A Polyphasic Approach to Compare the Genomic Profiles of Aflatoxigenic and Non-Aflatoxigenic Isolates of Aspergillus Section Flavi


Abstract
:1. Introduction
2. Results
2.1. Toxin Analysis
2.2. Morphological and Molecular Identification
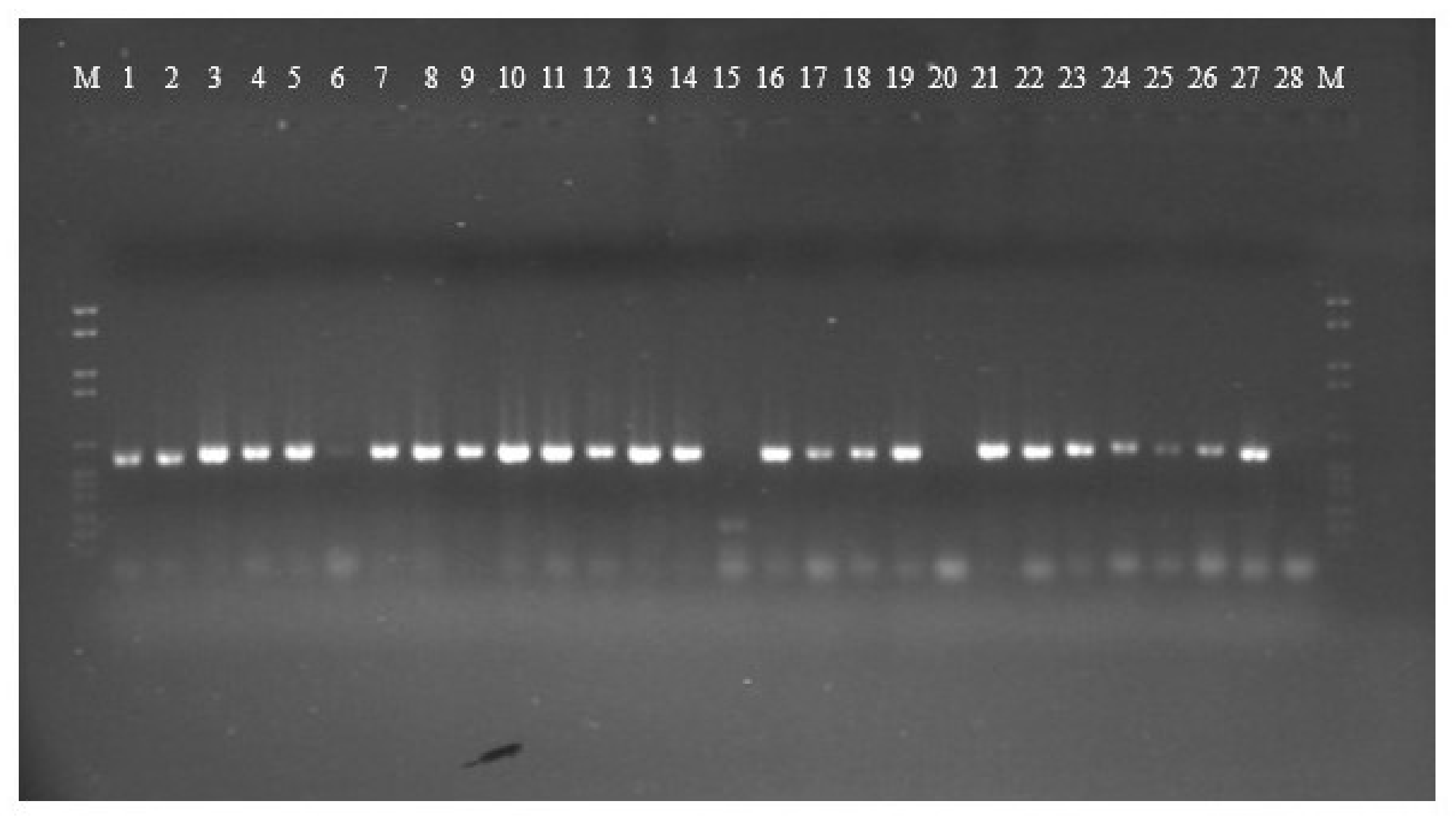
2.3. Genotyping Analysis of ITS Sequences
2.4. Correlation between aflR Gene Profile and Aflatoxin Production
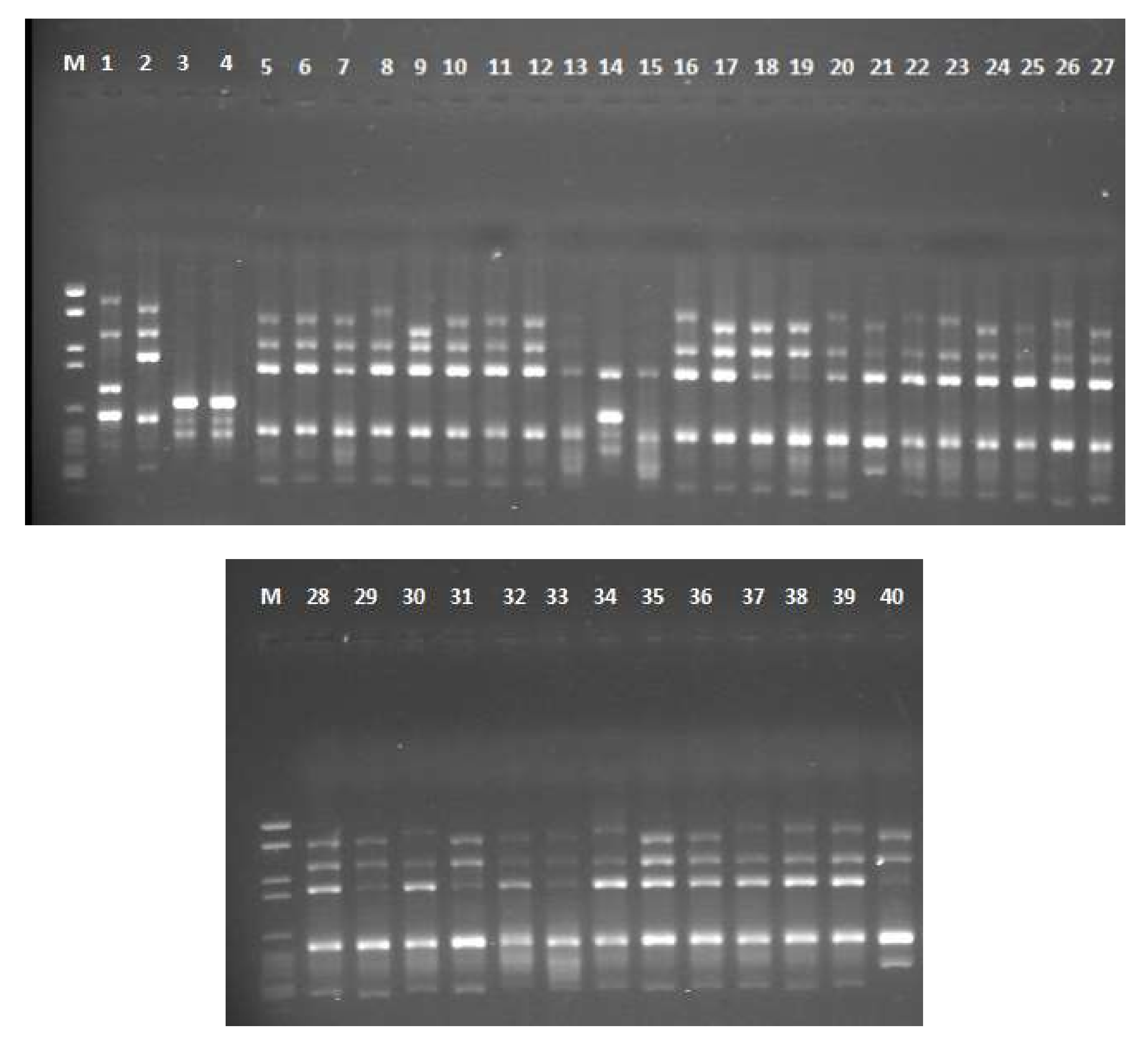
2.5. RAPD and ISSR Banding Pattern, PIC and MI
2.6. Allelic and Genetic Diversity among Aspergillus Isolates
2.7. AMOVA Analysis
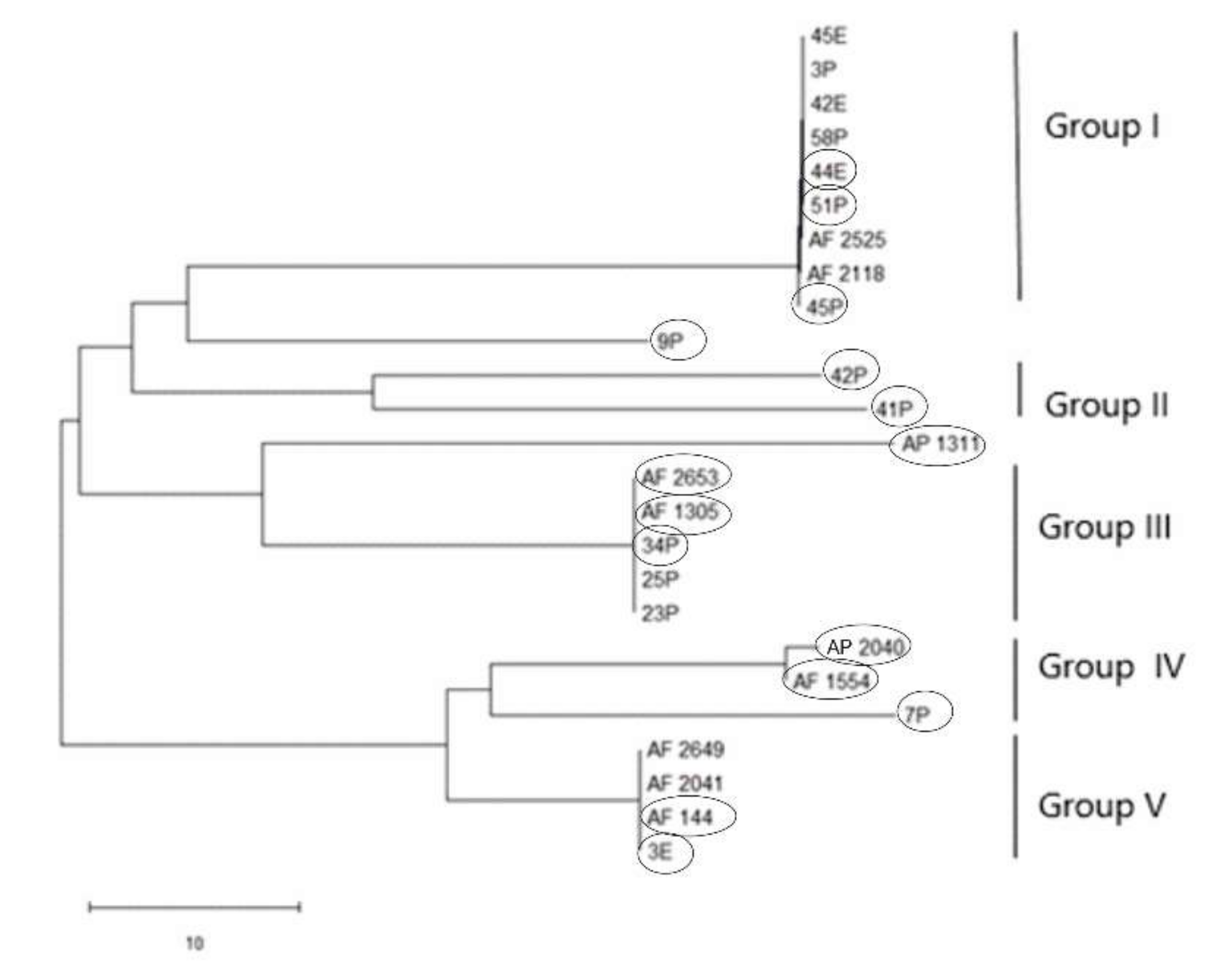
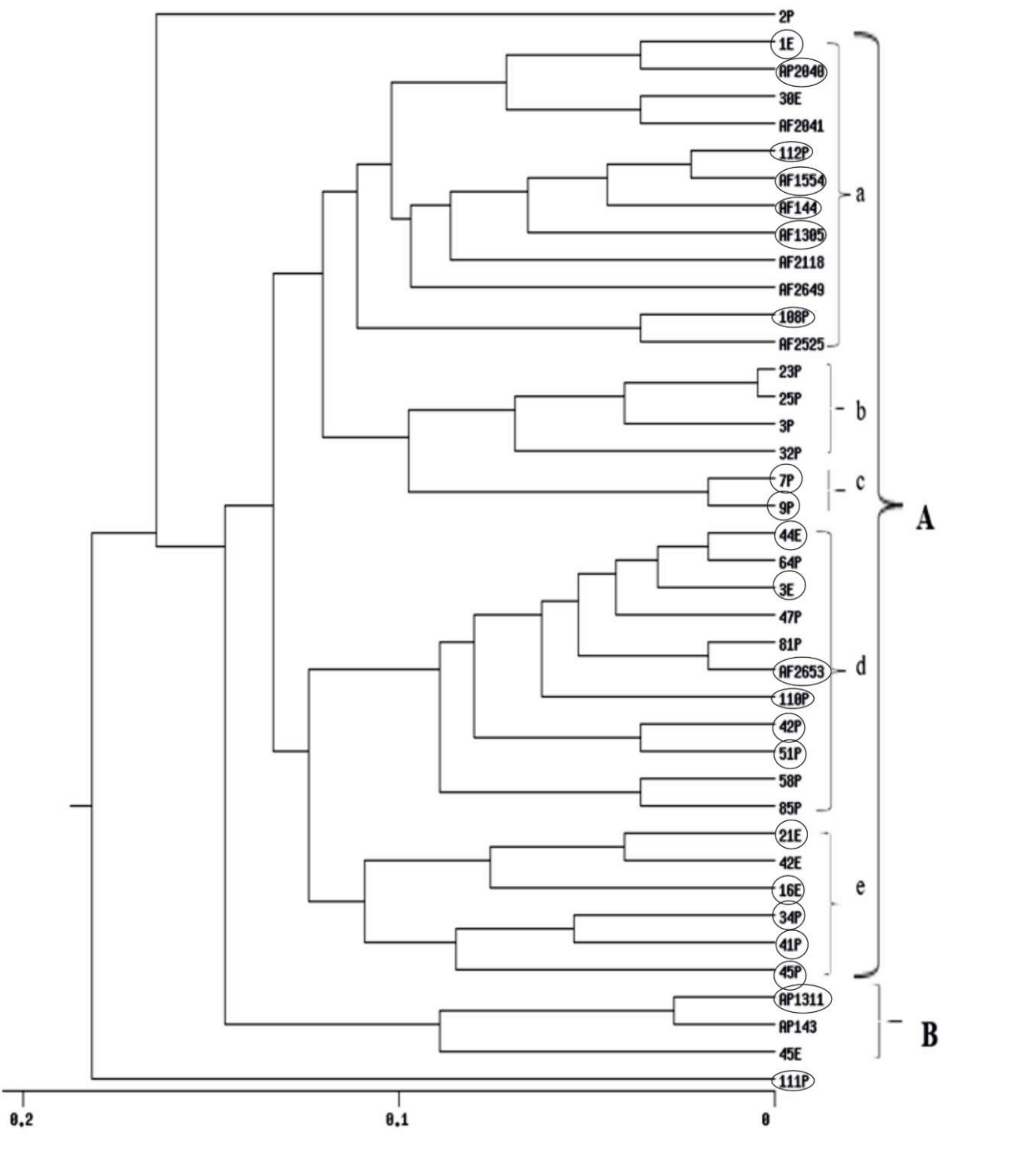
2.8. RAPD and ISSR Dendrogram Analyses
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Chemicals, Reagents, and Media
5.2. Source of Aspergillus Isolates
5.3. Detection of Aflatoxins (AFB1, AFB2, AFG1, and AFG2) using HPLC
5.4. Molecular Studies of 40 Examined Aspergillus Isolates (Genotypic Analyses)
5.4.1. DNA Extraction
5.4.2. PCR Amplification and Sequencing
5.4.3. RAPD and ISSR Amplifications
5.4.4. Gel Electrophoresis
5.5. Phylogenetic Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kumar, P.; Mahato, D.K.; Kamle, M.; Mohanta, T.K.; Kang, S.G. Aflatoxins: A Global Concern for Food Safety, Human Health and Their Management. Front. Microbiol. 2017, 7, 21–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amaike, S.; Keller, N.P. Aspergillus flavus . Annu. Rev. Phytopathol. 2011, 49, 107–133. [Google Scholar] [CrossRef] [PubMed]
- Risk Assessment on Aflatoxins; Joint FAO/WHO Expert Committee on Food Additives (JECFA): Geneva, Switzerland, 2018; REF. No.: WHO/NHM/FOS/RAM/18.1.
- Reddy, K.R.N. An overview of mycotoxin contamination in foods and its implications for human health. Toxin Rev. 2010, 29, 3–26. [Google Scholar] [CrossRef]
- Jang, J.H.; Lee, J.H.; Ki, C.S.; Lee, N.Y. Identification of Clinical Mold Isolates by Sequence Analysis of the Internal Transcribed Spacer Region, Ribosomal Large-Subunit D1/D2, and β-Tubulin. Ann. Lab. Med. 2012, 2, 126–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varga, J.; Frisvad, J.C.; Samson, R.A. Two new aflatoxin producing species and an overview of Aspergillus section. Flavi. Stud. Mycol. 2011, 69, 57–80. [Google Scholar] [CrossRef] [PubMed]
- Pildain, M.B.; Frisvad, J.C.; Vaamonde, G.; Cabral, D.; Varga, J.; Samson, R.A. Two novel Aflatoxin-producing Aspergillus species from Argentinean peanuts. Int. J. Syst. Evol. Microbiol. 2008, 58, 725–735. [Google Scholar] [CrossRef] [Green Version]
- Taylor, J.W.; Jacobson, D.J.; Kroken, S.; Kasuga, T.; Geiser, D.M.; Hibbett, D.S.; Fisher, M.C. Phylogenetic species recognition and species concepts in Fungi. Fungal Genet. Biol. 2000, 31, 21–31. [Google Scholar] [CrossRef] [Green Version]
- Yli-Mattila, T.; Nayaka, S.C.; Venkataramana, M.; Yörük, E. Multiplex Detection of Fusarium Species. Chapter 18; In Mycotoxigenic Fungi (Methods in Molecular Biology); Moretti, A., Susca, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2017; Volume 1542, pp. 269–291. ISBN 978-1-4939-6705-6. [Google Scholar]
- Okoth, S.; De Boevre, M.; Vidal, A.; Diana Di Mavungu, J.; Landschoot, S.; Kyallo, M.; Njuguna, J.; Harvey, J.; DeSaeger, S. Genetic and Toxigenic Variability within Aspergillus flavus Population Isolated from Maize in Two Diverse Environments in Kenya. Front. Microbiol. 2018, 9, 57. [Google Scholar] [CrossRef]
- Norlia, M.; Jinap, S.; Nor-Khaizura, M.A.R.; Radu, S.; Chin, C.K.; Samsudin, N.I.P.; Farawahida, A.H. Molecular Characterisation of Aflatoxigenic and Non-Aflatoxigenic Strains of Aspergillus Section Flavi Isolated from Imported Peanuts along the Supply Chain in Malaysia. Toxins 2019, 11, 501. [Google Scholar] [CrossRef] [Green Version]
- Cary, J.W.; Calvo, A.M. Regulation of Aspergillus Mycotoxin Biosynthesis. Toxin Rev. 2008, 27, 347–370. [Google Scholar] [CrossRef]
- Kesawat, M.S.; Kumar, B.D. Molecular markers: Its application in crop improvement. J. Crop. Sci. Biotechnol. 2009, 4, 169–181. [Google Scholar] [CrossRef]
- Moe, K.T.; Kwon, S.W.; Park, Y.J. Trends in genomics and molecular marker systems for the development of some underutilized crops. Genes Genom. 2012, 5, 451–466. [Google Scholar] [CrossRef]
- Leonova, I.N. Molecular markers: Implementation in crop plant breeding for identification, introgression and gene pyramiding. Russ. J. Genet. 2013, 6, 464–473. [Google Scholar] [CrossRef]
- Muranty, H.; Jorge, V.; Bastien, C.; Lepoittevin, C.; Bouffier, L.; Sanchez, L. Potential for marker-assisted selection for forest tree breeding: Lessons from 20 years of MAS in crops. Tree Genet. Genom. 2014, 6, 1491–1510. [Google Scholar] [CrossRef] [Green Version]
- Cui, C.; Li, Y.; Liu, Y.; Li, X.; Luo, S.; Zhang, Z.; Wu, R.; Liang, G.; Sun, J.; Peng, J.; et al. Determination of genetic diversity among Saccharina germplasm using ISSR and RAPD markers. C. R. Biol. 2016, 340, 76–86. [Google Scholar] [CrossRef]
- Ganie, S.H.; Upadhyaya, P.; Das, S.; Sharma, M.P. Authentication of medicinal plants by DNA markers. Plant Gene 2015, 4, 83–99. [Google Scholar] [CrossRef] [Green Version]
- Ercan, G.A.; Taskin, M.; Turgut, K. Analysis of genetic diversity in Turkish sesame (Sesamum indicum L.) populations using RAPD markers. Genet. Resour. Crop Evol. 2004, 51, 599–607. [Google Scholar] [CrossRef]
- Balendres, M.A.O.; Karlovsky, P.; Cumagun, C.J.R. Mycotoxigenic Fungi and Mycotoxins in Agricultural Crop Commodities in the Philippines: A Review. Foods 2019, 8, 249. [Google Scholar] [CrossRef] [Green Version]
- Hussien, T.A. Molecular Approaches for Mycotoxin Risk Reduction; University of Turku: Turku, Finland, 2019; ISBN 978-951-29-7694-2. Available online: http://urn.fi/URN:NBN:fi-fe2019052316835 (accessed on 10 June 2019).
- Sultan, Y.; Magan, N. Mycotoxigenic fungi in peanuts from different geographic regions of Egypt. Mycotoxin Res. 2010, 26, 133–140. [Google Scholar] [CrossRef] [Green Version]
- Al-Wadai, S.A.; Al-Othman, M.R.; Mahmoud, M.A.; Abd El-Aziz, A.R.M. Molecular characterization of Aspergillus flavus and aflatoxin contamination of wheat grains from Saudi Arabia. Genet. Mol. Res. 2013, 3, 3335–3352. [Google Scholar] [CrossRef]
- Mahmoud, M.A.; El-Samawaty, A.M.A.; Yassin, M.A.; Abd El-Aziz, A.R.M. Genetic diversity analysis of Aspergillus flavus isolates from plants and air by ISSR markers. Genet. Mol. Res. 2016, 2, 15028081. [Google Scholar] [CrossRef] [PubMed]
- Moretti, A.; Susca, A.; Mulé, G.; Logrieco, A.F.; Proctor, R.H. Molecular biodiversity of mycotoxigenic fungi that threaten food safety. Int. J. Food Microbiol. 2013, 167, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Cotty, P.J. Bio competitive exclusion of toxigenic fungi. In The Mycotoxin Factbook; Barug, D., Bhatnagar, D., van Egmond, H.P., van der Kamp, J.W., van Osenbruggen, W.A., Visconti, A., Eds.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2006; 400p. [Google Scholar]
- Dorner, J.W.; Horn, B.W. Separate and combined applications of non-toxigenic Aspergillus flavus and A. parasiticus for biocontrol of aflatoxin in peanuts. Mycopathologia 2007, 163, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Basaran, P.; Demirbas, R.M. Spectroscopic detection of pharmaceutical compounds from an aflatoxigenic strain of Aspergillus parasiticus. Microbiol. Res. 2010, 6, 516–522. [Google Scholar] [CrossRef]
- Rank, C.; Klejnstrup, M.L.; Petersen, L.M.; Kildgaard, S.; Frisvad, J.C.; Gotfredsen, H.C.; Ostenfeld, L.T. Comparative Chemistry of Aspergillus oryzae (RIB40) and A. flavus (NRRL 3357). Metabolites 2012, 2, 39–56. [Google Scholar] [CrossRef]
- Sales, A.C.; Yoshizawa, T. Updated profile of aflatoxin and Aspergillus section Flavi contamination in rice and its by-products from the Philippines. Food Addit. Contam. 2005, 5, 429–436. [Google Scholar] [CrossRef]
- Waing, K.G.; Abella, E.A.; Kalaw, S.P.; Waing, F.P.; Galvez, C.T. Studies on biodiversity of leaf litter fungi of Central Luzon State University and evaluation of their enzyme producing ability. Curr. Res. Environ. Appl. Mycol. 2015, 5, 269–276. [Google Scholar] [CrossRef]
- El-Kady, I.A.; Youssef, M.S. Survey of mycoflora and mycotoxins in Egyptian soybean seeds. J. Basic Microbiol. 1993, 33, 371–378. [Google Scholar] [CrossRef]
- El-Shanshoury, A.R.; El-Sabbagh, S.M.; Emara, H.A.; Saba, H.E. Occurrence of moulds, toxicogenic capability of Aspergillus flavus and levels of aflatoxins in maize, wheat, rice and peanut from markets in central delta provinces, Egypt. Int. J. Curr. Microbiol. App. Sci. 2014, 3, 852–865. [Google Scholar]
- Sabry, B.A.; Hathout, A.S.; Nooh, A.; Aly, S.E.; Shehata, M.G. The prevalence of aflatoxin and Aspergillus parasiticus in Egyptian sesame seeds. Int.J.Chem.Tech.Res. 2016, 11, 308–319. [Google Scholar]
- Liu, B.H.; Chu, F.S. Regulation of aflR and its product associated with aflatoxin biosynthesis. App. Environ. Microbiol. 1998, 10, 3718–3722. [Google Scholar] [CrossRef] [Green Version]
- Cary, J.W.; Ehrlich, K.C.; Wright, M.; Chang, P.K.; Bhatnagar, D. Generation of aflR disruption mutants of Aspergillus parasiticus. Appl. Microbiol. Biotechnol. 2000, 53, 680–684. [Google Scholar] [CrossRef] [PubMed]
- Hussain, A.; Afzal, A.; Irfan, M.; Abdulla, K. Molecular detection of aflatoxin producing strains of Aspergillus flavus from peanut (Arachis hypogaea). Turkish J. Agric. Food Sci. Technol. 2015, 5, 335–341. [Google Scholar] [CrossRef] [Green Version]
- Gallo, A.; Stea, G.; Battilani, P.; Logrieco, A.F.; Perrone, G. Molecular characterization of an Aspergillus flavus population isolated from maize during the first outbreak of aflatoxin contamination in Italy. Phytopathol. Mediterr. 2012, 51, 198–206. [Google Scholar]
- Levin, R.E. PCR detection of aflatoxin producing fungi and its limitations. Int. J. Food Microbiol. 2012, 156, 1–6. [Google Scholar] [CrossRef]
- Baruah, J.; Pandey, S.K.; Begum, T.; Sarma, N.; Paw, M.; La, M. Molecular diversity assessed amongst high dry rhizome recovery Ginger germplasm (Zingiber officinale Roscoe) from NE-India using RAPD and ISSR markers. Ind. Crops Prod. 2019, 129, 463–471. [Google Scholar] [CrossRef]
- Hussien, T.; Carlobos-Lopez, A.L.; Cumagun, C.J.; Yli-Mattila, T. Identification and quantification of fumonisin-producing Fusarium species in grain and soil samples from Egypt and the Philippines. Phytopathol. Mediterr. 2017, 56, 146–153. [Google Scholar]
- Sosa, M.J.; Cordoba, J.J.; Diaz, C.; Rodriguez, M.; Bermudez, E.; Asensio, M.A.; Nunez, F. Production of cyclopiazonic acid by Penicillium commune isolated from dry-cured ham on a meat extract-based substrate. J. Food Prot. 2002, 65, 988–992. [Google Scholar] [CrossRef]
- AOAC. Natural toxins. In Official Methods of Analysis, 17th ed.; Association of Official Analytical Chemists: Washington, DC, USA, 2000. [Google Scholar]
- Giray, B.; Girgin, G.; Engin, A.B.; Aydin, S. Aflatoxins levels in wheat samples consumed in some regions of Turkey. Food Control 2007, 18, 23–29. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Jurjević, Z.; Kubatova, A.; Kolarik, M.; Hubka, V. Taxonomy of Aspergillus section Petersonii sect. nov encompassing indoor and soil-borne species with predominant tropical distribution. Plant Syst. Evolution. 2015, 301, 2441–2462. [Google Scholar] [CrossRef]
- Geisen, R. Multiplex polymerase chain reaction for the detection of potential aflatoxin and sterigmatocystin producing fungi. Syst. Appl. Microbiol. 1996, 19, 388–392. [Google Scholar] [CrossRef]
- Mahmoud, M.A.; Al-Sohaibani, S.A.; Abdelbacki, A.M.; Al-Othman, M.R.; Abd El-Aziz, A.R.; Kasem, K.K.; Mikhail, M.S.; Sabet, K.K.; Omar, M.R.; Hussein, E.M. Molecular characterization of the pathogenic plant fungus Rhizoctonia solani (Ceratobasidiaceae) isolated from Egypt based on protein and PCRRAPD profiles. Genet. Mol. Res. 2012, 11, 3585–3600. [Google Scholar] [CrossRef] [PubMed]
- Batista, P.P.; Santos, J.F.; Oliveira, N.T.; Pires AP, D.; Motta CM, S.; Luna-Alves Lima, E.A. Genetic characterization of Brazilian strains of Aspergillus flavus using DNA markers. Genet. Mol. Res. 2008, 7, 706–717. [Google Scholar] [CrossRef] [PubMed]
- Amiryousefi, A.; Hyvönen, J.; Poczai, P. iMEC: Online Marker Efficiency Calculator. Appl. Plant. Sci. 2018, 6, e01159. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Geographic Origin | Source | Sample ID | Identification | GenBankaccession Number/Identification Sequence Type | Identity | Aflatoxin Production (ppb) | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| AFG1 | AFB1 | AFG2 | AFB2 | TotalAFs | ||||||
| SRRC culture collection | Cottonseed, USA | AF 144 | A. flavus | MH752568/aflR gene | 529/529 (100%) | ND | 30.68 ± 25.39 | ND | ND | 30.68 |
| Karnataka, India | AF 1305 | A. flavus | KF432854/ITS | 535/535 (100%) | ND | 67.5 ± 0.12 | ND | 0.9 ± 0.45 | 68.4 | |
| Pistachio, USA | AF 1554 | A. flavus | MH752566/aflR gene | 516/516 (100%) | ND | 40.38 ± 34.1 | ND | ND | 40.38 | |
| Peanut, Australia | AF 2041 | A. flavus | MH244421/ITS | 537/537 (100%) | ND | ND | ND | ND | ND | |
| Dried fish, Indonesia | AF 2118 | A. flavus | MN511750/ITS | - | ND | ND | ND | ND | ND | |
| Dead termites, China | AF 2525 | A. flavus | FN398160/aflR gene | 525/525 (100%) | ND | ND | ND | ND | ND | |
| Lung tissue, USA | AF 2649 | A. flavus | AY510451/aflR gene | 528/528 (100%) | ND | ND | ND | ND | ND | |
| Corneal ulcer, USA | AF 2653 | A. flavus | KY630136/aflR gene | 525/525 (100%) | ND | 3.585 ± 0.03 | ND | 2336 ± 6.48 | 2339.6 | |
| Peanuts, Uganda, Africa | AP 143 | A. parasiticus | MN511749/ITS | - | ND | ND | ND | ND | ND | |
| Rice, USA | AP 1311 | A. parasiticus | KC769508/aflR gene | 526/526 (100%) | 157.8 ± 2.57 | 559.1 ± 22.35 | 1422 ± 17.20 | 171.9 ± 3.315 | 2310.8 | |
| Peanut, Australia | AP 2040 | A. parasiticus | MH752575/aflR gene | 524/524 (100%) | 903.4 ± 17.49 | 4.755 ± 0.17 | 23.50 ± 0.47 | ND | 931.7 | |
| Philippines | Soil | 2P | A. tamarii | MN511748/ITS | - | ND | ND | ND | ND | ND |
| Soil | 3P | A. flavus | LN482489/ITS | 539/539 (100%) | ND | ND | ND | ND | ND | |
| Soil | 7P | A. nomius | MH752557/ITS | 505/505 (100%) | 3549 ± 28.3 | 315.2 ± 15.47 | 777.5 ± 7.48 | 72.53 ± 2.03 | 4714.23 | |
| Soil | 9P | A. nomius | AY510454/ITS | 512/512 (100%) | 10705 ± 3.4 | 1334 ± 4.52 | 2035 ± 5.36 | 342.1 ± 20.33 | 14,416.1 | |
| Soil | 23P | A. flavus | KX426971/ITS | 536/536 (100%) | ND | ND | ND | ND | ND | |
| Soil | 25P | A. flavus | MN511747/ITS | - | ND | ND | ND | ND | ND | |
| Soil | 32P | A. flavus | KF432854/ITS | 535/535 (100%) | ND | ND | ND | ND | ND | |
| Soil | 34P | A. flavus | KY630136/aflR gene | 528/528 (100%) | ND | 13.28 ± 2.87 | ND | ND | 13.28 | |
| Soil | 41P | A. flavus | MF094441/aflR gene | 524/524 (100%) | ND | 32.89 ± 12.9 | ND | ND | 32.89 | |
| Soil | 42P | A. flavus | MG720232/aflR gene | 524/524 (100%) | ND | 6.614 ± 1.4 | ND | ND | 6.614 | |
| Soil, peanuts | 45P | A. flavus | FN398161/aflR gene | 475/475 (100%) | ND | 51.04 ± 22.59 | ND | ND | 51.04 | |
| Soil | 47P | A. flavus | MH595954/ITS | 537/537 (100%) | ND | ND | ND | ND | ND | |
| Soil | 51P | A. flavus | FN398157/aflR gene | 529/529 (100%) | ND | 5.058 ± 1.11 | ND | ND | 5.058 | |
| Soil | 58P | A. flavus | MK791661/ITS | 511/511 (100%) | ND | ND | ND | ND | ND | |
| Soil | 64P | A. flavus | KX426971/ITS | 536/536 (100%) | ND | ND | ND | ND | ND | |
| Soil | 81P | A. flavus | MN511746/ITS | - | ND | ND | ND | ND | ND | |
| Soil | 85P | A. flavus | LN482481/ITS | 538/538 (100%) | ND | ND | ND | ND | ND | |
| Soil, coconut | 108P | A. flavus | MN511745/ITS | - | ND | ND | ND | 68.28 ± 20.25 | 68.28 | |
| Soil, Coconut | 110P | A. nomius | MN511744/ITS | - | ND | 87.50 ± 10.58 | 204.8 ± 22.29 | ND | 292.3 | |
| Soil | 111P | A. flavus | KU561938/ITS | 49/51 (94%) | ND | 5.055 ± 0.59 | ND | ND | 5.055 | |
| Peanuts | 112P | A. flavus | MN511743/ITS | - | ND | 58.01 ± 2.57 | ND | 14.11 ± 1.08 | 72.12 | |
| Egypt | Maize | 1E | A. flavus | MN511742/ITS | - | ND | 37.71 ± 13.77 | ND | ND | 37.71 |
| Maize | 3E | A. flavus | MH752568/aflR gene | 531/531 (100%) | ND | 658.1 ± 66.20 | ND | 114.1 ± 14.69 | 772.2 | |
| Maize | 16E | A. flavus | JF729324/ITS | 529/529 (100%) | ND | 91.06 ± 13.64 | ND | 29.94 ± 1.3 | 121 | |
| Maize | 21E | A. flavus | MG554234/ITS | 547/547 (100%) | ND | 844.0 ± 15.41 | ND | 313.6 ± 12.6 | 1157.6 | |
| Soil | 30E | A. flavus | MH595954/ITS | 535/535 (100%) | ND | ND | ND | ND | ND | |
| Soil | 42E | A. flavus | MH595954/ITS | 531/531 (100%) | ND | ND | ND | ND | ND | |
| Bench sample | 44E | A. flavus | FN398156/aflR gene | 526/526 (100%) | ND | ND | ND | 66.16 ± 9.06 | 66.16 | |
| Air sample | 45E | A. flavus | MH595954/ITS | 532/532 (100%) | ND | ND | ND | ND | ND | |
| Number of Bands | Number of Polymorphic Bands | PPB (%) | PIC Value | MI | |
|---|---|---|---|---|---|
| RAPD markers | |||||
| RAPD 1 | 7 | 6 | 85.7 | 0.76 | 0.55 |
| RAPD 2 | 5 | 5 | 100 | 0.75 | 0.48 |
| RAPD 5 | 5 | 3 | 60 | 0.45 | 0.39 |
| Average | 5.67 | 4.67 | 81.9 | 0.65 | 0.47 |
| ISSR markers | |||||
| (GTG) 5 | 6 | 4 | 66.7 | 0.77 | 0.57 |
| (GACA) 4 | 7 | 5 | 71.4 | 0.79 | 0.55 |
| (AGAG) 4G | 8 | 8 | 100 | 0.81 | 0.56 |
| Average | 7 | 5.67 | 79.37 | 0.79 | 0.56 |
| Marker | Number of Alleles (Mean Na) | Effective Number of Alleles (Mean Ne) | Nei’s Gene Diversity (Mean H) | Shannon’s Diversity Index (Mean I) |
|---|---|---|---|---|
| RAPD | 2.00 | 1.13 | 0.11 | 0.223 |
| ISSR | 2.00 | 1.33 | 0.24 | 0.408 |
| Marker | Source | Df | Ss | Var | % | P-Value |
|---|---|---|---|---|---|---|
| RAPD | Among Pops | 2 | 27.342 | 0.562 | 8% | 0.076 ns |
| Within Pops | 37 | 252.433 | 6.823 | 92% | 0.001 * | |
| Total | 39 | 279.775 | 7.385 | 100% | ||
| ISSR | Among Pops | 2 | 54.423 | 1.516 | 15% | 0.148 ns |
| Within Pops | 37 | 323.777 | 8.751 | 85% | 0.001 * | |
| Total | 39 | 378.200 | 10.267 | 100% |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abbas, A.; Hussien, T.; Yli-Mattila, T. A Polyphasic Approach to Compare the Genomic Profiles of Aflatoxigenic and Non-Aflatoxigenic Isolates of Aspergillus Section Flavi. Toxins 2020, 12, 56. https://doi.org/10.3390/toxins12010056
Abbas A, Hussien T, Yli-Mattila T. A Polyphasic Approach to Compare the Genomic Profiles of Aflatoxigenic and Non-Aflatoxigenic Isolates of Aspergillus Section Flavi. Toxins. 2020; 12(1):56. https://doi.org/10.3390/toxins12010056
Chicago/Turabian StyleAbbas, Asmaa, Taha Hussien, and Tapani Yli-Mattila. 2020. "A Polyphasic Approach to Compare the Genomic Profiles of Aflatoxigenic and Non-Aflatoxigenic Isolates of Aspergillus Section Flavi" Toxins 12, no. 1: 56. https://doi.org/10.3390/toxins12010056
APA StyleAbbas, A., Hussien, T., & Yli-Mattila, T. (2020). A Polyphasic Approach to Compare the Genomic Profiles of Aflatoxigenic and Non-Aflatoxigenic Isolates of Aspergillus Section Flavi. Toxins, 12(1), 56. https://doi.org/10.3390/toxins12010056