4?-O-methylpyridoxine: Preparation from Ginkgo biloba Seeds and Cytotoxicity in GES-1 Cells

 , ,
, ,
Abstract
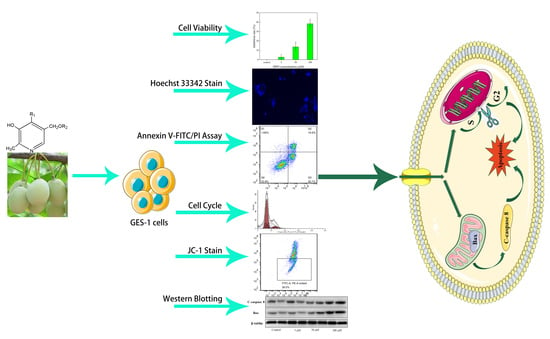
:
1. Introduction
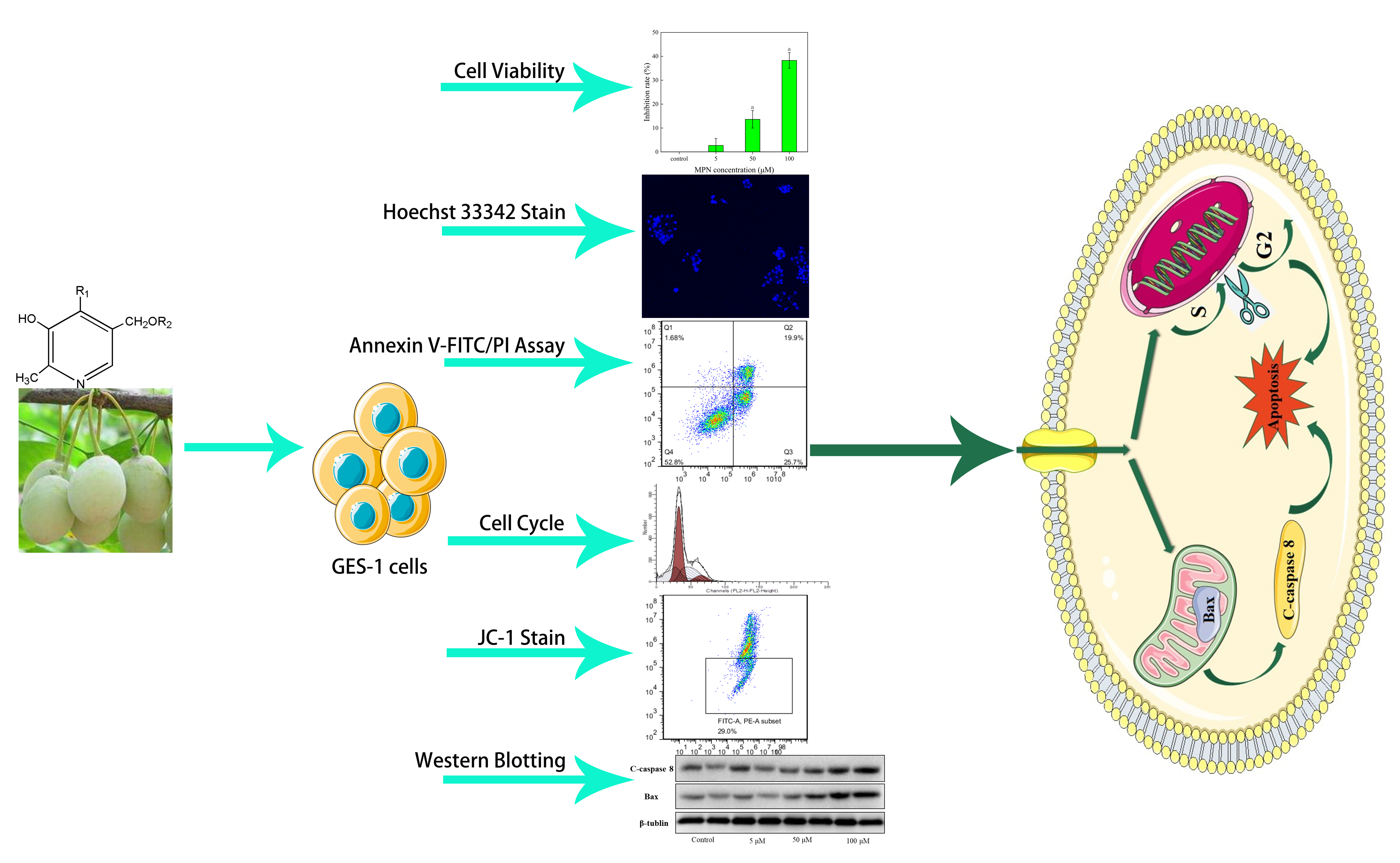
2. Results
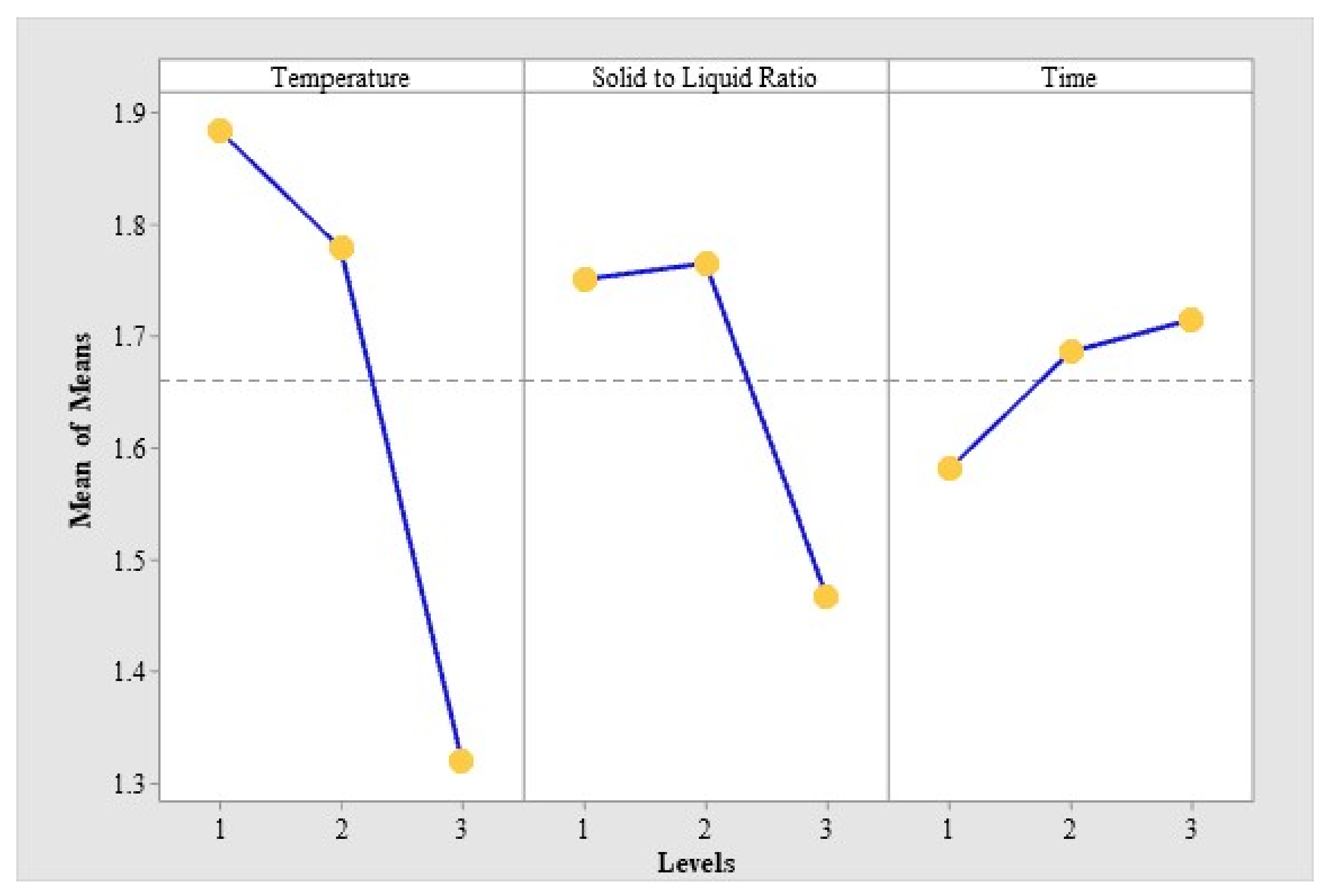
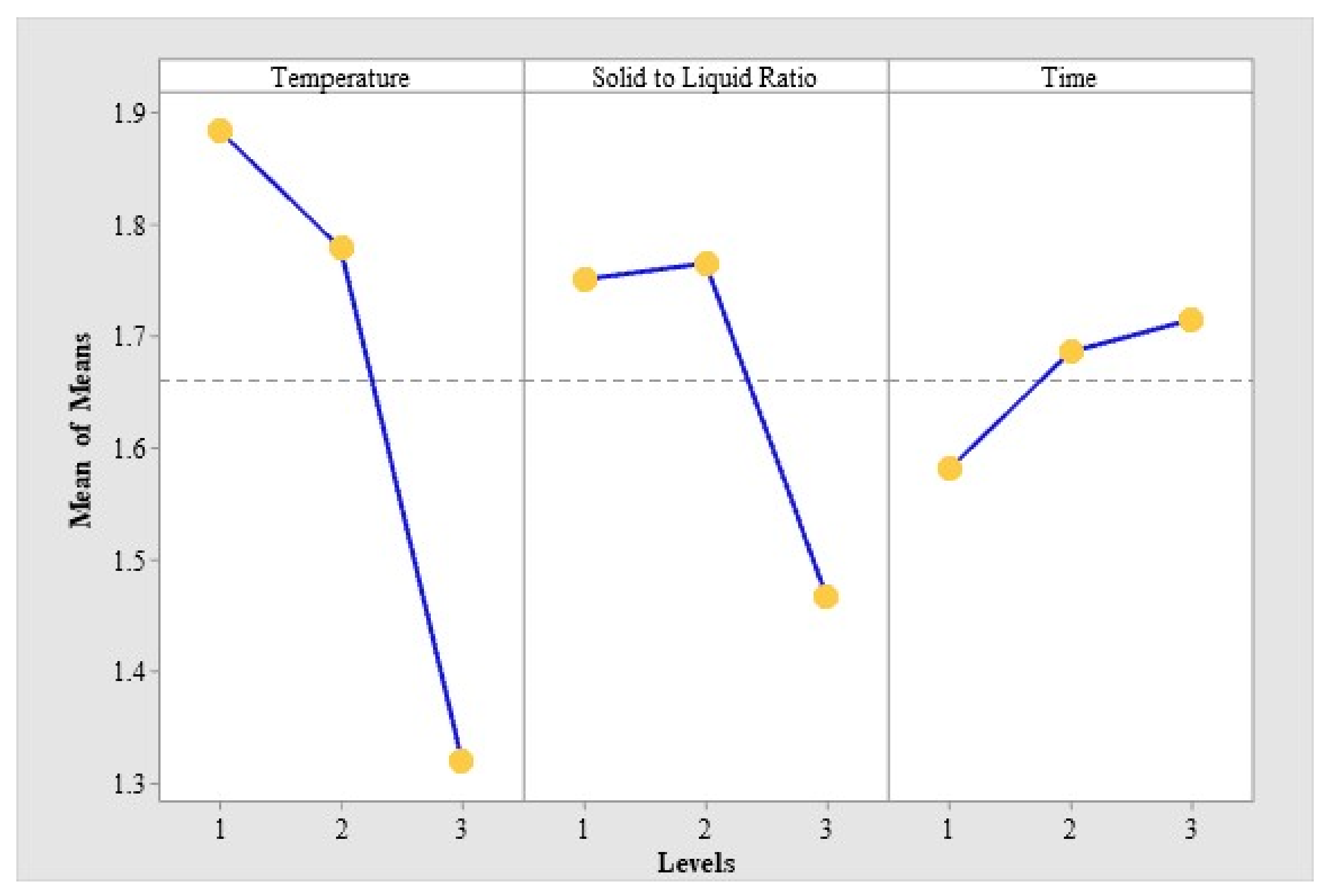
2.1. Optimization of MPN Extraction Conditions by Orthogonal Test
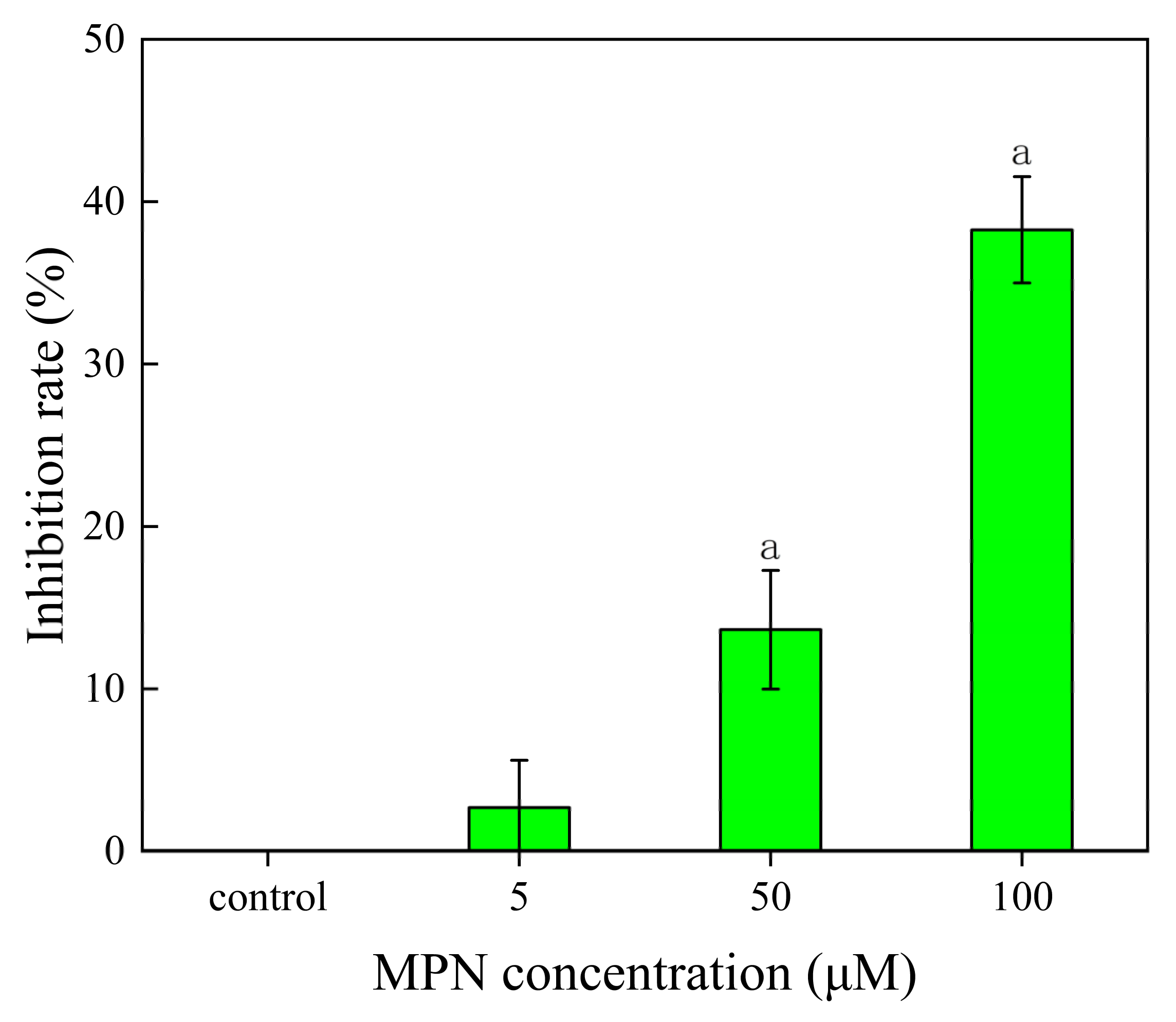
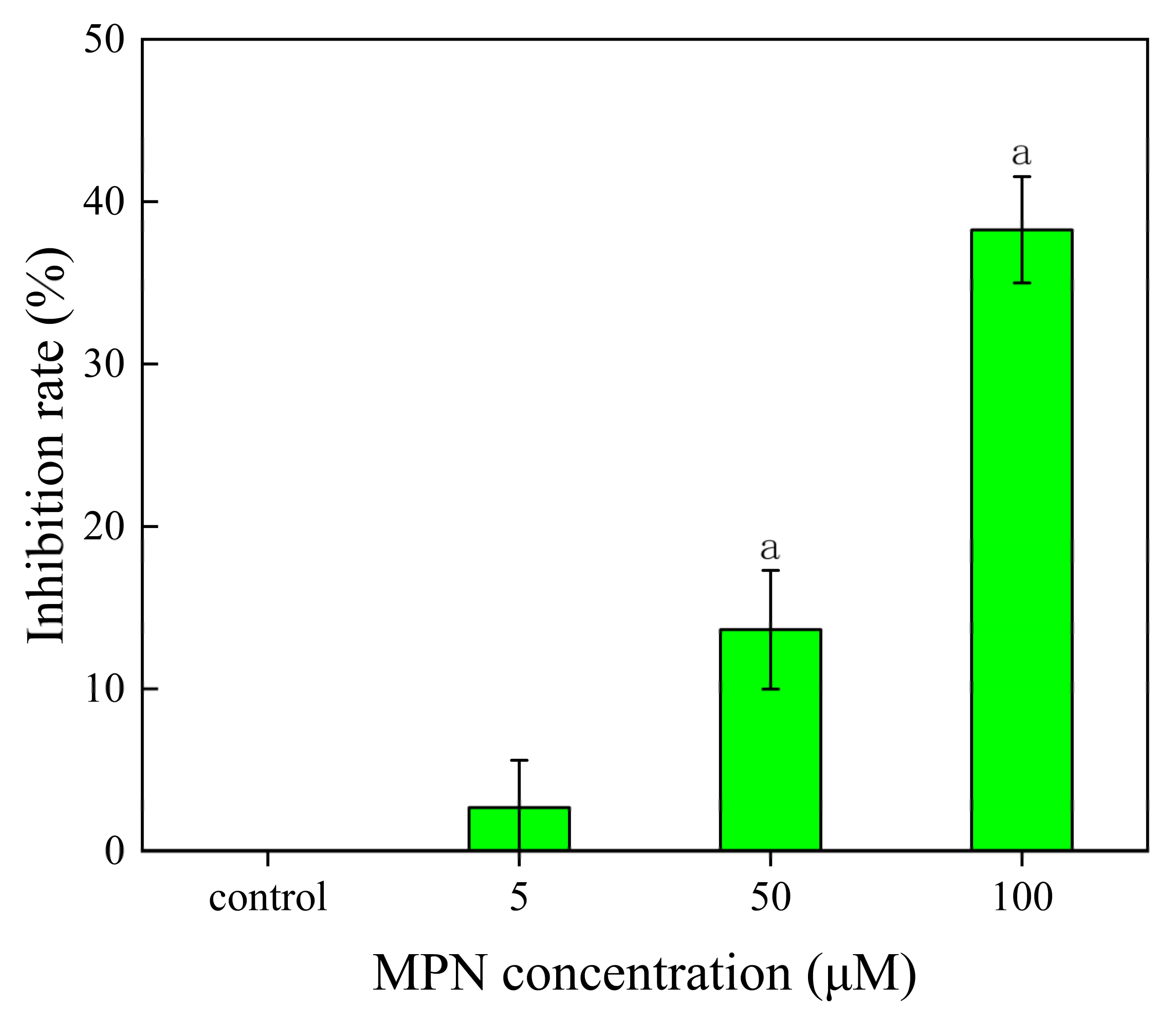
2.2. Inhibition of MPN at Different Concentrations on GES-1 Cells Activity
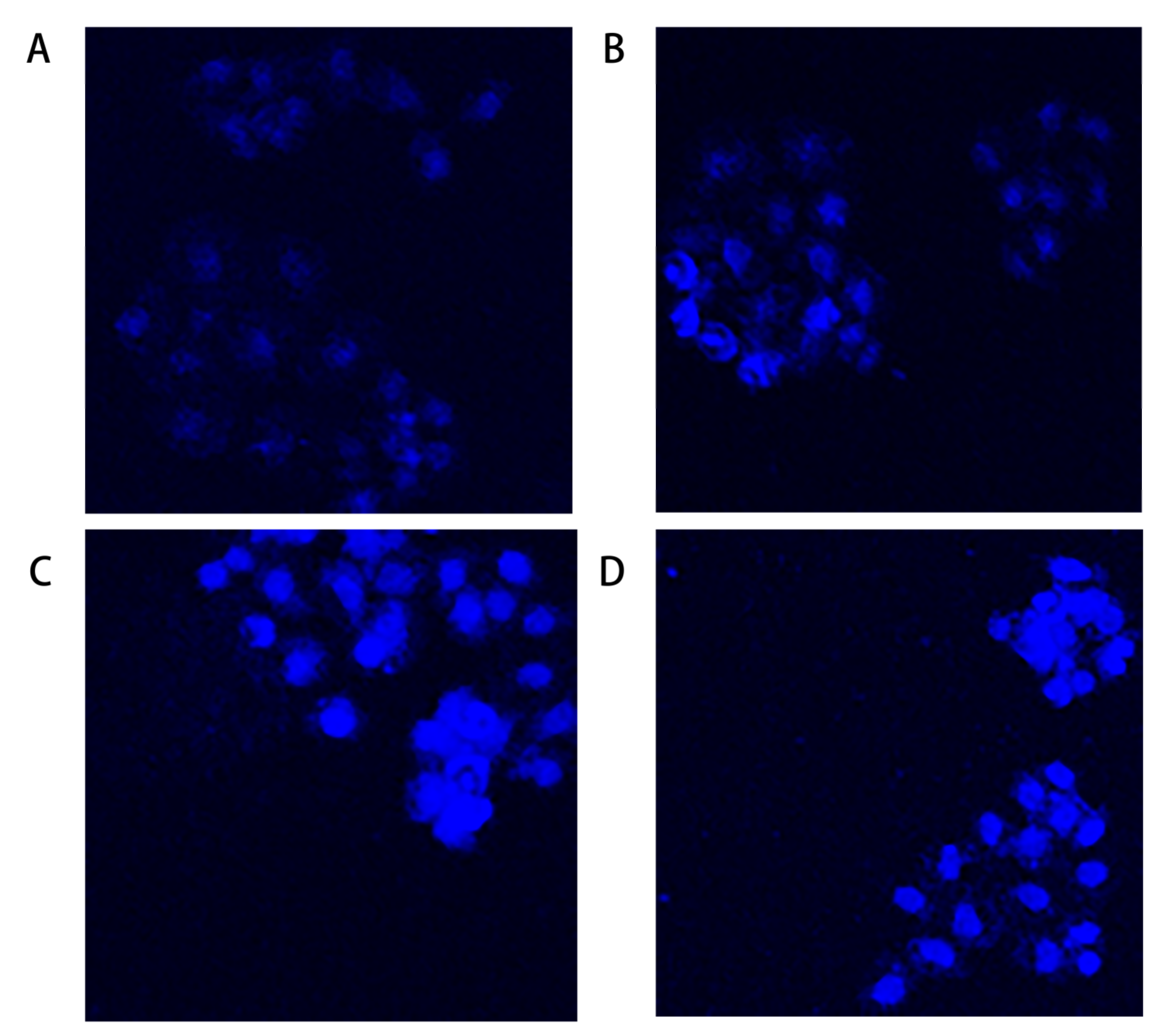
2.3. Hoechst 33342 Staining Assay
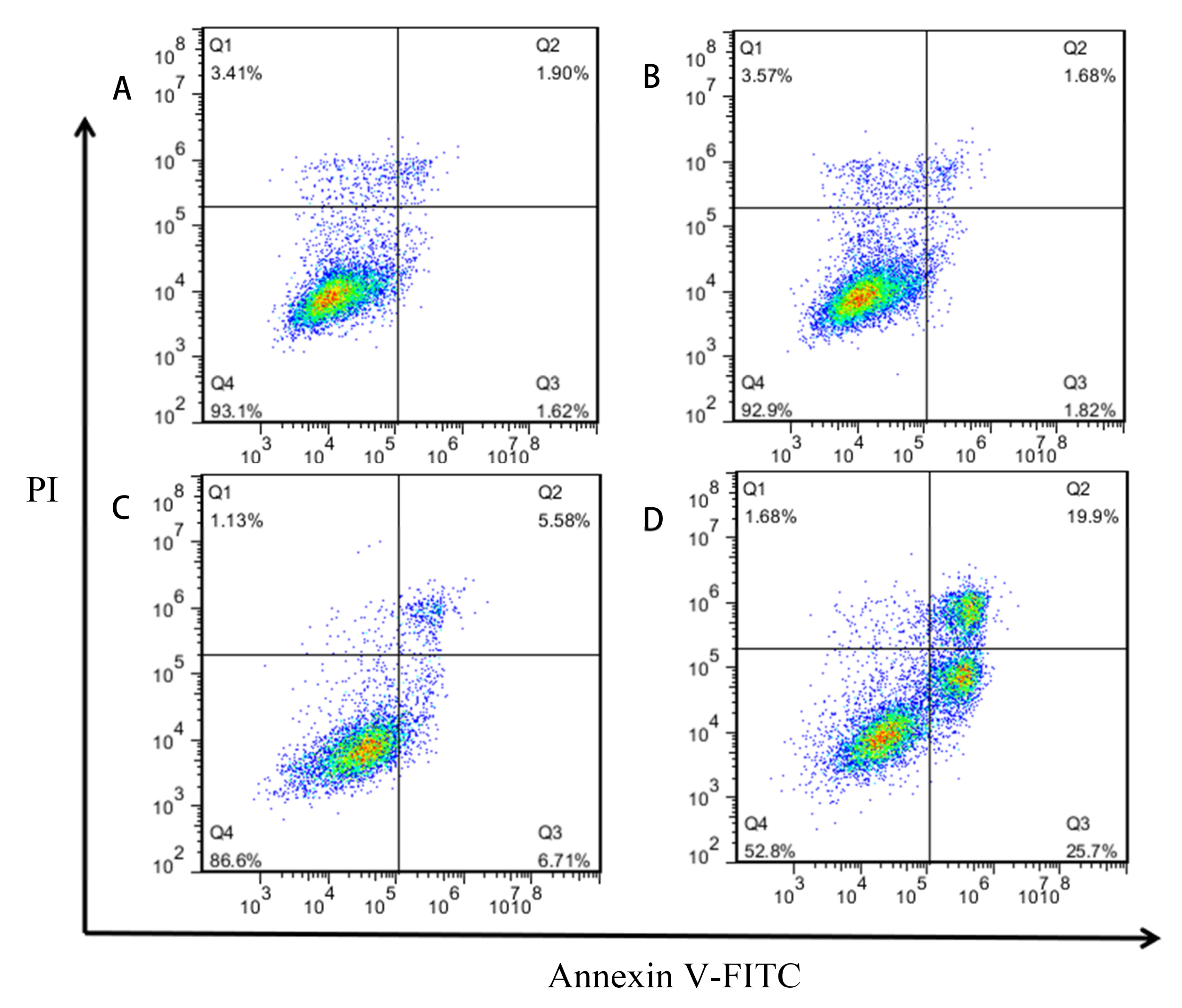
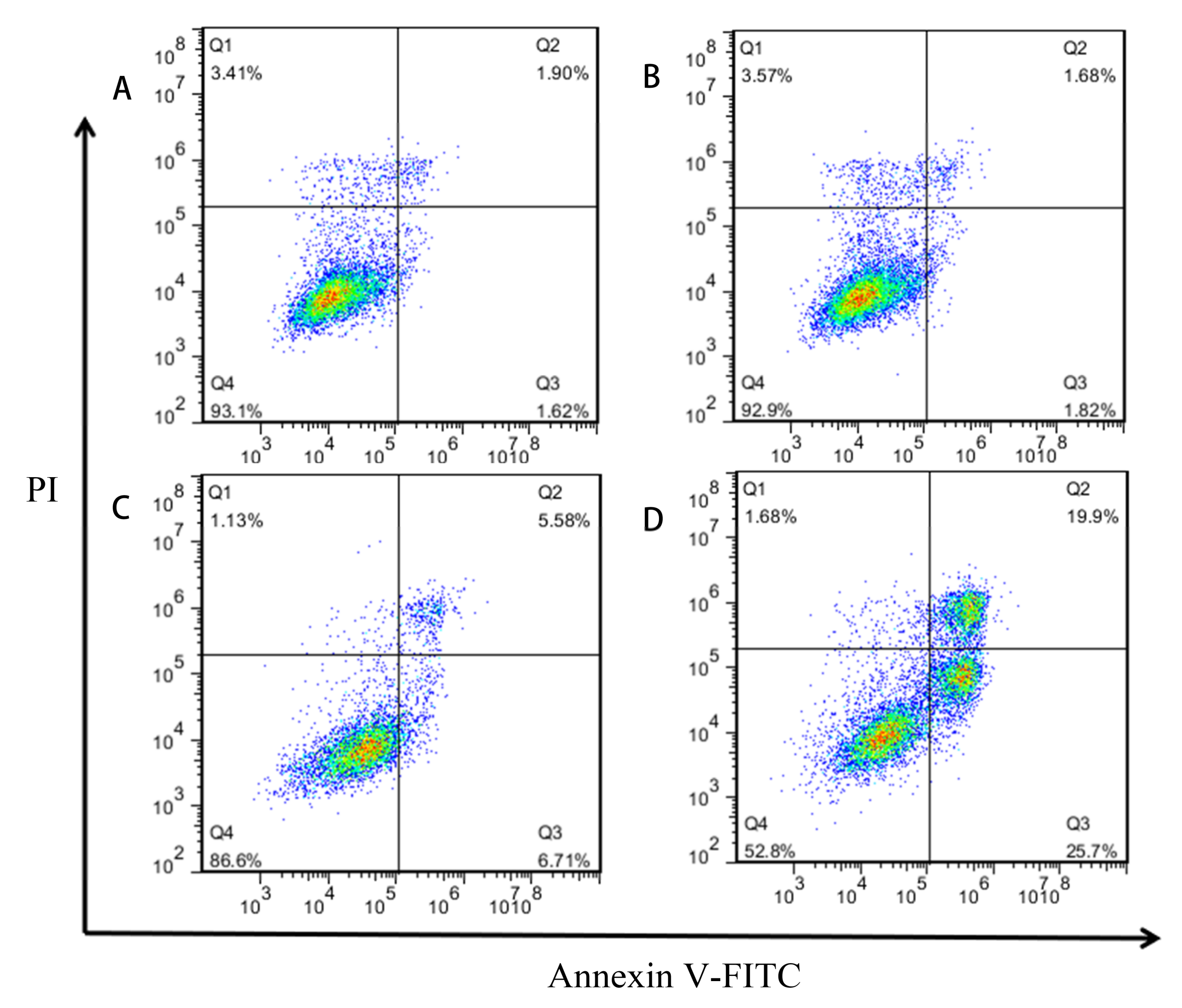
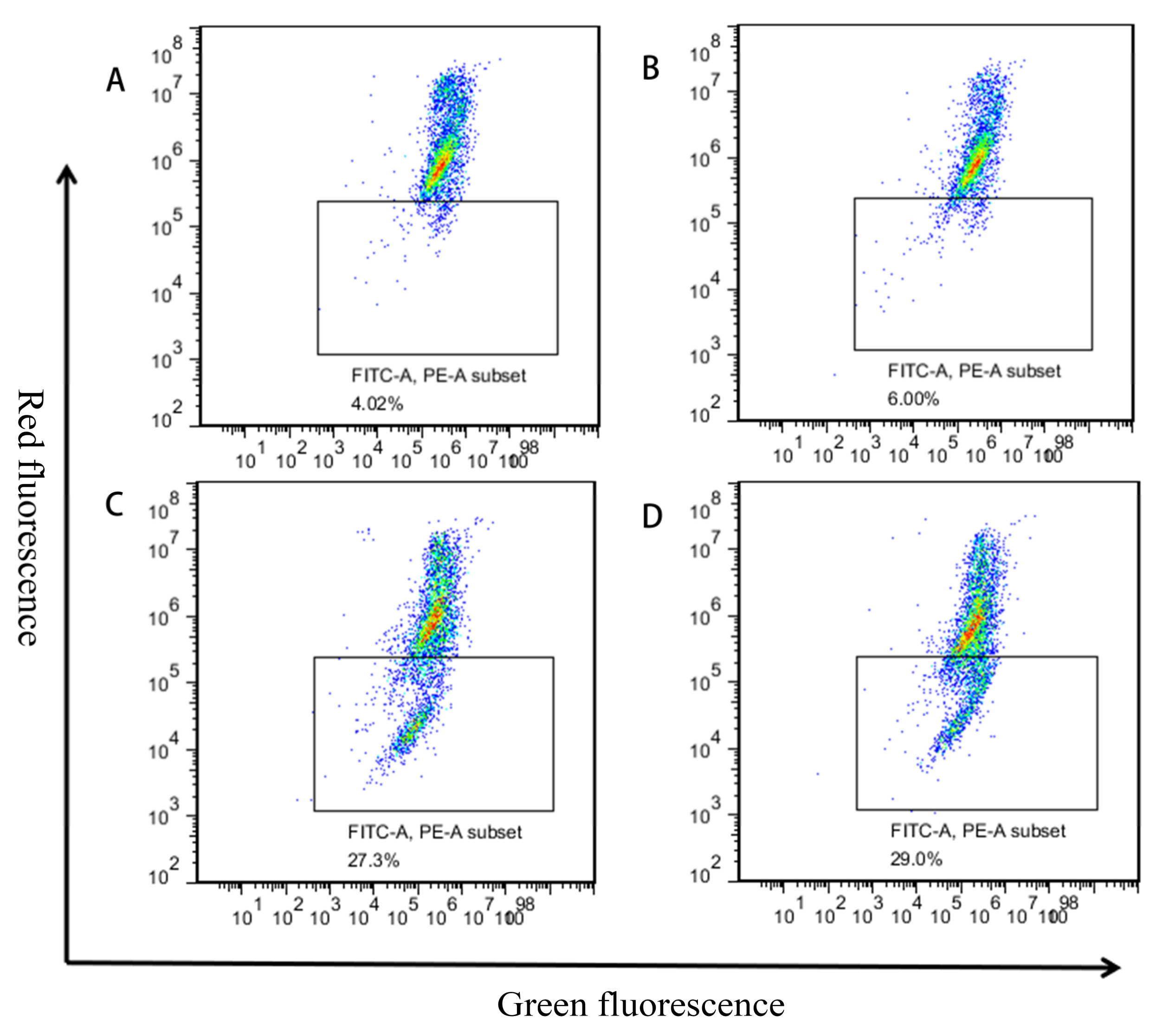
2.4. Induction of Apoptosis of GES-1 Cells by Different Concentrations of MPN
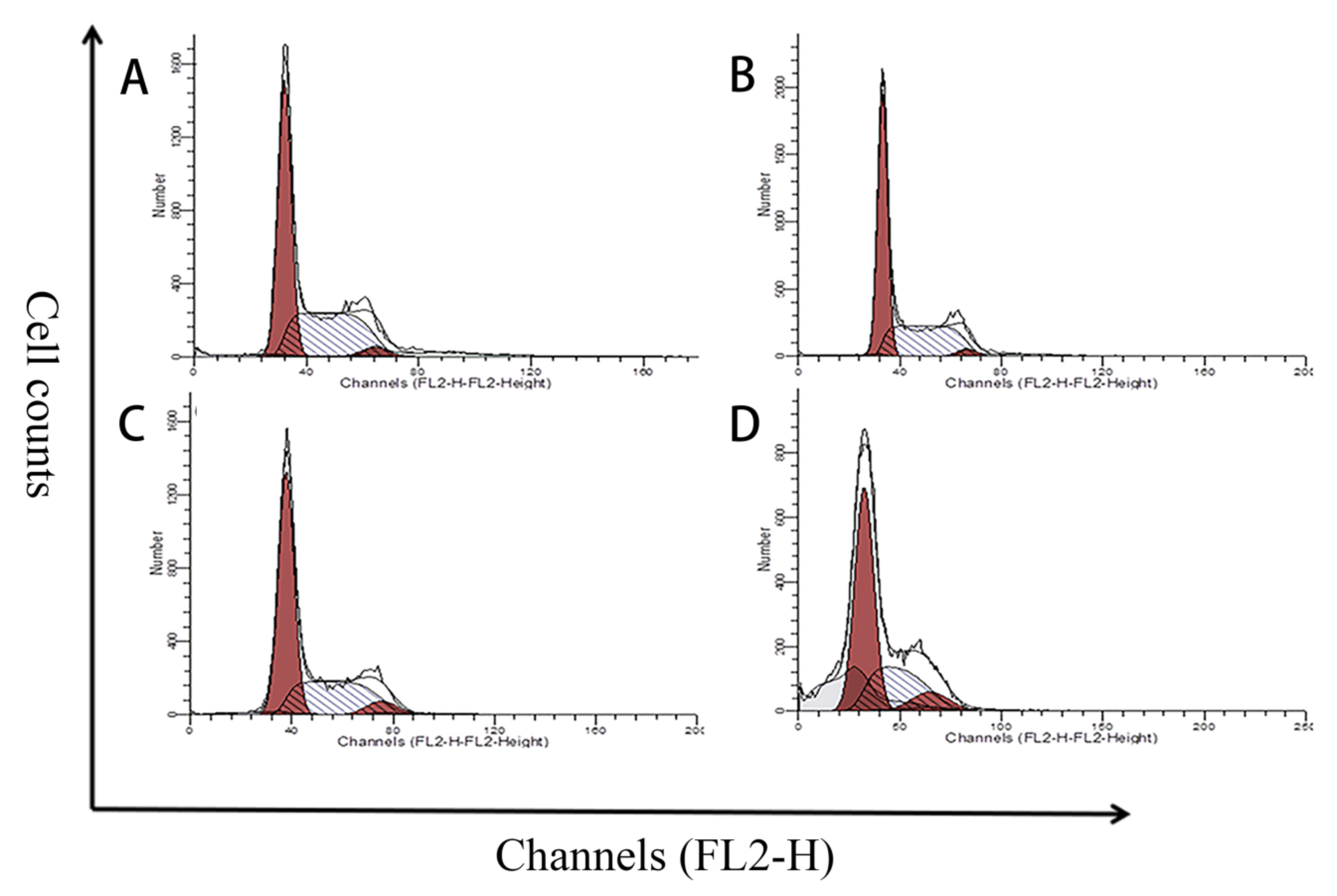
2.5. Effects of MPN at Different Concentrations on GES-1 Cells Cycle
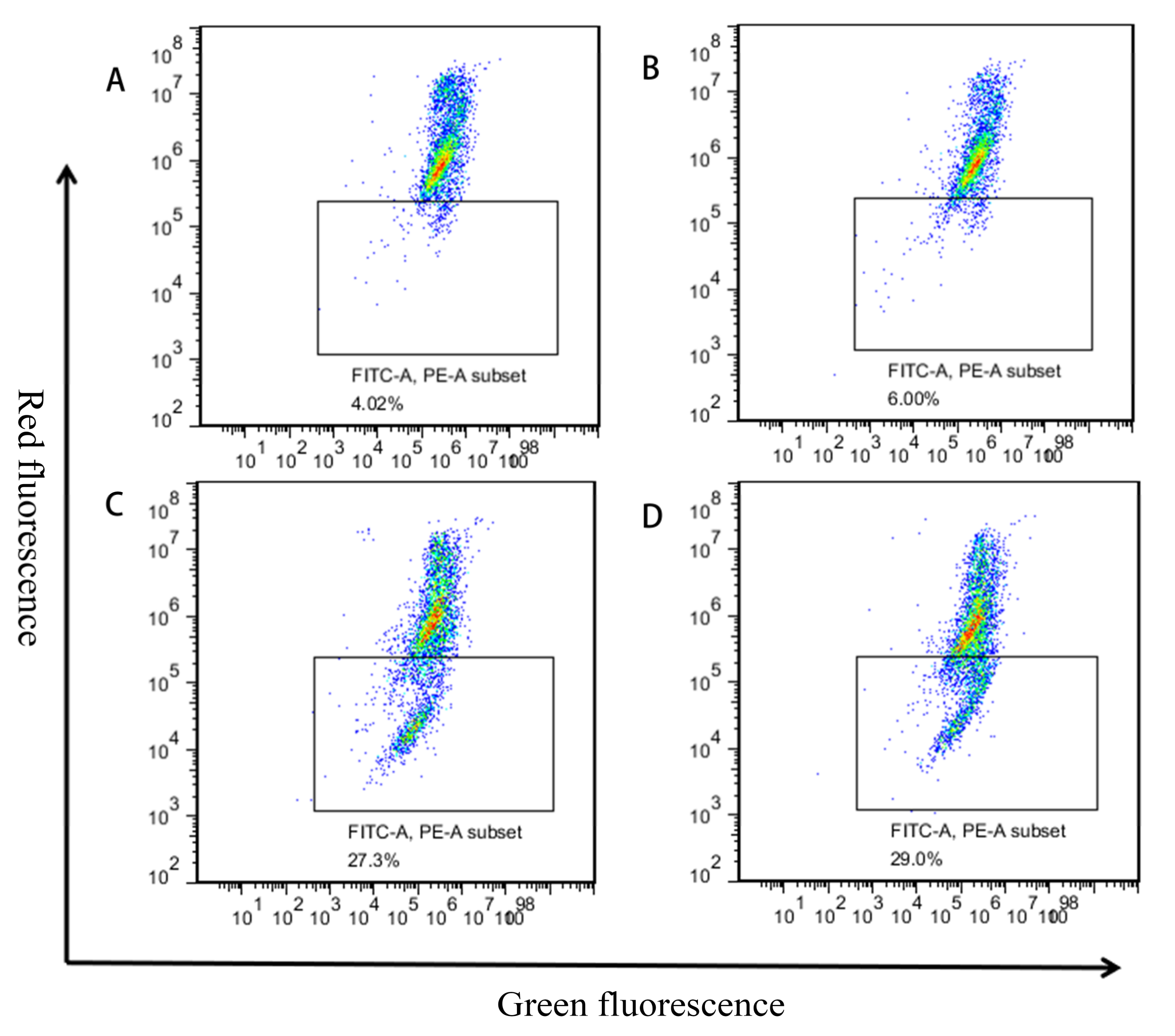
2.6. Effects of MPN at Different Concentrations on the Mitochondrial Membrane Potential of GES-1 Cells
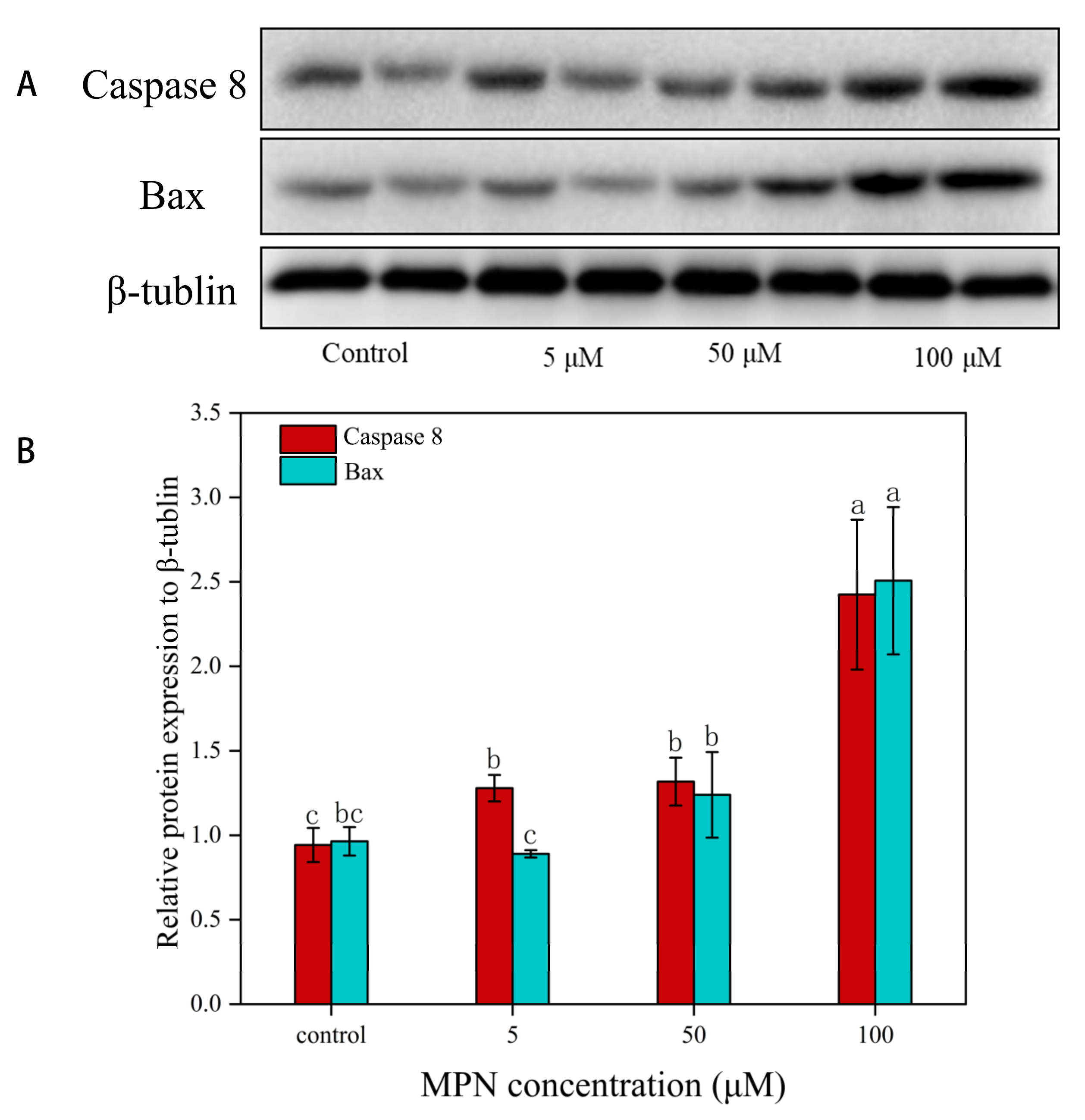
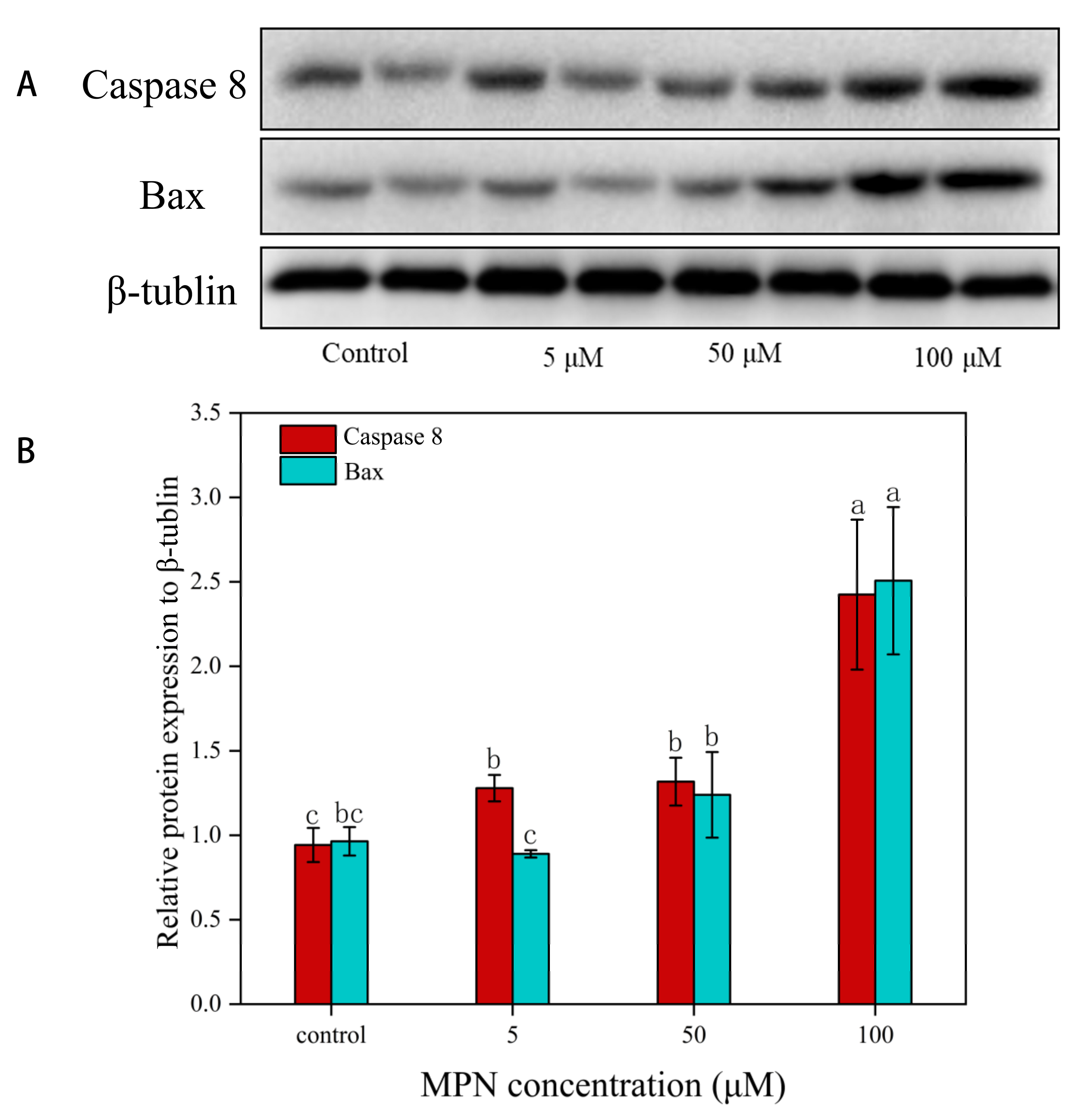
2.7. Effects of MPN at Different Concentrations on the Expression of Caspase 8 and Bax in GES-1 Cells
2.8. Correlation Analysis
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Sample Preparation
5.2. Optimization of MPN Water Extraction Process
5.3. Cell Culture
5.4. Cell Viability Assay
5.5. Hoechst 33342 Stain
5.6. Annexin V-FITC/Propidium Iodide (PI) Assay
5.7. Cell Cycle Analysis
5.8. Detection of Mitochondrial Membrane Potential
5.9. Western Blotting Analysis
5.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, W.; Zou, M.; Wu, R.; Jiang, H.; Cao, F.; Su, E. Insight into the transformation of 4′-O-methylpyridoxine and 4′-O-methylpyridoxine-5′-glucoside in Ginkgo biloba seeds undergoing the heat treatment. Ind. Crop. Prod. 2019, 140. [Google Scholar] [CrossRef]
- Gong, H.; Wu, C.; Kou, X.-H.; Fan, G.-J.; Li, T.-T.; Wang, J.-H.; Wang, T. Comparison study of 4′-O-methylpyridoxine analogues in Ginkgo biloba seeds from different regions of China. Ind. Crop. Prod. 2019, 129, 45–50. [Google Scholar] [CrossRef]
- Wada, K.; Ishigaki, S.; Ueda, K.; Sakata, M.; Haga, M. An antivitamin B6, 4′-methoxypyridoxine, from the seed of Ginkgo biloba L. Chem. Pharm. Bull. 1985, 33, 3555–3557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott, P.M.; Lau, B.P.-Y.; A Lawrence, G.; A Lewis, D. Analysis of Ginkgo biloba for the Presence of Ginkgotoxin and Ginkgotoxin 5′-Glucoside. J. AOAC Int. 2000, 83, 1313–1320. [Google Scholar] [CrossRef] [Green Version]
- Leistner, E.; Drewke, C. Ginkgo biloba and Ginkgotoxin. J. Nat. Prod. 2010, 73, 86–92. [Google Scholar] [CrossRef]
- Arenz, A.; Klein, M.; Fiehe, K.; Groß, J.; Drewke, C.; Hemscheidt, T.; Leistner, E. Occurrence of Neurotoxic 4′-O-Methylpyridoxine in Ginkgo biloba Leaves, Ginkgo Medications and Japanese Ginkgo Food. Planta Med. 1996, 62, 548–551. [Google Scholar] [CrossRef]
- Hori, Y.; Fujisawa, M.; Shimada, K.; Oda, A.; Katsuyama, S.; Wada, K. Rapid Analysis of 4-O-Methylpyridoxine in the Serum of Patients with Ginkgo Biloba Seed Poisoning by Ion-Pair High-Performance Liquid Chromatography. Biol. Pharm. Bull. 2004, 27, 486–491. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, G.A.; Scott, P.M. Improved extraction of ginkgotoxin (4′-O-methylpyridoxine) from Ginkgo biloba products. J. Aoac Int. 2005, 88, 26–29. [Google Scholar] [CrossRef] [Green Version]
- Yoshimura, T.; Udaka, N.; Morita, J.; Zhang, J.Y.; Sasaki, K.; Kobayashi, D.; Wada, K.; Hori, Y. High performance liquid chroma-tographic determination of ginkgotoxin and ginkgotoxin-5′-glucoside in Ginkgo biloba seeds. J. Liquid Chromatogr. Relat. Technol. 2006, 29, 605–616. [Google Scholar] [CrossRef]
- Kobayashi, D.; Yoshimura, T.; Johno, A.; Sasaki, K.; Wada, K. Toxicity of 4′-O-methylpyridoxine-5′-glucoside in Ginkgo biloba seeds. Food Chem. 2011, 126, 1198–1202. [Google Scholar] [CrossRef]
- Havaux, M.; Ksas, B.; Szewczyk, A.; Rumeau, D.; Franck, F.; Caffarri, S.; Triantaphylidès, C. Vitamin B6 deficient plants display increased sensitivity to high light and photo-oxidative stress. BMC Plant Biol. 2009, 9, 130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mei, N.; Guo, X.; Ren, Z.; Kobayashi, D.; Wada, K.; Guo, L. Review of Ginkgo biloba-induced toxicity, from experimental studies to human case reports. J. Environ. Sci. Health Part C 2017, 35, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Miwa, H.; Iijima, M.; Tanaka, S.; Mizuno, Y. Generalized convulsions after consuming a large amount of Gingko nuts. Epilepsia 2001, 42, 280–281. [Google Scholar] [PubMed]
- Kajiyama, Y.; Fujii, K.; Takeuchi, H.; Manabe, Y. Ginkgo Seed Poisoning. Pediatrics 2002, 109, 325–327. [Google Scholar] [CrossRef]
- Kerr, J.F.R.; Wyllie, A.H.; Currie, A.R. Apoptosis: A basic biological phenomenon with wide-ranging implication in tissue kinetics. Br. J. Cancer 1972, 26, 239–257. [Google Scholar] [CrossRef] [Green Version]
- Baar, M.P.; Brandt, R.M.C.; Putavet, D.A.; Klein, J.D.D.; Derks, K.W.J.; Bourgeois, B.R.M.; Stryeck, S.; Rijksen, Y.; Van Willigenburg, H.; Feijtel, D.A.; et al. Targeted Apoptosis of Senescent Cells Restores Tissue Homeostasis in Response to Chemotoxicity and Aging. Cell 2017, 169, 132–147.e6. [Google Scholar] [CrossRef] [Green Version]
- Candeias, E.; Sebastiao, I.; Cardoso, S.; Carvalho, C.; Santos, M.S.; Oliveira, C.R.; Moreira, P.I.; Duarte, A.I. Brain GLP-1/IGF-1 Sig-naling and Autophagy Mediate Exendin-4 Protection Against Apoptosis in Type 2 Diabetic Rats. Mol. Neurobiol. 2018, 55, 4030–4050. [Google Scholar]
- Pfeffer, C.M.; Singh, A.T.K. Apoptosis: A Target for Anticancer Therapy. Int. J. Mol. Sci. 2018, 19, 448. [Google Scholar] [CrossRef] [Green Version]
- Pena-Blanco, A.; Garcia-Saez, A.J. Bax, Bak and beyond—Mitochondrial performance in apoptosis. Febs J. 2018, 285, 416–431. [Google Scholar] [CrossRef] [Green Version]
- Levine, S.; Saltzman, A. Pyridoxine (vitamin B6) toxicity: Enhancement by uremia in rats. Food Chem. Toxicol. 2002, 40, 1449–1451. [Google Scholar] [CrossRef]
- Schaumburg, H.; Kaplan, J.; Windebank, A.; Vick, N.; Rasmus, S.; Pleasure, D.; Brown, M.J. Sensory neuropathy from pyridoxine abuse. A new megavitamin syndrome. N. Engl. J. Med. 1983, 309, 445–448. [Google Scholar] [CrossRef] [PubMed]
- Fiehe, K.; Arenz, A.; Drewke, C.; Hemscheidt, T.; Williamson, R.T.; Leistner, E. Biosynthesis of 4′-O-methylpyridoxine (Gink-gotoxin) from primary precursors. J. Nat. Prod. 2000, 63, 185–189. [Google Scholar] [CrossRef]
- Galluzzi, L.; Vitale, I.; Senovilla, L.; Olaussen, K.A.; Pinna, G.; Eisenberg, T.; Goubar, A.; Martins, I.; Michels, J.; Kratassiouk, G.; et al. Prognostic Impact of Vitamin B6 Metabolism in Lung Cancer. Cell Rep. 2012, 2, 257–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardwick, J.M.; Soane, L. Multiple functions of BCL-2 family proteins. Cold Spring Harb. Perspect. Biol. 2013, 5, 1–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Opdenbosch, N.; Lamkanfi, M. Caspases in cell death, inflammation, and disease. Immunity 2019, 50, 1352–1364. [Google Scholar] [CrossRef] [PubMed]
- Fan, T.-J.; Han, L.-H.; Cong, R.-S.; Liang, J. Caspase Family Proteases and Apoptosis. Acta Biochim. Biophys. Sin. 2005, 37, 719–727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akl, H.; Vervloessem, T.; Kiviluoto, S.; Bittremieux, M.; Parys, J.B.; De Smedt, H.; Bultynck, G. A dual role for the anti-apoptotic Bcl-2 protein in cancer: Mitochondria versus endoplasmic reticulum. Biochim. Biophys. Acta BBA Bioenerg. 2014, 1843, 2240–2252. [Google Scholar] [CrossRef] [Green Version]
- Birkinshaw, R.W.; Czabotar, P.E. The BCL-2 family of proteins and mitochondrial outer membrane permeabilisation. Semin. Cell Dev. Biol. 2017, 72, 152–162. [Google Scholar] [CrossRef] [PubMed]
- De Giorgi, F.; Lartigue, L.; Bauer, M.K.; Schubert, A.; Grimm, S.; Hanson, G.T.; Remington, S.J.; Youle, R.J.; Ichas, F. The perme ability transition pore signals apoptosis by directing Bax translocation and multimerization. FASEB J. 2002, 16, 607–609. [Google Scholar] [CrossRef]
- Darendelioglu, E.; Aykutoglu, G.; Tartik, M.; Baydas, G. Turkish propolis protects human endothelial cells In Vitro from ho-mocysteine-induced apoptosis. Acta Histochem. 2016, 118, 369–376. [Google Scholar] [CrossRef]
- Ma, L.; Wang, X.; Li, W.; Qu, F.; Liu, Y.; Lu, J.-C.; Su, G.; Zhao, Y. Conjugation of Ginsenoside with Dietary Amino Acids: A Promising Strategy to Suppress Cell Proliferation and Induce Apoptosis in Activated Hepatic Stellate Cells. J. Agric. Food Chem. 2019, 67, 10245–10255. [Google Scholar] [CrossRef] [PubMed]
- Sanderson, T.H.; Reynolds, C.A.; Kumar, R.; Przyklenk, K.; Hüttemann, M. Molecular Mechanisms of Ischemia–Reperfusion Injury in Brain: Pivotal Role of the Mitochondrial Membrane Potential in Reactive Oxygen Species Generation. Mol. Neurobiol. 2013, 47, 9–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayomy, N.A.; Abdelaziz, E.Z.; Said, M.A.; Badawi, M.S.; El-Bakary, R.H. Effect of pycnogenol and spirulina on vancomy-cin-induced renal cortical oxidative stress, apoptosis, and autophagy in adult male albino rat. Can. J. Physiol. Pharmacol. 2016, 94, 838–848. [Google Scholar] [CrossRef] [PubMed]
- Li, S.-Z.; Ren, J.; Fei, J.; Zhang, X.; Du, R.-L. Cordycepin induces Bax-dependent apoptosis in colorectal cancer cells. Mol. Med. Rep. 2018, 19, 901–908. [Google Scholar] [CrossRef] [PubMed]
- Fischer, U.M.; Janicke, R.U.; Schulze-Osthoff, K. Many cuts to ruin: A comprehensive update of caspase substrates. Cell Death Differ. 2003, 10, 76–100. [Google Scholar] [CrossRef]
- Vrolijk, M.F.; Opperhuizen, A.; Jansen, E.H.J.M.; Hageman, G.J.; Bast, A.; Haenen, G.R.M.M. The vitamin B6 paradox: Supplemen-tation with high concentrations of pyridoxine leads to decreased vitamin B6 function. Toxicol. Vitro 2017, 44, 206–212. [Google Scholar] [CrossRef]
- Ueland, P.M.; McCann, A.; Midttun, Ø.; Ulvik, A. Inflammation, vitamin B6 and related pathways. Mol. Asp. Med. 2017, 53, 10–27. [Google Scholar] [CrossRef]
- Škorňa, P.; Rimarčík, J.; Poliak, P.; Lukes, V.; Klein, E. Thermodynamic study of vitamin B6 antioxidant potential. Comput. Theor. Chem. 2016, 1077, 32–38. [Google Scholar]
- Fitzpatrick, T.B.; Amrhein, N.; Kappes, B.; Macheroux, P.; Tews, I.; Raschle, T. Two independent routes of de novo vitamin B6 biosynthesis: Not that different after all. Biochem. J. 2007, 407, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Lebiedzińska, A.; Marszałł, M.L.; Grembecka, M.; Czaja, J.; Szefer, P.; Kuta, J.; Garabato, B.D.; Kozlowski, P.M. Detection of vitamin B6 in grain products: Experimental and computational studies. Food Anal. Methods 2017, 20, 1–8. [Google Scholar]
- Schmidt, A.; Schreiner, M.; Mayer, H.K. Rapid determination of the various native forms of vitamin B6 and B2 in cow’s milk using ultra-high performance liquid chromatography. J. Chromatogr. A 2017, 1500, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Kästner, U.; Hallmen, C.; Wiese, M.; Leistner, E.; Drewke, C. The human pyridoxal kinase, a plausible target for ginkgotoxin fromGinkgo biloba. FEBS J. 2007, 274, 1036–1045. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, T.B.; Basset, G.J.; Borel, P.; Carrari, F.; DellaPenna, D.; Fraser, P.D.; Hellmann, H.; Osorio, S.; Rothan, C.; Valpuesta, V.; et al. Vitamin Deficiencies in Humans: Can Plant Science Help? Plant Cell 2012, 24, 395–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, D.; Yoshimura, T.; Johno, A.; Ishikawa, M.; Sasaki, K.; Wada, K. Decrease in pyridoxal-5′-phosphate concentration and increase in pyridoxal concentration in rat plasma by 4′-O-methylpyridoxine administration. Nutr. Res. 2015, 35, 637–642. [Google Scholar] [CrossRef] [PubMed]
- Buss, K.; Drewke, C.; Lohmann, S.; Piwonska, A.; Leistner, E. Properties and interaction of heterologously expressed glutamate decarboxylase isoenzymes GAD(65kDa) and GAD(67kDa) from human brain with ginkgotoxin and its 5′-phosphate. J. Med. Chem. 2001, 44, 3166–3174. [Google Scholar] [CrossRef]
- Gong, H.; Wu, C.; Fan, G.-J.; Li, T.-T.; Wang, J.; Wang, T. Determination and Comparison of 4′-O-Methylpyridoxine Analogues in Ginkgo biloba Seeds at Different Growth Stages. J. Agric. Food Chem. 2018, 66, 7916–7922. [Google Scholar] [CrossRef]
- Gong, H.; Wu, C.-E.; Kou, X.-H.; Fan, G.-J.; Li, T.-T.; Wang, J.-H.; Wang, T. Determination of the native contents of 4′-O-methylpyridoxine and its glucoside in Ginkgo biloba Seeds. J. Food Meas. Charact. 2020, 14, 917–924. [Google Scholar] [CrossRef]
- Madeo, F.; Fröhlich, E.; Fröhlich, K.-U. A Yeast Mutant Showing Diagnostic Markers of Early and Late Apoptosis. J. Cell Biol. 1997, 139, 729–734. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Test No. | Factors | Extraction Amount of 4′-O-methylpyridoxine (MPN) (μg/mg) | |||
|---|---|---|---|---|---|
| A: Temperature (°C) | B: Solid-to-Liquid Ratio (m/v) | C: Time(min) | Blank | ||
| 1 | 1(40) | 1(1:5) | 1(60) | 1 | 1.870 |
| 2 | 1 | 2(1:10) | 2(80) | 2 | 2.007 |
| 3 | 1 | 3(1:15) | 3(100) | 3 | 1.777 |
| 4 | 2(50) | 1 | 2 | 3 | 1.928 |
| 5 | 2 | 2 | 3 | 1 | 1.913 |
| 6 | 2 | 3 | 1 | 2 | 1.496 |
| 7 | 3(60) | 1 | 3 | 2 | 1.455 |
| 8 | 3 | 2 | 1 | 3 | 1.376 |
| 9 | 3 | 3 | 2 | 1 | 1.123 |
| Source of Variation | Sum of Squares | Degree of Freedom | F Value | F0.05(2.2) = 19 | F0.01(2.2) = 99 |
|---|---|---|---|---|---|
| A: Temperature | 0.545 | 2 | 109.000 | * | * * |
| B: Solid-to-liquid ratio | 0.172 | 2 | 34.400 | * | |
| C: Time | 0.030 | 2 | 6.000 | ||
| Blank | 0.005 | 2 | 1.000 | ||
| Error | 0.01 | 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, J.-P.; Gong, H.; Wu, C.-E.; Fan, G.-J.; Li, T.-T.; Wang, J.-H. 4?-O-methylpyridoxine: Preparation from Ginkgo biloba Seeds and Cytotoxicity in GES-1 Cells. Toxins 2021, 13, 95. https://doi.org/10.3390/toxins13020095
Zhu J-P, Gong H, Wu C-E, Fan G-J, Li T-T, Wang J-H. 4?-O-methylpyridoxine: Preparation from Ginkgo biloba Seeds and Cytotoxicity in GES-1 Cells. Toxins. 2021; 13(2):95. https://doi.org/10.3390/toxins13020095
Chicago/Turabian StyleZhu, Jin-Peng, Hao Gong, Cai-E Wu, Gong-Jian Fan, Ting-Ting Li, and Jia-Hong Wang. 2021. "4?-O-methylpyridoxine: Preparation from Ginkgo biloba Seeds and Cytotoxicity in GES-1 Cells" Toxins 13, no. 2: 95. https://doi.org/10.3390/toxins13020095
APA StyleZhu, J.-P., Gong, H., Wu, C.-E., Fan, G.-J., Li, T.-T., & Wang, J.-H. (2021). 4?-O-methylpyridoxine: Preparation from Ginkgo biloba Seeds and Cytotoxicity in GES-1 Cells. Toxins, 13(2), 95. https://doi.org/10.3390/toxins13020095