Exoproteomics for Better Understanding Pseudomonas aeruginosa Virulence



{kind=link}
{kind=link}
Abstract
1. Introduction
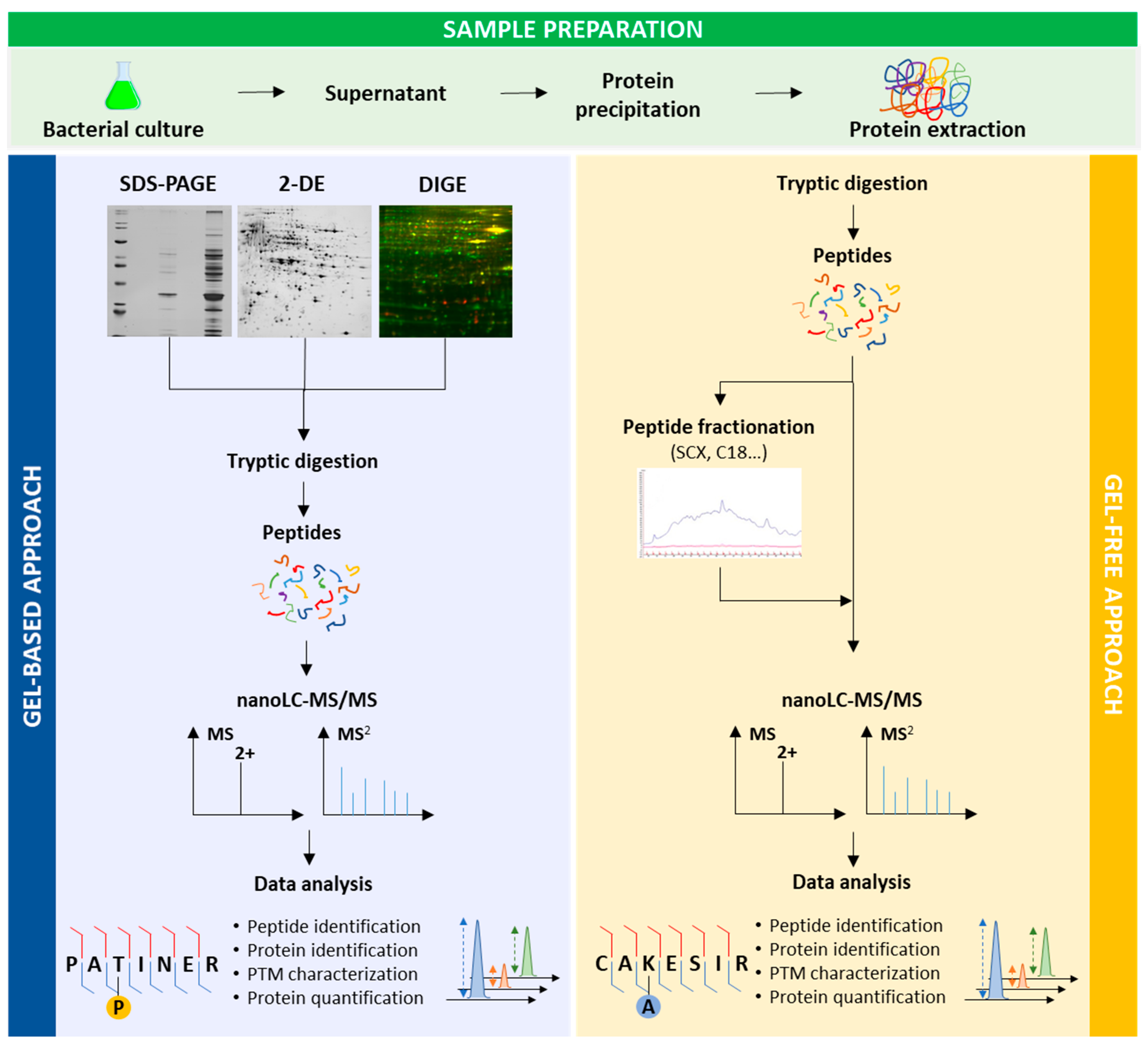
2. Keys to Successful Proteomic Analyses of Exoproteins
3. Proteomics for Exoproteins Characterisation
3.1. Gel-Based Proteomics
3.2. Gel-Free Proteomics
3.3. Exoproteins Subcellular Localisation
3.4. Hypothetical Proteins as New Virulence Factors
4. Proteomics for the Characterisation of PTMs Involved in Virulence
5. Proteomics for Host-Pathogen Studies
6. Conclusions
Funding
Conflicts of Interest
References
- Pachori, P.; Gothalwal, R.; Gandhi, P. Emergence of antibiotic resistance Pseudomonas aeruginosa in intensive care unit; a critical review. Genes Dis. 2019, 6, 109–119. [Google Scholar] [CrossRef]
- Pang, Z.; Raudonis, R.; Glick, B.R.; Lin, T.-J.; Cheng, Z. Antibiotic resistance in Pseudomonas aeruginosa: Mechanisms and alternative therapeutic strategies. Biotechnol. Adv. 2019, 37, 177–192. [Google Scholar] [CrossRef]
- World Health Organization. WHO Publishes List of Bacteria for Which New Antibiotics Are Urgently Needed. Available online: https://www.who.int/news-room/detail/27-02-2017-who-publishes-list-of-bacteria-for-which-new-antibiotics-are-urgently-needed (accessed on 17 April 2020).
- Azam, M.W.; Khan, A.U. Updates on the pathogenicity status of Pseudomonas aeruginosa. Drug Discov. Today 2019, 24, 350–359. [Google Scholar] [CrossRef] [PubMed]
- Rasko, D.A.; Sperandio, V. Anti-virulence strategies to combat bacterial-mediated disease. Nat. Rev. Drug Discov. 2010, 9, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Hauser, A.R. Pseudomonas aeruginosa: So many virulence factors, so little time. Crit. Care Med. 2011, 39, 2193–2194. [Google Scholar] [CrossRef] [PubMed]
- Kipnis, E.; Sawa, T.; Wiener-Kronish, J. Targeting mechanisms of Pseudomonas aeruginosa pathogenesis. Méd. Mal. Infect. 2006, 36, 78–91. [Google Scholar] [CrossRef]
- Newman, J.W.; Floyd, R.V.; Fothergill, J.L. The contribution of Pseudomonas aeruginosa virulence factors and host factors in the establishment of urinary tract infections. FEMS Microbiol. Lett. 2017, 364, fnx124. [Google Scholar] [CrossRef]
- De Kievit, T.R. Quorum sensing in Pseudomonas aeruginosa biofilms. Environ. Microbiol. 2009, 11, 279–288. [Google Scholar] [CrossRef]
- Saint-Criq, V.; Villeret, B.; Bastaert, F.; Kheir, S.; Hatton, A.; Cazes, A.; Xing, Z.; Sermet-Gaudelus, I.; Garcia-Verdugo, I.; Edelman, A.; et al. Pseudomonas aeruginosa LasB protease impairs innate immunity in mice and humans by targeting a lung epithelial cystic fibrosis transmembrane regulator-IL-6-antimicrobial-repair pathway. Thorax 2018, 73, 49–61. [Google Scholar] [CrossRef]
- Voulhoux, R.; Ball, G.; Ize, B.; Vasil, M.L.; Lazdunski, A.; Wu, L.-F.; Filloux, A. Involvement of the twin-arginine translocation system in protein secretion via the type II pathway. EMBO J. 2001, 20, 6735–6741. [Google Scholar] [CrossRef]
- Hauser, A.R. The type III secretion system of Pseudomonas aeruginosa: Infection by injection. Nat. Rev. Microbiol. 2009, 7, 654–665. [Google Scholar] [CrossRef] [PubMed]
- Ho, B.T.; Dong, T.G.; Mekalanos, J.J. A view to a kill: The bacterial type VI secretion system. Cell Host Microbe 2014, 15, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Allsopp, L.P.; Wood, T.E.; Howard, S.A.; Maggiorelli, F.; Nolan, L.M.; Wettstadt, S.; Filloux, A. RsmA and AmrZ orchestrate the assembly of all three type IV secretion systems in Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2017, 114, 7707–7712. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Wang, T.; Chen, G.; Pu, Q.; Liu, Q.; Zhang, Y.; Xu, L.; Wu, M.; Liang, H. A Pseudomonas aeruginosa type VI secretion system regulated by CueR facilitates copper acquisition. PLoS Pathog. 2019, 15, e1008198. [Google Scholar] [CrossRef] [PubMed]
- Lien, Y.-W.; Lai, E.-M. Type VI secretion effectors: Methodologies and biology. Front. Cell. Infect. Microbiol. 2017, 7, 254. [Google Scholar] [CrossRef]
- Chen, L.; Zou, Y.; She, P.; Wu, Y. Composition, function, and regulation of T6SS in Pseudomonas aeruginosa. Microbiol. Res. 2015, 172, 19–25. [Google Scholar] [CrossRef]
- Kang, Q.; Zhang, D. Principle and potential applications of the non-classical protein secretory pathway in bacteria. Appl. Microbiol. Biotechnol. 2020, 104, 953–965. [Google Scholar] [CrossRef]
- Henderson, B.; Martin, A. Bacterial virulence in the moonlight: Multitasking bacterial moonlighting proteins are virulence determinants in infectious disease. Infect. Immun. 2011, 79, 3476–3491. [Google Scholar] [CrossRef]
- Duport, C.; Alpha-Bazin, B.; Armengaud, J. Advanced proteomics as a powerful tool for studying toxins of human bacterial pathogens. Toxins 2019, 11, 576. [Google Scholar] [CrossRef]
- Anantharajah, A.; Mingeot-Leclercq, M.-P.; Van Bambeke, F. Targeting the type three secretion system in Pseudomonas aeruginosa. Trends Pharmacol. Sci. 2016, 37, 734–749. [Google Scholar] [CrossRef]
- Cezairliyan, B.; Ausubel, F.M. Investment in secreted enzymes during nutrient-limited growth is utility dependent. Proc. Natl. Acad. Sci. USA 2017, E7796–E7802. [Google Scholar] [CrossRef] [PubMed]
- Gaviard, C.; Cosette, P.; Jouenne, T.; Hardouin, J. LasB and CbpD virulence factors of Pseudomonas aeruginosa carry multiple post-translational modifications on their lysine residues. J. Proteome Res. 2019, 923–933. [Google Scholar] [CrossRef] [PubMed]
- Macek, B.; Forchhammer, K.; Hardouin, J.; Weber-Ban, E.; Grangeasse, C.; Mijakovic, I. Protein post-translational modifications in bacteria. Nat. Rev. Microbiol. 2019, 17, 651–664. [Google Scholar] [CrossRef] [PubMed]
- Sandri, A.; Ortombina, A.; Boschi, F.; Cremonini, E.; Boaretti, M.; Sorio, C.; Melotti, P.; Bergamini, G.; Lleo, M. Inhibition of Pseudomonas aeruginosa secreted virulence factors reduces lung inflammation in CF mice. Virulence 2018, 9, e1489198. [Google Scholar] [CrossRef] [PubMed]
- Hare, N.J.; Cordwell, S.J. Proteomics of bacterial pathogens: Pseudomonas aeruginosa infections in cystic fibrosis—A case study. Proteom. Clin. Appl. 2010, 4, 228–248. [Google Scholar] [CrossRef]
- Rao, P.S.S.; Yamada, Y.; Tan, Y.P.; Leung, K.Y. Use of proteomics to identify novel virulence determinants that are required for Edwardsiella tarda pathogenesis. Mol. Microbiol. 2004, 53, 573–586. [Google Scholar] [CrossRef]
- Fléchard, M.; Duchesne, R.; Tahrioui, A.; Bouffartigues, E.; Depayras, S.; Hardouin, J.; Lagy, C.; Maillot, O.; Tortuel, D.; Azuama, C.O.; et al. The absence of SigX results in impaired carbon metabolism and membrane fluidity in Pseudomonas aeruginosa. Sci. Rep. 2018, 8, 17212. [Google Scholar] [CrossRef]
- Clamens, T.; Rosay, T.; Crépin, A.; Grandjean, T.; Kentache, T.; Hardouin, J.; Bortolotti, P.; Neidig, A.; Mooij, M.; Hillion, M.; et al. The aliphatic amidase AmiE is involved in regulation of Pseudomonas aeruginosa virulence. Sci. Rep. 2017, 7, 41178. [Google Scholar] [CrossRef]
- Macé, C.; Seyer, D.; Chemani, C.; Cosette, P.; Di-Martino, P.; Guery, B.; Filloux, A.; Fontaine, M.; Molle, V.; Junter, G.-A.; et al. Identification of biofilm-associated cluster (bac) in Pseudomonas aeruginosa involved in biofilm formation and virulence. PLoS ONE 2008, 3, e3897. [Google Scholar] [CrossRef]
- Vilain, S.; Cosette, P.; Hubert, M.; Lange, C.; Junter, G.-A.; Jouenne, T. Comparative proteomic analysis of planktonic and immobilized Pseudomonas aeruginosa cells: A multivariate statistical approach. Anal. Biochem. 2004, 329, 120–130. [Google Scholar] [CrossRef]
- Armengaud, J.; Christie-Oleza, J.A.; Clair, G.; Malard, V.; Duport, C. Exoproteomics: Exploring the world around biological systems. Expert Rev. Proteom. 2012, 9, 561–575. [Google Scholar] [CrossRef] [PubMed]
- Brown, K.J.; Formolo, C.A.; Seol, H.; Marathi, R.L.; Duguez, S.; An, E.; Pillai, D.; Nazarian, J.; Rood, B.R.; Hathout, Y. Advances in the proteomic investigation of the cell secretome. Expert Rev. Proteom. 2012, 9, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Desvaux, M.; Hébraud, M.; Talon, R.; Henderson, I.R. Secretion and subcellular localizations of bacterial proteins: A semantic awareness issue. Trends Microbiol. 2009, 17, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Maffei, B.; Francetic, O.; Subtil, A. Tracking proteins secreted by bacteria: What’s in the toolbox? Front. Cell. Infect. Microbiol. 2017, 7, 221. [Google Scholar] [CrossRef]
- Burstein, D.; Satanower, S.; Simovitch, M.; Belnik, Y.; Zehavi, M.; Yerushalmi, G.; Ben-Aroya, S.; Pupko, T.; Banin, E. Novel type III effectors in Pseudomonas aeruginosa. mBio 2015, 6, e00161-15. [Google Scholar] [CrossRef]
- Fritsch, M.; Trunk, K.; Diniz, J.A.; Guo, M.; Trost, M.; Coulthurst, S.J. Proteomic identification of novel secreted antibacterial toxins of the Serratia marcescens type VI secretion system. Mol. Cell. Proteom. 2013, 12, 2735–2749. [Google Scholar] [CrossRef]
- Rolsma, S.L.; Frank, D.W. In vitro assays to monitor the activity of Pseudomonas aeruginosa type III secreted proteins. Methods Mol. Biol. 2014, 1149, 171–184. [Google Scholar] [CrossRef]
- Basler, M. Type VI secretion system: Secretion by a contractile nanomachine. Philos. Trans. R. Soc. B 2015, 370, 20150021. [Google Scholar] [CrossRef]
- Nouwens, A.S.; Willcox, M.D.P.; Walsh, B.J.; Cordwell, S.J. Proteomic comparison of membrane and extracellular proteins from invasive (PAO1) and cytotoxic (6206) strains of Pseudomonas aeruginosa. Proteomics 2002, 2, 1325–1346. [Google Scholar] [CrossRef]
- Wehmhöner, D.; Häussler, S.; Tümmler, B.; Jänsch, L.; Bredenbruch, F.; Wehland, J.; Steinmetz, I. Inter- and intraclonal diversity of the Pseudomonas aeruginosa proteome manifests within the secretome J. Bacteriol. 2003, 185. [Google Scholar] [CrossRef]
- Tan, A.S.P.; Worobec, E.A. Isolation and characterization of two immunochemically distinct alkaline phosphatases from Pseudomonas aeruginosa. FEMS Microbiol. Lett. 1993, 106, 281–286. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Arevalo-Ferro, C.; Hentzer, M.; Reil, G.; Görg, A.; Kjelleberg, S.; Givskov, M.; Riedel, K.; Eberl, L. Identification of quorum-sensing regulated proteins in the opportunistic pathogen Pseudomonas aeruginosa by proteomics. Environ. Microbiol. 2003, 5, 1350–1369. [Google Scholar] [CrossRef] [PubMed]
- Kropinski, A.M.B.; Lewis, V.; Berry, D. Effect of growth temperature on the lipids, outer membrane proteins, and lipopolysaccharides of Pseudomonas aeruginosa PAO. J. Bacteriol. 1987, 169, 1960–1966. [Google Scholar] [CrossRef]
- Barbier, M.; Damron, F.H.; Bielecki, P.; Suárez-Diez, M.; Puchałka, J.; Alberti, S.; Martins dos Santos, V.; Goldberg, J.B. From the environment to the host: Re-wiring of the transcriptome of Pseudomonas aeruginosa from 22 °C to 37 °C. PLoS ONE 2014, 9, e89941. [Google Scholar] [CrossRef]
- Termine, E.; Michel, G.P.F. Transcriptome and secretome analyses of the adaptative response of Pseudomonas aeruginosa to suboptimal growth temperature. Int. Microbiol. 2009, 12, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Scott, N.E.; Hare, N.J.; White, M.Y.; Manos, J.; Cordwell, S.J. Secretome of transmissible Pseudomonas aeruginosa AES-1R grown in a cystic fibrosis lung-Like environment J. Proteome Res. 2013, 12, 5357–5369. [Google Scholar] [CrossRef] [PubMed]
- Nouwens, A.S.; Beatson, S.A.; Whitchurch, C.B.; Walsh, B.J.; Schweizer, H.P.; Mattick, J.S.; Cordwell, S.J. Proteome analysis of extracellular proteins regulated by the las and rhl quorum sensing systems in Pseudomonas aeruginosa PAO1. Microbioly 2003, 149, 1311–1322. [Google Scholar] [CrossRef]
- Lampaki, D.; Diepold, A.; Glatter, T. A serial sample processing strategy with improved performance for in-depth quantitative analysis of type III secretion events in Pseudomonas aeruginosa. J. Proteome Res. 2020, 19, 543–553. [Google Scholar] [CrossRef]
- Gaviard, C.; Jouenne, T.; Hardouin, J. Proteomics of Pseudomonas aeruginosa: The increasing role of post-translational modifications. Expert Rev. Proteom. 2018, 15, 757–772. [Google Scholar] [CrossRef]
- Pérez-Llarena, F.J.; Bou, G. Proteomics as a tool for studying bacterial virulence and antimicrobial resistance. Front. Microbiol. 2016, 7, 410. [Google Scholar] [CrossRef]
- Lermyte, F.; Tsybin, Y.O.; O’Connor, P.B.; Loo, J.A. Top or middle? Up or down? Toward a standard lexicon for protein top-down and allied mass spectrometry approaches. J. Am. Soc. Mass Spectrom. 2019, 30, 1149–1157. [Google Scholar] [CrossRef] [PubMed]
- Typas, A.; Sourjik, V. Bacterial protein networks: Properties and functions. Nat. Rev. Microbiol. 2015, 13, 559–572. [Google Scholar] [CrossRef]
- Silverman, J.M.; Agnello, D.M.; Zheng, H.; Andrews, B.T.; Li, M.; Catalano, C.E.; Gonen, T.; Mougous, J.D. Haemolysin co-regulated protein is an exported receptor and chaperone of type VI secretion substrates. Mol. Cell 2013, 51, 584–593. [Google Scholar] [CrossRef] [PubMed]
- Nicod, C.; Banaei-Esfahani, A.; Collins, B.C. Elucidation of host-pathogen protein-protein interactions to uncover mechanisms of host cell rewiring. Curr. Opin. Microbiol. 2017, 39, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Nanao, M.; Ricard-Blum, S.; Di Guilmi, A.M.; Lemaire, D.; Lascoux, D.; Chabert, J.; Attree, I.; Dessen, A. Type III secretion proteins PcrV and PcrG from Pseudomonas aeruginosa form a I:I complex through high affinity interactions. BMC Microbiol. 2003, 3, 21. [Google Scholar] [CrossRef] [PubMed]
- Sriramulu, D.D.; Nimtz, M.; Romling, U. Proteome analysis reveals adaptation of Pseudomonas aeruginosa to the cystic fibrosis lung environment. Proteomics 2005, 5, 3712–3721. [Google Scholar] [CrossRef]
- Shankar, S.; Kavanaugh-Black, A.; Kamath, S.; Chakrabarty, A.M. Characterization of a phosphoprotein phosphatase for the phosphorylated form of nucleoside-diphosphate kinase from Pseudomonas aeruginosa. J. Biol. Chem. 1995, 270, 28246–28250. [Google Scholar] [CrossRef]
- Kamath, K.S.; Kumar, S.S.; Kaur, J.; Venkatakrishnan, V.; Paulsen, I.T.; Nevalainen, H.; Molloy, M.P. Proteomics of hosts and pathogens in cystic fibrosis. Proteom. Clin. Appl. 2015, 9, 134–146. [Google Scholar] [CrossRef]
- Spooner, R.; Yilmaz, Ö. Nucleoside-diphosphate-kinase: A pleiotropic effector in microbial colonization under interdisciplinary characterization. Microbes Infect. 2012, 14, 228–237. [Google Scholar] [CrossRef]
- Kim, Y.-J.; Lee, J.-H.; Lee, Y.; Jingyue, J.; Paek, S.-H.; Kim, H.-B.; Jin, S.; Ha, U.-H. Nucleoside diphosphate kinase and flagellin from Pseudomonas aeruginosa induce interleukin 1 Expression via the Akt/NF-κB signaling pathways. Infect. Immun. 2014, 82, 3252–3260. [Google Scholar] [CrossRef]
- Needl, D.; Jin, Y.; Bichsel, C.; Jia, J.; Guo, J.; Bai, F.; Wu, W.; Ha, U.-H.; Terada, N.; Jin, S. Pseudomonas aeruginosa injects NDK into host cells through a type III secretion system. Microbiology 2014, 160, 1417–1426. [Google Scholar] [CrossRef]
- Ouidir, T.; Jarnier, F.; Cosette, P.; Jouenne, T.; Hardouin, J. Extracellular Ser/Thr/Tyr phosphorylated proteins of Pseudomonas aeruginosa PA14 strain. Proteomics 2014, 14, 2017–2030. [Google Scholar] [CrossRef] [PubMed]
- Meleady, P. Two-Dimensional Gel Electrophoresis and 2D-DIGE. In Difference Gel Electrophoresis; Ohlendieck, K., Ed.; Humana Press: New York, NY, USA, 2018; Volume 1664, pp. 3–14. [Google Scholar]
- Ball, G.; Antelmann, H.; Imbert, P.R.C.; Gimenez, M.R.; Voulhoux, R.; Ize, B. Contribution of the twin arginine translocation system to the exoproteome of Pseudomonas aeruginosa. Sci. Rep. 2016, 6, 27675. [Google Scholar] [CrossRef] [PubMed]
- Hood, R.D.; Singh, P.; Hsu, F.; Güvener, T.; Carl, M.A.; Trinidad, R.R.S.; Silverman, J.M.; Ohlson, B.B.; Hicks, K.G.; Plemel, R.L.; et al. A type VI secretion system of Pseudomonas aeruginosa targets a toxin to bacteria. Cell Host Microbe 2010, 7, 25–37. [Google Scholar] [CrossRef]
- Lin, L.; Lezan, E.; Schmidt, A.; Basler, M. Abundance of bacterial Type VI secretion system components measured by targeted proteomics. Nat. Commun. 2019, 10, 2584. [Google Scholar] [CrossRef]
- Bergamini, G.; Di Silvestre, D.; Mauri, P.; Cigana, C.; Bragonzi, A.; De Palma, A.; Benazzi, L.; Döring, G.; Assael, B.M.; Melotti, P.; et al. MudPIT analysis of released proteins in Pseudomonas aeruginosa laboratory and clinical strains in relation to pro-inflammatory effects. Integr. Biol. 2012, 4, 270–279. [Google Scholar] [CrossRef]
- Hickey, C.; Schaible, B.; Nguyen, S.; Hurley, D.; Srikumar, S.; Fanning, S.; Brown, E.; Crifo, B.; Matallanas, D.; McClean, S.; et al. Increased virulence of bloodstream over peripheral isolates of P. aeruginosa identified through post-transcriptional regulation of virulence factors. Front. Cell. Infect. Microbiol. 2018, 8, 357. [Google Scholar] [CrossRef]
- Casilag, F.; Lorenz, A.; Krueger, J.; Klawonn, F.; Weiss, S.; Häussler, S. The LasB elastase of Pseudomonas aeruginosa acts in concert with alkaline protease AprA to prevent flagellin-mediated immune recognition. Infect. Immun. 2016, 84, 162–171. [Google Scholar] [CrossRef]
- Armengaud, J.; Bland, C.; Christie-Oleza, J.; Miotello, G. Microbial proteogenomics, gaining ground with the avalanche of genome sequences. J. Bacteriol. Parasitol. 2011, S3-001. [Google Scholar] [CrossRef]
- Jarocki, V.M.; Tacchi, J.L.; Djordjevic, S.P. Non-proteolytic functions of microbial proteases increase pathological complexity. Proteomics 2015, 15, 1075–1088. [Google Scholar] [CrossRef]
- Jeffery, C. The Use of Proteomics Studies in Identifying Moonlighting Proteins. In Functional Proteomics; Wang, X., Kuruc, M., Eds.; Humana Press: New York, NY, USA, 2019; Volume 1871, pp. 437–443. [Google Scholar]
- Wang, W.; Jeffery, C.J. An analysis of surface proteomics results reveals novel candidates for intracellular/surface moonlighting proteins in bacteria. Mol. Biosyst. 2016, 12, 1420–1431. [Google Scholar] [CrossRef] [PubMed]
- Shinagawa, M.; Kobayashi, D.; Goto, M.; Tanaka, M.; Kuribayashi, K.; Yanagihara, N.; Watanabe, N. Identification of a bacteriolysis-associated virulence factor against lung epithelial cells in Pseudomonas aeruginosa PAO-1 cell lysate. Microb. Pathog. 2014, 74, 35–40. [Google Scholar] [CrossRef] [PubMed]
- O’Callaghan, R.; Caballero, A.; Tang, A.; Bierdeman, M. Pseudomonas aeruginosa keratitis: Protease IV and PASP as corneal virulence mediators. Microorganisms 2019, 7, 281. [Google Scholar] [CrossRef] [PubMed]
- Ribet, D.; Cossart, P. Post-translational modifications in host cells during bacterial infection. FEBS Lett. 2010, 584, 2748–2758. [Google Scholar] [CrossRef] [PubMed]
- Pompeo, F.; Foulquier, E.; Serrano, B.; Grangeasse, C.; Galinier, A. Phosphorylation of the cell division protein GpsB regulates PrkC kinase activity through a negative feedback loop in Bacillus subtilis. Mol. Microbiol. 2015, 97, 139–150. [Google Scholar] [CrossRef]
- Hentchel, K.L.; Escalante-Semerena, J. Acylation of biomolecules in prokaryotes: A widespread strategy for the control of biological function and metabolic stress. Microbiol. Mol. Biol. Rev. 2015, 79, 321–346. [Google Scholar] [CrossRef]
- Libby, E.A.; Reuveni, S.; Dworkin, J. Multisite phosphorylation drives phenotypic variation in (p)ppGpp synthetase-dependent antibiotic tolerance. Nat. Commun. 2019, 10, 5133. [Google Scholar] [CrossRef]
- Liu, F.; Yang, M.; Wang, X.; Yang, S.; Gu, J.; Zhou, J.; Zhang, X.-E.; Deng, J.; Ge, F. Acetylome analysis reveals diverse functions of lysine acetylation in Mycobacterium tuberculosis. Mol. Cell. Proteom. 2014, 13, 3352–3366. [Google Scholar] [CrossRef]
- Khan, M.Z.; Bhaskar, A.; Upadhyay, S.; Kumari, P.; Rajmani, R.S.; Jain, P.; Singh, A.; Kumar, D.; Bhavesh, N.S.; Nandicoori, V.K. Protein kinase G confers survival advantage to Mycobacterium tuberculosis during latency-like conditions. J. Biol. Chem. 2017, 292, 16093–16108. [Google Scholar] [CrossRef]
- Ren, J.; Sang, Y.; Tan, Y.; Tao, J.; Ni, J.; Liu, S.; Fan, X.; Zhao, W.; Lu, J.; Wu, W.; et al. Acetylation of lysine 201 inhibits the DNA-binding ability of PhoP to regulate Salmonella virulence. PLoS Pathog. 2016, 12, e1005458. [Google Scholar] [CrossRef]
- Bielecki, P.; Glik, J.; Kawecki, M.; Martins dos Santos, V.A.P. Towards understanding Pseudomonas aeruginosa burn wound infections by profiling gene expression. Biotechnol. Lett. 2008, 30, 777–790. [Google Scholar] [CrossRef] [PubMed]
- Ouidir, T.; Jouenne, T.; Hardouin, J. Post-translational modifications in Pseudomonas aeruginosa revolutionized by proteomic analysis. Biochimie 2016, 125, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Ravichandran, A.; Sugiyama, N.; Tomita, M.; Swarup, S.; Ishihama, Y. Ser/Thr/Tyr phosphoproteome analysis of pathogenic and non-pathogenic Pseudomonas species. Proteomics 2009, 9, 2764–2775. [Google Scholar] [CrossRef] [PubMed]
- Ouidir, T.; Cosette, P.; Jouenne, T.; Hardouin, J. Proteomic profiling of lysine acetylation in Pseudomonas aeruginosa reveals the diversity of acetylated proteins. Proteomics 2015, 15, 2152–2157. [Google Scholar] [CrossRef] [PubMed]
- Gaviard, C.; Broutin, I.; Cosette, P.; Dé, E.; Jouenne, T.; Hardouin, J. Lysine succinylation and acetylation in Pseudomonas aeruginosa. J. Proteome Res. 2018, 17, 2449–2459. [Google Scholar] [CrossRef] [PubMed]
- Tran, B.Q.; Barton, C.; Feng, J.; Sandjong, A.; Yoon, S.H.; Awasthi, S.; Liang, T.; Khan, M.M.; Kilgour, D.P.A.; Goodlett, D.R.; et al. Glycosylation characterization of therapeutic mAbs by top- and middle-down mass spectrometry. Data Brief 2015, 6, 68–76. [Google Scholar] [CrossRef]
- Chamot-Rooke, J.; Mikaty, G.; Malosse, C.; Soyer, M.; Dumont, A.; Gault, J.; Imhaus, A.-F.; Martin, P.; Trellet, M.; Clary, G.; et al. Posttranslational modification of pili upon cell contact triggers N. meningitidis dissemination. Science 2011, 331, 778–782. [Google Scholar] [CrossRef]
- Jin, Y.; Diffee, G.M.; Colman, R.J.; Anderson, R.M.; Ge, Y. Top-down mass spectrometry of sarcomeric protein post-translational modifications from non-human primate skeletal muscle. J. Am. Soc. Mass Spectrom. 2019, 30, 2460–2469. [Google Scholar] [CrossRef]
- Miao, E.A.; Mao, D.P.; Yudkovsky, N.; Bonneau, R.; Lorang, C.G.; Warren, S.E.; Leaf, I.A.; Aderem, A. Innate immune detection of the type III secretion apparatus through the NLRC4 inflammasome. Proc. Natl. Acad. Sci. USA 2010, 107, 3076–3080. [Google Scholar] [CrossRef]
- Suriyanarayanan, T.; Periasamy, S.; Lin, M.-H.; Ishihama, Y.; Swarup, S. Flagellin FliC phosphorylation affects type 2 protease secretion and biofilm dispersal in Pseudomonas aeruginosa PAO1. PLoS ONE 2016, 11, e0164155. [Google Scholar] [CrossRef]
- Kelly-Wintenberg, K.; South, S.L.; Montie, T.C. Tyrosine phosphate in a- and b-type flagellins of Pseudomonas aeruginosa. J. Bacteriol. 1993, 175, 2458–2461. [Google Scholar] [CrossRef][Green Version]
- Schirm, M.; Arora, S.K.; Verma, A.; Vinogradov, E.; Thibault, P.; Ramphal, R.; Logan, S.M. Structural and genetic characterization of glycosylation of type a flagellin in Pseudomonas aeruginosa. J. Bacteriol. 2004, 186, 2523–2531. [Google Scholar] [CrossRef] [PubMed]
- Brimer, C.D.; Montie, T.C. Cloning and comparison of fliC genes and identification of glycosylation in the flagellin of Pseudomonas aeruginosa a-type strains. J. Bacteriol. 1998, 180, 3209–3217. [Google Scholar] [CrossRef]
- Verma, A.; Schirm, M.; Arora, S.K.; Thibault, P.; Logan, S.M.; Ramphal, R. Glycosylation of b-type flagellin of Pseudomonas aeruginosa: Structural and genetic basis. J. Bacteriol. 2006, 188, 4395–4403. [Google Scholar] [CrossRef][Green Version]
- Mougous, J.D.; Gifford, C.A.; Ramsdell, T.L.; Mekalanos, J.J. Threonine phosphorylation post-translationally regulates protein secretion in Pseudomonas aeruginosa. Nat. Cell Biol. 2007, 9, 797–803. [Google Scholar] [CrossRef] [PubMed]
- Hsu, F.; Schwarz, S.; Mougous, J.D. TagR promotes PpkA-catalyzed type VI secretion activation in Pseudomonas aeruginosa. Mol. Microbiol. 2009, 75, 1111–1125. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Barbieri, J.T. Pseudomonas aeruginosa ExoT ADP-ribosylates CT10 regulator of kinase (Crk) proteins. J. Biol. Chem. 2003, 278, 32794–32800. [Google Scholar] [CrossRef]
- UniProt Consortium. UniProt: A hub for protein information. Nucleic Acids Res. 2015, 43, D204–D212. [Google Scholar] [CrossRef] [PubMed]
- Fulton, K.M.; Baltat, I.; Twine, S.M. Immunoproteomics Methods and Techniques. In Immunoproteomics, 2nd ed.; Fulton, K.M., Twine, S.M., Eds.; Humana: New York, NY, USA, 2019; Volume 2024, pp. 22–58. [Google Scholar]
- Upritchard, H.G.; Cordwell, S.J.; Lamont, I.L. Immunoproteomics to examine cystic fibrosis host interactions with extracellular Pseudomonas aeruginosa proteins. Infect. Immun. 2008, 76, 4624–4632. [Google Scholar] [CrossRef][Green Version]
- Dennehy, R.; McClean, S. Immunoproteomics: The key to discovery of new vaccine antigens against bacterial respiratory infections. Curr. Protein Pept. Sci. 2012, 13, 807–815. [Google Scholar] [CrossRef]
- Bastaert, F.; Kheir, S.; Saint-Criq, V.; Villeret, B.; Dang, P.M.-C.; El-Benna, J.; Sirard, J.-C.; Voulhoux, R.; Sallenave, J.-M. Pseudomonas aeruginosa LasB subverts alveolar macrophage activity by interfering with bacterial killing through downregulation of innate immune defense, reactive oxygen species generation, and complement activation. Front. Immunol. 2018, 9, 1675. [Google Scholar] [CrossRef] [PubMed]
- Vigneshkumar, B.; Durai, S.; Kundu, S.; Balamurugan, K. Proteome analysis reveals translational inhibition of Caenorhabditis elegans enhances susceptibility to Pseudomonas aeruginosa PAO1 pathogenesis. J. Proteom. 2016, 145, 141–152. [Google Scholar] [CrossRef]
- Díaz-Pascual, F.; Ortíz-Severín, J.; Varas, M.A.; Allende, M.L.; Chávez, F.P. In vivo host-pathogen interactions as revealed by global proteomic profiling of zebrafish larvae. Front. Cell. Infect. Microbiol. 2017, 7, 334. [Google Scholar] [CrossRef] [PubMed]
- Llamas, M.A.; van der Sar, A.M. Assessing Pseudomonas Virulence with Nonmammalian Host: Zebrafish. In Pseudomonas Methods and Protocols; Filloux, A., Ramos, J.-L., Eds.; Humana Press: New York, NY, USA, 2014; Volume 1149, pp. 709–721. [Google Scholar]
- Koppenol-Raab, M.; Sjoelund, V.; Manes, N.P.; Gottschalk, R.A.; Dutta, B.; Benet, Z.L.; Fraser, I.D.C.; Nita-Lazar, A. Proteome and secretome analysis reveals differential post-transcriptional regulation of toll-like receptor responses. Mol. Cell. Proteom. 2017, 16, 172–186. [Google Scholar] [CrossRef] [PubMed]
- Yeung, J.; Gadjeva, M.; Geddes-McAlister, J. Label-free quantitative proteomics distinguishes general and site-specific host responses to Pseudomonas aeruginosa infection at the ocular surface. Proteomics 2020, 20, e1900290. [Google Scholar] [CrossRef]
- Millette, G.; Langlois, J.-P.; Brouillette, E.; Frost, E.H.; Cantin, A.M.; Malouin, F. Despite antagonism in vitro, Pseudomonas aeruginosa enhances Staphylococcus aureus colonization in a murine lung infection model. Front. Microbiol. 2019, 10, 2880. [Google Scholar] [CrossRef]
- Serra, R.; Grande, R.; Butrico, L.; Rossi, A.; Settimio, F.U.; Caroleo, B.; Amato, B.; Gallelli, L.; de Franciscis, S. Chronic wound infections: The role of Pseudomonas aeruginosa and Staphylococcus aureus. Expert Rev. Anti Infect. Ther. 2015, 13, 605–613. [Google Scholar] [CrossRef]
- Pallett, R.; Leslie, L.J.; Lambert, P.A.; Milic, I.; Devitt, A.; Marshall, L.J. Anaerobiosis influences virulence properties of Pseudomonas aeruginosa cystic fibrosis isolates and the interaction with Staphylococcus aureus. Sci. Rep. 2019, 9, 6748. [Google Scholar] [CrossRef]
- Tielen, P.; Rosenau, F.; Wilhelm, S.; Jaeger, K.-E.; Flemming, H.-C.; Wingender, J. Extracellular enzymes affect biofilm formation of mucoid Pseudomonas aeruginosa. Microbiology 2010, 156, 2239–2252. [Google Scholar] [CrossRef]
- Purschke, F.G.; Hiller, E.; Trick, I.; Rupp, S. Flexible survival strategies of Pseudomonas aeruginosa in biofilms result in increased fitness compared to Candida albicans. Mol. Cell. Proteom. 2012, 11, 1652–1669. [Google Scholar] [CrossRef]
- Galle, M.; Jin, S.; Bogaert, P.; Haegman, M.; Vandenabeele, P.; Beyaert, R. The Pseudomonas aeruginosa type III secretion system has an exotoxin S/T/Y independent pathogenic role during acute lung infection. PLoS ONE 2012, 7, e41547. [Google Scholar] [CrossRef][Green Version]
- Bartels, K.-M.; Funken, H.; Knapp, A.; Brocker, M.; Bott, M.; Wilhelm, S.; Jaeger, K.-E.; Rosenau, F. Glycosylation is required for outer membrane localization of the lectin LecB in Pseudomonas aeruginosa. J. Bacteriol. 2011, 193, 1107–1113. [Google Scholar] [CrossRef] [PubMed]
- Beltran, P.M.J.; Federspiel, J.D.; Sheng, X.; Cristea, I.M. Proteomics and integrative omic approaches for understanding host-pathogen interactions and infectious diseases. Mol. Syst. Biol. 2017, 13, 922. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, R.; Thorsell, A.; Gomila, M.; Salvà-Serra, F.; Jakobsson, H.E.; Gonzales-Siles, L.; Jaén-Luchoro, D.; Skovbjerg, S.; Fuchs, J.; Karlsson, A.; et al. Discovery of species-unique peptide biomarkers of bacterial pathogens by tandem mass spectrometry-based proteotyping. Mol. Cell. Proteom. 2020, 19, 518–528. [Google Scholar] [CrossRef] [PubMed]
- Jagusztyn-Krynicka, E.K.; Dadlez, M.; Grabowska, A.; Roszczenko, P. Proteomic technology in the design of new effective antibacterial vaccines. Expert Rev. Proteom. 2009, 6, 315–330. [Google Scholar] [CrossRef] [PubMed]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sauvage, S.; Hardouin, J. Exoproteomics for Better Understanding Pseudomonas aeruginosa Virulence. Toxins 2020, 12, 571. https://doi.org/10.3390/toxins12090571
Sauvage S, Hardouin J. Exoproteomics for Better Understanding Pseudomonas aeruginosa Virulence. Toxins. 2020; 12(9):571. https://doi.org/10.3390/toxins12090571
Chicago/Turabian StyleSauvage, Salomé, and Julie Hardouin. 2020. "Exoproteomics for Better Understanding Pseudomonas aeruginosa Virulence" Toxins 12, no. 9: 571. https://doi.org/10.3390/toxins12090571
APA StyleSauvage, S., & Hardouin, J. (2020). Exoproteomics for Better Understanding Pseudomonas aeruginosa Virulence. Toxins, 12(9), 571. https://doi.org/10.3390/toxins12090571