Ionophores: Potential Use as Anticancer Drugs and Chemosensitizers

Abstract
:1. Introduction
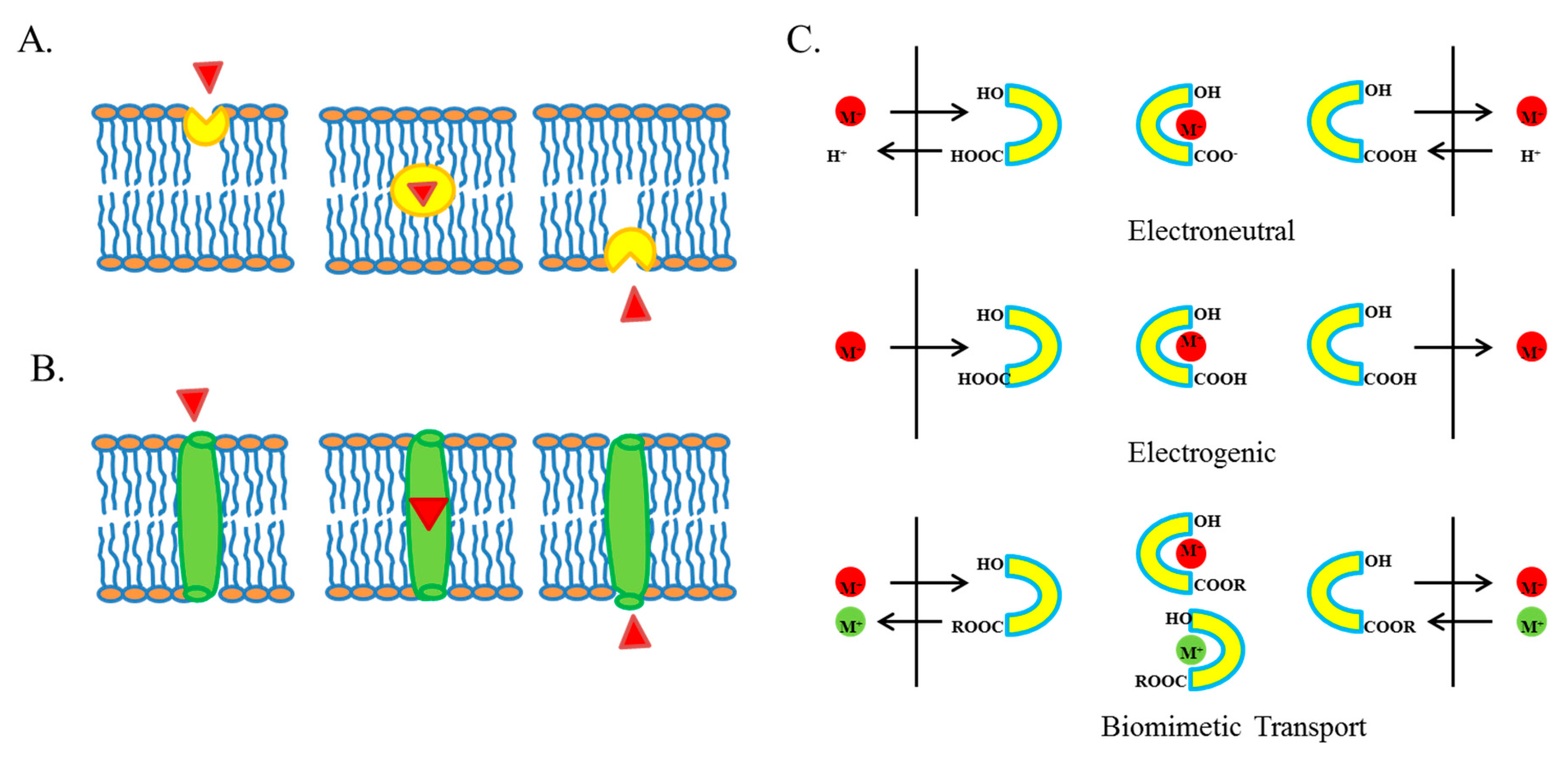
Ion Transport
2. Ion Transport in Cancer Cells and Its Targeting to Develop Novel Anticancer Therapies
2.1. Ion Transport in Cancer Cells
2.2. Ion Transport and Chemotherapy
3. Ionophores
3.1. Salinomycin (SAL)
3.2. Nigericin (NIG)
3.3. Obatoclax (OBT)
4. Ionophores in Ongoing Clinical Trials for Cancer Treatment
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Govorunova, E.G.; Sineshchekov, O.A.; Janz, R.; Liu, X.; Spudich, J.L. Natural light-gated anion channels: A family of microbial rhodopsins for advanced optogenetics. Science 2015, 349, 647–650. [Google Scholar] [CrossRef] [PubMed]
- Alberts, B.; Johnson, A.; Lewis, J.; Raff, M.; Roberts, K.; Walter, P. Carrier Proteins and Active Membrane Transport. In Molecular Biology of the Cell, 4th ed.; Garland Science: New York, NY, USA, 2002. [Google Scholar]
- Manago, A.; Leanza, L.; Carraretto, L.; Sassi, N.; Grancara, S.; Quintana-Cabrera, R.; Trimarco, V.; Toninello, A.; Scorrano, L.; Trentin, L.; et al. Early effects of the antineoplastic agent salinomycin on mitochondrial function. Cell Death Dis. 2015, 6, e1930. [Google Scholar] [CrossRef] [PubMed]
- Boesch, M.; Zeimet, A.G.; Rumpold, H.; Gastl, G.; Sopper, S.; Wolf, D. Drug Transporter-Mediated Protection of Cancer Stem Cells from Ionophore Antibiotics. Stem Cells Transl. Med. 2015, 4, 1028–1032. [Google Scholar] [CrossRef] [PubMed]
- Deng, C.C.; Liang, Y.; Wu, M.S.; Feng, F.T.; Hu, W.R.; Chen, L.Z.; Feng, Q.S.; Bei, J.X.; Zeng, Y.X. Nigericin selectively targets cancer stem cells in nasopharyngeal carcinoma. Int. J. Biochem. Cell Biol. 2013, 45, 1997–2006. [Google Scholar] [CrossRef] [PubMed]
- Yakisich, J.S.; Azad, N.; Kaushik, V.; O’Doherty, G.A.; Iyer, A.K. Nigericin decreases the viability of multidrug-resistant cancer cells and lung tumorspheres and potentiates the effects of cardiac glycosides. Tumour Biol. 2017, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, P.B.; Onder, T.T.; Jiang, G.; Tao, K.; Kuperwasser, C.; Weinberg, R.A.; Lander, E.S. Identification of selective inhibitors of cancer stem cells by high-throughput screening. Cell 2009, 138, 645–659. [Google Scholar] [CrossRef] [PubMed]
- Magrath, J.W.; Kim, Y. Salinomycin’s potential to eliminate glioblastoma stem cells and treat glioblastoma multiforme (Review). Int. J. Oncol. 2017, 51, 753–759. [Google Scholar] [CrossRef] [PubMed]
- Naujokat, C.; Steinhart, R. Salinomycin as a drug for targeting human cancer stem cells. J. Biomed. Biotechnol 2012, 2012, 950658. [Google Scholar] [CrossRef] [PubMed]
- Berghauser Pont, L.M.; Spoor, J.K.; Venkatesan, S.; Swagemakers, S.; Kloezeman, J.J.; Dirven, C.M.; van der Spek, P.J.; Lamfers, M.L.; Leenstra, S. The Bcl-2 inhibitor Obatoclax overcomes resistance to histone deacetylase inhibitors SAHA and LBH589 as radiosensitizers in patient-derived glioblastoma stem-like cells. Genes Cancer 2014, 5, 445–459. [Google Scholar] [PubMed]
- Blatt, K.; Herrmann, H.; Stefanzl, G.; Sperr, W.R.; Valent, P. Evaluation of in vitro effects of various targeted drugs on plasma cells and putative neoplastic stem cells in patients with multiple myeloma. Oncotarget 2016, 7, 65627–65642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruickshanks, N.; Hamed, H.A.; Booth, L.; Tavallai, S.; Syed, J.; Sajithlal, G.B.; Grant, S.; Poklepovic, A.; Dent, P. Histone deacetylase inhibitors restore toxic BH3 domain protein expression in anoikis-resistant mammary and brain cancer stem cells, thereby enhancing the response to anti-ERBB1/ERBB2 therapy. Cancer Biol. Ther. 2013, 14, 982–996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishida, C.T.; Bianchetti, E.; Shu, C.; Halatsch, M.E.; Westhoff, M.A.; Karpel-Massler, G.; Siegelin, M.D. BH3-mimetics and BET-inhibitors elicit enhanced lethality in malignant glioma. Oncotarget 2017, 8, 29558–29573. [Google Scholar] [CrossRef] [PubMed]
- Lagadinou, E.D.; Sach, A.; Callahan, K.; Rossi, R.M.; Neering, S.J.; Minhajuddin, M.; Ashton, J.M.; Pei, S.; Grose, V.; O’Dwyer, K.M.; et al. BCL-2 inhibition targets oxidative phosphorylation and selectively eradicates quiescent human leukemia stem cells. Cell Stem Cell 2013, 12, 329–341. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Bora-Singhal, N.; Kroeger, J.; Laklai, H.; Chellappan, S.P. βArrestin-1 and Mcl-1 modulate self-renewal growth of cancer stem-like side-population cells in non-small cell lung cancer. PLoS ONE 2013, 8, e55982. [Google Scholar] [CrossRef] [PubMed]
- David, J.M.; Owens, T.A.; Inge, L.J.; Bremner, R.M.; Rajasekaran, A.K. Gramicidin A blocks tumor growth and angiogenesis through inhibition of hypoxia-inducible factor in renal cell carcinoma. Mol. Cancer Ther. 2014, 13, 788–799. [Google Scholar] [CrossRef] [PubMed]
- Rao, D.K.; Liu, H.; Ambudkar, S.V.; Mayer, M. A combination of curcumin with either gramicidin or ouabain selectively kills cells that express the multidrug resistance-linked ABCG2 transporter. J. Biol. Chem. 2014, 289, 31397–31410. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Tie, X.; Meng, L.; Wang, Y.; Wu, A. PMA and ionomycin induce glioblastoma cell death: Activation-induced cell-death-like phenomena occur in glioma cells. PLoS ONE 2013, 8, e76717. [Google Scholar] [CrossRef] [PubMed]
- Miyake, H.; Hara, I.; Yamanaka, K.; Arakawa, S.; Kamidono, S. Calcium ionophore, ionomycin inhibits growth of human bladder cancer cells both in vitro and in vivo with alteration of Bcl-2 and Bax expression levels. J. Urol 1999, 162, 916–921. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Zhang, J.; Wang, Z.; Yan, Z.; Qiao, M.; Ye, J.; Wei, Q.; Wang, J.; Wang, X.; Zhao, L.; et al. Antibiotic monensin synergizes with EGFR inhibitors and oxaliplatin to suppress the proliferation of human ovarian cancer cells. Sci. Rep. 2015, 5, 17523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.H.; Kim, K.Y.; Yu, S.N.; Park, S.G.; Yu, H.S.; Seo, Y.K.; Ahn, S.C. Monensin Induces PC-3 Prostate Cancer Cell Apoptosis via ROS Production and Ca2+ Homeostasis Disruption. Anticancer Res. 2016, 36, 5835–5843. [Google Scholar] [CrossRef] [PubMed]
- Tumova, L.; Pombinho, A.R.; Vojtechova, M.; Stancikova, J.; Gradl, D.; Krausova, M.; Sloncova, E.; Horazna, M.; Kriz, V.; Machonova, O.; et al. Monensin inhibits canonical Wnt signaling in human colorectal cancer cells and suppresses tumor growth in multiple intestinal neoplasia mice. Mol. Cancer Ther. 2014, 13, 812–822. [Google Scholar] [CrossRef] [PubMed]
- Ryoo, I.J.; Park, H.R.; Choo, S.J.; Hwang, J.H.; Park, Y.M.; Bae, K.H.; Shin-Ya, K.; Yoo, I.D. Selective cytotoxic activity of valinomycin against HT-29 Human colon carcinoma cells via down-regulation of GRP78. Biol. Pharm. Bull. 2006, 29, 817–820. [Google Scholar] [CrossRef] [PubMed]
- Daoud, S.S.; Forde, N.H. Synergistic cytotoxic actions of cisplatin and liposomal valinomycin on human ovarian carcinoma cells. Cancer Chemother. Pharmacol. 1991, 28, 370–376. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.Y.; Kim, S.H.; Yu, S.N.; Park, S.G.; Kim, Y.W.; Nam, H.W.; An, H.H.; Yu, H.S.; Kim, Y.W.; Ji, J.H.; et al. Lasalocid induces cytotoxic apoptosis and cytoprotective autophagy through reactive oxygen species in human prostate cancer PC-3 cells. Biomed. Pharmacother. 2017, 88, 1016–1024. [Google Scholar] [CrossRef] [PubMed]
- Dornetshuber, R.; Heffeter, P.; Kamyar, M.R.; Peterbauer, T.; Berger, W.; Lemmens-Gruber, R. Enniatin exerts p53-dependent cytostatic and p53-independent cytotoxic activities against human cancer cells. Chem. Res. Toxicol. 2007, 20, 465–473. [Google Scholar] [CrossRef] [PubMed]
- Dornetshuber-Fleiss, R.; Heilos, D.; Mohr, T.; Richter, L.; Sussmuth, R.D.; Zlesak, M.; Novicky, A.; Heffeter, P.; Lemmens-Gruber, R.; Berger, W. The naturally born fusariotoxin enniatin B and sorafenib exert synergistic activity against cervical cancer in vitro and in vivo. Biochem. Pharmacol. 2015, 93, 318–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, C.L.; Lin, H.I.; Chen, B.F.; Jow, G.M. Beauvericin-induced cell apoptosis through the mitogen-activated protein kinase pathway in human nonsmall cell lung cancer A549 cells. J. Toxicol. Sci. 2016, 41, 429–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watjen, W.; Debbab, A.; Hohlfeld, A.; Chovolou, Y.; Proksch, P. The mycotoxin beauvericin induces apoptotic cell death in H4IIE hepatoma cells accompanied by an inhibition of NF-κB-activity and modulation of MAP-kinases. Toxicol. Lett. 2014, 231, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Prevarskaya, N.; Skryma, R.; Shuba, Y. Ion channels and the hallmarks of cancer. Trends Mol. Med. 2010, 16, 107–121. [Google Scholar] [CrossRef] [PubMed]
- Roderick, H.L.; Cook, S.J. Ca2+ signalling checkpoints in cancer: Remodelling Ca2+ for cancer cell proliferation and survival. Nat. Rev. Cancer 2008, 8, 361–375. [Google Scholar] [CrossRef] [PubMed]
- Kahl, C.R.; Means, A.R. Regulation of cell cycle progression by calcium/calmodulin-dependent pathways. Endocr. Rev. 2003, 24, 719–736. [Google Scholar] [CrossRef] [PubMed]
- Thebault, S.; Flourakis, M.; Vanoverberghe, K.; Vandermoere, F.; Roudbaraki, M.; Lehen’kyi, V.; Slomianny, C.; Beck, B.; Mariot, P.; Bonnal, J.L.; et al. Differential role of transient receptor potential channels in Ca2+ entry and proliferation of prostate cancer epithelial cells. Cancer Res. 2006, 66, 2038–2047. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.; Wang, X.; Chen, M.; Ouyang, K.; Zheng, M.; Cheng, H. Flickering calcium microdomains signal turning of migrating cells. Can. J. Physiol. Pharmacol. 2010, 88, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Deliot, N.; Constantin, B. Plasma membrane calcium channels in cancer: Alterations and consequences for cell proliferation and migration. Biochim. Biophys. Acta 2015, 1848 Pt B, 2512–2522. [Google Scholar] [CrossRef]
- Dubois, C.; Vanden Abeele, F.; Prevarskaya, N. Targeting apoptosis by the remodelling of calcium-transporting proteins in cancerogenesis. FEBS J. 2013, 280, 5500–5510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orrenius, S.; Zhivotovsky, B.; Nicotera, P. Regulation of cell death: The calcium-apoptosis link. Nat. Rev. Mol. Cell Biol. 2003, 4, 552–565. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, E.K.; Lambert, I.H. Ion channels and transporters in the development of drug resistance in cancer cells. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369, 20130109. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z. Roles of K+ channels in regulating tumour cell proliferation and apoptosis. Pflugers Arch. 2004, 448, 274–286. [Google Scholar] [CrossRef] [PubMed]
- Fraser, S.P.; Ozerlat-Gunduz, I.; Brackenbury, W.J.; Fitzgerald, E.M.; Campbell, T.M.; Coombes, R.C.; Djamgoz, M.B. Regulation of voltage-gated sodium channel expression in cancer: Hormones, growth factors and auto-regulation. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369, 20130105. [Google Scholar] [CrossRef] [PubMed]
- Lopez, V.; Kelleher, S.L. Zip6-attenuation promotes epithelial-to-mesenchymal transition in ductal breast tumor (T47D) cells. Exp. Cell Res. 2010, 316, 366–375. [Google Scholar] [CrossRef] [PubMed]
- Matsui, C.; Takatani-Nakase, T.; Hatano, Y.; Kawahara, S.; Nakase, I.; Takahashi, K. Zinc and its transporter ZIP6 are key mediators of breast cancer cell survival under high glucose conditions. FEBS Lett. 2017, 591, 3348–3359. [Google Scholar] [CrossRef] [PubMed]
- Donadelli, M.; Dalla Pozza, E.; Scupoli, M.T.; Costanzo, C.; Scarpa, A.; Palmieri, M. Intracellular zinc increase inhibits p53(−/−) pancreatic adenocarcinoma cell growth by ROS/AIF-mediated apoptosis. Biochim. Biophys. Acta 2009, 1793, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wei, X.; Yan, P.; Han, Y.; Sun, S.; Wu, K.; Fan, D. Human mitochondrial Mrs2 protein promotes multidrug resistance in gastric cancer cells by regulating p27, cyclin D1 expression and cytochrome C release. Cancer Biol. Ther. 2009, 8, 607–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castiglioni, S.; Cazzaniga, A.; Trapani, V.; Cappadone, C.; Farruggia, G.; Merolle, L.; Wolf, F.I.; Iotti, S.; Maier, J.A. Magnesium homeostasis in colon carcinoma LoVo cells sensitive or resistant to doxorubicin. Sci. Rep. 2015, 5, 16538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furukawa, T.; Komatsu, M.; Ikeda, R.; Tsujikawa, K.; Akiyama, S. Copper transport systems are involved in multidrug resistance and drug transport. Curr. Med. Chem. 2008, 15, 3268–3278. [Google Scholar] [CrossRef] [PubMed]
- Bompiani, K.M.; Tsai, C.Y.; Achatz, F.P.; Liebig, J.K.; Howell, S.B. Copper transporters and chaperones CTR1, CTR2, ATOX1, and CCS as determinants of cisplatin sensitivity. Metallomics 2016, 8, 951–962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.Y.; Choi, C.H.; Do, I.G.; Song, S.Y.; Lee, W.; Park, H.S.; Song, T.J.; Kim, M.K.; Kim, T.J.; Lee, J.W.; et al. Prognostic value of the copper transporters, CTR1 and CTR2, in patients with ovarian carcinoma receiving platinum-based chemotherapy. Gynecol. Oncol. 2011, 122, 361–365. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, T.; Inoue, Y.; Kodama, H.; Yamazaki, H.; Kawai, K.; Suemizu, H.; Masuda, R.; Iwazaki, M.; Yamada, S.; Ueyama, Y.; et al. Expression of copper-transporting p-type adenosine triphosphatase (ATP7B) correlates with cisplatin resistance in human non-small cell lung cancer xenografts. Oncol. Rep. 2008, 20, 265–270. [Google Scholar] [PubMed]
- Samimi, G.; Safaei, R.; Katano, K.; Holzer, A.K.; Rochdi, M.; Tomioka, M.; Goodman, M.; Howell, S.B. Increased expression of the copper efflux transporter ATP7A mediates resistance to cisplatin, carboplatin, and oxaliplatin in ovarian cancer cells. Clin. Cancer Res. 2004, 10, 4661–4669. [Google Scholar] [CrossRef] [PubMed]
- Ober, S.S.; Pardee, A.B. Intracellular pH is increased after transformation of Chinese hamster embryo fibroblasts. Proc. Natl. Acad. Sci. USA 1987, 84, 2766–2770. [Google Scholar] [CrossRef] [PubMed]
- Larsen, A.K.; Escargueil, A.E.; Skladanowski, A. Resistance mechanisms associated with altered intracellular distribution of anticancer agents. Pharmacol. Ther. 2000, 85, 217–229. [Google Scholar] [CrossRef]
- Cardone, R.A.; Casavola, V.; Reshkin, S.J. The role of disturbed pH dynamics and the Na+/H+ exchanger in metastasis. Nat. Rev. Cancer 2005, 5, 786–795. [Google Scholar] [CrossRef] [PubMed]
- Abdoul-Azize, S.; Dubus, I.; Vannier, J.P. Improvement of dexamethasone sensitivity by chelation of intracellular Ca2+ in pediatric acute lymphoblastic leukemia cells through the prosurvival kinase ERK1/2 deactivation. Oncotarget 2017, 8, 27339–27352. [Google Scholar] [CrossRef] [PubMed]
- Hsia, T.C.; Huang, Y.P.; Jiang, Y.W.; Chen, H.Y.; Cheng, Z.Y.; Hsiao, Y.T.; Chen, C.Y.; Peng, S.F.; Chueh, F.S.; Chou, Y.C.; et al. Phenethyl Isothiocyanate Induces Apoptotic Cell Death Through the Mitochondria-dependent Pathway in Gefitinib-resistant NCI-H460 Human Lung Cancer Cells In Vitro. Anticancer Res. 2018, 38, 2137–2147. [Google Scholar] [PubMed]
- Chen, S.J.; Kuo, C.C.; Pan, H.Y.; Tsou, T.C.; Yeh, S.C.; Chang, J.Y. Desferal regulates hCtr1 and transferrin receptor expression through Sp1 and exhibits synergistic cytotoxicity with platinum drugs in oxaliplatin-resistant human cervical cancer cells in vitro and in vivo. Oncotarget 2016, 7, 49310–49321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, E.L.; Shimizu, T.; Ise, T.; Numata, T.; Kohno, K.; Okada, Y. Impaired activity of volume-sensitive Cl-channel is involved in cisplatin resistance of cancer cells. J. Cell. Physiol. 2007, 211, 513–521. [Google Scholar] [CrossRef] [PubMed]
- Goh, W.; Sleptsova-Freidrich, I.; Petrovic, N. Use of proton pump inhibitors as adjunct treatment for triple-negative breast cancers. An introductory study. J. Pharm. Pharm. Sci. 2014, 17, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Kaushik, V.; Yakisich, J.S.; Azad, N.; Kulkarni, Y.; Venkatadri, R.; Wright, C.; Rojanasakul, Y.; Iyer, A.K.V. Anti-Tumor Effects of Cardiac Glycosides on Human Lung Cancer Cells and Lung Tumorspheres. J. Cell. Physiol. 2017, 232, 2497–2507. [Google Scholar] [CrossRef] [PubMed]
- Kaushik, V.; Azad, N.; Yakisich, J.S.; Iyer, A.K. Antitumor effects of naturally occurring cardiac glycosides convallatoxin and peruvoside on human ER+ and triple-negative breast cancers. Cell Death Discov. 2017, 3, 17009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulkarni, Y.M.; Kaushik, V.; Azad, N.; Wright, C.; Rojanasakul, Y.; O’Doherty, G.; Iyer, A.K. Autophagy-Induced Apoptosis in Lung Cancer Cells by a Novel Digitoxin Analog. J. Cell. Physiol. 2016, 231, 817–828. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, Y.M.; Yakisich, J.S.; Azad, N.; Venkatadri, R.; Kaushik, V.; O’Doherty, G.; Iyer, A.K.V. Anti-tumorigenic effects of a novel digitoxin derivative on both estrogen receptor-positive and triple-negative breast cancer cells. Tumour Biol. 2017, 39. [Google Scholar] [CrossRef] [PubMed]
- Babcock, J.J.; Li, M. hERG channel function: Beyond long QT. Acta Pharmacol. Sin. 2013, 34, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Pillozzi, S.; Masselli, M.; De Lorenzo, E.; Accordi, B.; Cilia, E.; Crociani, O.; Amedei, A.; Veltroni, M.; D’Amico, M.; Basso, G.; et al. Chemotherapy resistance in acute lymphoblastic leukemia requires hERG1 channels and is overcome by hERG1 blockers. Blood 2011, 117, 902–914. [Google Scholar] [CrossRef] [PubMed]
- Arcangeli, A.; Becchetti, A. Novel perspectives in cancer therapy: Targeting ion channels. Drug Resist. Updat. 2015, 21–22, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Yakisich, J.S.; Kulkarni, Y.; Azad, N.; Iyer, A.K.V. Selective and Irreversible Induction of Necroptotic Cell Death in Lung Tumorspheres by Short-Term Exposure to Verapamil in Combination with Sorafenib. Stem Cells Int. 2017, 2017, 5987015. [Google Scholar] [CrossRef] [PubMed]
- Helson, L. Calcium channel blocker enhancement of anticancer drug cytotoxicity—A review. Cancer Drug Deliv. 1984, 1, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Vilpo, J.; Koski, T.; Vilpo, L. Calcium antagonists potentiate p-glycoprotein-independent anticancer drugs in chronic lymphocytic leukemia cells in vitro. Haematologica 2000, 85, 806–813. [Google Scholar] [PubMed]
- Hilgenfeld, R.; Saenger, W. Structural chemistry of natural and synthetic ionophores and their complexes with cations. Top. Curr. Chem. 1982, 101, 1–82. [Google Scholar] [PubMed]
- Antonenko, Y.N.; Rokitskaya, T.I.; Huczynski, A. Electrogenic and nonelectrogenic ion fluxes across lipid and mitochondrial membranes mediated by monensin and monensin ethyl ester. Biochim. Biophys. Acta 2015, 1848, 995–1004. [Google Scholar] [CrossRef] [PubMed]
- Huczynski, A. Polyether ionophores-promising bioactive molecules for cancer therapy. Bioorg. Med. Chem. Lett. 2012, 22, 7002–7010. [Google Scholar] [CrossRef] [PubMed]
- Versini, A.; Saier, L.; Sindikubwabo, F.; Müller, S.; Cañeque, T.; Rodriguez, R. Chemical biology of salinomycin. Rodriguez 2018, 74, 5585–5614. [Google Scholar] [CrossRef]
- Rutkowski, J.; Brzezinski, B. Structures and properties of naturally occurring polyether antibiotics. Biomed. Res. Int. 2013, 2013, 162513. [Google Scholar] [CrossRef] [PubMed]
- An, H.; Kim, J.Y.; Lee, N.; Cho, Y.; Oh, E.; Seo, J.H. Salinomycin possesses anti-tumor activity and inhibits breast cancer stem-like cells via an apoptosis-independent pathway. Biochem. Biophys. Res. Commun. 2015, 466, 696–703. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Yi, L.; Li, F.; Hu, R.; Hu, S.; Yin, Y.; Lan, C.; Li, Z.; Fu, C.; Cao, L.; et al. Salinomycin inhibits the tumor growth of glioma stem cells by selectively suppressing glioma-initiating cells. Mol. Med. Rep. 2015, 11, 2407–2412. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.; Kim, Y.H.; Kwon, M.; Shin, S.J.; Kwon, S.H.; Cha, S.D.; Cho, C.H. The effect of salinomycin on ovarian cancer stem-like cells. Obstet. Gynecol. Sci. 2016, 59, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.G.; Shin, S.J.; Chung, H.W.; Kwon, S.H.; Cha, S.D.; Lee, J.E.; Cho, C.H. Salinomycin reduces stemness and induces apoptosis on human ovarian cancer stem cell. J. Gynecol. Oncol. 2017, 28, e14. [Google Scholar] [CrossRef] [PubMed]
- Mao, J.; Fan, S.; Ma, W.; Fan, P.; Wang, B.; Zhang, J.; Wang, H.; Tang, B.; Zhang, Q.; Yu, X.; et al. Roles of Wnt/beta-catenin signaling in the gastric cancer stem cells proliferation and salinomycin treatment. Cell Death Dis. 2014, 5, e1039. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.L.; Zhao, Z.Q.; Li, J.C.; Liang, Y.; Yin, J.Q.; Zou, C.Y.; Xie, X.B.; Zeng, Y.X.; Shen, J.N.; Kang, T.; et al. Salinomycin inhibits osteosarcoma by targeting its tumor stem cells. Cancer Lett. 2011, 311, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Tian, Y.; Song, F.; Fu, C.; Han, B.; Wang, Y. Salinomycin inhibits the growth of colorectal carcinoma by targeting tumor stem cells. Oncol. Rep. 2015, 34, 2469–2476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antoszczak, M.; Huczynski, A. Anticancer Activity of Polyether Ionophore-Salinomycin. Anticancer Agents Med. Chem. 2015, 15, 575–591. [Google Scholar] [CrossRef] [PubMed]
- Dewangan, J.; Srivastava, S.; Rath, S.K. Salinomycin: A new paradigm in cancer therapy. Tumour Biol. 2017, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaplan, F.; Teksen, F. Apoptotic effects of salinomycin on human ovarian cancer cell line (OVCAR-3). Tumour Biol. 2016, 37, 3897–3903. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.Y.; Yu, S.N.; Lee, S.Y.; Chun, S.S.; Choi, Y.L.; Park, Y.M.; Song, C.S.; Chatterjee, B.; Ahn, S.C. Salinomycin-induced apoptosis of human prostate cancer cells due to accumulated reactive oxygen species and mitochondrial membrane depolarization. Biochem. Biophys. Res. Commun. 2011, 413, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.G.; Lee, J.M.; Shin, S.J.; Kwon, S.H.; Lee, G.S.; Song, C.H.; Choi, E.S.; Cha, S.D.; Cho, C.H. Salinomycin inhibited cell proliferation and induced apoptosis in human uterine leiomyoma cells. Obstet. Gynecol. Sci. 2014, 57, 501–506. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.; Choi, M.Y.; Yu, J.; Castro, J.E.; Kipps, T.J.; Carson, D.A. Salinomycin inhibits Wnt signaling and selectively induces apoptosis in chronic lymphocytic leukemia cells. Proc. Natl. Acad. Sci. USA 2011, 108, 13253–13257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, F.; He, L.; Dai, W.Q.; Xu, Y.P.; Wu, D.; Lin, C.L.; Wu, S.M.; Cheng, P.; Zhang, Y.; Shen, M.; et al. Salinomycin inhibits proliferation and induces apoptosis of human hepatocellular carcinoma cells in vitro and in vivo. PLoS ONE 2012, 7, e50638. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Zhang, Y.; Huang, J.; Fan, Z.; Shi, F.; Wang, S. Salinomycin inhibits proliferation and induces apoptosis of human nasopharyngeal carcinoma cell in vitro and suppresses tumor growth in vivo. Biochem. Biophys. Res. Commun. 2014, 443, 712–717. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Su, L.; Zhong, N.; Hao, X.; Zhong, D.; Singhal, S.; Liu, X. Salinomycin induces cell death with autophagy through activation of endoplasmic reticulum stress in human cancer cells. Autophagy 2013, 9, 1057–1068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verdoodt, B.; Vogt, M.; Schmitz, I.; Liffers, S.T.; Tannapfel, A.; Mirmohammadsadegh, A. Salinomycin induces autophagy in colon and breast cancer cells with concomitant generation of reactive oxygen species. PLoS ONE 2012, 7, e44132. [Google Scholar] [CrossRef] [PubMed]
- Endo, S.; Nakata, K.; Sagara, A.; Koikawa, K.; Ando, Y.; Kibe, S.; Takesue, S.; Nakayama, H.; Abe, T.; Okumura, T.; et al. Autophagy inhibition enhances antiproliferative effect of salinomycin in pancreatic cancer cells. Pancreatology 2017, 17, 990–996. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.Y.; Park, K.I.; Kim, S.H.; Yu, S.N.; Park, S.G.; Kim, Y.W.; Seo, Y.K.; Ma, J.Y.; Ahn, S.C. Inhibition of Autophagy Promotes Salinomycin-Induced Apoptosis via Reactive Oxygen Species-Mediated PI3K/AKT/mTOR and ERK/p38 MAPK-Dependent Signaling in Human Prostate Cancer Cells. Int. J. Mol. Sci. 2017, 18, 1088. [Google Scholar] [CrossRef] [PubMed]
- Klose, J.; Stankov, M.V.; Kleine, M.; Ramackers, W.; Panayotova-Dimitrova, D.; Jager, M.D.; Klempnauer, J.; Winkler, M.; Bektas, H.; Behrens, G.M.; et al. Inhibition of autophagic flux by salinomycin results in anti-cancer effect in hepatocellular carcinoma cells. PLoS ONE 2014, 9, e95970. [Google Scholar] [CrossRef] [PubMed]
- Xipell, E.; Gonzalez-Huarriz, M.; Martinez de Irujo, J.J.; Garcia-Garzon, A.; Lang, F.F.; Jiang, H.; Fueyo, J.; Gomez-Manzano, C.; Alonso, M.M. Salinomycin induced ROS results in abortive autophagy and leads to regulated necrosis in glioblastoma. Oncotarget 2016, 7, 30626–30641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yue, W.; Hamai, A.; Tonelli, G.; Bauvy, C.; Nicolas, V.; Tharinger, H.; Codogno, P.; Mehrpour, M. Inhibition of the autophagic flux by salinomycin in breast cancer stem-like/progenitor cells interferes with their maintenance. Autophagy 2013, 9, 714–729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.Y.; Park, K.I.; Kim, S.H.; Yu, S.N.; Lee, D.; Kim, Y.W.; Noh, K.T.; Ma, J.Y.; Seo, Y.K.; Ahn, S.C. Salinomycin Induces Reactive Oxygen Species and Apoptosis in Aggressive Breast Cancer Cells as Mediated with Regulation of Autophagy. Anticancer Res. 2017, 37, 1747–1758. [Google Scholar] [PubMed]
- Kim, S.H.; Choi, Y.J.; Kim, K.Y.; Yu, S.N.; Seo, Y.K.; Chun, S.S.; Noh, K.T.; Suh, J.T.; Ahn, S.C. Salinomycin simultaneously induces apoptosis and autophagy through generation of reactive oxygen species in osteosarcoma U2OS cells. Biochem. Biophys. Res. Commun. 2016, 473, 607–613. [Google Scholar] [CrossRef] [PubMed]
- Qin, L.S.; Jia, P.F.; Zhang, Z.Q.; Zhang, S.M. ROS-p53-cyclophilin-D signaling mediates salinomycin-induced glioma cell necrosis. J. Exp. Clin. Cancer Res. 2015, 34, 57. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Wang, F.; Dai, W.Q.; Wu, D.; Lin, C.L.; Wu, S.M.; Cheng, P.; Zhang, Y.; Shen, M.; Wang, C.F.; et al. Mechanism of action of salinomycin on growth and migration in pancreatic cancer cell lines. Pancreatology 2013, 13, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Klose, J.; Eissele, J.; Volz, C.; Schmitt, S.; Ritter, A.; Ying, S.; Schmidt, T.; Heger, U.; Schneider, M.; Ulrich, A. Salinomycin inhibits metastatic colorectal cancer growth and interferes with Wnt/beta-catenin signaling in CD133+ human colorectal cancer cells. BMC Cancer 2016, 16, 896. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Dong, T.; Hu, C.; Lu, J.; Dai, J.; Liu, P. Salinomycin repressed the epithelial-mesenchymal transition of epithelial ovarian cancer cells via downregulating Wnt/beta-catenin pathway. Onco Targets Ther. 2017, 10, 1317–1325. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.; Li, Y. Salinomycin suppresses LRP6 expression and inhibits both Wnt/beta-catenin and mTORC1 signaling in breast and prostate cancer cells. J. Cell. Biochem. 2014, 115, 1799–1807. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.Z.; Yan, Y.Y.; He, M.; Xiao, Q.H.; Yao, W.F.; Zhao, L.; Wu, H.Z.; Yu, Z.J.; Zhou, M.Y.; Lv, M.T.; et al. Salinomycin induces selective cytotoxicity to MCF-7 mammosphere cells through targeting the Hedgehog signaling pathway. Oncol. Rep. 2016, 35, 912–922. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Zhang, C.; Li, Q.; Mao, J.; Ma, W.; Yu, X.; Hou, Z.; Li, L. Inhibitory effect of salinomycin on human breast cancer cells MDA-MB-231 proliferation through Hedgehog signaling pathway. Zhonghua Bing Li Xue Za Zhi 2015, 44, 395–398. [Google Scholar] [PubMed]
- Kim, K.Y.; Kim, S.H.; Yu, S.N.; Park, S.K.; Choi, H.D.; Yu, H.S.; Ji, J.H.; Seo, Y.K.; Ahn, S.C. Salinomycin enhances doxorubicin-induced cytotoxicity in multidrug resistant MCF-7/MDR human breast cancer cells via decreased efflux of doxorubicin. Mol. Med. Rep. 2015, 12, 1898–1904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oak, P.S.; Kopp, F.; Thakur, C.; Ellwart, J.W.; Rapp, U.R.; Ullrich, A.; Wagner, E.; Knyazev, P.; Roidl, A. Combinatorial treatment of mammospheres with trastuzumab and salinomycin efficiently targets HER2-positive cancer cells and cancer stem cells. Int. J. Cancer 2012, 131, 2808–2819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sommer, A.K.; Hermawan, A.; Mickler, F.M.; Ljepoja, B.; Knyazev, P.; Brauchle, C.; Ullrich, A.; Wagner, E.; Roidl, A. Salinomycin co-treatment enhances tamoxifen cytotoxicity in luminal A breast tumor cells by facilitating lysosomal degradation of receptor tyrosine kinases. Oncotarget 2016, 7, 50461–50476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, G.N.; Liang, Y.; Zhou, L.J.; Chen, S.P.; Chen, G.; Zhang, T.P.; Kang, T.; Zhao, Y.P. Combination of salinomycin and gemcitabine eliminates pancreatic cancer cells. Cancer Lett. 2011, 313, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Venkatadri, R.; Iyer, A.K.V.; Kaushik, V.; Azad, N. A novel resveratrol-salinomycin combination sensitizes ER-positive breast cancer cells to apoptosis. Pharmacol. Rep. 2017, 69, 788–797. [Google Scholar] [CrossRef] [PubMed]
- Manmuan, S.; Sakunrangsit, N.; Ketchart, W. Salinomycin overcomes acquired tamoxifen resistance through AIB1 and inhibits cancer cell invasion in endocrine resistant breast cancer. Clin. Exp. Pharmacol. Physiol. 2017, 44, 1042–1052. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Li, P.; Xue, X.; He, S.; Kuang, Y.; Zhao, H.; Chen, S.; Zhi, Q.; Guo, X. Salinomycin induces apoptosis in cisplatin-resistant colorectal cancer cells by accumulation of reactive oxygen species. Toxicol. Lett. 2013, 222, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Liang, C.; Xue, F.; Chen, W.; Zhi, X.; Feng, X.; Bai, X.; Liang, T. Salinomycin decreases doxorubicin resistance in hepatocellular carcinoma cells by inhibiting the beta-catenin/TCF complex association via FOXO3a activation. Oncotarget 2015, 6, 10350–10365. [Google Scholar] [PubMed]
- Hermawan, A.; Wagner, E.; Roidl, A. Consecutive salinomycin treatment reduces doxorubicin resistance of breast tumor cells by diminishing drug efflux pump expression and activity. Oncol. Rep. 2016, 35, 1732–1740. [Google Scholar] [CrossRef] [PubMed]
- Riccioni, R.; Dupuis, M.L.; Bernabei, M.; Petrucci, E.; Pasquini, L.; Mariani, G.; Cianfriglia, M.; Testa, U. The cancer stem cell selective inhibitor salinomycin is a p-glycoprotein inhibitor. Blood Cells Mol. Dis. 2010, 45, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Chen, H.; Yu, C.; Zhang, Y.; Chen, M.; Tian, S.; Sun, C. The promotion of salinomycin delivery to hepatocellular carcinoma cells through EGFR and CD133 aptamers conjugation by PLGA nanoparticles. Nanomedicine (Lond.) 2015, 10, 1863–1879. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Liu, Y.; Li, S.; Rohrs, J.; Zhang, R.; Zhang, X.; Wang, P. Co-Eradication of Breast Cancer Cells and Cancer Stem Cells by Cross-Linked Multilamellar Liposomes Enhances Tumor Treatment. Mol. Pharm. 2015, 12, 2811–2822. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.; Liu, J.; Gong, Z.; Zhang, H.; Lu, Y.; Zou, H.; Yu, Y.; Chen, Y.; Sun, Z.; Li, W.; et al. iRGD-conjugated DSPE-PEG2000 nanomicelles for targeted delivery of salinomycin for treatment of both liver cancer cells and cancer stem cells. Nanomedicine (Lond.) 2015, 10, 2677–2695. [Google Scholar] [CrossRef] [PubMed]
- Ni, M.; Xiong, M.; Zhang, X.; Cai, G.; Chen, H.; Zeng, Q.; Yu, Z. Poly (lactic-co-glycolic acid) nanoparticles conjugated with CD133 aptamers for targeted salinomycin delivery to CD133+ osteosarcoma cancer stem cells. Int. J. Nanomed. 2015, 10, 2537–2554. [Google Scholar]
- Yao, H.J.; Zhang, Y.G.; Sun, L.; Liu, Y. The effect of hyaluronic acid functionalized carbon nanotubes loaded with salinomycin on gastric cancer stem cells. Biomaterials 2014, 35, 9208–9223. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Deng, Z.; Tian, J.; Liu, T. Synthesis and biological evaluation of salinomycin triazole analogues as anticancer agents. Eur. J. Med. Chem. 2017, 127, 900–908. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Borgstrom, B.; Mansson, L.; Persson, L.; Oredsson, S.; Hegardt, C.; Strand, D. Semisynthesis of SY-1 for investigation of breast cancer stem cell selectivity of C-ring-modified salinomycin analogues. ACS Chem. Biol. 2014, 9, 1587–1594. [Google Scholar] [CrossRef] [PubMed]
- Benedict, R.G. Antibiotics produced by actinomycetes. Bot. Rev. 1953, 19. [Google Scholar] [CrossRef]
- Harned, R.L.; Hidy, P.H.; Corum, C.J.; Jones, K.L. Nigericin a new crystalline antibiotic from an unidentified Streptomyces. Antibiot. Chemother. (Northfield) 1951, 1, 594–596. [Google Scholar] [PubMed]
- Pouyssegur, J.; Franchi, A.; L’Allemain, G.; Paris, S. Cytoplasmic pH, a key determinant of growth factor-induced DNA synthesis in quiescent fibroblasts. FEBS Lett. 1985, 190, 115–119. [Google Scholar] [CrossRef] [Green Version]
- Moolenaar, W.H. Effects of growth factors on intracellular pH regulation. Annu. Rev. Physiol. 1986, 48, 363–376. [Google Scholar] [CrossRef] [PubMed]
- Damaghi, M.; Wojtkowiak, J.W.; Gillies, R.J. pH sensing and regulation in cancer. Front. Physiol. 2013, 4, 370. [Google Scholar] [CrossRef] [PubMed]
- Margolis, L.B.; Rozovskaja, I.A.; Skulachev, V.P. Acidification of the interior of Ehrlich ascites tumor cells by nigericin inhibits DNA synthesis. FEBS Lett. 1987, 220, 288–290. [Google Scholar] [CrossRef] [Green Version]
- Hegazy, A.M.; Yamada, D.; Kobayashi, M.; Kohno, S.; Ueno, M.; Ali, M.A.; Ohta, K.; Tadokoro, Y.; Ino, Y.; Todo, T.; et al. Therapeutic Strategy for Targeting Aggressive Malignant Gliomas by Disrupting Their Energy Balance. J. Biol. Chem. 2016, 291, 21496–21509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varnes, M.E.; Clay, M.E.; Freeman, K.; Antunez, A.R.; Oleinick, N.L. Enhancement of photodynamic cell killing (with chloroaluminum phthalocyanine) by treatment of V79 cells with the ionophore nigericin. Cancer Res. 1990, 50, 1620–1625. [Google Scholar] [PubMed]
- Xue, L.Y.; Agarwal, M.L.; Varnes, M.E. Elevation of GRP-78 and loss of HSP-70 following photodynamic treatment of V79 cells: Sensitization by nigericin. Photochem. Photobiol. 1995, 62, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Varnes, M.E.; Bayne, M.T.; Menegay, H.J.; Tuttle, S.W. Effect of the K+/H+ ionophore nigericin on response of A549 cells to photodynamic therapy and tert-butylhydroperoxide. Free Radic. Biol. Med. 1993, 15, 395–405. [Google Scholar] [CrossRef]
- Yang, L.; Mei, Y.; Xie, Q.; Han, X.; Zhang, F.; Gu, L.; Zhang, Y.; Chen, Y.; Li, G.; Gao, Z. Acidification induces Bax translocation to the mitochondria and promotes ultraviolet light-induced apoptosis. Cell. Mol. Biol. Lett. 2008, 13, 119–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varnes, M.E.; Menegay, H.J.; McKenna, D.S. Inhibition of recovery from potentially lethal radiation damage in A549 cells by the K+/H+ ionophore nigericin. Int. J. Radiat. Oncol. Biol. Phys. 1991, 20, 281–285. [Google Scholar] [CrossRef]
- Wood, P.J.; Sansom, J.M.; Newell, K.; Tannock, I.F.; Stratford, I.J. Reduction of tumour intracellular pH and enhancement of melphalan cytotoxicity by the ionophore Nigericin. Int. J. Cancer 1995, 60, 264–268. [Google Scholar] [CrossRef] [PubMed]
- Jahde, E.; Glusenkamp, K.H.; Rajewsky, M.F. Nigericin enhances mafosfamide cytotoxicity at low extracellular pH. Cancer Chemother. Pharmacol. 1991, 27, 440–444. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhao, Y.; Yao, S.; Cui, X.; Pan, W.; Huang, W.; Gao, J.; Dong, T.; Zhang, S. Nigericin Inhibits Epithelial Ovarian Cancer Metastasis by Suppressing the Cell Cycle and Epithelial-Mesenchymal Transition. Biochemistry 2017, 82, 933–941. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Li, W.; Hua, S.; Han, Y.; Xu, Z.; Wan, D.; Wang, Y.; Chen, W.; Kuang, Y.; Shi, J.; et al. Nigericin Exerts Anticancer Effects on Human Colorectal Cancer Cells by Inhibiting Wnt/β-catenin Signaling Pathway. Mol. Cancer Ther. 2018, 17, 952–965. [Google Scholar] [CrossRef] [PubMed]
- Vu, H.T.; Kobayashi, M.; Hegazy, A.M.; Tadokoro, Y.; Ueno, M.; Kasahara, A.; Takase, Y.; Nomura, N.; Peng, H.; Ito, C.; et al. Autophagy inhibition synergizes with calcium mobilization to achieve efficient therapy of malignant gliomas. Cancer Sci. 2018, 109, 2497–2508. [Google Scholar] [CrossRef] [PubMed]
- Diaz de Grenu, B.; Iglesias Hernandez, P.; Espona, M.; Quinonero, D.; Light, M.E.; Torroba, T.; Perez-Tomas, R.; Quesada, R. Synthetic prodiginine obatoclax (GX15-070) and related analogues: Anion binding, transmembrane transport, and cytotoxicity properties. Chemistry 2011, 17, 14074–14083. [Google Scholar] [CrossRef] [PubMed]
- Cruickshanks, N.; Tang, Y.; Booth, L.; Hamed, H.; Grant, S.; Dent, P. Lapatinib and obatoclax kill breast cancer cells through reactive oxygen species-dependent endoplasmic reticulum stress. Mol. Pharmacol. 2012, 82, 1217–1229. [Google Scholar] [CrossRef] [PubMed]
- Martin, A.P.; Mitchell, C.; Rahmani, M.; Nephew, K.P.; Grant, S.; Dent, P. Inhibition of MCL-1 enhances lapatinib toxicity and overcomes lapatinib resistance via BAK-dependent autophagy. Cancer Biol. Ther. 2009, 8, 2084–2096. [Google Scholar] [CrossRef] [PubMed]
- Brem, E.A.; Thudium, K.; Khubchandani, S.; Tsai, P.C.; Olejniczak, S.H.; Bhat, S.; Riaz, W.; Gu, J.; Iqbal, A.; Campagna, R.; et al. Hernandez-Ilizaliturri, F.J. Distinct cellular and therapeutic effects of obatoclax in rituximab-sensitive and -resistant lymphomas. Br. J. Haematol. 2011, 153, 599–611. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.Z.; Ma, B.; Liang, Y.J.; Liu, H.C.; Zhang, T.H.; Zheng, G.S.; Su, Y.X.; Liao, G.Q. Obatoclax induces Beclin 1- and ATG5-dependent apoptosis and autophagy in adenoid cystic carcinoma cells. Oral Dis. 2015, 21, 470–477. [Google Scholar] [CrossRef] [PubMed]
- McCoy, F.; Hurwitz, J.; McTavish, N.; Paul, I.; Barnes, C.; O’Hagan, B.; Odrzywol, K.; Murray, J.; Longley, D.; McKerr, G.; et al. Obatoclax induces Atg7-dependent autophagy independent of beclin-1 and BAX/BAK. Cell. Death Dis. 2010, 1, e108. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Hamed, H.A.; Cruickshanks, N.; Fisher, P.B.; Grant, S.; Dent, P. Obatoclax and lapatinib interact to induce toxic autophagy through NOXA. Mol. Pharmacol. 2012, 81, 527–540. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Kadia, T.; Tong, W.; Zhang, M.; Jia, Y.; Yang, H.; Hu, Y.; Viallet, J.; O’Brien, S.; Garcia-Manero, G. The combination of a histone deacetylase inhibitor with the BH3-mimetic GX15-070 has synergistic antileukemia activity by activating both apoptosis and autophagy. Autophagy 2010, 6, 976–978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamelos, V.A.; Fisher, N.; Bamrah, H.; Voisey, C.; Price, J.C.; Farrell, W.E.; Redman, C.W.; Richardson, A. The BH3 Mimetic Obatoclax Accumulates in Lysosomes and Causes Their Alkalinization. PLoS ONE 2016, 11, e0150696. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Wu, W.K.; Gu, C.; Zhong, D.; Zhao, X.; Kong, Y.; Lin, Q.; Chan, M.T.; Zhou, Z.; Liu, S. Obatoclax impairs lysosomal function to block autophagy in cisplatin-sensitive and -resistant esophageal cancer cells. Oncotarget 2016, 7, 14693–14707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basit, F.; Cristofanon, S.; Fulda, S. Obatoclax (GX15-070) triggers necroptosis by promoting the assembly of the necrosome on autophagosomal membranes. Cell Death Differ. 2013, 20, 1161–1173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Wang, G.; Niu, X.; Zhao, J.; Tan, W.; Wang, H.; Zhao, L.; Ge, Y. Combination of AZD2281 (Olaparib) and GX15-070 (Obatoclax) results in synergistic antitumor activities in preclinical models of pancreatic cancer. Cancer Lett. 2014, 348, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.Y.; Lin, Q.H.; Que, F.C.; Gu, C.P.; Yu, L.; Liu, S.W. Synergistic anti-tumor effect of obatoclax and MG-132 in esophageal cancer cell line CaES-17. Nan Fang Yi Ke Da Xue Xue Bao 2016, 36, 506–513. [Google Scholar] [PubMed]
- Li, J.; Viallet, J.; Haura, E.B. A small molecule pan-Bcl-2 family inhibitor, GX15-070, induces apoptosis and enhances cisplatin-induced apoptosis in non-small cell lung cancer cells. Cancer Chemother. Pharmacol. 2008, 61, 525–534. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Zhao, Z.; Wu, K.; Xu, Z.; Liu, K. MCL-1 is the key target of adjuvant chemotherapy to reverse the cisplatin-resistance in NSCLC. Gene 2016, 587, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.J.; Sun, Z.K.; Shen, C.T.; Song, H.J.; Zhang, X.Y.; Qiu, Z.L.; Luo, Q.Y. Obatoclax and LY3009120 Efficiently Overcome Vemurafenib Resistance in Differentiated Thyroid Cancer. Theranostics 2017, 7, 987–1001. [Google Scholar] [CrossRef] [PubMed]
- Heidari, N.; Hicks, M.A.; Harada, H. GX15-070 (obatoclax) overcomes glucocorticoid resistance in acute lymphoblastic leukemia through induction of apoptosis and autophagy. Cell Death Dis. 2010, 1, e76. [Google Scholar] [CrossRef] [PubMed]
- Bonapace, L.; Bornhauser, B.C.; Schmitz, M.; Cario, G.; Ziegler, U.; Niggli, F.K.; Schafer, B.W.; Schrappe, M.; Stanulla, M.; Bourquin, J.P. Induction of autophagy-dependent necroptosis is required for childhood acute lymphoblastic leukemia cells to overcome glucocorticoid resistance. J. Clin. Investig. 2010, 120, 1310–1323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Brien, S.M.; Claxton, D.F.; Crump, M.; Faderl, S.; Kipps, T.; Keating, M.J.; Viallet, J.; Cheson, B.D. Phase I study of obatoclax mesylate (GX15-070), a small molecule pan-Bcl-2 family antagonist, in patients with advanced chronic lymphocytic leukemia. Blood 2009, 113, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Hrgovic, I.; Glavic, Z.; Kovacic, Z.; Mulic, S.; Zunic, L.; Hrgovic, Z. Repeated administration of inhibitors for ion pumps reduce markedly tumor growth in vivo. Med. Arch. 2014, 68, 76–78. [Google Scholar] [CrossRef] [PubMed]
- Naujokat, C.; Fuchs, D.; Opelz, G. Salinomycin in cancer: A new mission for an old agent. Mol. Med. Rep. 2010, 3, 555–559. [Google Scholar] [CrossRef] [PubMed]
- Schimmer, A.D.; Raza, A.; Carter, T.H.; Claxton, D.; Erba, H.; DeAngelo, D.J.; Tallman, M.S.; Goard, C.; Borthakur, G. A multicenter phase I/II study of obatoclax mesylate administered as a 3- or 24-hour infusion in older patients with previously untreated acute myeloid leukemia. PLoS ONE 2014, 9, e108694. [Google Scholar] [CrossRef] [PubMed]
- Oki, Y.; Copeland, A.; Hagemeister, F.; Fayad, L.E.; Fanale, M.; Romaguera, J.; Younes, A. Experience with obatoclax mesylate (GX15-070), a small molecule pan-Bcl-2 family antagonist in patients with relapsed or refractory classical Hodgkin lymphoma. Blood 2012, 119, 2171–2172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arellano, M.L.; Borthakur, G.; Berger, M.; Luer, J.; Raza, A. A phase II, multicenter, open-label study of obatoclax mesylate in patients with previously untreated myelodysplastic syndromes with anemia or thrombocytopenia. Clin. Lymphoma Myeloma Leuk 2014, 14, 534–539. [Google Scholar] [CrossRef] [PubMed]
- Langer, C.J.; Albert, I.; Ross, H.J.; Kovacs, P.; Blakely, L.J.; Pajkos, G.; Somfay, A.; Zatloukal, P.; Kazarnowicz, A.; Moezi, M.M.; et al. Randomized phase II study of carboplatin and etoposide with or without obatoclax mesylate in extensive-stage small cell lung cancer. Lung Cancer 2014, 85, 420–428. [Google Scholar] [CrossRef] [PubMed]
- Paik, P.K.; Rudin, C.M.; Pietanza, M.C.; Brown, A.; Rizvi, N.A.; Takebe, N.; Travis, W.; James, L.; Ginsberg, M.S.; Juergens, R.; et al. A phase II study of obatoclax mesylate, a Bcl-2 antagonist, plus topotecan in relapsed small cell lung cancer. Lung Cancer 2011, 74, 481–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urtishak, K.A.; Edwards, A.Y.; Wang, L.S.; Hudome, A.; Robinson, B.W.; Barrett, J.S.; Cao, K.; Cory, L.; Moore, J.S.; Bantly, A.D.; et al. Potent obatoclax cytotoxicity and activation of triple death mode killing across infant acute lymphoblastic leukemia. Blood 2013, 121, 2689–2703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boehmerle, W.; Endres, M. Salinomycin induces calpain and cytochrome c-mediated neuronal cell death. Cell Death Dis. 2011, 2, e168. [Google Scholar] [CrossRef] [PubMed]
- Delwar, Z.M.; Avramidis, D.; Siden, A.; Cruz, M.; Yakisich, J.S. Depletion of drug-surviving glioma cells by a second phase treatment with low concentration of salinomycin. Drugs Ther. Stud. 2011, 1, 21–25. [Google Scholar] [CrossRef]
- Huang, X.; Borgstrom, B.; Stegmayr, J.; Abassi, Y.; Kruszyk, M.; Leffler, H.; Persson, L.; Albinsson, S.; Massoumi, R.; Scheblykin, I.G.; et al. The Molecular Basis for Inhibition of Stemlike Cancer Cells by Salinomycin. ACS Cent. Sci. 2018, 4, 760–767. [Google Scholar] [CrossRef] [PubMed]
- Najumudeen, A.K.; Jaiswal, A.; Lectez, B.; Oetken-Lindholm, C.; Guzman, C.; Siljamaki, E.; Posada, I.M.; Lacey, E.; Aittokallio, T.; Abankwa, D. Cancer stem cell drugs target K-ras signaling in a stemness context. Oncogene 2016, 35, 5248–5262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.; Ma, W.; Mao, J.; Yu, X.; Hou, Z.; Fan, S.; Song, B.; Wang, H.; Li, J.; Kang, L.; et al. Salinomycin exerts anticancer effects on human breast carcinoma MCF-7 cancer stem cells via modulation of Hedgehog signaling. Chem. Biol. Interact. 2015, 228, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Jing, H.; Qu, X.; Liu, L.; Xia, H. A Novel Long Noncoding RNA (lncRNA), LL22NC03-N64E9.1, Promotes the Proliferation of Lung Cancer Cells and is a Potential Prognostic Molecular Biomarker for Lung Cancer. Med. Sci. Monit. 2018, 24, 4317–4323. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.F.; Su, J.C.; Liu, C.Y.; Huang, J.W.; Chen, K.C.; Chen, W.L.; Tai, W.T.; Shiau, C.W. A novel obatoclax derivative, SC-2001, induces apoptosis in hepatocellular carcinoma cells through SHP-1-dependent STAT3 inactivation. Cancer Lett. 2012, 321, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Su, J.C.; Chen, K.F.; Chen, W.L.; Liu, C.Y.; Huang, J.W.; Tai, W.T.; Chen, P.J.; Kim, I.; Shiau, C.W. Synthesis and biological activity of obatoclax derivatives as novel and potent SHP-1 agonists. Eur. J. Med. Chem. 2012, 56, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Antoszczak, M.; Urbaniak, A.; Delgado, M.; Maj, E.; Borgstrom, B.; Wietrzyk, J.; Huczynski, A.; Yuan, Y.; Chambers, T.C.; Strand, D. Biological activity of doubly modified salinomycin analogs—Evaluation in vitro and ex vivo. Eur. J. Med. Chem. 2018, 156, 510–523. [Google Scholar] [CrossRef] [PubMed]
- Borgstrom, B.; Huang, X.; Chygorin, E.; Oredsson, S.; Strand, D. Salinomycin Hydroxamic Acids: Synthesis, Structure, and Biological Activity of Polyether Ionophore Hybrids. ACS Med. Chem. Lett. 2016, 7, 635–640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huczynski, A.; Janczak, J.; Antoszczak, M.; Wietrzyk, J.; Maj, E.; Brzezinski, B. Antiproliferative activity of salinomycin and its derivatives. Bioorg. Med. Chem. Lett. 2012, 22, 7146–7150. [Google Scholar] [CrossRef] [PubMed]
- Urbaniak, A.; Delgado, M.; Antoszczak, M.; Huczynski, A.; Chambers, T.C. Salinomycin derivatives exhibit activity against primary acute lymphoblastic leukemia (ALL) cells in vitro. Biomed. Pharmacother. 2018, 99, 384–390. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Wu, J.; Li, B.; Lian, X.; Xia, J.; Zhou, Q.; Wu, S. Design and synthesis of conformationally constrained salinomycin derivatives. Eur. J. Med. Chem. 2017, 138, 353–356. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Wu, J.; Li, B.; Wu, H.; Wang, L.; Hao, J.; Wu, S.; Zhou, Q. Structure-activity & structure-toxicity relationship study of salinomycin diastereoisomers and their benzoylated derivatives. Org. Biomol. Chem. 2016, 14, 2840–2845. [Google Scholar] [PubMed]
- Zhou, J.; Sun, J.; Chen, H.; Peng, Q. Promoted delivery of salinomycin sodium to lung cancer cells by dual targeting PLGA hybrid nanoparticles. Int. J. Oncol. 2018, 53, 1289–1300. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Zeng, Y.; Qi, X.; Chen, Y.; Ge, Z.; Jiang, Z.; Zhang, X.; Dong, Y.; Chen, H.; Yu, Z. Targeted salinomycin delivery with EGFR and CD133 aptamers based dual-ligand lipid-polymer nanoparticles to both osteosarcoma cells and cancer stem cells. Nanomedicine 2018, 14, 2115–2127. [Google Scholar] [CrossRef] [PubMed]
- Mi, Y.; Huang, Y.; Deng, J. The enhanced delivery of salinomycin to CD133(+) ovarian cancer stem cells through CD133 antibody conjugation with poly(lactic-co-glycolic acid)-poly(ethylene glycol) nanoparticles. Oncol. Lett. 2018, 15, 6611–6621. [Google Scholar] [CrossRef] [PubMed]
- Daman, Z.; Faghihi, H.; Montazeri, H. Salinomycin nanoparticles interfere with tumor cell growth and the tumor microenvironment in an orthotopic model of pancreatic cancer. Drug Dev. Ind. Pharm. 2018, 44, 1434–1442. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.J.; Gurunathan, S.; Kim, J.H. Graphene Oxide-Silver Nanocomposite Enhances Cytotoxic and Apoptotic Potential of Salinomycin in Human Ovarian Cancer Stem Cells (OvCSCs): A Novel Approach for Cancer Therapy. Int. J. Mol. Sci. 2018, 19, 710. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Chen, F.; Qi, X.; Dong, Y.; Zhang, Y.; Ge, Z.; Cai, G.; Zhang, X. Epidermal growth factor receptor aptamer-conjugated polymer-lipid hybrid nanoparticles enhance salinomycin delivery to osteosarcoma and cancer stem cells. Exp. Ther. Med. 2018, 15, 1247–1256. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.B.; Yu, Z.C.; He, Y.N.; Zhang, T.; Du, L.B.; Dong, Y.M.; Chen, H.W.; Zhang, Y.Y.; Wang, W.Q. Salinomycin-loaded lipid-polymer nanoparticles with anti-CD20 aptamers selectively suppress human CD20+ melanoma stem cells. Acta Pharmacol. Sin. 2018, 39, 261–274. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, Q.; Sun, J.; Liu, H.; Li, Q. The combination therapy of salinomycin and gefitinib using poly(d,l-lactic-co-glycolic acid)-poly(ethylene glycol) nanoparticles for targeting both lung cancer stem cells and cancer cells. Onco Targets Ther. 2017, 10, 5653–5666. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Xu, W.; Yuan, X.; Chen, H.; Song, H.; Wang, B.; Han, J. Polymer-lipid hybrid anti-HER2 nanoparticles for targeted salinomycin delivery to HER2-positive breast cancer stem cells and cancer cells. Int. J. Nanomed. 2017, 12, 6909–6921. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Cui, D.; Ye, L.; Li, Y.; Zhu, L.; Yang, L.; Bai, B.; Nie, Z.; Gao, J.; Cao, Y. Codelivery of salinomycin and docetaxel using poly(d,l-lactic-co-glycolic acid)-poly(ethylene glycol) nanoparticles to target both gastric cancer cells and cancer stem cells. Anticancer Drugs 2017, 28, 989–1001. [Google Scholar] [CrossRef] [PubMed]
- Gong, Z.; Chen, D.; Xie, F.; Liu, J.; Zhang, H.; Zou, H.; Yu, Y.; Chen, Y.; Sun, Z.; Wang, X.; et al. Codelivery of salinomycin and doxorubicin using nanoliposomes for targeting both liver cancer cells and cancer stem cells. Nanomedicine (Lond.) 2016, 11, 2565–2579. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Xia, G.; Dong, S.; Jiang, Z.X.; Chen, M. An iTEP-salinomycin nanoparticle that specifically and effectively inhibits metastases of 4T1 orthotopic breast tumors. Biomaterials 2016, 93, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muntimadugu, E.; Kumar, R.; Saladi, S.; Rafeeqi, T.A.; Khan, W. CD44 targeted chemotherapy for co-eradication of breast cancer stem cells and cancer cells using polymeric nanoparticles of salinomycin and paclitaxel. Colloids Surf. B Biointerfaces 2016, 143, 532–546. [Google Scholar] [CrossRef] [PubMed]
- Tigli Aydin, R.S.; Kaynak, G.; Gumusderelioglu, M. Salinomycin encapsulated nanoparticles as a targeting vehicle for glioblastoma cells. J. Biomed. Mater. Res. A 2016, 104, 455–464. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Ionophore | Transported Ion | Cancer Type | Target CSCs | Reference |
|---|---|---|---|---|
| Nigericin | K+, H+ | Several | Yes | [4,5,6] |
| Salinomycin | K+, Ca2+ | Several | Yes | [7,8,9] |
| Obatoclax | HCO3−, Cl− | Several | Yes | [10,11,12,13,14,15] |
| Gramicidin | H+, Na+, K+ | Renal cell carcinoma, | Not known | [16,17] |
| Ionomycin | Ca2+ | Breast | Not known | [18,19] |
| Monensin | Na+, H+ | Glioblastoma, Bladder | Not known | [20,21,22] |
| Valinomycin | K+ | Ovarian, Colorectal, | Likely | [3,23,24] |
| Lasalocid | K+, Na+, Ca2+, | Prostate | Not known | [25] |
| Enniatin | Mg2+ | Colon, Ovarian | Not known | [26,27] |
| Beauvericin | NH4+, Ca2+, Ba2+ | Prostate Cervical, Colorectal Hepatoma, Lung | Not known | [28,29] |
| Ionophore | Condition | Other Drugs | Phase | Clinical Trial | Published Results |
|---|---|---|---|---|---|
| Salinomycin | - | ||||
| Nigericin | - | ||||
| Obatoclax | AML | - | NCT00684918 | [160] | |
| Chronic Lymphocytic Leukemia | - | I/II | NCT00600964 | [157] | |
| Extensive-stage Small-Cell Lung Cancer | Carboplatin/etoposide | I/II | NCT00682981 | [163] | |
| Lymphoma, Follicular | Rituximab | NCT00427856 | - | ||
| Non-Hodgkin Lymphoma Recurrent Adult Diffuse Large Cell Lymphoma Recurrent Grade 1 Follicular Lymphoma (and 5 more…) | Bortezomib | NCT00538187 | - | ||
| B-cell Chronic Lymphocytic Leukemia Leukemia Prolymphocytic Leukemia (and 5 more…) | Fludarabine rituximab | NCT00612612 | - | ||
| Leukemia Systemic Mastocytosis | NCT00918931 | - | |||
| Mantle-Cell Lymphoma | Bortezomib | NCT00407303 | - | ||
| Hodgkin’s Lymphoma | NCT00359892 | [161] | |||
| Extensive-stage Small-Cell Lung Cancer | Carboplatine and Etoposide | NCT01563601 | - | ||
| Lung Cancer | Docetaxel | NCT00405951 | - | ||
| Myelodysplastic Syndromes | NCT00413114 | [162] | |||
| Acute Leukemias of Ambiguous Lineage Acute Undifferentiated Leukemia Angioimmunoblastic T-cell Lymphoma (and 26 more…) | Dexrazoxane hydrochloride Doxorubicin hydrochloride (and 3 more…) | NCT00933985 | [165] | ||
| Refractory Multiple Myeloma Stage I Multiple Myeloma Stage II Multiple Myeloma Stage III Multiple Myeloma | Bortezomib | NCT00719901 | - | ||
| Recurrent Small-Cell Lung Cancer Unspecified Adult Solid Tumor | Topotecan hydrochloride | NCT00521144 | [164] | ||
| Extranodal Marginal Zone B-cell Lymphoma of Mucosa-associated Lymphoid Tissue Nodal Marginal Zone B-cell Lymphoma Recurrent Grade 1 Follicular Lymphoma (and 4 more…) | Bendamustine hydrochloride | NCT01238146 | - | ||
| Myelofibrosis | NCT00360035 | - | |||
| Hematological Malignancies | NCT00438178 | - | |||
| Leukemia (samples) | NCT01150656 | - | |||
| Metastatic Melanoma | Temozolomide | I/II | NCT00724841 | - |
| Nanoparticle | Cancer Type | Efficacy | Reference |
|---|---|---|---|
| SS lipid-polymer hybrid nanoparticles | Lung | ↑ | [180] |
| CESP * | Osteosarcoma | ↑ | [181] |
| CD133-SAL-NP | CD133+ ovarian cancer stem cells and nude mice bearing ovarian cancer xenografts | ↑ | [182] |
| Poly (lactic-co-glycolic acid) (PLGA) nanoparticles | Pancreatic cancer | Blocked tumor growth by 52% compared to the control. | [183] |
| rGO-Ag | Human ovarian cancer stem cells | ↑ | [184] |
| EGFR-SNPs | Osteosarcoma and cancer stem cells | ↑ | [185] |
| CD20-SA-NPs | Human CD20+ melanoma stem cells | ↑ | [186] |
| Salinomycin-NPs + gefitinib-NPs | Lung cancer and lung cancer stem cells | ↑ | [187] |
| Sali-NP-HER2 | HER2-positive breast cancer stem cells and cancer cells | ↑ | [188] |
| Salinomycin-NPs + docetaxel-NPs | Gastric cancer cells and cancer stem cells | ↑ | [189] |
| SDLN | Liver cancer cells and cancer stem cells | ↑ | [190] |
| iTEP-Sali-ABA NP + iTEP NP-delivered paclitaxel | Metastases of 4T1 orthotopic breast tumors | ↑ | [191] |
| Salinomycin-NPs + Paclitaxell-NPs | Breast cancer stem cells and cancer cells | ↑ | [192] |
| P80-SAL-PLGA | Glioblastoma | ↑ | [193] |
| CESN | Hepatocellular carcinoma | ↑ | [115] |
| Ap-SAL-NP | Osteosarcoma cancer stem cells | ↑ | [118] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaushik, V.; Yakisich, J.S.; Kumar, A.; Azad, N.; Iyer, A.K.V. Ionophores: Potential Use as Anticancer Drugs and Chemosensitizers. Cancers 2018, 10, 360. https://doi.org/10.3390/cancers10100360
Kaushik V, Yakisich JS, Kumar A, Azad N, Iyer AKV. Ionophores: Potential Use as Anticancer Drugs and Chemosensitizers. Cancers. 2018; 10(10):360. https://doi.org/10.3390/cancers10100360
Chicago/Turabian StyleKaushik, Vivek, Juan Sebastian Yakisich, Anil Kumar, Neelam Azad, and Anand K. V. Iyer. 2018. "Ionophores: Potential Use as Anticancer Drugs and Chemosensitizers" Cancers 10, no. 10: 360. https://doi.org/10.3390/cancers10100360
APA StyleKaushik, V., Yakisich, J. S., Kumar, A., Azad, N., & Iyer, A. K. V. (2018). Ionophores: Potential Use as Anticancer Drugs and Chemosensitizers. Cancers, 10(10), 360. https://doi.org/10.3390/cancers10100360