Ageratina adenophora Inhibits Spleen Immune Function in Rats via the Loss of the FRC Network and Th1–Th2 Cell Ratio Elevation


 ,
,
Abstract
:1. Introduction
2. Results
2.1. Effect of A. adenophora Exposure on Rat Weight and Spleen
2.2. A. adenophora Exposure Induced Abnormal CD3+ T-Lymphocyte Levels in the Blood and Spleen
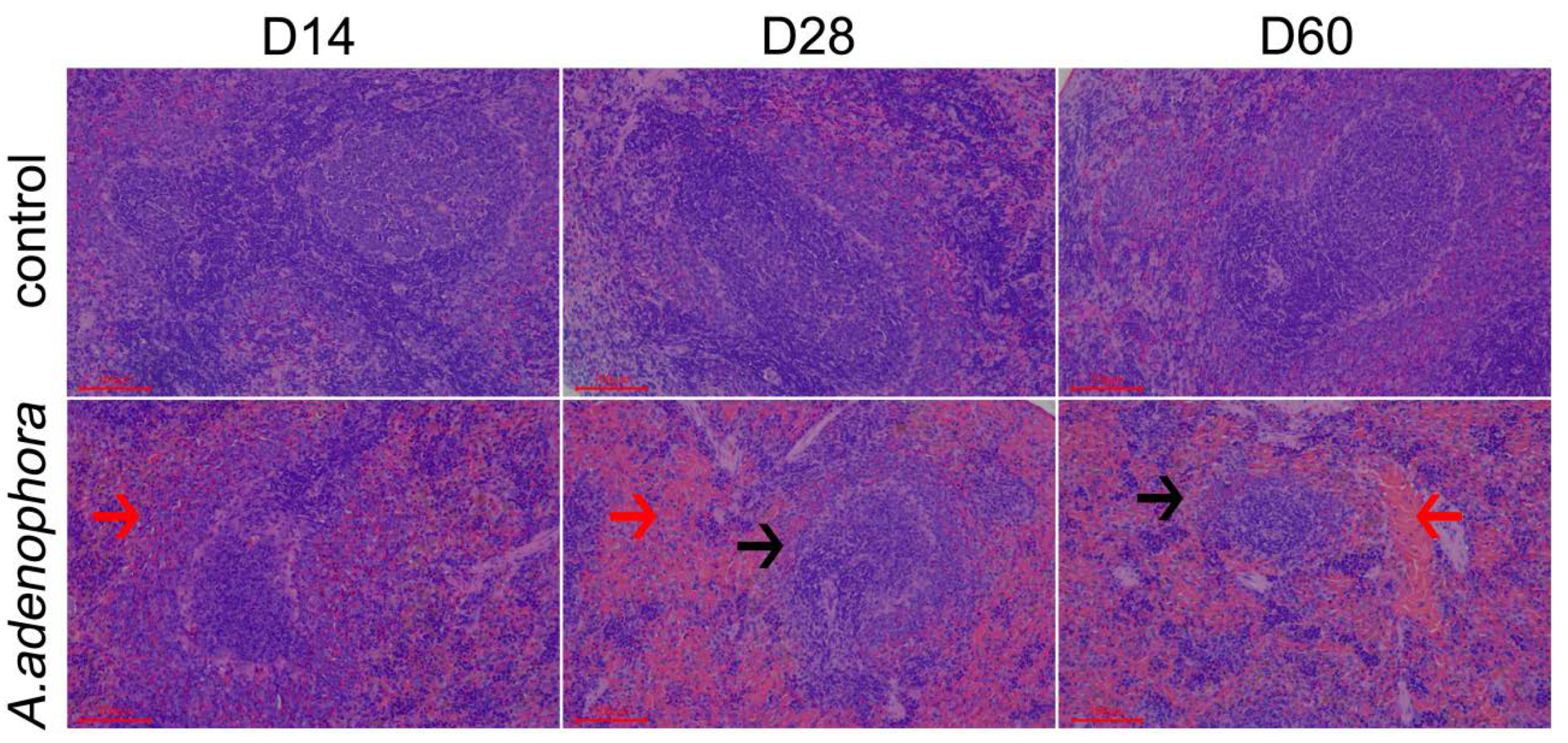
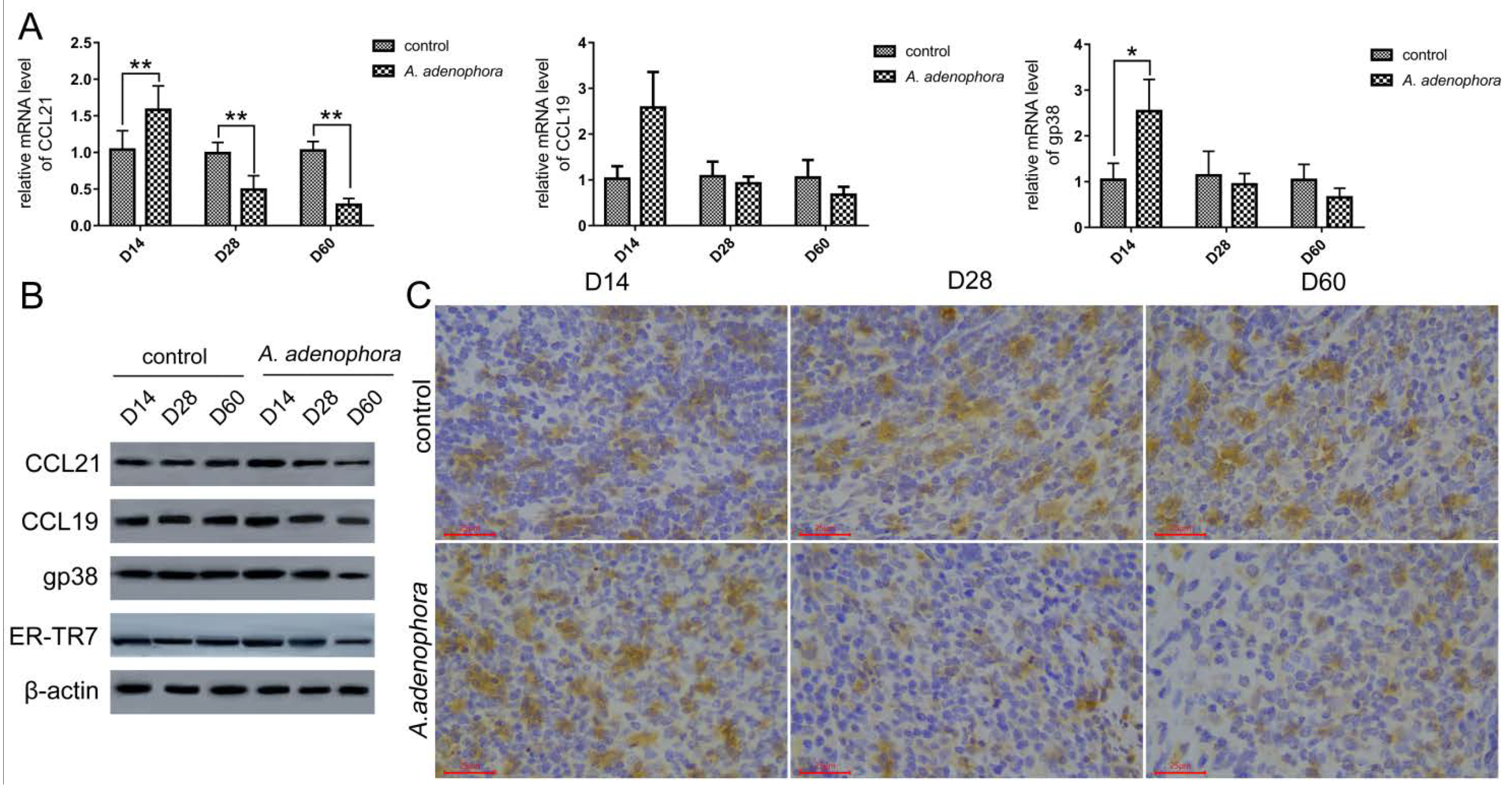
2.3. A. adenophora Exposure Caused Disruption of Splenic Fibroblast Reticulocyte (FRC) Network
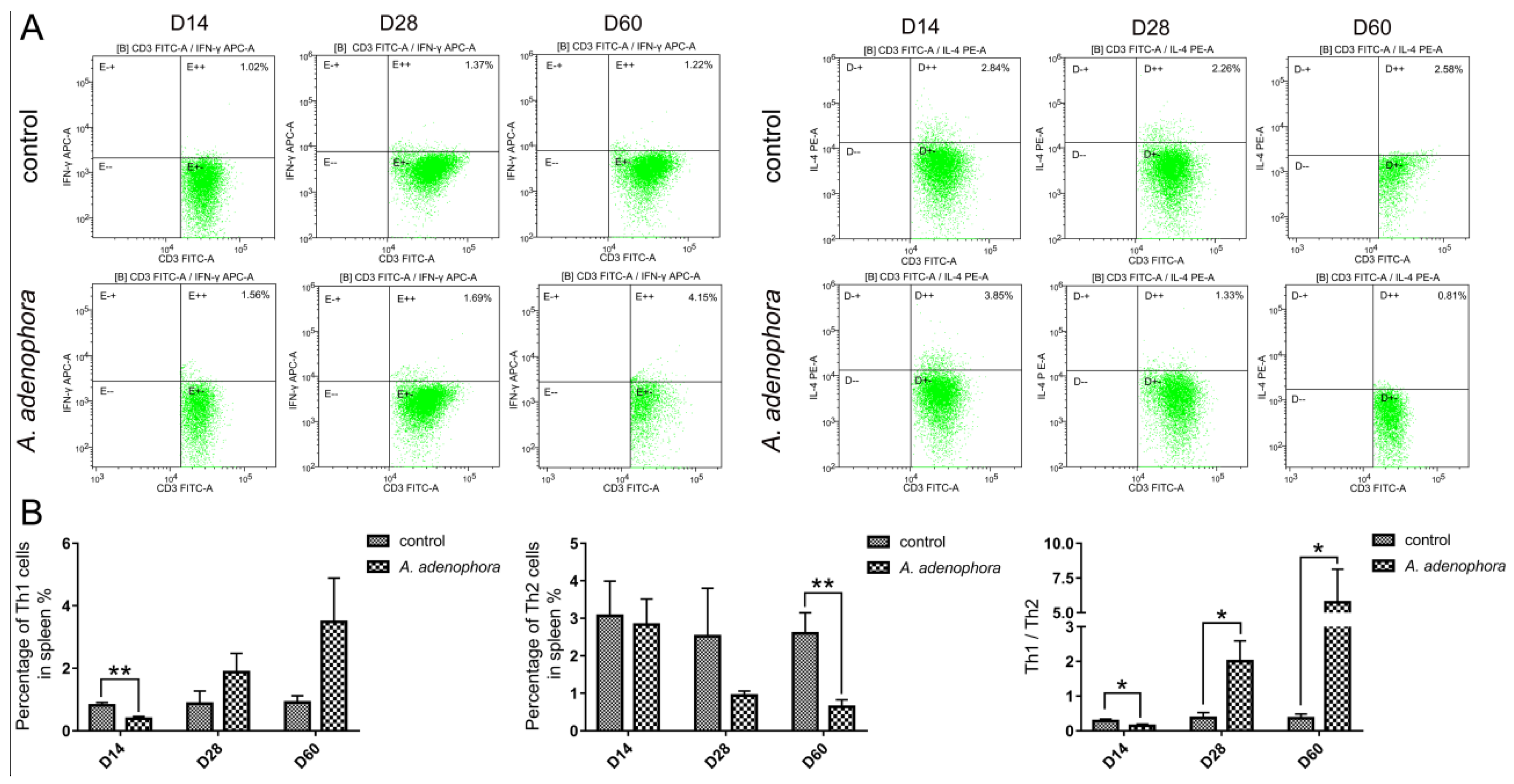
2.4. A. adenophora-Induced Splenic Th1 and Th2 Cell Imbalance and Increased Th1-Type Response
3. Discussion
4. Conclusions
5. Material and Methods
5.1. Plant Material
5.2. Animals and Experimental Design
5.3. Histological Analysis
5.4. Flow Cytometry Analysis
5.5. Extraction of Total RNA and Quantitative Real Time RT-PCR (qRT-PCR)
5.6. Enzyme-Linked Immunosorbent Assay (ELISA)
5.7. Western Blot Analysis
5.8. Immunohistochemical Staining
5.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Qiang, S. The history and status of the study on Crofton weed (Eupatorium adenophorum Spreng), a worldwide weed. J. Wuhan Botan. Res. 1998, 16, 366–372. [Google Scholar]
- Wang, R.; Wang, Y. Invasion dynamics and potential spread of the invasive alien plant species Ageratina adenophora (asteraceae) in China. Divers. Distrib. 2006, 12, 397–408. [Google Scholar] [CrossRef]
- Yan, Q.; Yang, J.; Li, H.; Cao, A.; Chen, Q.; Wen, Y.; He, L. Advances in the studies on the chemical components and bioactivity of Eupatorium adenophorum Spreng as a intruding species. J. Beijing Normal Univ. 2006, 42, 70–73. [Google Scholar]
- He, L.; Hou, J.; Gan, M.L.; Shi, J.G.; Chantrapromma, S.; Fun, H.K.; Williams, I.D.; Sung, H.H.Y. Cadinane sesquiterpenes from the leaves of Eupatorium adenophorum. J. Nat. Prod. 2008, 71, 1485–1488. [Google Scholar] [CrossRef]
- Li, M.; Li, Z.; Ye, M. The chemical compositions and their bioactivities in the different parts of Eupatorium adenophorum Spreng. J. Yunnan Agric. Univ. 2008, 23, 42–46. [Google Scholar]
- Zhang, M.; Liu, W.; Zheng, M.; Xu, Q.; Wan, F.; Wang, J.; Tan, J. Bioactive quinic acid derivatives from Ageratina adenophora. Molecules 2013, 18, 14096–14104. [Google Scholar] [CrossRef] [Green Version]
- Kurade, N.; Jaitak, V.; Kaul, K.; Sharma, O. Chemical composition and anti-bacterial activity of essential oils of Lantana camara, Ageratum houstonianum and Eupatorium adenophorum. Pharmaceutical. Biol. 2010, 48, 539–544. [Google Scholar] [CrossRef] [Green Version]
- Wan, F.; Liu, W.; Guo, J.; Qiang, S.; Li, B.; Wang, J.; Yang, G.; Niu, H.; Gui, F.; Huang, W. Invasive mechanism and control strategy of Ageratina adenophora (Sprengel). Sci. China Life Sci. 2010, 53, 1291–1298. [Google Scholar] [CrossRef]
- O’Sullivan, B. Crofton weed (Eupatorium adenophorum) toxicity in horses. Austral. Vet. J. 1979, 55, 19–21. [Google Scholar] [CrossRef]
- Verma, A.; Yadava, B.; Sampath, K. Possible use of Spreng (Eupatorium adenophorum) in animal feeding. Indian J. Anim. Nutr. 1987, 4, 189–192. [Google Scholar]
- He, Y.; Mo, Q.; Luo, B.; Qiao, Y.; Xu, R.; Zuo, Z.; Deng, J.; Nong, X.; Peng, G.; He, W.; et al. Induction of apoptosis and autophagy via mitochondria- and PI3K/Akt/mTOR-mediated pathways by E. adenophorum in hepatocytes of Saanen goat. Oncotarget 2016, 7, 54537–54548. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, Y.; Saini, N.; Anjum, N.; Verma, P. A review of ethnomedicinal, phytochemical, pharmacological and toxicological aspects of Eupatorium adenophorum spreng. Asian J. Biomed. Pharm. Sci. 2018, 8, 25–35. [Google Scholar]
- Bhardwaj, R.; Singh, A.; Sharma, O.; Dawra, R.; Kurade, N.; Mahato, S. Hepatotoxicity and cholestasis in rats induced by the sesquiterpene 9-oxo-10,11-dehydroageraphorone isolated from Eupatorium adenophorum. J. Biochem. Mol. Toxicol. 2001, 15, 279–286. [Google Scholar] [CrossRef]
- Kaushal, V.; Dawra, R.; Sharma, P.; Kurade, N. Biochemical alterations in the blood plasma of rats associated with hepatotoxicity induced by Eupatorium adenophorum. Vet. Res. Commun. 2001, 25, 601–608. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Zeng, C.; Liu, S.; Fu, J.; Hu, L.; Shi, Z.; Yue, D.; Ren, Z.; Zhong, Z.; Zuo, Z.; et al. Ageratina adenophora induces mice hepatotoxicity via ROS-NLRP3-mediated pyroptosis. Sci. Rep. 2018, 8, 16032. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, C.; Liu, X.; Yan, D.; Li, Y.; Wang, Q.; Cao, A.; Guo, M. Immunotoxicity assessment of cadinene sesquiterpenes from Eupatorium adenophorum in mice. J. Integr. Agric. 2016, 15, 2319–2325. [Google Scholar] [CrossRef] [Green Version]
- Mo, Q.; Hu, L.; Weng, J.; Zhang, Y.; Zhou, Y.; Xu, R.; Zuo, Z.; Deng, J.; Ren, Z.; Zhong, Z.; et al. Euptox A induces G1 arrest and autophagy via p38 MAPK- and PI3K/Akt/mTOR-mediated pathways in mouse splenocytes. J. Histochem. Cytochem. 2017, 65, 543–558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, W.; Zeng, C.; Yue, D.; Liu, S.; Ren, Z.; Zuo, Z.; Deng, J.; Peng, G.; Hu, Y. Ageratina adenophora causes spleen toxicity by inducing oxidative stress and pyroptosis in mice. Royal Soc. Open Sci. 2019, 6, 190127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poudel, R.; Neupane, N.P.; Mukeri, I.H.; Alok, S.; Verma, A. An updated review on invasive nature, phytochemical evaluation, & pharmacological activity of Ageratina adenophora. Int. J. Pharm. Sci. Res. 2020, 11, 2510–2520. [Google Scholar]
- He, L.; Yang, J.; Cao, A.; Liu, Y.; An, Y.; Shi, J. A new sequiterpenoid from Eupatorium adenophorum Spreng. Chin. J. Chem. 2006, 24, 1375–1377. [Google Scholar] [CrossRef]
- Ouyang, C.; Liu, X.; Liu, Q.; Bai, J.; Li, H.; Wang, Q.; Yan, D.; Mao, L.; Cao, A.; Guo, M. Toxicity assessment of cadinene sesquiterpenes from Eupatorium adenophorum in mice. Nat. Prod. Bioprospect. 2014, 5, 29–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; Cao, L.; Zhang, L.; Yuan, X.; Zhao, B. Hepatotoxicity of Eupatorium adenophorum extracts and identification of major hepatotoxic components. Nat. Prod. Res. 2017, 31, 2788–2792. [Google Scholar] [CrossRef]
- Oelrichs, P.; Calanasan, C.; Macleod, J.; Seawright, A.; Ng, J. The isolation of a compound from Eupatorium adenophorum (Spreng) (Ageratina adenophora) causing hepatotoxicity in mice. Nat. Toxins. 1995, 3, 350–354. [Google Scholar] [CrossRef]
- Okyere, S.K.; Mo, Q.; Pei, G.; Ren, Z.; Deng, J.; Hu, Y. Euptox A induces G0/G1 arrest and apoptosis of hepatocyte via ROS, mitochondrial dysfunction and caspases-dependent path- ways in vivo. J. Toxicol. Sci. 2020, 45, 661–671. [Google Scholar] [CrossRef] [PubMed]
- Mebius, R.E.; Kraal, G. Structure and function of the spleen. Nat. Rev. Immunol. 2005, 5, 606–616. [Google Scholar] [CrossRef] [PubMed]
- Bajénoff, M.; Glaichenhaus, N.; Germain, R. Fibroblastic reticular cells guide T-lymphocyte entry into and migration within the splenic T-cell zone. J. Immunol. 2008, 181, 3947–3954. [Google Scholar] [CrossRef] [Green Version]
- Haan, J.; Mebius, R.; Kraal, G. Stromal cells of the mouse spleen. Front. Immunol. 2012, 3, 201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Astarita, J.; Acton, S.; Turley, S. Podoplanin: Emerging functions in development, the immune system, and cancer. Front. Immunol. 2012, 3, 283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aw, D.; Hilliard, L.; Nishikawa, Y.; Cadman, E.; Lawrence, R.; Palmer, D. Disorganization of the splenic microanatomy in ageing mice. Immunology 2016, 148, 92–101. [Google Scholar] [CrossRef] [Green Version]
- Masters, A.; Jellison, E.; Puddington, L.; Khanna, K.; Haynes, L. Attrition of T-cell zone fibroblastic reticular cell number and function in aged spleens. ImmunoHorizons 2018, 2, 155–163. [Google Scholar] [CrossRef] [Green Version]
- Katakai, T.; Hara, T.; Sugai, M.; Gonda, H.; Shimizu, A. Lymph node fibroblastic reticular cells construct the stromal reticulum via contact with lymphocytes. J. Exp. Med. 2004, 200, 783–795. [Google Scholar] [CrossRef] [PubMed]
- Lokmic, Z.; Lämmermann, T.; Sixt, M.; Cardell, S.; Hallmann, R.; Sorokin, L. The extracellular matrix of the spleen as a potential organizer of immune cell compartments. Semin. Immunol. 2008, 20, 4–13. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Yamane, H.; Paul, W.E. Differentiation of Effector CD4 T Cell Populations. Annu. Rev. Immunol. 2010, 28, 445–489. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Mo, Q.; Hu, Y.; Chen, W.; Luo, B.; Wu, L.; Qiao, Y.; Xu, R.; Zhou, Y.; Zuo, Z.; et al. E. adenophorum induces cell cycle arrest and apoptosis of splenocytes through the mitochon- drial pathway and caspase activation in Saanen goats. Sci. Rep. 2015, 5, 15967. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zhang, Y.; Gu, W.; He, L.; Sun, B. TH1/TH2 cell function in immune system. Adv. Exp. Med. Biol. 2014, 841, 845. [Google Scholar]
- Sun, B.; Zhang, Y. Overview of orchestration of CD4+ T-cell subsets in immune responses. Adv. Exp. Med. Biol. 2014, 841, 1. [Google Scholar]
- Raphael, I.; Nalawade, S.; Eagar, T.; Forsthuber, T. T-Cell subsets and their signature cytokines in autoimmune and inflammatory diseases. Cyotkine 2015, 74, 5–17. [Google Scholar] [CrossRef] [Green Version]
- Newton, K.; Dixit, V. Signaling in innate immunity and inflammation. Cold Spring Harbor Perspect. Biol. 2012, 4, a6049. [Google Scholar] [CrossRef] [PubMed]
- Rathinam, V.; Fitzgerald, K. Inflammasome complexes: Emerging mechanisms and effector functions. Cell 2016, 165, 792–800. [Google Scholar] [CrossRef] [Green Version]
- Rahimi, K.; Ahmadi, A.; Hassanzadeh, K.; Soleimani, Z.; Sathyapalan, T.; Mohammedi, A.; Sahebkar, A. Targeting the balance of T-helper cell responses by curcumin in inflammatory and autoimmune states. Autoimmun. Rev. 2019, 18, 738–748. [Google Scholar] [CrossRef] [PubMed]
- Cameron, P.U.; Jones, P.; Gorniak, M.; Dunster, K.; Paul, E.; Lewin, S.; Woolley, L. Splenectomy Associated Changes in IgM Memory B Cells in an Adult Spleen Registry Cohort. PLoS ONE 2011, 6, e23164. [Google Scholar] [CrossRef] [Green Version]
- Bai, J.; Cao, A.; Guo, M.; Liu, X.; Liu, X.; Liang, H.; Zhou, B. Identification of 9-oxo-10,11-dehydroagerophorone in Eupatorium adenophorum by HPLC. Chinese Bull. Botany 2011, 46, 470–475. [Google Scholar]
- Bai, J. Foundational Studies of Toxicity Substances in Exotic Invasive Plant Eupatorium adenophorum Spreng. Ph.D. Thesis, China Academy of Agricultural Sciences, Beijing, China, 2012. [Google Scholar]
- Tan, J.; Huang, C.; Luo, Q.; Liu, W.; Cheng, D.; Li, Y.; Xia, Y.; Li, C.; Tang, L.; Fang, J.; et al. Soy isoflavones ameliorate fatty acid metabolism of visceral adipose tissue by increasing AMPK activity in male rats with diet-induced obesity (DIO). Molecules 2019, 24, 2809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeyabalan, J.; Aqil, F.; Munagala, R.; Annamalai, L.; Vadhanam, M.; Gupta, R. Chemopreventive and therapeutic activity of dietary blueberry against estrogen-mediated breast cancer. J. Agric. Food Chem. 2014, 62, 3963–3971. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | D0 | D14 | D28 | D42 | D60 |
|---|---|---|---|---|---|
| Control | 225.99 ± 7.57 | 276.03 ± 9.33 | 312.77 ± 11.54 | 347.71 ± 15.91 | 366.60 ± 15.65 |
| A. adenophora | 227.37 ± 11.70 | 239.14 ± 16.97 | 207.27 ± 10.29 * | 199.20 ± 14.64 ** | 190.78 ± 11.86 ** |
| Time | Spleen Weight/g | Spleen–Body Ratio % | ||
|---|---|---|---|---|
| Control | A. adenophora | Control | A. adenophora | |
| D14 | 0.53 ± 0.092 | 0.53 ± 0.055 | 0.19 ± 0.035 | 0.23 ± 0.040 |
| D28 | 0.62 ± 0.092 | 0.76 ± 0.058 * | 0.20 ± 0.033 | 0.37 ± 0.041 |
| D60 | 0.66 ± 0.085 | 1.14 ± 0.160 ** | 0.18 ± 0.019 | 0.60 ± 0.110 ** |
| Factors | D14 | D28 | D60 | |||
|---|---|---|---|---|---|---|
| Control | A. adenophora | Control | A. adenophora | Control | A. adenophora | |
| CCL21 | 0.72 ± 0.20 | 1.05 ± 0.29 | 0.67 ± 0.28 | 0.43 ± 0.18 | 0.78 ± 0.03 | 0.21 ± 0.07 ** |
| CCL19 | 0.99 ± 0.24 | 1.29 ± 0.08 * | 0.94 ± 0.08 | 0.79 ± 0.16 | 0.90 ± 0.08 | 0.46 ± 0.07 ** |
| ER-TR7 | 1.10 ± 0.15 | 1.12 ± 0.26 | 1.00 ± 0.32 | 0.77 ± 0.29 | 0.94 ± 0.13 | 0.38 ± 0.08 * |
| gp38 (WB) | 0.61 ± 0.20 | 1.21 ± 0.32 ** | 0.63 ± 0.23 | 0.65 ± 0.25 | 0.65 ± 0.17 | 0.25 ± 0.05 * |
| gp38 (immune-chemistry) | 0.22 ± 0.04 | 0.34 ± 0.04 ** | 0.22 ± 0.02 | 0.13 ± 0.01 ** | 0.21 ± 0.03 | 0.08 ± 0.01 ** |
| Factors | D14 | D28 | D60 | |||
|---|---|---|---|---|---|---|
| Control | A. adenophora | Control | A. adenophora | Control | A. adenophora | |
| T-bet | 0.96 ± 0.25 | 1.44 ± 0.42 | 1.05 ± 0.26 | 1.51 ± 0.31 | 1.09 ± 0.33 | 1.84 ± 0.74 ** |
| IFN-γ | 1.10 ± 0.43 | 1.37 ± 0.48 | 1.13 ± 0.49 | 2.67 ± 1.79 | 0.94 ± 0.22 | 3.95 ± 0.80 * |
| GATA3 | 0.92 ± 0.24 | 0.78 ± 0.09 | 0.99 ± 0.32 | 0.88 ± 0.29 | 1.03 ± 0.15 | 0.50 ± 0.09 * |
| IL-4 | 1.05 ± 0.34 | 1.17 ± 0.63 | 1.06 ± 0.45 | 0.76 ± 0.30 | 1.08 ± 0.32 | 0.54 ± 0.22 * |
| Factors | D14 | D28 | D60 | |||
|---|---|---|---|---|---|---|
| Control | A. adenophora | Control | A. adenophora | Control | A. adenophora | |
| IFN-γ | 1614.62 ± 32.34 | 1621.06 ± 87.80 | 1615.07 ± 112.19 | 1614.96 ± 38.27 | 1609.92 ± 97.87 | 1767.11 ± 66.85 ** |
| IL-12 | 13.30 ± 0.97 | 13.84 ± 0.71 | 13.14 ± 0.84 | 14.08 ± 0.29 | 13.24 ± 0.82 | 15.27 ± 0.50 ** |
| IL-4 | 89.20 ± 7.79 | 97.87 ± 4.71 * | 85.07 ± 4.79 | 80.78 ± 5.84 | 89.32 ± 3.64 | 72.25 ± 3.82 ** |
| IL-10 | 41.84 ± 3.11 | 37.40 ± 1.66* | 41.42 ± 3.77 | 37.44 ± 2.20* | 41.54 ± 2.95 | 34.69 ± 1.50 ** |
| Ingredients | Content % |
|---|---|
| Water | 10.00 |
| Crude protein | 18.00 |
| Crude fat | 4.00 |
| Crude fiber | 5.00 |
| Crude ash | 8.00 |
| Calcium | 1.50 |
| Phosphorus | 1.00 |
| Lysine | 0.82 |
| Methionine + cystine | 0.53 |
| Gene | Primer Sequence (5′→3′) | Sequence Number |
|---|---|---|
| IFN γ | F: GGCAAAAGGACGGTAACACG R: TCTGTGGGTTGTTCACCTCG | NM_138880.3 |
| IL-4 | F: AAGGAACACCACGGAGAACG R: CAGACCGCTGACACCTCTAC | NM_201270.1 |
| T-bet | F: GTATGCCAGGGAACCGCTTA R: ATTGTTGGAAGCCCCCTTGT | NM_001107043.1 |
| GATA3 | F: CTCTTCCCTCCCAGCAGCCTAC R: AGTACCATCTCGCCGCCACAG | NM_133293.1 |
| CCL21 | F: ACAGGAAGCAAGAACCGAGC R: TCTGTCTGTTCAGTCCCCTTG | NM_001008513.1 |
| CCL19 | F: GTGCTAACGATGCGGAAGAC R: AGAGCTGGTAGCCCCTTAGT | NM_001108661.1 |
| gp38 | F: TAAGAGAGCTTCCACCTTGCC R: GGCTTGTCCCGTATCTTTCCT | NM_019358.2 |
| β-actin | F: ACGGTCAGGTCATCACTATCG R: GGCATAGAGGTCTTTACGGATG | NM_031144.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ren, Z.; Gao, P.; Okyere, S.K.; Cui, Y.; Wen, J.; Jing, B.; Deng, J.; Hu, Y. Ageratina adenophora Inhibits Spleen Immune Function in Rats via the Loss of the FRC Network and Th1–Th2 Cell Ratio Elevation. Toxins 2021, 13, 309. https://doi.org/10.3390/toxins13050309
Ren Z, Gao P, Okyere SK, Cui Y, Wen J, Jing B, Deng J, Hu Y. Ageratina adenophora Inhibits Spleen Immune Function in Rats via the Loss of the FRC Network and Th1–Th2 Cell Ratio Elevation. Toxins. 2021; 13(5):309. https://doi.org/10.3390/toxins13050309
Chicago/Turabian StyleRen, Zhihua, Pei Gao, Samuel Kumi Okyere, Yujing Cui, Juan Wen, Bo Jing, Junliang Deng, and Yanchun Hu. 2021. "Ageratina adenophora Inhibits Spleen Immune Function in Rats via the Loss of the FRC Network and Th1–Th2 Cell Ratio Elevation" Toxins 13, no. 5: 309. https://doi.org/10.3390/toxins13050309
APA StyleRen, Z., Gao, P., Okyere, S. K., Cui, Y., Wen, J., Jing, B., Deng, J., & Hu, Y. (2021). Ageratina adenophora Inhibits Spleen Immune Function in Rats via the Loss of the FRC Network and Th1–Th2 Cell Ratio Elevation. Toxins, 13(5), 309. https://doi.org/10.3390/toxins13050309