Bitis arietans Snake Venom and Kn-Ba, a Snake Venom Serine Protease, Induce the Production of Inflammatory Mediators in THP-1 Macrophages

 , and
, and 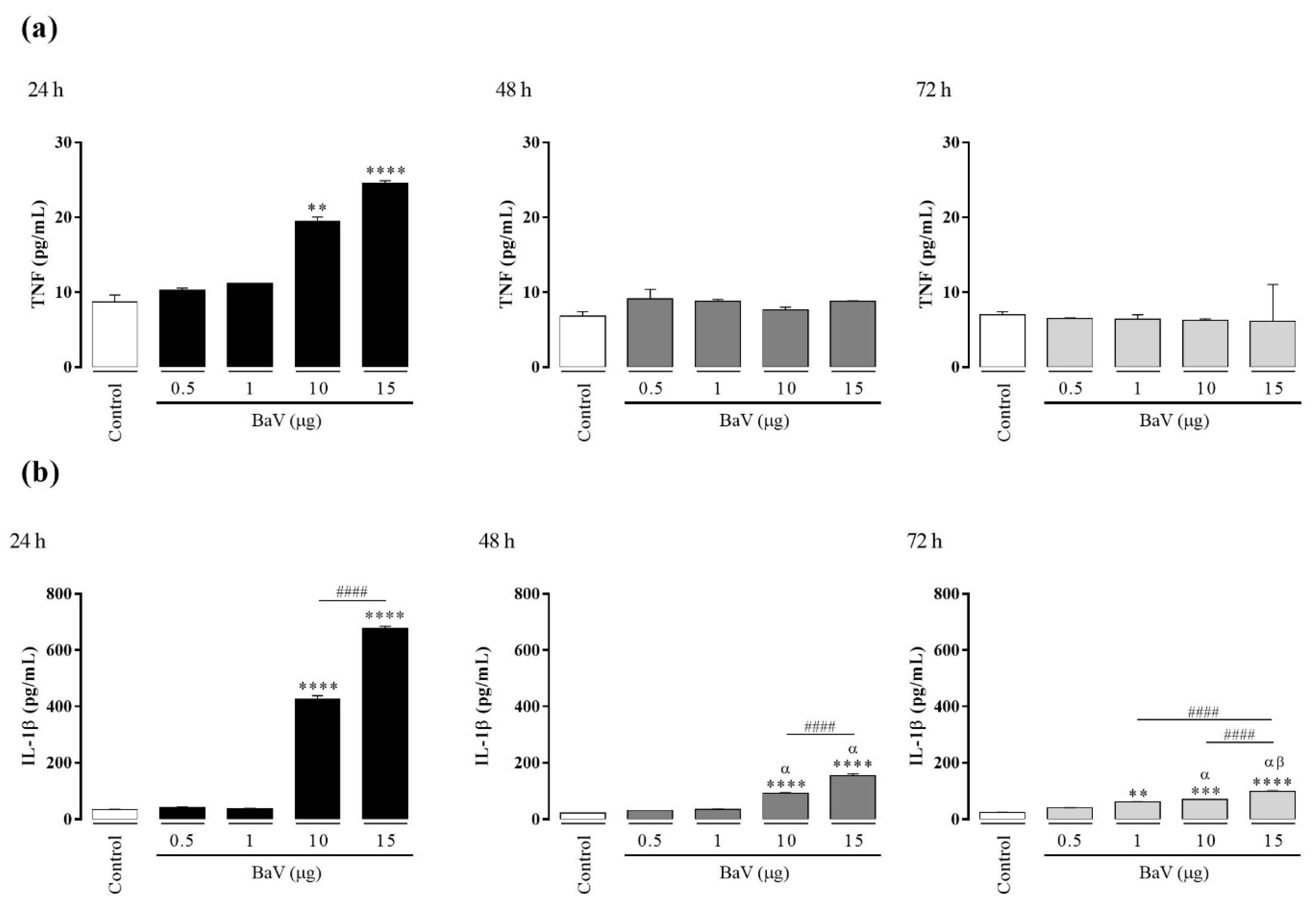{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. THP-1 Macrophage Differentiation
2.2. Total Protein and Endotoxin Contents Determination in BaV and Kn-Ba
2.3. Effects of BaV and Kn-Ba on THP-1 Macrophage Viabilities
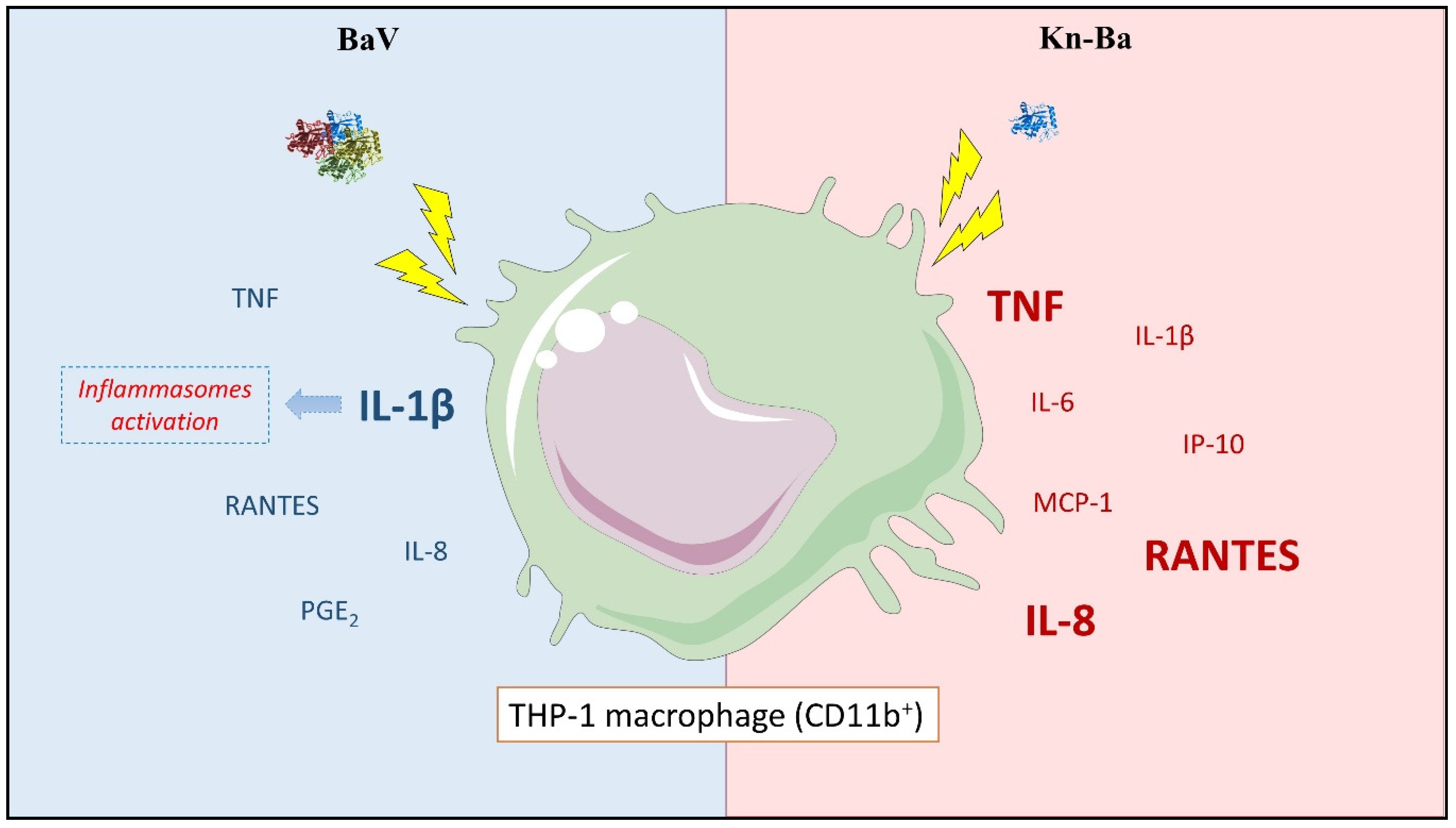
2.4. BaV Induces the Production of TNF and IL-1β in THP-1 Macrophages
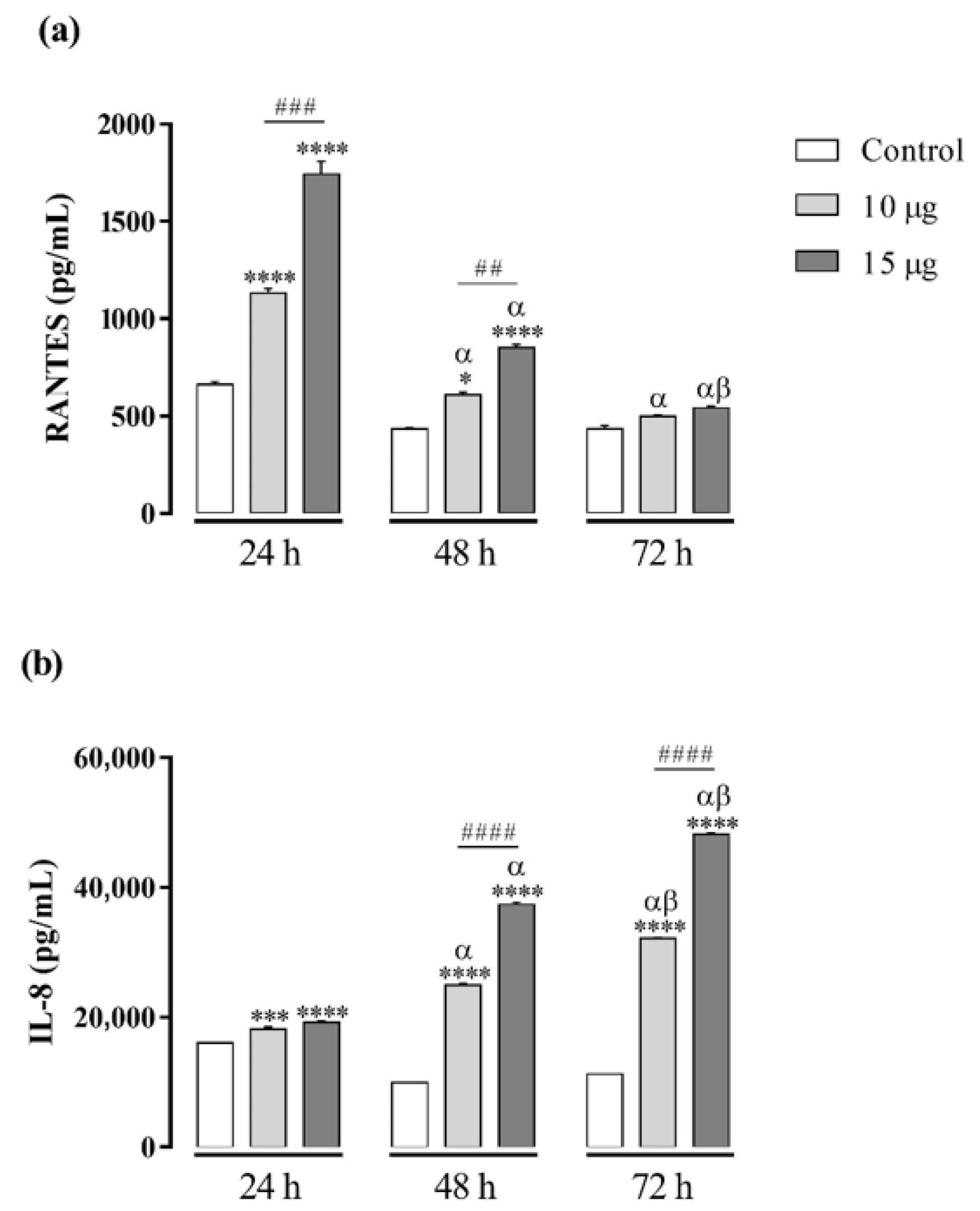
2.5. BaV Induces the Production of RANTES and IL-8 in THP-1 Macrophages
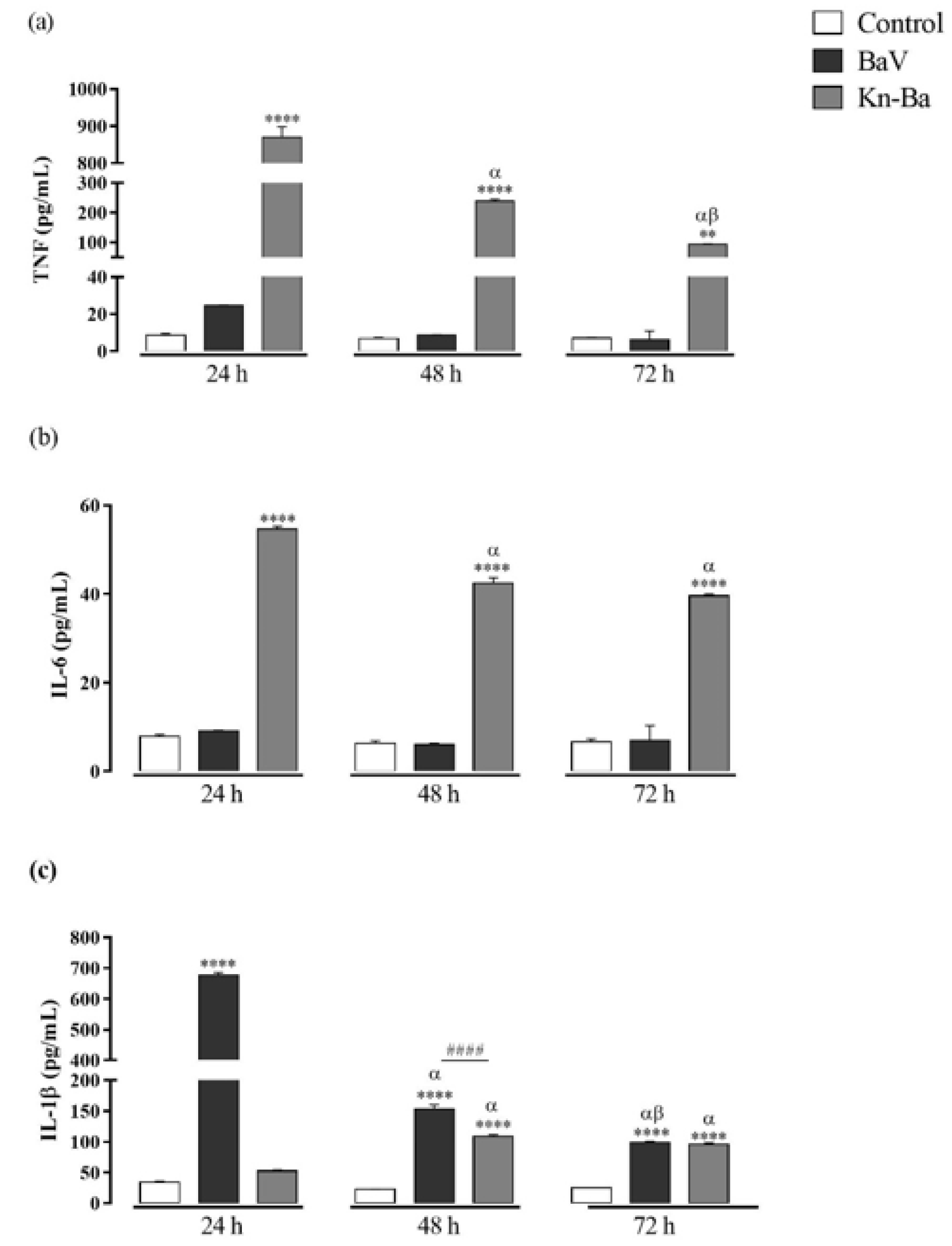
2.6. Kn-Ba Induces the Production of TNF, IL-6, and IL-1β in THP-1 Macrophages
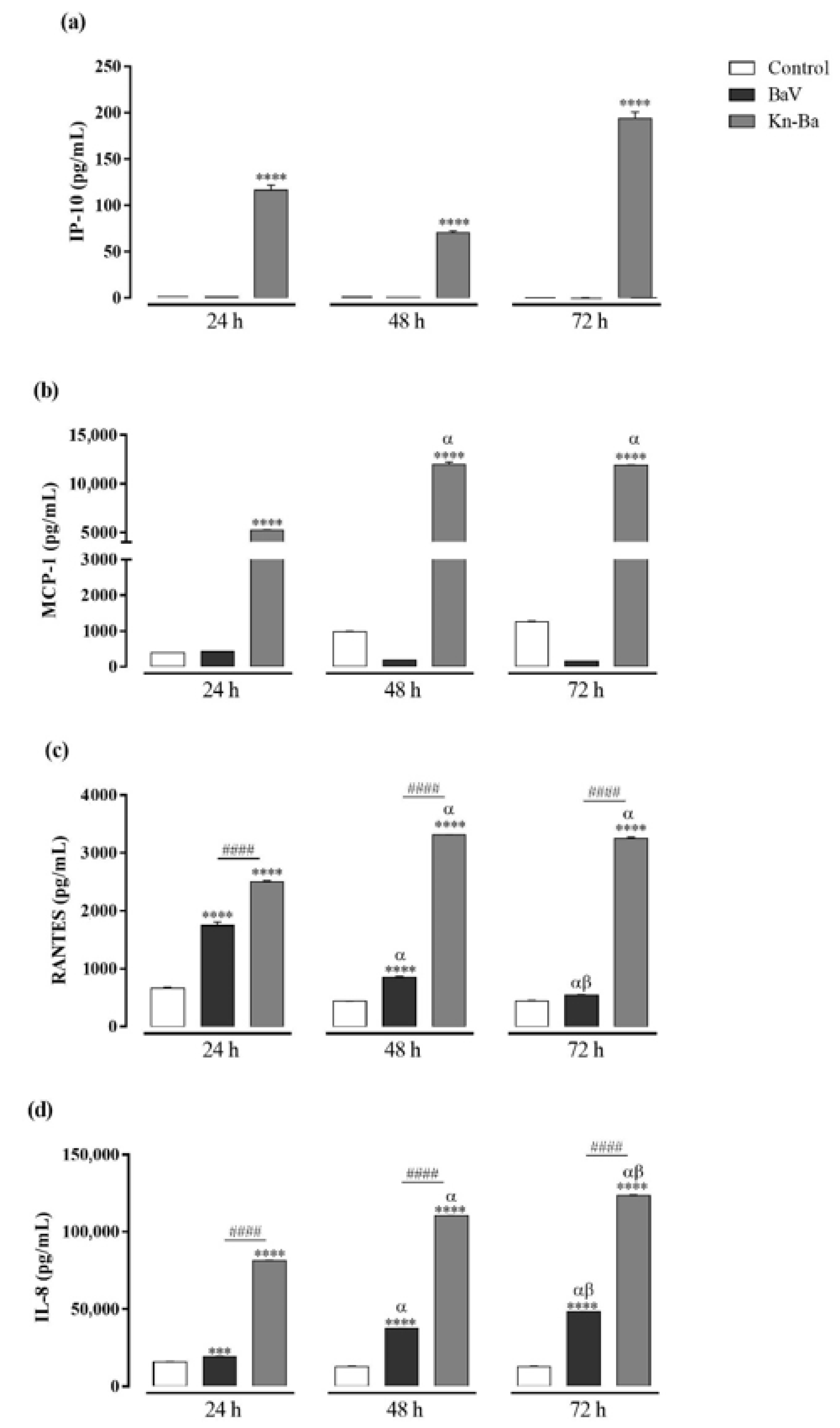
2.7. Kn-Ba Induces the Production of IP-10, MCP-1, RANTES, and IL-8 in THP-1 Macrophages
2.8. Kn-Ba Induced High Levels of TNF and IL-6, Whereas BaV Is Involved in the IL-1β Production in THP-1 Macrophages
2.9. Kn-Ba Induced the Production of Highest Levels of all Evaluated Chemokines in THP-1 Macrophages
2.10. BaV, But Not Kn-Ba, Induced the Production of PGE2
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. BaV
5.2. Kn-Ba
5.3. Total Protein Quantification
5.4. Endotoxin Contents Determination
5.5. Human THP-1 Pre-Monocyte Culture
5.6. Human THP-1 Pre-Monocyte Differentiation into THP-1 Macrophages
5.7. CD11b Expression in Differentiated THP-1 Macrophages
5.8. Incubation of THP-1 Macrophages with BaV and Kn-Ba
5.9. Release of Lactate Dehydrogenase (LDH)
5.10. Quantification of Cytokines and Chemokines Produced by THP-1 Macrophages
5.11. Quantification of Lipid Inflammatory Mediators Produced by THP-1 Macrophages
5.12. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Kasturiratne, A.; Wickremasinghe, A.R.; de Silva, N.; Gunawardena, N.K.; Pathmeswaran, A.; Premaratna, R.; Savioli, L.; Lalloo, D.G.; de Silva, H.J. The global burden of snakebite: A literature analysis and modelling based on regional estimates of envenoming and deaths. PLoS Med. 2008, 5, 1591–1604. [Google Scholar] [CrossRef] [Green Version]
- Chippaux, J.P. Estimate of the burden of snakebites in sub-Saharan Africa: A meta-analytic approach. Toxicon 2011, 57, 586–599. [Google Scholar] [CrossRef] [PubMed]
- Swaroop, S.; Grab, B. Snakebite mortality in the world. Bull. World Health Organ. 1954, 10, 35–76. [Google Scholar] [PubMed]
- WHO. Guidelines for the Production, Control and Regulation of Snake Antivenom Immunoglobulins; World Health Organization: Geneva, Switzerland, 2016; pp. 1–146. [Google Scholar]
- Shupe, S. Venomous Snakes of the World: A Manual for Use by US Amphibious Forces; Skyhorse Publishing Inc.: New York, NY, USA, 2013. [Google Scholar]
- Shupe, S. Generic and Species Descriptions; Skyhorse Publishing: New York, NY, USA, 2013. [Google Scholar]
- WHO. Guidelines for the Prevention and Clinical Management of Snakebite in Africa; World Health Organization: Brazzaville, Congo, 2010; pp. 1–145. [Google Scholar]
- Currier, R.B.; Harrison, R.A.; Rowley, P.D.; Laing, G.D.; Wagstaff, S.C. Intra-specific variation in venom of the African Puff Adder (Bitis arietans): Differential expression and activity of snake venom metalloproteinases (SVMPs). Toxicon 2010, 55, 864–873. [Google Scholar] [CrossRef] [PubMed]
- Warrell, D.A.; Ormerod, L.D.; Davidson, N.M. Bites by puff-adder (Bitis arietans) in Nigeria, and value of antivenom. Br. Med. J. 1975, 4, 697–700. [Google Scholar] [CrossRef] [Green Version]
- Bey, T.A.; Boyer, L.V.; Walter, F.G.; McNally, J.; Desai, H. Exotic snakebite: Envenomation by an African puff adder (Bitis arietans). J. Emerg. Med. 1997, 15, 827–831. [Google Scholar] [CrossRef]
- Langhorn, R.; Persson, F.; Åblad, B.; Goddard, A.; Schoeman, J.P.; Willesen, J.L.; Tarnow, I.; Kjelgaard-Hansen, M. Myocardial injury in dogs with snake envenomation and its relation to systemic inflammation. J. Vet. Emerg. Crit. Care 2014, 24, 174–181. [Google Scholar] [CrossRef] [Green Version]
- Lavonas, E.J.; Tomaszewski, C.A.; Ford, M.D.; Rouse, A.M.; Kerns, W.P., 2nd. Severe puff adder (Bitis arietans) envenomation with coagulopathy. J. Toxicology. Clin. Toxicol. 2002, 40, 911–918. [Google Scholar] [CrossRef]
- Theakston, R.; Wyatt, G. Venom antibody levels in a patient bitten by a young puff adder (Bitis arietans) during a world record attempt. Ann. Trop. Med. Parasitol. 1985, 79, 305–307. [Google Scholar] [CrossRef]
- Markland, F.S. Snake venoms and the hemostatic system. Toxicon 1998, 36, 1749–1800. [Google Scholar] [CrossRef]
- Juarez, P.; Wagstaff, S.C.; Oliver, J.; Sanz, L.; Harrison, R.A.; Calvete, J.J. Molecular cloning of disintegrin-like transcript BA-5A from a Bitis arietans venom gland cDNA library: A putative intermediate in the evolution of the long-chain disintegrin bitistatin. J. Mol. Evol. 2006, 63, 142–152. [Google Scholar] [CrossRef]
- Fasoli, E.; Sanz, L.; Wagstaff, S.; Harrison, R.A.; Righetti, P.G.; Calvete, J.J. Exploring the venom proteome of the African puff adder, Bitis arietans, using a combinatorial peptide ligand library approach at different pHs. J. Proteom. 2010, 73, 932–942. [Google Scholar] [CrossRef] [Green Version]
- Archer, J.; Whiteley, G.; Casewell, N.R.; Harrison, R.A.; Wagstaff, S.C. VTBuilder: A tool for the assembly of multi isoform transcriptomes. BMC Bioinform. 2014, 15, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Kodama, R.T.; Cajado-Carvalho, D.; Kuniyoshi, A.K.; Kitano, E.S.; Tashima, A.K.; Barna, B.F.; Takakura, A.C.; Serrano, S.M.; Dias-Da-Silva, W.; Tambourgi, D.V.; et al. New proline-rich oligopeptides from the venom of African adders: Insights into the hypotensive effect of the venoms. Biochim. Biophys. Acta 2015, 1850, 1180–1187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petretski, J.; Kanashiro, M.; Silva, C.; Alves, E.; Kipnis, T. Two related thrombin-like enzymes present in Bothrops atrox venom. Braz. J. Med Biol. Res. Rev. Bras. Pesqui. Med. E Biol. 2000, 33, 1293–1300. [Google Scholar] [CrossRef] [Green Version]
- Kanashiro, M.M.; Rita de Cássia, M.E.; Petretski, J.H.; Prates, M.V.; Alves, E.W.; Machado, O.L.; da Silva, W.D.; Kipnis, T.L. Biochemical and biological properties of phospholipases A2 from Bothrops atrox snake venom. Biochem. Pharmacol. 2002, 64, 1179–1186. [Google Scholar] [CrossRef]
- Stocker, K.; Barlow, G. The coagulant enzyme from Bothrops atrox venom (batroxobin). Methods Enzymol. 1976, 45, 214–223. [Google Scholar]
- Clissa, P.B.; Laing, G.D.; Theakston, R.D.G.; Mota, I.; Taylor, M.J.; Moura-da-Silva, A.M. The effect of jararhagin, a metalloproteinase from Bothrops jararaca venom, on pro-inflammatory cytokines released by murine peritoneal adherent cells. Toxicon 2001, 39, 1567–1573. [Google Scholar] [CrossRef]
- Costa, E.; Clissa, P.; Teixeira, C.; Moura-da-Silva, A. Importance of metalloproteinases and macrophages in viper snake envenomation–induced local inflammation. Inflammation 2002, 26, 13–17. [Google Scholar] [CrossRef]
- Serrano, S.M.; Sampaio, C.A.; Mentele, R.; Camargo, A.C.; Fink, E. A novel fibrinogen-clotting enzyme, TL-BJ, from the venom of the snake Bothrops jararaca: Purification and characterization. Thromb. Haemost. 2000, 83, 438–444. [Google Scholar]
- Megale, Â.A.A.; Magnoli, F.C.; Kuniyoshi, A.K.; Iwai, L.K.; Tambourgi, D.V.; Portaro, F.C.; da Silva, W.D. Kn-Ba: A novel serine protease isolated from Bitis arietans snake venom with fibrinogenolytic and kinin-releasing activities. J. Venom. Anim. Toxins 2018, 24, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, C.F.; Cury, Y.; Moreira, V.; Picolo, G.; Chaves, F. Inflammation induced by Bothrops asper venom. Toxicon 2009, 54, 67–76. [Google Scholar] [CrossRef]
- Teixeira, C.F.; Fernandes, C.M.; Zuliani, J.P.; Zamuner, S.F. Inflammatory effects of snake venom metalloproteinases. Mem. Do Inst. Oswaldo Cruz 2005, 100, 181–184. [Google Scholar] [CrossRef]
- Voronov, E.; Apte, R.; Sofer, S. The systemic inflammatory response syndrome related to the release of cytokines following severe envenomation. J. Venom. Anim. Toxins 1999, 5, 5–33. [Google Scholar] [CrossRef]
- Szold, O.; Ben-Abraham, R.; Frolkis, I.; Sorkine, M.; Sorkine, P. Tumor necrosis factor as a mediator of cardiac toxicity following snake envenomation. Crit. Care Med. 2003, 31, 1449–1453. [Google Scholar] [CrossRef]
- Megale, Â.A.A.; Portaro, F.C.; Da Silva, W.D. Bitis arietans snake venom induces an inflammatory response which is partially dependent on lipid mediators. Toxins 2020, 12, 594. [Google Scholar] [CrossRef]
- Schwende, H.; Fitzke, E.; Ambs, P.; Dieter, P. Differences in the state of differentiation of THP-1 cells induced by phorbol ester and 1, 25-dihydroxyvitamin D3. J. Leukoc. Biol. 1996, 59, 555–561. [Google Scholar] [CrossRef]
- Mittar, D.; Paramban, R.; McIntyre, C. Flow cytometry and high-content imaging to identify markers of monocyte-macrophage differentiation. J. BD Biosci. 2011, 1, 1–20. [Google Scholar]
- Daigneault, M.; Preston, J.A.; Marriott, H.M.; Whyte, M.K.; Dockrell, D.H. The identification of markers of macrophage differentiation in PMA-stimulated THP-1 cells and monocyte-derived macrophages. PLoS ONE 2010, 5, 1–10. [Google Scholar] [CrossRef]
- Zoccal, K.F. A Peçonha do Escorpião Tityus serrulatus é Reconhecida por Receptores de Reconhecimento Padrão e Induz Ativação Celular e Inflamação. Ph.D. Thesis, Universidade de São Paulo, São Paulo, Brazil, 2014. [Google Scholar]
- Gutiérrez, J.M.; Chaves, F.; Cerdas, L. Inflammatory infiltrate in skeletal muscle injected with Bothrops asper venom. Rev. Biol. Trop. 1986, 34, 209–214. [Google Scholar]
- Zamuner, S.R.; Zuliani, J.P.; Fernandes, C.M.; Gutiérrez, J.M.; Teixeira, C.d.F.P. Inflammation induced by Bothrops asper venom: Release of proinflammatory cytokines and eicosanoids, and role of adhesion molecules in leukocyte infiltration. Toxicon 2005, 46, 806–813. [Google Scholar] [CrossRef] [PubMed]
- Barros, S.; Friedlanskaia, I.; Petricevich, V.; Kipnis, T. Local inflammation, lethality and cytokine release in mice injected with Bothrops atrox venom. Mediat. Inflamm. 1998, 7, 339–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreira, V.; Dos-Santos, M.C.; Nascimento, N.G.; da Silva, H.B.; Fernandes, C.M.; Lima, M.R.D.I.; Teixeira, C. Local inflammatory events induced by Bothrops atrox snake venom and the release of distinct classes of inflammatory mediators. Toxicon 2012, 60, 12–20. [Google Scholar] [CrossRef]
- Farsky, S.H.; Walber, J.; Costa-Cruz, M.; Curry, Y.; Teixeira, C.F. Leukocyte response induced by Bothrops jararaca crude venom: In vivo and in vitro studies. Toxicon 1997, 35, 185–193. [Google Scholar] [CrossRef]
- Zamuner, S.R.; Gutiérrez, J.M.; Muscará, M.N.; Teixeira, S.A.; Teixeira, C.F. Bothrops asper and Bothrops jararaca snake venoms trigger microbicidal functions of peritoneal leukocytes in vivo. Toxicon 2001, 39, 1505–1513. [Google Scholar] [CrossRef]
- Murray, P.J.; Wynn, T.A.J. Protective and pathogenic functions of macrophage subsets. Nat. Rev. Immunol. 2011, 11, 723–737. [Google Scholar] [CrossRef]
- Laing, G.D.; Clissa, P.B.; Theakston, R.D.G.; Moura-da-Silva, A.M.; Taylor, M.J. Inflammatory pathogenesis of snake venom metalloproteinase-induced skin necrosis. Eur. J. Immunol. 2003, 33, 3458–3463. [Google Scholar] [CrossRef] [Green Version]
- Lentschat, A.; Karahashi, H.; Michelsen, K.S.; Thomas, L.S.; Zhang, W.; Vogel, S.N.; Arditi, M. Mastoparan, a G protein agonist peptide, differentially modulates TLR4- and TLR2-mediated signaling in human endothelial cells and murine macrophages. J. Immunol. 2005, 174, 4252–4261. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Zhan, Z.; Li, D.; Xu, L.; Ma, F.; Zhang, P.; Yao, H.; Cao, X. Intracellular MHC class II molecules promote TLR-triggered innate immune responses by maintaining activation of the kinase Btk. Nat. Immunol. 2011, 12, 416–424. [Google Scholar] [CrossRef] [PubMed]
- Medzhitov, R. Recognition of microorganisms and activation of the immune response. Nature 2007, 449, 819–826. [Google Scholar] [CrossRef]
- Zoccal, K.F.; Bitencourt Cda, S.; Paula-Silva, F.W.; Sorgi, C.A.; de Castro Figueiredo Bordon, K.; Arantes, E.C.; Faccioli, L.H. TLR2, TLR4 and CD14 recognize venom-associated molecular patterns from Tityus serrulatus to induce macrophage-derived inflammatory mediators. PLoS ONE 2014, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Takashiba, S.; Van Dyke, T.E.; Amar, S.; Murayama, Y.; Soskolne, A.W.; Shapira, L. Differentiation of monocytes to macrophages primes cells for lipopolysaccharide stimulation via accumulation of cytoplasmic nuclear factor κB. J. Infect. Immun. 1999, 67, 5573–5578. [Google Scholar] [CrossRef] [Green Version]
- Kohro, T.; Tanaka, T.; Murakami, T.; Wada, Y.; Aburatani, H.; Hamakubo, T.; Kodama, T. A comparison of differences in the gene expression profiles of phorbol 12-myristate 13-acetate differentiated THP-1 cells and human monocyte-derived macrophage. J. Atheroscler. Trhromb. 2004, 11, 88–97. [Google Scholar] [CrossRef] [Green Version]
- Aderem, A.; Underhill, D.M. Mechanisms of phagocytosis in macrophages. Annu. Rev. Immunol. 1999, 17, 593–623. [Google Scholar] [CrossRef]
- Mamede, C.C.N.; de Sousa, B.B.; da Cunha Pereira, D.F.; Matias, M.S.; de Queiroz, M.R.; de Morais, N.C.G.; Vieira, S.A.P.B.; Stanziola, L.; de Oliveira, F. Comparative analysis of local effects caused by Bothrops alternatus and Bothrops moojeni snake venoms: Enzymatic contributions and inflammatory modulations. Toxicon 2016, 117, 37–45. [Google Scholar] [CrossRef]
- Saravia-Otten, P.; Frisan, T.; Thelestam, M.; Gutierrez, J.M. Membrane independent activation of fibroblast proMMP-2 by snake venom: Novel roles for venom proteinases. Toxicon 2004, 44, 749–764. [Google Scholar] [CrossRef]
- Costa, C.; Belchor, M.; Rodrigues, C.; Toyama, D.; de Oliveira, M.; Novaes, D.; Toyama, M. Edema induced by a Crotalus durissus terrificus venom serine protease (Cdtsp 2) involves the PAR pathway and PKC and PLC activation. Int. J. Mol. Sci. 2018, 19, 2405. [Google Scholar] [CrossRef] [Green Version]
- Matsui, T.; Fujimura, Y.; Titani, K. Snake venom proteases affecting hemostasis and thrombosis. Biochim. Biophys. Acta 2000, 1477, 146–156. [Google Scholar] [CrossRef]
- Menaldo, D.L.; Bernardes, C.P.; Pereira, J.C.; Silveira, D.S.; Mamede, C.C.; Stanziola, L.; de Oliveira, F.; Pereira-Crott, L.S.; Faccioli, L.H.; Sampaio, S.V. Effects of two serine proteases from Bothrops pirajai snake venom on the complement system and the inflammatory response. Int. Immunopharmacol. 2013, 15, 764–771. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, T. The nuclear factor NF-kappaB pathway in inflammation. Cold Spring Harb. Perspect. Biol. 2009, 1, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Arias-Salvatierra, D.; Silbergeld, E.K.; Acosta-Saavedra, L.C.; Calderon-Aranda, E.S. Role of nitric oxide produced by iNOS through NF-κB pathway in migration of cerebellar granule neurons induced by lipopolysaccharide. Cell. Signal. 2011, 23, 425–435. [Google Scholar] [CrossRef]
- Eder, C. Mechanisms of interleukin-1β release. J. Immunobiol. 2009, 214, 543–553. [Google Scholar] [CrossRef]
- Martín-Sánchez, F.; Diamond, C.; Zeitler, M.; Gomez, A.; Baroja-Mazo, A.; Bagnall, J.; Spiller, D.; White, M.; Daniels, M.; Mortellaro, A. Inflammasome-dependent IL-1 β release depends upon membrane permeabilisation. J. Cell Death Differ. 2016, 23, 1219–1231. [Google Scholar] [CrossRef] [Green Version]
- Martinon, F.; Tschopp, J. Inflammatory caspases: Linking an intracellular innate immune system to autoinflammatory diseases. Cell 2004, 117, 561–574. [Google Scholar] [CrossRef] [Green Version]
- Meylan, E.; Tschopp, J.; Karin, M. Intracellular pattern recognition receptors in the host response. Nature 2006, 442, 39–44. [Google Scholar] [CrossRef]
- Zheng, H.; Fletcher, D.; Kozak, W.; Jiang, M.; Hofmann, K.J.; Corn, C.A.; Soszynski, D.; Grabiec, C.; Trumbauer, M.E.; Shaw, A. Resistance to fever induction and impaired acute-phase response in interleukin-1β-deficient mice. Immunity 1995, 3, 9–19. [Google Scholar] [CrossRef] [Green Version]
- Rider, P.; Carmi, Y.; Guttman, O.; Braiman, A.; Cohen, I.; Voronov, E.; White, M.R.; Dinarello, C.A.; Apte, R.N. IL-1α and IL-1β recruit different myeloid cells and promote different stages of sterile inflammation. J. Immunol. 2011, 187, 4835–4843. [Google Scholar] [CrossRef] [Green Version]
- Dinarello, C.A. Interleukin-1 in the pathogenesis and treatment of inflammatory diseases. Blood 2011, 117, 3720–3732. [Google Scholar] [CrossRef] [Green Version]
- Broderick, L.; De Nardo, D.; Franklin, B.S.; Hoffman, H.M.; Latz, E. The inflammasomes and autoinflammatory syndromes. Annu. Rev. Pathol. 2015, 10, 395–424. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.K.; Krohn, R.I.; Hermanson, G.; Mallia, A.; Gartner, F.; Provenzano, M.; Fujimoto, E.; Goeke, N.; Olson, B.; Klenk, D. Measurement of protein using bicinchoninic acid. Anal. Biochem. 1985, 150, 76–85. [Google Scholar] [CrossRef]
- Delafontaine, M.; Villas-Boas, I.; Mathieu, L.; Josset, P.; Blomet, J.; Tambourgi, D. Enzymatic and pro-inflammatory activities of Bothrops lanceolatus venom: Relevance for envenomation. Toxins 2017, 9, 244. [Google Scholar] [CrossRef] [PubMed]
- Rucavado, A.; Flores-Sánchez, E.; Franceschi, A.; Magalhaes, A.; Gutiérrez, J.M. Characterization of the local tissue damage induced by LHF-II, a metalloproteinase with weak hemorrhagic activity isolated from Lachesis muta muta snake venom. Toxicon 1999, 37, 1297–1312. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Megale, Â.A.A.; Magnoli, F.C.; Guidolin, F.R.; Godoi, K.S.; Portaro, F.C.V.; Dias-da-Silva, W. Bitis arietans Snake Venom and Kn-Ba, a Snake Venom Serine Protease, Induce the Production of Inflammatory Mediators in THP-1 Macrophages. Toxins 2021, 13, 906. https://doi.org/10.3390/toxins13120906
Megale ÂAA, Magnoli FC, Guidolin FR, Godoi KS, Portaro FCV, Dias-da-Silva W. Bitis arietans Snake Venom and Kn-Ba, a Snake Venom Serine Protease, Induce the Production of Inflammatory Mediators in THP-1 Macrophages. Toxins. 2021; 13(12):906. https://doi.org/10.3390/toxins13120906
Chicago/Turabian StyleMegale, Ângela Alice Amadeu, Fabio Carlos Magnoli, Felipe Raimondi Guidolin, Kemily Stephanie Godoi, Fernanda Calheta Vieira Portaro, and Wilmar Dias-da-Silva. 2021. "Bitis arietans Snake Venom and Kn-Ba, a Snake Venom Serine Protease, Induce the Production of Inflammatory Mediators in THP-1 Macrophages" Toxins 13, no. 12: 906. https://doi.org/10.3390/toxins13120906
APA StyleMegale, Â. A. A., Magnoli, F. C., Guidolin, F. R., Godoi, K. S., Portaro, F. C. V., & Dias-da-Silva, W. (2021). Bitis arietans Snake Venom and Kn-Ba, a Snake Venom Serine Protease, Induce the Production of Inflammatory Mediators in THP-1 Macrophages. Toxins, 13(12), 906. https://doi.org/10.3390/toxins13120906