Effect of Seaweed-Derived Fucoidans from Undaria pinnatifida and Fucus vesiculosus on Coagulant, Proteolytic, and Phospholipase A2 Activities of Snake Bothrops jararaca, B. jararacussu, and B. neuwiedi Venom

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. Toxicity of FVF and UPF
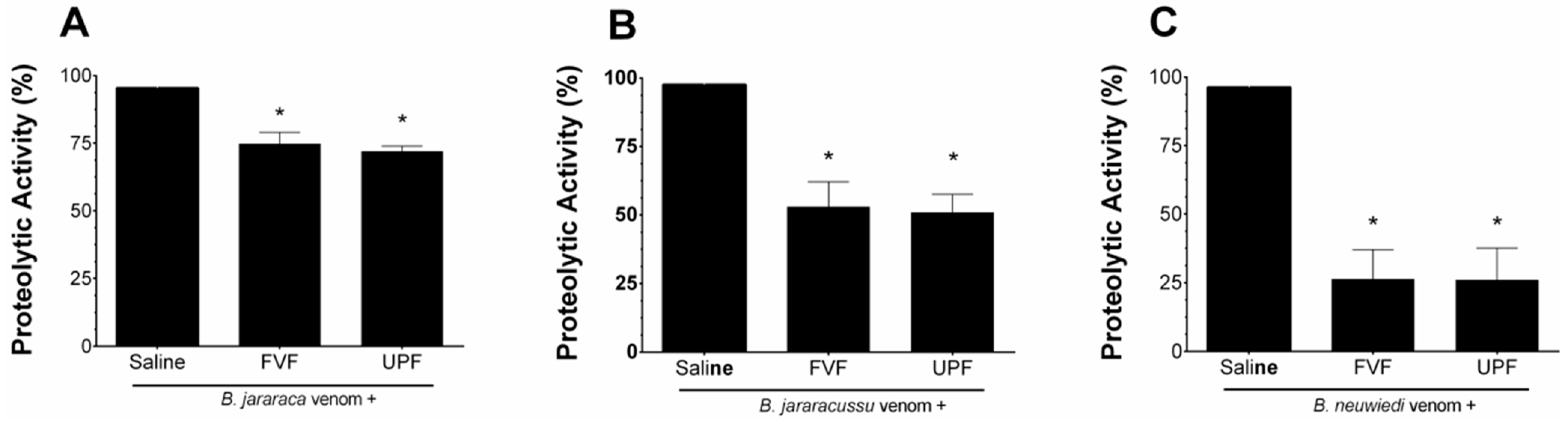
2.2. Inhibitory Effect of FVF and UPF against the Proteolytic Activity of the Venom of B. jararaca, B. jararacussu, and B. neuwiedi
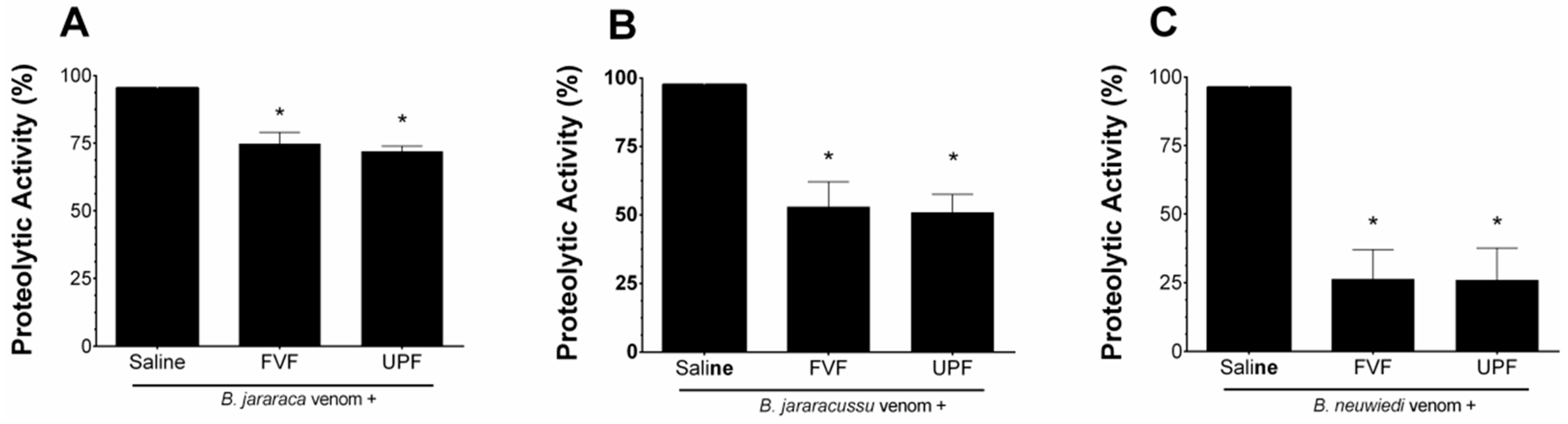
2.3. Inhibitory Effect of FVF and UPF on the Coagulant Activity of the Venom of B. jararaca, B. jararacussu, and B. neuwiedi
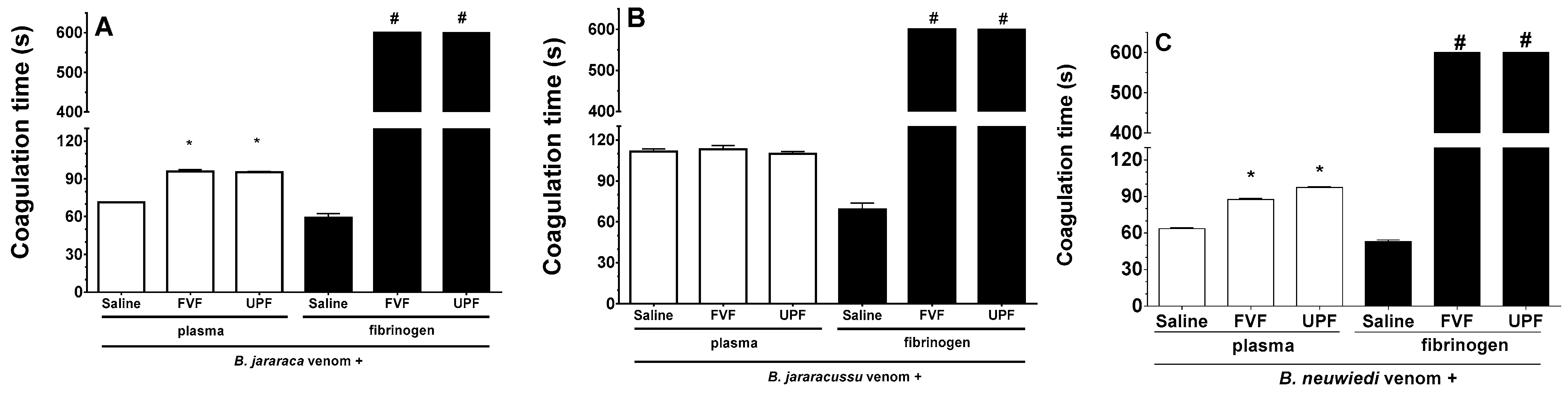
2.4. Inhibitory Effect of FVF and UPF on the PLA2 Activity of the Venom of B. jararaca, B. jararacussu, and B. neuwiedi
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Reagents and Venoms
5.2. Toxicity of FVP and UPF
5.3. Fucoidan Material
5.4. Effect of FVF and UPF on the Proteolytic Activity of B. jararaca, B. jararacussu, and B. neuwiedi Venom
5.5. Effect of FVF and UPF on the Coagulant Activity of B. jararaca, B. jararacussu, and B. neuwiedi Venom
5.6. Effect of FVF and UPF on the Phospholipase A2 Activity of B. jararaca, B. jararacussu, and B. neuwiedi Venom
5.7. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organization. Target Product Profiles for Animal Plasma-Derived Antivenoms: Antivenoms for Treatment of Snakebite Envenoming in Sub-Saharan Africa; WHO: Geneva, Switzerland, 2023; 25p. [Google Scholar]
- World Health Organization. WHO Guidelines for the Production, Control and Regulation of Snake Antivenom Immunoglobulins; WHO: Geneva, Switzerland, 2017; 388p. [Google Scholar]
- Williams, D.J.; Faiz, M.A.; Abela-Ridder, B.; Ainsworth, S.; Bulfone, T.C.; Nickerson, A.D.; Habib, A.G.; Junghanss, T.; Fan, H.W.; Turner, M.; et al. Strategy for a globally coordinated response to a priority neglected tropical disease: Snakebite envenoming. PLoS Negl. Trop. Dis. 2019, 13, e0007059. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, J.M.; Calvete, J.J.; Habib, A.G.; Harrison, R.A.; Williams, D.J.; Warrell, D.A. Snakebite envenoming. Nat. Rev. Dis. Primers 2017, 3, 17063. [Google Scholar] [CrossRef]
- Chippaux, J.P. Snakebite envenomation turns again into a neglected tropical disease! J. Venom. Anim. Toxins Incl. Trop. Dis. 2017, 23, 38–39. [Google Scholar] [CrossRef] [PubMed]
- Queiroz, G.P.; Pessoa, L.A.; Portaro, F.C.V.; Furtado, M.d.F.D.; Tambourgi, D.V. Interspecific variation in venom composition and toxicity of Brazilian snakes from Bothrops genus. Toxicon 2008, 52, 842–851. [Google Scholar] [CrossRef] [PubMed]
- Cavecci-Mendonça, B.; Luciano, K.M.; Vaccas, T.; de Oliveira, L.A.; Clemente, E.F.; Rossini, B.C.; Vieira, J.C.S.; de Barros, L.C.; Biondi, I.; de Magalhães, P.P.; et al. Preliminary insights of Brazilian snake venom metalloproteomics. Toxins 2023, 15, 648. [Google Scholar] [CrossRef] [PubMed]
- Chippaux, J.P. Incidence and mortality due to snakebite in the Americas. PLoS Negl. Trop. Dis. 2017, 11, e0005662. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, F.N.; Sachetto, A.T.A.; Santoro, M.L. Two-dimensional blue native/SDS polyacrylamide gel electrophoresis for analysis of Brazilian Bothrops snake venoms. Toxins 2022, 14, 661. [Google Scholar] [CrossRef] [PubMed]
- Camey, K.U.; Velarde, D.T.; Sanchez, E.F. Pharmacological characterization and neutralization of the venoms used in the production of Bothropic antivenom in Brazil. Toxicon 2002, 40, 501–509. [Google Scholar] [CrossRef] [PubMed]
- de Lima, M.E.; Fortes-Dias, C.L.; Carlini, C.R.; Guimarães, J.A. Toxinology in Brazil: A big challenge for a rich biodiversity. Toxicon 2010, 56, 1084–1091. [Google Scholar] [CrossRef]
- WHO. Snakebite Envenoming. 2021. Available online: https://www.who.int/news-room/fact-sheets/detail/snakebite-envenoming (accessed on 17 February 2021).
- Waiddyanatha, S.; Silva, A.; Siribaddana, S.; Isbister, G.K. Long-term effects of snake envenoming. Toxins 2019, 11, 193. [Google Scholar] [CrossRef]
- Solano, G.; Ainsworth, S.; Sánchez, A.; Villalta, M.; Sánchez, P.; Durán, G.; Gutiérrez, J.A.; León, G. Analysis of commercially available snake antivenoms reveals high contents of endotoxins in some products. Toxicon X 2024, 211, 100187–100189. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, J.M.; Albulescu, L.O.; Clare, R.H.; Casewell, N.R.; Abd El-Aziz, T.M.; Escalante, T.; Rucavado, A. The search for natural and synthetic inhibitors that would complement antivenoms as therapeutics for snakebite envenoming. Toxins 2021, 13, 451. [Google Scholar] [CrossRef]
- Marshall, W.R.; Soffa, A.N. Considerations for the development of a field-based medical device for the administration of adjunctive therapies for snakebite envenoming. Toxicon X 2023, 20, 100169–100190. [Google Scholar]
- Liaqat, A.; Mallhi, T.H.; Khan, Y.H.; Khokhar, A.; Chaman, S.; Ali, M. Anti-Snake venom properties of medicinal plants: A comprehensive systematic review of literature. Braz. J. Pharm. Sci. 2022, 58, e191124. [Google Scholar] [CrossRef]
- Adrião, A.A.X.; Dos Santos, A.O.; de Lima, E.J.S.P.; Maciel, J.B.; Paz, W.H.P.; da Silva, F.M.A.; Pucca, M.B.; Moura-da-Silva, A.M.; Monteiro, W.M.; Sartim, M.A.; et al. Plant-derived toxin inhibitors as potential candidates to complement antivenom treatment in snakebite envenomations. Front. Immunol. 2022, 9, 842576–842605. [Google Scholar] [CrossRef] [PubMed]
- Mekinić, I.G.; Skroza, D.; Šimat, V.; Hamed, I.; Čagalj, I.M.; Perković, Z.P. Phenolic content of brown algae (Pheophyceae) species: Extraction, identification, and quantification. Biomolecules 2019, 22, 244. [Google Scholar] [CrossRef]
- Lomartire, S.; Gonçalves, A.M.M. An overview of potential seaweed-derived bioactive compounds for pharmaceutical applications. Mar. Drugs 2022, 20, 141. [Google Scholar] [CrossRef]
- Costa, B.B.; Gianelli, J.L.D.; Moreira, T.A.; Soares, A.R.; Glauser, B.F.; Mourão, P.A.S.; Neto, C.; Barros, C.M.; Cinelli, L.P. Partial characterization and anticoagulant activity of sulfated galactan from the green seaweed Halimeda opuntia. An. Acad. Bras. Cienc. 2023, 95, e20211002. [Google Scholar] [CrossRef] [PubMed]
- Yinyi, F.; Jiao, H.; Sun, J.; Okoye, C.O.; Zhang, H.; Li, Y.; Lu, X.; Wang, Q.; Liu, J. Structure-activity relationships of bioactive polysaccharides extracted from macroalgae towards biomedical application: A review. Carbohydr. Polym. 2024, 324, 121533–121558. [Google Scholar]
- El-Sheekh, M.M.; Ward, F.; Deyab, M.A.; Al-Zahrani, M.; Touliabah, H.E. Chemical composition, antioxidant, and antitumor activity of fucoidan from the brown alga Dictyota dichotoma. Molecules 2023, 19, 7175. [Google Scholar] [CrossRef]
- Ferreira, L.G.; da Silva, A.C.R.; Noseda, M.D.; Fuly, A.L.; de Carvalho, M.M.; Fujii, M.T.; Sanchez, E.F.; Carneiro, J.; Duarte, M.E.R. Chemical structure and snake antivenom properties of sulfated agarans obtained from Laurencia dendroidea (Ceramiales, Rhodophyta). Carbohydr. Polym. 2019, 15, 136–144. [Google Scholar] [CrossRef] [PubMed]
- da Silva, A.C.R.; Ferreira, L.G.; Duarte, M.E.R.; Noseda, M.D.; Sanchez, E.F.; Fuly, A.L. Sulfated Galactan from Palisada flagellifera Inhibits Toxic Effects of Lachesis muta Snake Venom. Mar. Drugs 2015, 13, 3761–3775. [Google Scholar] [CrossRef] [PubMed]
- da Silva, A.C.R.; Pereira, K.K.G.; Critchley, A.T.; Sanchez, E.F.; Fuly, A.L. Potential utilization of a lambda carrageenan polysaccharide, derived from a cultivated, clonal strain of the red seaweed Chondrus crispus (Irish moss) against toxic actions of venom of Bothrops jararaca and B. jararacussu snakes. J. Appl. Phycol. 2020, 32, 4309–4320. [Google Scholar] [CrossRef]
- da Silva, A.C.R.; Duarte, M.E.R.; Noseda, M.D.; Ferreira, L.G.; Cassolato, J.E.F.; Sanchez, E.F.; Fuly, A.L. Potential utilization of a polysaccharide from the marine algae Gayralia oxysperma, as an antivenom for Viperidae snakebites. Mar. Drugs 2018, 16, 412. [Google Scholar] [CrossRef] [PubMed]
- Obluchinskaya, E.D.; Pozharitskaya, O.N.; Shevyrin, V.A.; Kovaleva, E.G.; Flisyuk, E.V.; Shikov, A.N. Optimization of Extraction of phlorotannins from the Arctic Fucus vesiculosus using natural deep eutectic solvents and their HPLC profiling with Tandem High-Resolution Mass Spectrometry. Mar. Drugs 2023, 21, 263. [Google Scholar] [CrossRef] [PubMed]
- Tagliapietra, B.L.; Clerici, M.T.P.S. Brown algae and their multiple applications as functional ingredient in food production. Food Res. Int. 2023, 167, 112655–112665. [Google Scholar] [CrossRef] [PubMed]
- Azofeifa, K.; Angulo, Y.; Lomonte, B. Ability of fucoidan to prevent muscle necrosis induced by snake venom myotoxins: Comparison of high- and low-molecular weight fractions. Toxicon 2008, 51, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Angulo, Y.; Lomonte, B. Inhibitory effect of fucoidan on the activities of crotaline snake venom myotoxic phospholipases A(2). Biochem. Pharmacol. 2003, 66, 1993–2000. [Google Scholar] [CrossRef] [PubMed]
- Bauer, M.; Lautenschlaeger, C.; Kempe, K.; Tauhardt, L.; Schubert, U.S.; Fischer, D. Poly(2-ethyl-2-oxazoline) as alternative for the stealth polymer poly(ethylene glycol): Comparison of in vitro cytotoxicity and hemocompatibility. Macromol. Biosci. 2012, 12, 986–998. [Google Scholar] [CrossRef]
- Olaoba, O.T.; dos Santos, P.K.; Selistre-de-Arujo, H.S.; de Souza, D.H.F. Snake Venom Metalloproteinases (SVMPs): A structure-function update. Toxicon X 2020, 21, 100052–100067. [Google Scholar] [CrossRef]
- Serrano, S.M. The long road of research on snake venom serine proteinases. Toxicon 2013, 62, 19–26. [Google Scholar] [CrossRef] [PubMed]
- de Paula, R.C.; Castro, H.C.; Rodrigues, C.R.; Melo, P.A.; Fuly, A.L. Structural and pharmacological features of phospholipases A2 from snake venoms. Protein Pept. Lett. 2009, 16, 899–907. [Google Scholar] [CrossRef] [PubMed]
- Aloulou, A.; Rahier, R.; Arhab, Y.; Noiriel, A.; Abousalham, A. Phospholipases: An Overview. Methods Mol. Biol. 2018, 1835, 69–105. [Google Scholar] [PubMed]
- Laing, G.D.; Moura-da-Silva, A.M. Jararhagin and its multiple effects on hemostasis. Toxicon 2005, 45, 987–996. [Google Scholar] [CrossRef] [PubMed]
- Molina, D.A.; Guerra-Duarte, C.; Naves de Souza, D.L.; Costal-Oliveira, F.; Ávila, G.R.; Soccol, V.T.; Machado-de-Ávila, R.A.; Chávez-Olórtegui, C. Identification of a linear B-cell epitope in the catalytic domain of bothropasin, a metalloproteinase from Bothrops jararaca snake venom. Mol. Immunol. 2018, 104, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, F.F.; Salvador, G.H.M.; Cavalcante, W.L.G.; Dal-Pai, M.; Fontes, M.R.M. BthTX-I, a phospholipase A2-like toxin, is inhibited by the plant cinnamic acid derivative: Chlorogenic acid. Biochim. Biophys. Acta Proteins Proteom. 2024, 1872, 140988–141001. [Google Scholar] [CrossRef] [PubMed]
- Borges, R.J.; Salvador, G.H.M.; Campanelli, H.B.; Pimenta, D.C.; de Oliveira, N.M.; Usón, I.; Fontes, M.R.M. BthTX-II from Bothrops jararacussu venom has variants with different oligomeric assemblies: An example of snake venom phospholipases A2 versatility. Int. J. Biol. Macromol. 2021, 191, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Belo, A.A.; de Souza, D.L.N.; de Melo-Braga, M.N.; de Souza, L.L.; Molina, M.D.A.; de Melo, P.D.V.; Larsen, M.R.; Guerra-Duarte, C.; Chávez-Olórtegui, C. Production of a murine mAb against Bothrops alternatus and B. neuwiedi snake venoms and its use to isolate a thrombin-like serine protease fraction. Int. J. Biol. Macromol. 2022, 214, 530–541. [Google Scholar] [CrossRef] [PubMed]
- Thumtecho, S.; Burlet, N.J.; Ljungars, A.; Laustsen, A.H. Towards better antivenoms: Navigating the road to new types of snakebite envenoming therapies. J. Venom. Anim. Toxins Incl. Trop. Dis. 2023, 29, e20230057. [Google Scholar] [CrossRef]
- Haggag, Y.A.; Elrahman, A.A.A.; Ulber, R.; Zayed, A. Fucoidan in Pharmaceutical Formulations: A Comprehensive Review for Smart Drug Delivery Systems. Mar. Drugs 2023, 21, 112. [Google Scholar] [CrossRef]
- Trujillo, E.; Monreal-Escalante, E.; Ramos-Vega, A.; Ângulo, C. Macroalgae: Marine players in vaccinology. Algal Res. 2024, 78, 103392–103403. [Google Scholar] [CrossRef]
- Pozharitskaya, O.N.; Shikov, A.N.; Obluchinskaya, E.D.; Vuorela, H. The pharmacokinetics of fucoidan after topical application to rats. Mar. Drugs 2019, 17, 687. [Google Scholar] [CrossRef]
- Shikov, A.N.; Flisyuk, E.V.; Obluchinskaya, E.D.; Pozharitskaya, O.N. Pharmacokinetics of marine-derived drugs. Mar. Drugs 2020, 18, 557. [Google Scholar] [CrossRef] [PubMed]
- George, A.; Shrivastav, P.S. Fucoidan, a brown seaweed polysaccharide in nanodrug delivery. Drug Deliv. Transl. Res. 2023, 13, 2427–2446. [Google Scholar] [CrossRef] [PubMed]
- Ullah, A.; Masood, R.; Ali, I.; Ullah, K.; Ali, H.; Akbar, H.; Betzel, C. Thrombin-like enzymes from snake venom: Structural characterization and mechanism of action. Int. J. Biol. Macromol. 2018, 15, 788–811. [Google Scholar] [CrossRef]
- da Silva, G.M.; de Souza, D.H.B.; Waitman, K.B.; Ebram, M.C.; Fessel, M.R.; Zainescu, I.C.; Portaro, F.C.; Heras, M.; de Andrade, S.A. Design, synthesis, and evaluation of Bothrops venom serine protease peptidic inhibitors. J. Venom. Anim. Toxins Incl. Trop. Dis. 2021, 27, e20200066. [Google Scholar] [CrossRef] [PubMed]
- Fuly, A.L.; Calil-Elias, S.; Zingali, R.B.; Guimarães, J.A.; Melo, P.A. Myotoxic activity of an acidic phospholipase A2 isolated from Lachesis muta (Bushmaster) snake venom. Toxicon 2000, 38, 961–972. [Google Scholar] [CrossRef]
- Fuly, A.L.; de Miranda, A.L.; Zingali, R.B.; Guimarães, J.A. Purification and characterization of a phospholipase A2 isoenzyme isolated from Lachesis muta snake venom. Biochem. Pharmacol. 2002, 63, 1589–1597. [Google Scholar] [CrossRef]
- Lewin, M.R.; Carter, R.W.; Matteo, I.A.; Samuel, S.P.; Rao, S.; Fry, B.G.; Bickler, P.E. Varespladib in the Treatment of Snakebite Envenoming: Development History and Preclinical Evidence Supporting Advancement to Clinical Trials in Patients Bitten by Venomous Snakes. Toxins 2022, 14, 783. [Google Scholar] [CrossRef]
- Salvador, G.H.M.; Pinto, Ê.K.R.; Ortolani, P.L.; Fortes-Dias, C.L.; Cavalcante, W.L.G.; Soares, A.M.; Lomonte, B.; Lewin, M.R.; Fontes, M.R.M. Structural basis of the myotoxic inhibition of the Bothrops pirajai PrTX-I by the synthetic Varespladib. Biochimie 2023, 207, 1–10. [Google Scholar] [CrossRef]
- Lewin, M.; Samuel, S.; Merkel, J.; Bickler, P. Varespladib (LY315920) Appears to Be a Potent, Broad-Spectrum, Inhibitor of Snake Venom Phospholipase A2 and a Possible Pre-Referral Treatment for Envenomation. Toxins 2016, 25, 248. [Google Scholar] [CrossRef] [PubMed]
- Re: GRAS Notice, No. GRN 000661. Available online: https://www.fda.gov/downloads/Food/IngredientsPackagingLabeling/GRAS/NoticeInventory/ucm549588.pdf (accessed on 10 February 2024).
- Citkowska, A.; Szekalska, M.; Winnicka, K. Possibilities of Fucoidan Utilization in the Development of Pharmaceutical Dosage Forms. Mar. Drugs 2019, 17, 458. [Google Scholar] [CrossRef] [PubMed]
- DuBois, M.K.; Gilles, K.J.; Hamilton, J.K.; Rebers, P.A.; Smith, F.S. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Filisetti-Cozzi, T.M.; Carpita, N.C. Measurement of uronic acids without interference from neutral sugars. Anal. Biochem. 1991, 97, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Dodgson, K.S.; Price, R.G. A note on the determination of the ester sulphate content of sulphated polysaccharides. Biochem. J. 1962, 84, 106–110. [Google Scholar] [CrossRef] [PubMed]
- Garcia, E.S.; Guimarães, J.A.; Prado, J.L. Purification and characterization of a sulfhydryl-dependent protease from Rhodnius prolixus midgut. Arch. Biochem. Biophys. 1978, 188, 315–322. [Google Scholar] [CrossRef]
- Marinetti, G.V. The action of phospholipase A2 on lipoproteins. Biochim. Biophys. Acta Lipids Lipid Metab. 1965, 98, 554–565. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Components | FVF | UPF |
|---|---|---|
| Fucoidan | 96.1 | 97 |
| Total carbohydrates | 71.7 | 60.5 |
| Fucose | 49.5 | 28.2 |
| Galactose | 2.7 | 25.6 |
| Uronic acid | 3.8 | 0.8 |
| Polyphenol | <2 | <2 |
| Sulfate | 30.7 | 30.4 |
| Cations | ~5 | 6.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castro-Pinheiro, C.; Junior, L.C.S.P.; Sanchez, E.F.; da Silva, A.C.R.; Dwan, C.A.; Karpiniec, S.S.; Critchley, A.T.; Fuly, A.L. Effect of Seaweed-Derived Fucoidans from Undaria pinnatifida and Fucus vesiculosus on Coagulant, Proteolytic, and Phospholipase A2 Activities of Snake Bothrops jararaca, B. jararacussu, and B. neuwiedi Venom. Toxins 2024, 16, 188. https://doi.org/10.3390/toxins16040188
Castro-Pinheiro C, Junior LCSP, Sanchez EF, da Silva ACR, Dwan CA, Karpiniec SS, Critchley AT, Fuly AL. Effect of Seaweed-Derived Fucoidans from Undaria pinnatifida and Fucus vesiculosus on Coagulant, Proteolytic, and Phospholipase A2 Activities of Snake Bothrops jararaca, B. jararacussu, and B. neuwiedi Venom. Toxins. 2024; 16(4):188. https://doi.org/10.3390/toxins16040188
Chicago/Turabian StyleCastro-Pinheiro, Camila, Luiz Carlos Simas Pereira Junior, Eladio Flores Sanchez, Ana Cláudia Rodrigues da Silva, Corinna A. Dwan, Samuel S. Karpiniec, Alan Trevor Critchley, and Andre Lopes Fuly. 2024. "Effect of Seaweed-Derived Fucoidans from Undaria pinnatifida and Fucus vesiculosus on Coagulant, Proteolytic, and Phospholipase A2 Activities of Snake Bothrops jararaca, B. jararacussu, and B. neuwiedi Venom" Toxins 16, no. 4: 188. https://doi.org/10.3390/toxins16040188
APA StyleCastro-Pinheiro, C., Junior, L. C. S. P., Sanchez, E. F., da Silva, A. C. R., Dwan, C. A., Karpiniec, S. S., Critchley, A. T., & Fuly, A. L. (2024). Effect of Seaweed-Derived Fucoidans from Undaria pinnatifida and Fucus vesiculosus on Coagulant, Proteolytic, and Phospholipase A2 Activities of Snake Bothrops jararaca, B. jararacussu, and B. neuwiedi Venom. Toxins, 16(4), 188. https://doi.org/10.3390/toxins16040188