ascochitine in phoma leafspot-wilt disease of Clematis. Physiol. Mol. Plant Pathol. 1994, 45, 333–348. [Google Scholar] [CrossRef]
- Venkatasubbaiah, P.; Chilton, W.S. Phytotoxins of Ascochyta hyalospora, causal agent of lambsquarters leaf spot. J. Nat. Prod. 1992, 55, 461–467. [Google Scholar] [CrossRef]
- Mafezoli, J.; Xu, Y.; Hilário, F.; Freidhof, B.; Espinosa-Artiles, P.; dos Santos, L.C.; de Oliveira, M.C.F.; Gunatilaka, A.A.L. Modulation of polyketide biosynthetic pathway of the endophytic fungus, Anteaglonium sp. FL0768, by copper (II) and anacardic acid. Phytochem. Lett. 2018, 28, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Seibert, S.F.; Eguereva, E.; Krick, A.; Kehraus, S.; Voloshina, E.; Raabe, G.; Fleischhauer, J.; Leistner, E.; Wiese, M.; Prinz, H.; et al. Polyketides from the marine-derived fungus Ascochyta salicorniae and their potential to inhibit protein phosphatases. Org. Biomol. Chem. 2006, 4, 2233–2240. [Google Scholar] [CrossRef] [PubMed]
- Oku, H.; Nakanishi, T. Reductive detoxification of an antibiotic, ascochitine, by an insensitive fungus, Fusarium lycopersici Sacc. Naturwissenschaften 1964, 51, 538. [Google Scholar] [CrossRef]
- Nakanishi, T.; Oku, H. Mechanism of selective toxicity: Absorption and detoxication of an antibiotic, ascochitine, by sensitive and insensitive fungi. Phytopathology 1969, 59, 1563–1565. [Google Scholar]
- Evidente, A.; Capasso, R.; Vurro, M.; Bottalico, A. Ascosalitoxin, a phytotoxic trisubstituted salicylic aldehyde from Ascochyta pisi. Phytochemistry 1993, 34, 995–998. [Google Scholar] [CrossRef]
- Kusnick, C.; Jansen, R.; Liberra, K.; Lindequist, U. Ascochital, a new metabolite from the marine ascomycete Kirschsteiniothelia maritima. Pharmazie 2002, 57, 510–512. [Google Scholar] [PubMed]
- Dewick, P.M. Medicinal Natural Products: A Biosynthetic Approach, 3rd ed.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2009. [Google Scholar]
- Osbourn, A.E.; Lanzotti, V. Plant-Derived Natural Products: Synthesis, Function, and Application; Springer: New York, NY, USA, 2009. [Google Scholar]
- Bick, I.; Rhee, C. Anthraquinone pigments from Phoma foveata Foister. Biochem. J. 1966, 98, 112–116. [Google Scholar] [CrossRef]
- Davies, G.D.; Hodge, P. Chemistry of quinones. Part, V. Structure of cladofulvin, a bianthraquinone from Cladosporium fulvum Cooke. J. Chem. Soc. Perkin Trans. 1974, 1974, 2403–2405. [Google Scholar] [CrossRef]
- Jalal, M.A.F.; Hossain, M.B.; Robeson, D.J.; Van der Helm, D. Cercospora beticola phytotoxins: Cebetins that are photoactive, magnesium ion-binding, chlorinated anthraquinone-xanthone conjugates. J. Am. Chem. Soc. 1992, 114, 5967–5971. [Google Scholar] [CrossRef]
- Bouras, N.; Strelkov, S.E. The anthraquinone catenarin is phytotoxic and produced in leaves and kernels of wheat infected by Pyrenophora tritici-repentis. Physiol. Mol. Plant Pathol. 2008, 72, 87–95. [Google Scholar] [CrossRef]
- Miethbauer, S.; Gaube, F.; Möllmann, U.; Dahse, H.-M.; Schmidtke, M.; Gareis, M.; Pickhardt, M.; Liebermann, B. Antimicrobial, antiproliferative, cytotoxic, and tau inhibitory activity of rubellins and caeruleoramularin produced by the phytopathogenic fungus Ramularia collo-cygni. Planta Med. 2009, 75, 1523–1525. [Google Scholar] [CrossRef]
- Lin, Y.-R.; Lo, C.-T.; Liu, S.-Y.; Peng, K.-C. Involvement of pachybasin and emodin in self-regulation of Trichoderma harzianum mycoparasitic coiling. J. Agric. Food Chem. 2012, 60, 2123–2128. [Google Scholar] [CrossRef]
- Andolfi, A.; Cimmino, A.; Villegas-Fernández, A.M.; Tuzi, A.; Santini, A.; Melck, D.; Rubiales, D.; Evidente, A. Lentisone, a new phytotoxic anthraquinone produced by Ascochyta lentis, the causal agent of Ascochyta blight in Lens culinaris. J. Agric. Food Chem. 2013, 61, 7301–7308. [Google Scholar] [CrossRef]
- Masi, M.; Nocera, P.; Zonno, M.C.; Tuzi, A.; Pescitelli, G.; Cimmino, A.; Boari, A.; Infantino, A.; Vurro, M.; Evidente, A. Lentiquinones A, B, and C, phytotoxic anthraquinone derivatives isolated from Ascochyta lentis, a pathogen of lentil. J. Nat. Prod. 2018, 81, 2700–2709. [Google Scholar] [CrossRef]
- Evidente, A.; Lanzetta, R.; Capasso, R.; Vurro, M.; Botralico, A. Pinolidoxin, a phytotoxic nonenolide from Ascochyta pinodes. Phytochemistry 1993, 34, 999–1003. [Google Scholar] [CrossRef]
- Cimmino, A.; Andolfi, A.; Fondevilla, S.; Abouzeid, M.A.; Rubiales, D.; Evidente, A. Pinolide, a new nonenolide produced by Didymella pinodes, the causal agent of Ascochyta blight on Pisum sativum. J. Agric. Food Chem. 2012, 60, 5273–5278. [Google Scholar] [CrossRef] [PubMed]
- Evidente, A.; Lanzetta, R.; Capasso, R.; Andolfi, A.; Bottalico, A.; Vurro, M.; Zonno, M.C. Putaminoxin, a phytotoxic nonenolide from Phoma putaminum. Phytochemistry 1995, 40, 1637–1641. [Google Scholar] [CrossRef]
- Vurro, M.; Ellis, B.E. Effect of fungal toxins on induction of phenylalanine ammonia-lyase activity in elicited cultures of hybrid poplar. Plant Sci. 1997, 126, 29–38. [Google Scholar] [CrossRef]
- Evidente, A.; Capasso, R.; Andolfi, A.; Vurro, M.; Zonno, M.C. Structure–Activity relationship studies of putaminoxins and pinolidoxins: Phytotoxic nonenolides produced by phytopathogenic Phoma and Ascochyta species. Nat. Toxins 1998, 6, 183–188. [Google Scholar] [CrossRef]
- Kim, W.; Park, J.-J.; Dugan, F.M.; Peever, T.L.; Gang, D.R.; Vandemark, G.; Chen, W. Production of the antibiotic secondary metabolite solanapyrone A by the fungal plant pathogen Ascochyta rabiei during fruiting body formation in saprobic growth. Environ. Microbiol. 2017, 19, 1822–1835. [Google Scholar] [CrossRef] [PubMed]
- Tietjen, K.G.; Schaller, E.; Matern, U. Phytotoxins from Alternaria carthami chowdhury: Structural identification and physiological significance. Physiol. Plant Pathol. 1983, 23, 387–400. [Google Scholar] [CrossRef]
- Härri, E.; Loeffler, W.; Sigg, H.P.; Stähelin, H.; Tamm, C. Über die Isolierung neuer Stoffwechselprodukte aus Penicillium brefeldianum DODGE. Helv. Chim. Acta 1963, 46, 1235–1243. [Google Scholar] [CrossRef]
- Silletta, M.G.; Colanzi, A.; Weigert, R.; Di Girolamo, M.; Santone, I.; Fiucci, G.; Mironov, A.; Matteis, M.A.D.; Luini, A.; Corda, D. Role of brefeldin A-dependent ADP-ribosylation in the control of intracellular membrane transport. In ADP-Ribosylation Reactions: From Bacterial Pathogenesis to Cancer; Alvarez-Gonzalez, R., Ed.; Springer: Boston, MA, USA, 1999; pp. 43–51. [Google Scholar]
- Colanzi, A.; Grimaldi, G.; Catara, G.; Valente, C.; Cericola, C.; Liberali, P.; Ronci, M.; Lalioti, V.S.; Bruno, A.; Beccari, A.R.; et al. Molecular mechanism and functional role of brefeldin A-mediated ADP-ribosylation of CtBP1/BARS. Proc. Natl. Acad. Sci. USA 2013, 110, 9794–9799. [Google Scholar] [CrossRef] [Green Version]
- Weber, R.W.S.; Stenger, E.; Meffert, A.; Hahn, M. Brefeldin A production by Phoma medicaginis in dead pre-colonized plant tissue: A strategy for habitat conquest? Mycol. Res. 2004, 108, 662–671. [Google Scholar] [CrossRef]
- Fausto Rivero-Cruz, J.; Garcı́a-Aguirre, G.; Cerda-Garcı́a-Rojas, C.M.; Mata, R. Conformational behavior and absolute stereostructure of two phytotoxic nonenolides from the fungus Phoma herbarum. Tetrahedron 2000, 56, 5337–5344. [Google Scholar] [CrossRef]
- Boruwa, J.; Gogoi, N.; Barua, C.N. A short enantioselective total synthesis of the phytotoxic lactone herbarumin III. Org. Biomol. Chem. 2006, 4, 3521–3525. [Google Scholar] [CrossRef] [PubMed]
- Jangili, P.; Kashanna, J.; Kumar, C.G.; Poornachandra, Y.; Das, B. Stereoselective total synthesis of a novel regiomer of herbarumin I and its cytotoxic and antimicrobial activities. Bioorg. Med. Chem. Lett. 2014, 24, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Tamura, G.; Suzuki, S.; Takatsuki, A.; Ando, K.; Arima, K. Ascochlorin, a new antibiotic, found by paper-disc agar-diffusion method. J. Antibiot. 1968, 21, 539–544. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, H.; Hosokawa, T.; Sawada, M.; Ando, K. Isolation and structure of ascofuranone and ascofranol, antibiotics with hypolipidemic activity. J. Antibiot. 1973, 26, 676–680. [Google Scholar] [CrossRef] [PubMed]
- Hijikawa, Y.; Matsuzaki, M.; Suzuki, S.; Inaoka, D.K.; Tatsumi, R.; Kido, Y.; Kita, K. Re-identification of the ascofuranone-producing fungus Ascochyta viciae as Acremonium sclerotigenum. J. Antibiot. 2017, 70, 304–307. [Google Scholar] [CrossRef]
- Ellestad, G.A.; Evans, R.H.; Kunstmann, M.P. Some new terpenoid metabolites from an unidentified fusarium species. Tetrahedron 1969, 25, 1323–1334. [Google Scholar] [CrossRef]
- Kato, A.; Ando, K.; Tamura, G.; Arima, K. Cylindrochlorin, a new antibiotic produced by Cylindrocladium. J. Antibiot. 1970, 23, 168–169. [Google Scholar] [CrossRef]
- Kosuge, Y.; Suzuki, A.; Hirata, S.; Tamura, S. Structure of colletochlorin from Colletotrichum nicotianae. Agric. Biol. Chem. 1973, 37, 455–456. [Google Scholar] [CrossRef]
- Cagnoli-Bellavita, N.; Ceccherelli, P.; Fringuelli, R.; Ribaldi, M. Ascochlorin: A terpenoid metabolite from Acremonium luzulae. Phytochemistry 1975, 14, 807. [Google Scholar] [CrossRef]
- Takamatsu, S.; Rho, M.-C.; Masuma, R.; Hayashi, M.; Komiyama, K.; Tanaka, H.; Omura, S. A novel testosterone 5α-reductase inhibitor, 8’, 9’-dehydroascochlorin produced by Verticillium sp. FO-2787. Chem. Pharm. Bull. 1994, 42, 953–956. [Google Scholar] [CrossRef]
- Takatsuki, A.; Tamura, G.; Arima, K. Antiviral and antitumor antibiotics. XIV. Effects of ascochlorin and other respiration inhibitors on multiplication of Newcastle disease virus in cultured cells. Appl. Microbiol. 1969, 17, 825–829. [Google Scholar] [PubMed]
- Minagawa, N.; Yabu, Y.; Kita, K.; Nagai, K.; Ohta, N.; Meguro, K.; Sakajo, S.; Yoshimoto, A. An antibiotic, ascofuranone, specifically inhibits respiration and in vitro growth of long slender bloodstream forms of Trypanosoma brucei brucei. Mol. Biochem. Parasitol. 1996, 81, 127–136. [Google Scholar] [CrossRef]
- Nihei, C.; Fukai, Y.; Kawai, K.; Osanai, A.; Yabu, Y.; Suzuki, T.; Ohta, N.; Minagawa, N.; Nagai, K.; Kita, K. Purification of active recombinant trypanosome alternative oxidase. FEBS Lett. 2003, 538, 35–40. [Google Scholar] [CrossRef]
- Mogi, T.; Ui, H.; Shiomi, K.; Ōmura, S.; Miyoshi, H.; Kita, K. Antibiotics LL-Z1272 identified as novel inhibitors discriminating bacterial and mitochondrial quinol oxidases. Biochim. Biophys. Acta-Bioenerg. 2009, 1787, 129–133. [Google Scholar] [CrossRef] [PubMed]
- Berry, E.A.; Huang, L.; Lee, D.-W.; Daldal, F.; Nagai, K.; Minagawa, N. Ascochlorin is a novel, specific inhibitor of the mitochondrial cytochrome bc1 complex. Biochim. Biophys. Acta Bioenerget. 2010, 1797, 360–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ichihara, A.; Tazaki, H.; Sakamura, S. Solanapyrones A, B and C, phytotoxic metabolites from the fungus Alternaria solani. Tetrahedron Lett. 1983, 24, 5373–5376. [Google Scholar] [CrossRef]
- Alam, S.S.; Bilton, J.N.; Slawin, A.M.Z.; Williams, D.J.; Sheppard, R.N.; Strange, R.N. Chickpea blight: Production of the phytotoxins solanapyrones A and C by Ascochyta rabiei. Phytochemistry 1989, 28, 2627–2630. [Google Scholar] [CrossRef]
- Kaur, S. Phytotoxicity of solanapyrones produced by the fungus Ascochyta rabiei and their possible role in blight of chickpea (Cicer arietinum). Plant Sci. 1995, 109, 23–29. [Google Scholar] [CrossRef]
- Höhl, B.; Weidemann, C.; Höhl, U.; Barz, W. Isolation of solanapyrones A, B and C from culture filture and spore germination fluids of Ascochyta rabiei and aspects of phytotoxin action. J. Phytopathol. 1991, 132, 193–206. [Google Scholar] [CrossRef]
- Hamid, K.; Strange, R.N. Phytotoxicity of solanapyrones A and B produced by the chickpea pathogen Ascochyta rabiei (Pass.) Labr. and the apparent metabolism of solanapyrone A by chickpea tissues. Physiol. Mol. Plant Pathol. 2000, 56, 235–244. [Google Scholar] [CrossRef]
- Kim, W.; Park, C.-M.; Park, J.-J.; Akamatsu, H.O.; Peever, T.L.; Xian, M.; Gang, D.R.; Vandemark, G.; Chen, W. Functional analyses of the Diels-Alderase gene sol5 of Ascochyta rabiei and Alternaria solani indicate that the solanapyrone phytotoxins are not required for pathogenicity. Mol. Plant Microbe Interact. 2014, 28, 482–496. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, K.M.; Toske, S.G.; Jensen, P.R.; Fenical, W. Solanapyrones e-g, antialgal metabolites produced by a marine fungus. Phytochemistry 1998, 49, 2299–2304. [Google Scholar] [CrossRef]
- Schmidt, L.E.; Gloer, J.B.; Wicklow, D.T. Solanapyrone analogues from a Hawaiian fungicolous fungus. J. Nat. Prod. 2007, 70, 1317–1320. [Google Scholar] [CrossRef] [PubMed]
- Trisuwan, K.; Rukachaisirikul, V.; Sukpondma, Y.; Preedanon, S.; Phongpaichit, S.; Sakayaroj, J. Pyrone derivatives from the marine-derived fungus Nigrospora sp. PSU-F18. Phytochemistry 2009, 70, 554–557. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.-H.; Chen, Y.-W.; Shao, S.-C.; Wang, L.-D.; Yu, Y.; Li, Z.-Y.; Yang, L.-Y.; Li, S.-L.; Huang, R. Two new solanapyrone analogues from the endophytic fungus Nigrospora sp. YB-141 of Azadirachta indica. Chem. Biodivers. 2009, 6, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-Z.; Luo, X.-H.; Xiao, J.; Zhai, M.-M.; Yuan, Y.; Zhu, Y.; Crews, P.; Yuan, C.-S.; Wu, Q.-X. Pyrone derivatives from the endophytic fungus Alternaria tenuissima SP-07 of Chinese herbal medicine Salvia przewalskii. Fitoterapia 2014, 99, 184–190. [Google Scholar] [CrossRef]
- Mizushina, Y.; Kamisuki, S.; Kasai, N.; Shimazaki, N.; Takemura, M.; Asahara, H.; Linn, S.; Yoshida, S.; Matsukage, A.; Koiwai, O.; et al. A plant phytotoxin, solanapyrone A, is an inhibitor of DNA polymerase β and λ. J. Biol. Chem. 2002, 277, 630–638. [Google Scholar] [CrossRef]
- Garcı́a-Dı́az, M.; Domı́nguez, O.; López-Fernández, L.A.; de Lera, L.T.; Sanı́ger, M.L.; Ruiz, J.F.; Párraga, M.; Garcı́a-Ortiz, M.J.; Kirchhoff, T.; del Mazo, J.; et al. DNA polymerase lambda (Pol λ), a novel eukaryotic DNA polymerase with a potential role in meiosis. J. Mol. Biol. 2000, 301, 851–867. [Google Scholar] [CrossRef]
- Yamtich, J.; Sweasy, J.B. DNA polymerase family X: Function, structure, and cellular roles. Biochim. Biophys. Acta Proteins Proteom. 2010, 1804, 1136–1150. [Google Scholar] [CrossRef]
- Durrant, W.E.; Wang, S.; Dong, X. Arabidopsis SNI1 and RAD51D regulate both gene transcription and DNA recombination during the defense response. Proc. Natl. Acad. Sci. USA 2007, 104, 4223–4227. [Google Scholar] [CrossRef]
- Song, J.; Durrant, W.E.; Wang, S.; Yan, S.; Tan, E.H.; Dong, X. DNA repair proteins are directly involved in regulation of gene expression during plant immune response. Cell Host Microbe 2011, 9, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Durrant, W.E.; Song, J.; Spivey, N.W.; Dong, X. Arabidopsis BRCA2 and RAD51 proteins are specifically involved in defense gene transcription during plant immune responses. Proc. Natl. Acad. Sci. USA 2010, 107, 22716–22721. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Wang, W.; Marqués, J.; Mohan, R.; Saleh, A.; Durrant, W.E.; Song, J.; Dong, X. Salicylic acid activates DNA damage responses to potentiate plant immunity. Mol. Cell 2013, 52, 602–610. [Google Scholar] [CrossRef] [PubMed]
- Wenke, J.; Anke, H.; Sterner, O. Pseurotin A and 8-O-demethylpseurotin A from Aspergillus fumigatus and their inhibitory activities on chitin synthase. Biosci. Biotechnol. Biochem. 1993, 57, 961–964. [Google Scholar] [CrossRef]
- Ishikawa, M.; Ninomiya, T.; Akabane, H.; Kushida, N.; Tsujiuchi, G.; Ohyama, M.; Gomi, S.; Shito, K.; Murata, T. Pseurotin A and its analogues as inhibitors of immunoglobuline E production. Bioorg. Med. Chem. Lett. 2009, 19, 1457–1460. [Google Scholar] [CrossRef]
- Vurro, M.; Zonno, M.C.; Evidente, A.; Capasso, R.; Bottaiico, A. Isolation of cytochalasins A and B from Ascochyta lathyri. Mycotox Res. 1992, 8, 17–20. [Google Scholar] [CrossRef]
- Latif, Z.; Strange, R.N.; Bilton, J.; Riazuddin, S. Production of the phytotoxins, solanapyrones A and C and cytochalasin D among nine isolates of Ascochyta rabiei. Plant Pathol. 1993, 42, 172–180. [Google Scholar] [CrossRef]
- Masi, M.; Nocera, P.; Boari, A.; Cimmino, A.; Zonno, M.C.; Infantino, A.; Vurro, M.; Evidente, A. Lathyroxins A and B, phytotoxic monosubstituted phenols isolated from Ascochyta lentis var. lathyri, a fungal pathogen of grass pea (Lathyrus sativus). J. Nat. Prod. 2018, 81, 1093–1097. [Google Scholar]
- Chen, H.; Fujita, M.; Feng, Q.; Clardy, J.; Fink, G.R. Tyrosol is a quorum-sensing molecule in Candida albicans. Proc. Natl. Acad. Sci. USA 2004, 101, 5048–5052. [Google Scholar] [CrossRef]
- Kasahara, K.; Miyamoto, T.; Fujimoto, T.; Oguri, H.; Tokiwano, T.; Oikawa, H.; Ebizuka, Y.; Fujii, I. Solanapyrone synthase, a possible Diels–Alderase and iterative type I polyketide synthase encoded in a biosynthetic gene cluster from Alternaria solani. ChemBioChem 2010, 11, 1245–1252. [Google Scholar] [CrossRef]
- Kim, W.; Park, J.-J.; Gang, D.R.; Peever, T.L.; Chen, W. A novel type pathway-specific regulator and dynamic genome environments of a solanapyrone biosynthesis gene cluster in the fungus Ascochyta rabiei. Eukaryot. Cell 2015, 14, 1102–1113. [Google Scholar] [CrossRef] [PubMed]
- Fujii, I.; Yoshida, N.; Shimomaki, S.; Oikawa, H.; Ebizuka, Y. An iterative type I polyketide synthase PKSN catalyzes synthesis of the decaketide alternapyrone with regio-specific octa-methylation. Chem. Biol. 2005, 12, 1301–1309. [Google Scholar] [CrossRef] [PubMed]
- Kasahara, K.; Fujii, I.; Oikawa, H.; Ebizuka, Y. Expression of Alternaria solani PKSF generates a set of complex reduced-type polyketides with different carbon-lengths and cyclization. ChemBioChem 2006, 7, 920–924. [Google Scholar] [CrossRef] [PubMed]
- Ostenfeld Larsen, T.; Perry, N.B.; Andersen, B. Infectopyrone, a potential mycotoxin from Alternaria infectoria. Tetrahedron Lett. 2003, 44, 4511–4513. [Google Scholar] [CrossRef]
- Kono, Y.; Gardner, J.M.; Suzuki, Y.; Takeuchi, S. Plant pathotoxins from Alternaria citri: The minor ACRL toxins. Phytochemistry 1985, 24, 2869–2874. [Google Scholar] [CrossRef]
- Katayama, K.; Kobayashi, T.; Oikawa, H.; Honma, M.; Ichihara, A. Enzymatic activity and partial purification of solanapyrone synthase: First enzyme catalyzing Diels–Alder reaction. Biochim. Biophys. Acta Protein Struct. Mol. Enzymol. 1998, 1384, 387–395. [Google Scholar] [CrossRef]
- Shimizu, T.; Kinoshita, H.; Ishihara, S.; Sakai, K.; Nagai, S.; Nihira, T. Polyketide synthase gene responsible for citrinin biosynthesis in Monascus purpureus. Appl. Environ. Microbiol. 2005, 71, 3453–3457. [Google Scholar] [CrossRef]
- He, Y.; Cox, R.J. The molecular steps of citrinin biosynthesis in fungi. Chem. Sci. 2016, 7, 2119–2127. [Google Scholar] [CrossRef]
- Kim, W.; Lichtenzveig, J.; Syme, R.A.; Williams, A.H.; Peever, T.L.; Chen, W. Identification of a polyketide synthase gene responsible for ascochitine biosynthesis in Ascochyta fabae and its abrogation in sister taxa. mSphere 2019, 4, e00622-19. [Google Scholar] [CrossRef]
- Beed, F.D.; Strange, R.N.; Onfroy, C.; Tivoli, B. Virulence for faba bean and production of ascochitine by Ascochyta fabae. Plant Pathol. 2018, 43, 987–997. [Google Scholar] [CrossRef]
- Araki, Y.; Awakawa, T.; Matsuzaki, M.; Cho, R.; Matsuda, Y.; Hoshino, S.; Shinohara, Y.; Yamamoto, M.; Kido, Y.; Inaoka, D.K.; et al. Complete biosynthetic pathways of ascofuranone and ascochlorin in Acremonium egyptiacum. Proc. Natl. Acad. Sci. USA 2019, 116, 8269–8274. [Google Scholar] [CrossRef] [PubMed]
- Chiang, Y.-M.; Szewczyk, E.; Davidson, A.D.; Entwistle, R.; Keller, N.P.; Wang, C.C.C.; Oakley, B.R. Characterization of the Aspergillus nidulans monodictyphenone gene cluster. Appl. Environ. Microbiol. 2010, 76, 2067–2074. [Google Scholar] [CrossRef] [PubMed]
- Zabala, A.O.; Chooi, Y.-H.; Choi, M.S.; Lin, H.-C.; Tang, Y. Fungal polyketide synthase product chain-length control by partnering thiohydrolase. ACS Chem. Biol. 2014, 9, 1576–1586. [Google Scholar] [CrossRef] [PubMed]
- Turo, C.J. Genomic Analysis of Fungal Species Causing Ascochyta Blight in Field Pea. Ph.D. Thesis, Curtin University, Bentley, WA, Australia, February 2016. [Google Scholar]
- Qiao, K.; Chooi, Y.-H.; Tang, Y. Identification and engineering of the cytochalasin gene cluster from Aspergillus clavatus NRRL 1. Metab. Eng. 2011, 13, 723–732. [Google Scholar] [CrossRef] [PubMed]
- Maiya, S.; Grundmann, A.; Li, X.; Li, S.-M.; Turner, G. Identification of a hybrid PKS/NRPS required for pseurotin A biosynthesis in the human pathogen Aspergillus fumigatus. ChemBioChem 2007, 8, 1736–1743. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Class | Biological Function | Producing Fungi (Host) [Reference] |
|---|---|---|---|
| ascochitine | polyketide | antifungal phytotoxic | Ascochyta fabae (faba bean) [16,23,102]; A. hyalospora (lambsquarters) [25]; A. pisi (pea) [16,22]; A. salicorniae (Ulva sp., an alga) [27]; A. viciae-villosae (hairy vetch) [16,102]; Phoma clematidina (Clematis spp.) [24]; P. koolunga (pea) [16]; many Ascochyta-like spp. isolated from different legumes [16] |
| ascochlorin | meroterpenoid | antitrypanosomal | A. viciae (Vicia spp., vetches) [56,64] |
| ascofuranone | meroterpenoid | antitrypanosomal | A. viciae (Vicia spp., vetches) [57,65] |
| ascosalitoxin | polyketide | phytotoxic | A. salicorniae (Ulva sp., an alga) [27] |
| brefeldin A | polyketide | phytotoxic | P. medicaginis (Medicago spp.) [52] |
| cytochalasin A/B | hybrid 1 | cytotoxic | A. lathyri (grass pea) [89] |
| herbarumin II | polyketide | phytotoxic | A. pinodes (pea) [43,53] |
| lathyroxin A/B | simple phenolics | phytotoxic | A. lentis var. lathyri (grass pea) [91] |
| lentiquinone A | polyketide | antibacterial (mild) phytotoxic | A. lentis (lentil) [41] |
| lentiquinone C | polyketide | antibacterial phytotoxic (mild) | A. lentis (lentil) [41] |
| lentisone | polyketide | antibacterial, phytotoxic | A. lentis (lentil) [40] |
| pachybasin | polyketide | induction of mycoparasitic coiling | A. lentis (lentil) [43] |
| pinolidoxin | polyketide | phytotoxic | A. pinodes (pea) [16,42]; A. pinodella (chickpea, lentil) [16] |
| pseurotin A | hybrid 1 | phytotoxic (mild) | A. lentis (lentil) [40] |
| putaminoxin | polyketide | phytotoxic | A. pinodes (pea) [43] |
| pyrenolide A | polyketide | antifungal | A. hyalospora (lambsquarters) [25] |
| solanapyrone A | polyketide | phytotoxic, antibacterial antifungal | A. rabiei (chickpea) [16,47,70] |
| solanapyrone C | polyketide | algistatic, antibacterial | A. rabiei (chickpea) [16,47,70] |
| tyrosol | simple phenolics | phytotoxic (mild) quorum sensing | A. lentis (lentil) [40,92] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, W.; Chen, W. Phytotoxic Metabolites Produced by Legume-Associated Ascochyta and Its Related Genera in the Dothideomycetes. Toxins 2019, 11, 627. https://doi.org/10.3390/toxins11110627
Kim W, Chen W. Phytotoxic Metabolites Produced by Legume-Associated Ascochyta and Its Related Genera in the Dothideomycetes. Toxins. 2019; 11(11):627. https://doi.org/10.3390/toxins11110627
Chicago/Turabian StyleKim, Wonyong, and Weidong Chen. 2019. "Phytotoxic Metabolites Produced by Legume-Associated Ascochyta and Its Related Genera in the Dothideomycetes" Toxins 11, no. 11: 627. https://doi.org/10.3390/toxins11110627
APA StyleKim, W., & Chen, W. (2019). Phytotoxic Metabolites Produced by Legume-Associated Ascochyta and Its Related Genera in the Dothideomycetes. Toxins, 11(11), 627. https://doi.org/10.3390/toxins11110627