A Cancer-Favoring, Engineered Vaccinia Virus for Cholangiocarcinoma



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
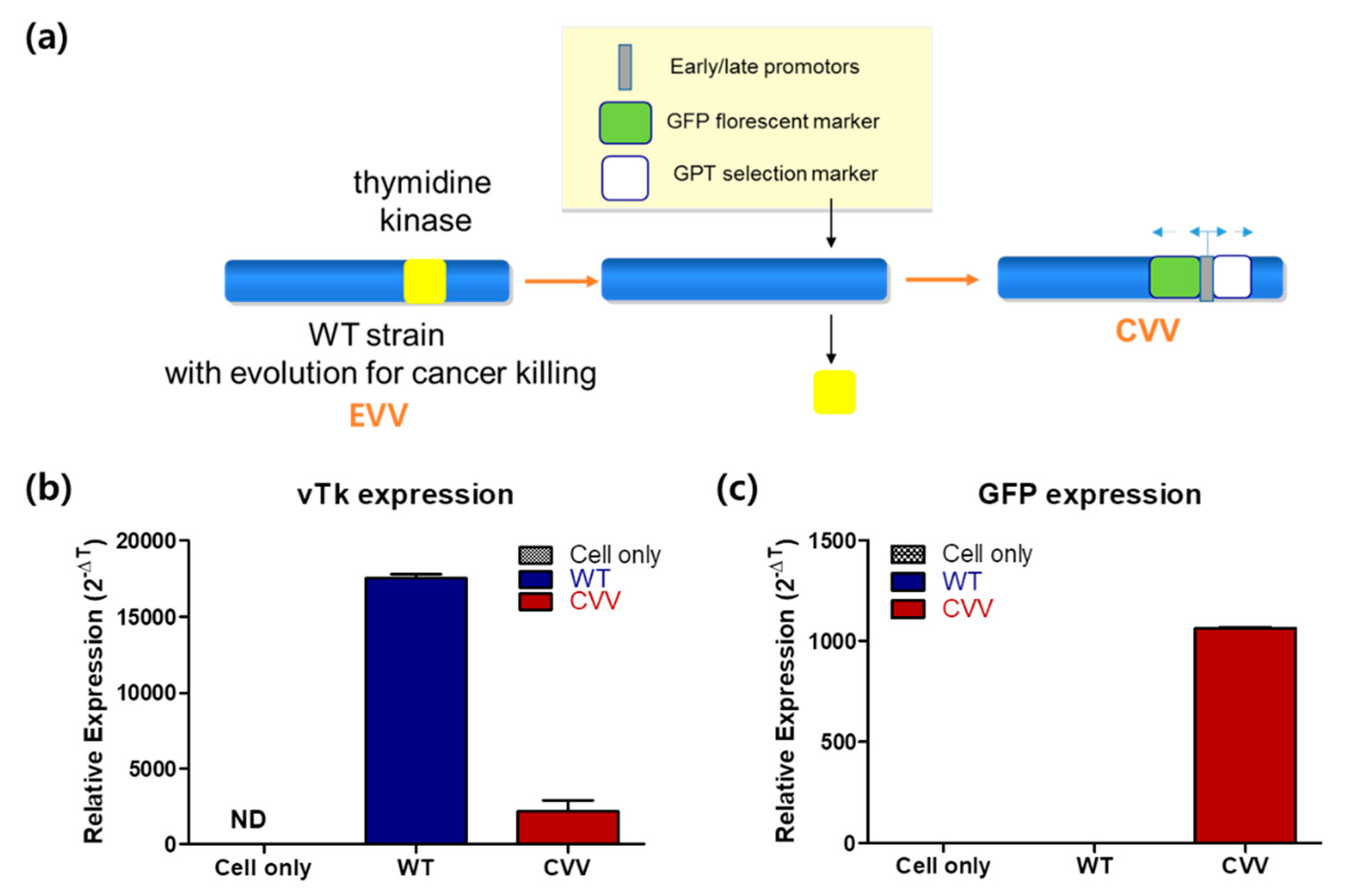
2.1. Cancer-Favoring, Evolutionarily-Engineered Vaccinia Virus
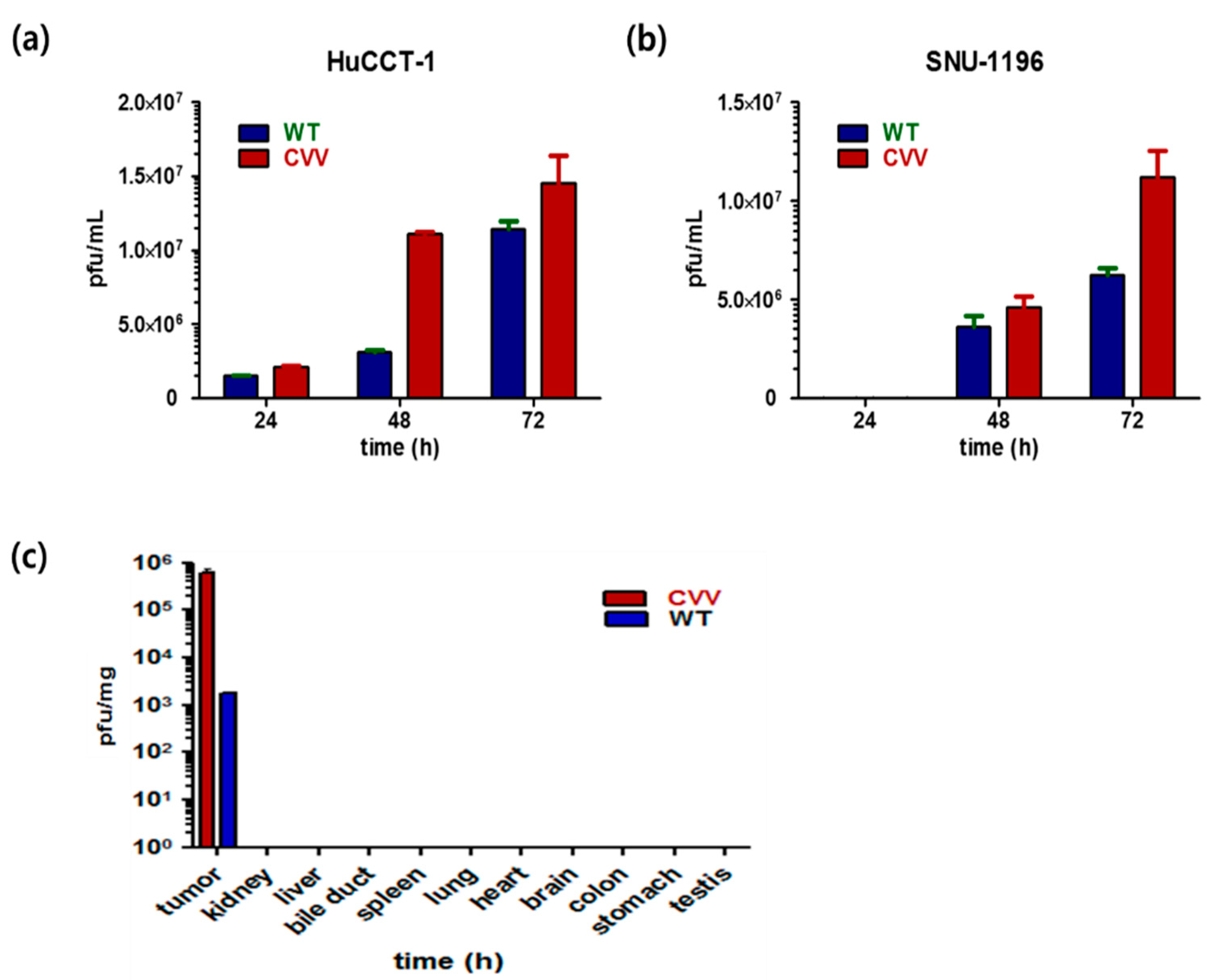
2.2. High Replication Efficacy of the CVV in CCA Cell Lines and Tumors
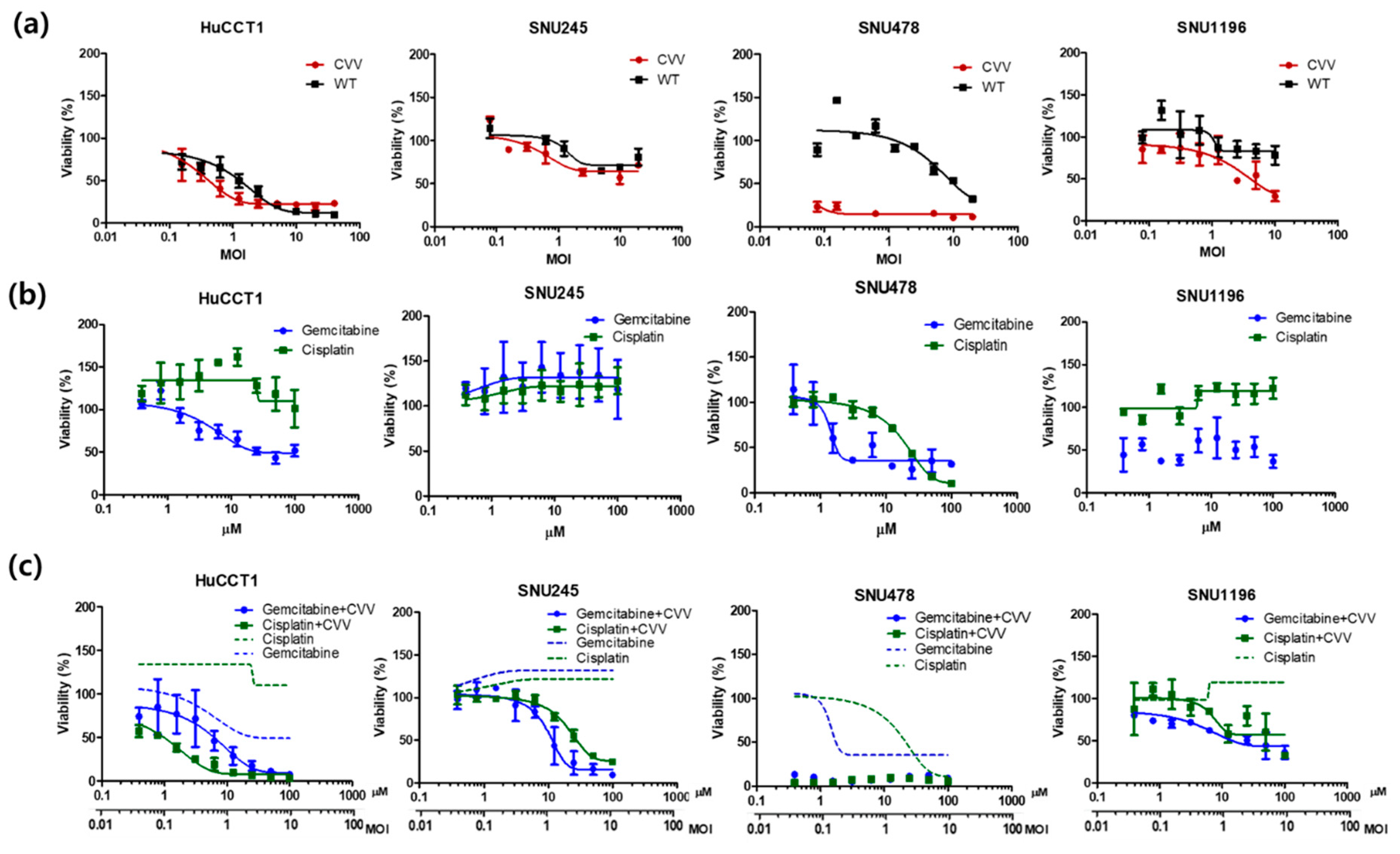
2.3. Oncolytic Efficacy of the CVV Enhances the Therapeutic Efficacy of Chemotherapeutic Drugs In Vitro
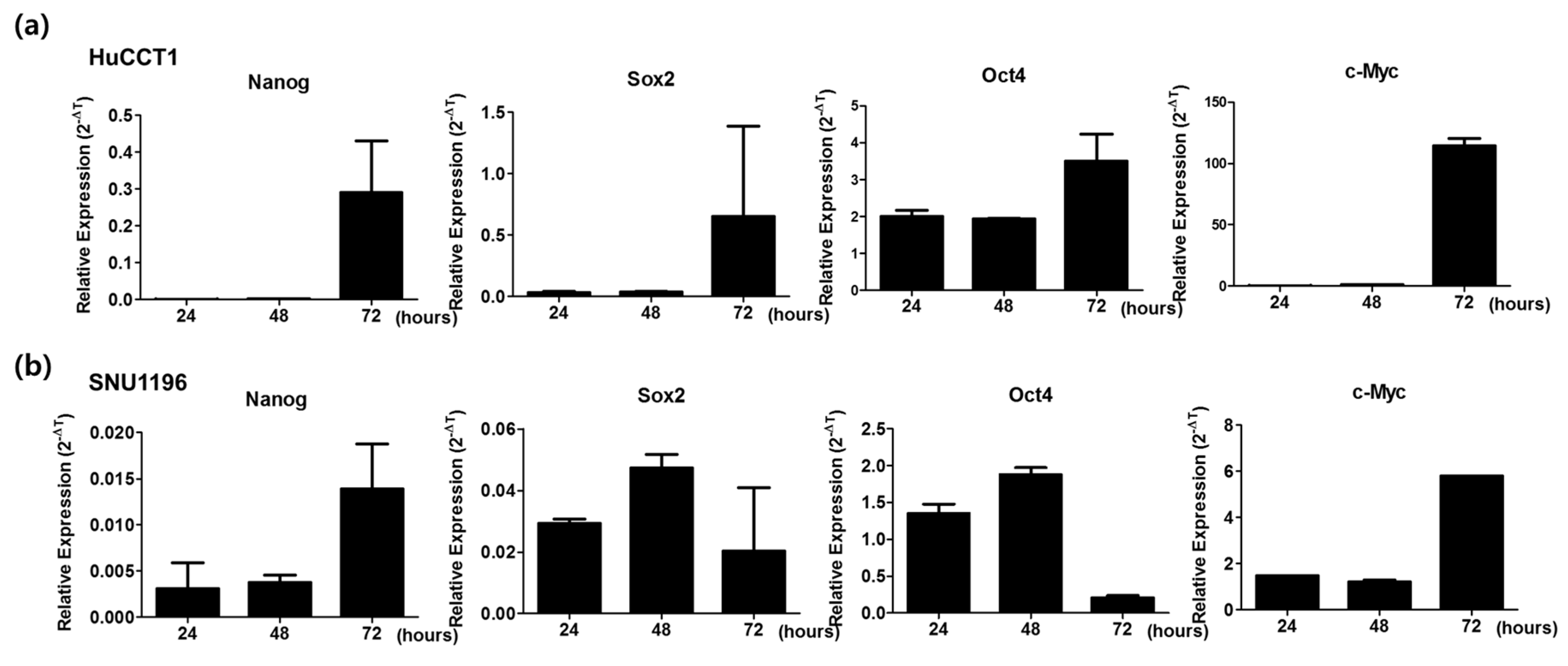
2.4. High Expression of Stemness Markers in CCA Cell Lines
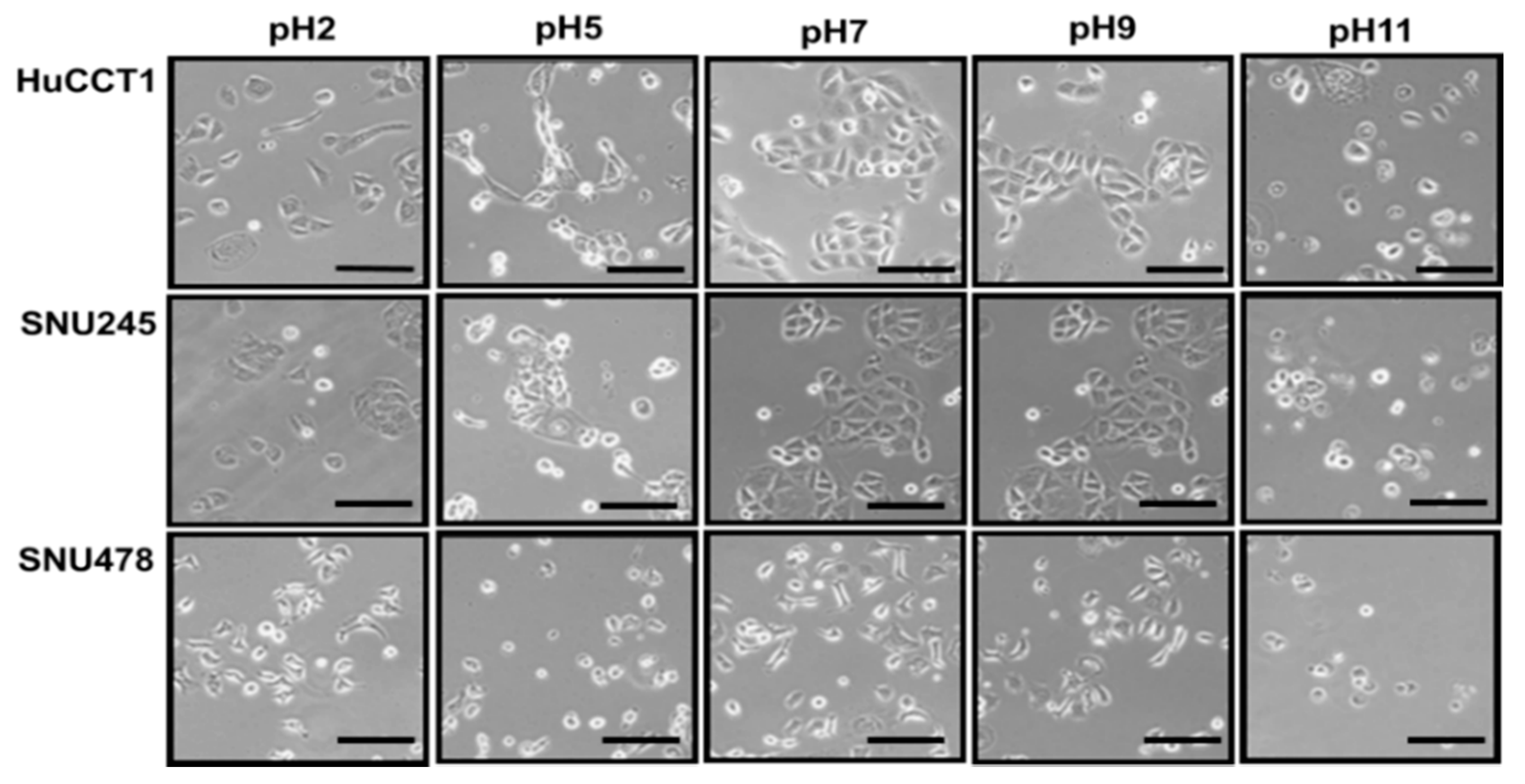
2.5. The Effect of pH on CCA Cell Lines
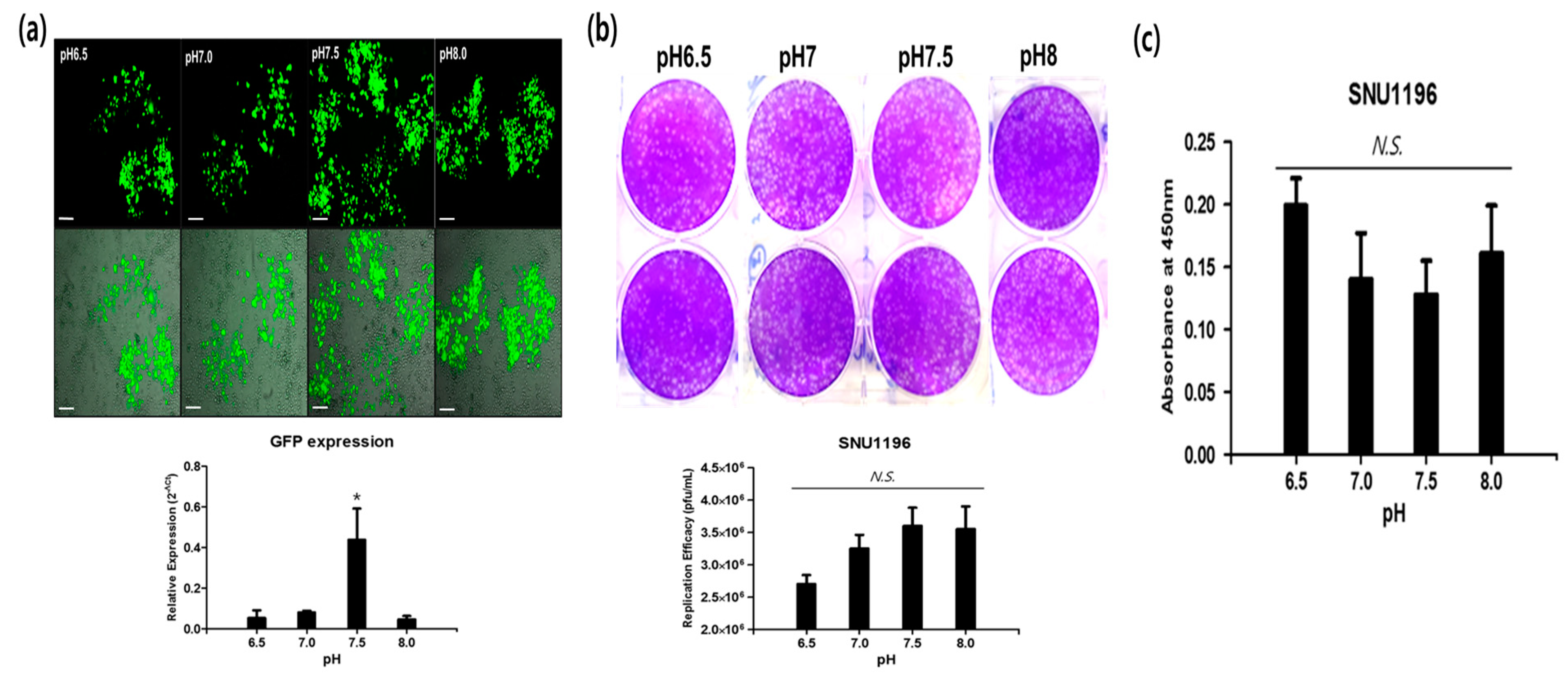
2.6. Alkaline pH Favors CVV Infection in the CCA Cell Line
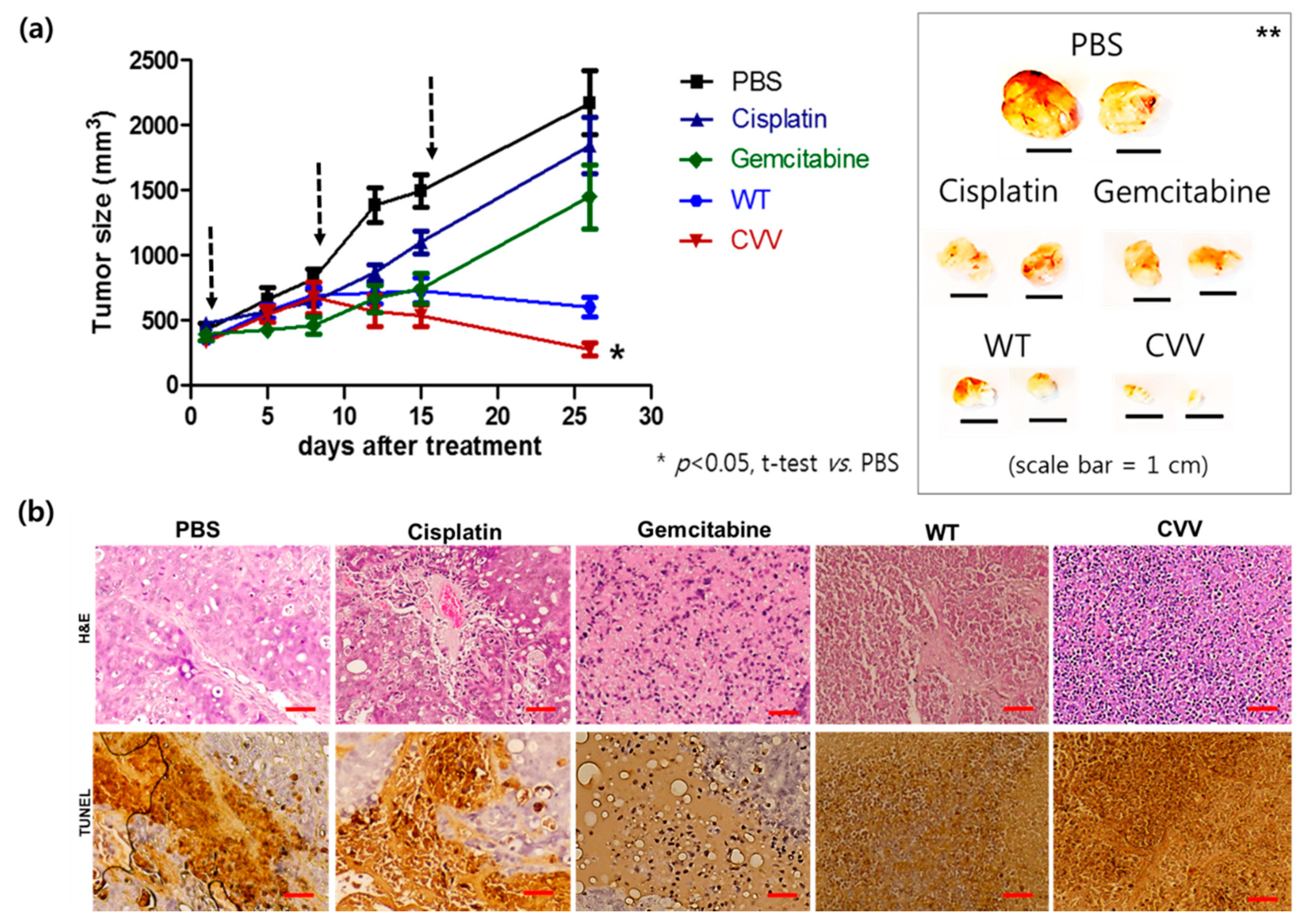
2.7. Therapeutic Efficacy of CVV in the Xenograft Model
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Virus
4.2. Replication Efficacy Assay
4.3. WST-1 Assay
4.4. Cells Culturing in pH-Adjusted Media
4.5. Real-Time PCR Assay
4.6. Animal Study
4.7. H&E staining and TUNEL Assay
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Lee, H.-S.; Kang, J.-I.; Chung, W.-J.; Lee, D.H.; Lee, B.Y.; Lee, S.-W.; Yoo, S.Y. Engineered Phage Matrix Stiffness-Modulating Osteogenic Differentiation. ACS Appl. Mater. Interfaces 2018, 10, 4349–4358. [Google Scholar] [CrossRef] [PubMed]
- Moon, J.-S.; Kim, W.-G.; Shin, D.-M.; Lee, S.-Y.; Kim, C.; Lee, Y.; Han, J.; Kim, K.; Yoo, S.Y.; Oh, J.-W. Bioinspired M-13 bacteriophage-based photonic nose for differential cell recognition. Chem. Sci. 2017, 8, 921–927. [Google Scholar] [CrossRef] [PubMed]
- Yoo, S.Y.; Badrinath, N.; Woo, H.Y.; Heo, J. Oncolytic Virus-Based Immunotherapies for Hepatocellular Carcinoma. Mediat. Inflamm. 2017, 2017, 5198798. [Google Scholar] [CrossRef] [PubMed]
- Yoo, S.Y.; Jeong, S.N.; Kang, J.I.; Lee, S.W. Chimeric Adeno-Associated Virus-Mediated Cardiovascular Reprogramming for Ischemic Heart Disease. ACS Omega 2018, 3, 5918–5925. [Google Scholar] [CrossRef]
- Yoo, S.Y.; Jin, H.-E.; Choi, D.S.; Kobayashi, M.; Farouz, Y.; Wang, S.; Lee, S.-W. M13 Bacteriophage and Adeno-Associated Virus Hybrid for Novel Tissue Engineering Material with Gene Delivery Functions. Adv. Healthc. Mater. 2016, 5, 88–93. [Google Scholar] [CrossRef]
- Yoo, S.Y.; Shrestha, K.R.; Jeong, S.N.; Kang, J.I.; Lee, S.W. Engineered phage nanofibers induce angiogenesis. Nanoscale 2017, 9, 17109–17117. [Google Scholar] [CrossRef]
- Heo, J.; Reid, T.; Ruo, L.; Breitbach, C.J.; Rose, S.; Bloomston, M.; Cho, M.; Lim, H.Y.; Chung, H.C.; Kim, C.W.; et al. Randomized dose-finding clinical trial of oncolytic immunotherapeutic vaccinia JX-594 in liver cancer. Nat. Med. 2013, 19, 329–336. [Google Scholar] [CrossRef]
- Varela-Guruceaga, M.; Tejada-Solís, S.; García-Moure, M.; Fueyo, J.; Gomez-Manzano, C.; Patiño-García, A.; Alonso, M.M. Oncolytic Viruses as Therapeutic Tools for Pediatric Brain Tumors. Cancers 2018, 10, 226. [Google Scholar] [CrossRef]
- Badrinath, N.; Heo, J.; Yoo, S.Y. Viruses as nanomedicine for cancer. Int. J. Nanomed. 2016, 11, 4835–4847. [Google Scholar] [CrossRef]
- Ricordel, M.; Foloppe, J.; Antoine, D.; Findeli, A.; Kempf, J.; Cordier, P.; Gerbaud, A.; Grellier, B.; Lusky, M.; Quemeneur, E.; et al. Vaccinia Virus Shuffling: deVV5, a Novel Chimeric Poxvirus with Improved Oncolytic Potency. Cancers 2018, 10, 231. [Google Scholar] [CrossRef]
- Guse, K.; Cerullo, V.; Hemminki, A. Oncolytic vaccinia virus for the treatment of cancer. Expert Opin. Biol. Ther. 2011, 11, 595–608. [Google Scholar] [CrossRef] [PubMed]
- Parato, K.A.; Breitbach, C.J.; Le Boeuf, F.; Wang, J.; Storbeck, C.; Ilkow, C.; Diallo, J.S.; Falls, T.; Burns, J.; Garcia, V.; et al. The oncolytic poxvirus JX-594 selectively replicates in and destroys cancer cells driven by genetic pathways commonly activated in cancers. Mol. Ther. J. Am. Soc. Gene Ther. 2012, 20, 749–758. [Google Scholar] [CrossRef] [PubMed]
- Furuse, J.; Okusaka, T. Targeted Therapy for Biliary Tract Cancer. Cancers 2011, 3, 2243–2254. [Google Scholar] [CrossRef] [PubMed]
- Kokuryo, T.; Yokoyama, Y.; Nagino, M. Recent advances in cancer stem cell research for cholangiocarcinoma. J. Hepato Biliary Pancreat. Sci. 2012, 19, 606–613. [Google Scholar] [CrossRef] [PubMed]
- Yoo, S.Y.; Bang, S.Y.; Jeong, S.N.; Kang, D.H.; Heo, J. A cancer-favoring oncolytic vaccinia virus shows enhanced suppression of stem-cell like colon cancer. Oncotarget 2016, 7, 16479–16489. [Google Scholar] [CrossRef]
- Yoo, S.Y.; Jeong, S.N.; Kang, D.H.; Heo, J. Evolutionary cancer-favoring engineered vaccinia virus for metastatic hepatocellular carcinoma. Oncotarget 2017, 8, 71489–71499. [Google Scholar] [CrossRef]
- Gillies, R.J.; Liu, Z.; Bhujwalla, Z. 31P-MRS measurements of extracellular pH of tumors using 3-aminopropylphosphonate. Am. J. Physiol. 1994, 267, C195–C203. [Google Scholar] [CrossRef]
- Vaupel, P.; Kallinowski, F.; Okunieff, P. Blood flow, oxygen and nutrient supply, and metabolic microenvironment of human tumors: A review. Cancer Res. 1989, 49, 6449–6465. [Google Scholar]
- Raggi, C.; Invernizzi, P.; Andersen, J.B. Impact of microenvironment and stem-like plasticity in cholangiocarcinoma: Molecular networks and biological concepts. J. Hepatol. 2015, 62, 198–207. [Google Scholar] [CrossRef]
- Griffiths, J.R. Are cancer cells acidic? Br. J. Cancer 1991, 64, 425–427. [Google Scholar] [CrossRef]
- Moeini, A.; Sia, D.; Bardeesy, N.; Mazzaferro, V.; Llovet, J.M. Molecular Pathogenesis and Targeted Therapies for Intrahepatic Cholangiocarcinoma. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2016, 22, 291–300. [Google Scholar] [CrossRef] [PubMed]
- Badrinath, N.; Yoo, S.Y. Recent Advances in Cancer Stem Cell-Targeted Immunotherapy. Cancers 2019, 11, 310. [Google Scholar] [CrossRef] [PubMed]
- Coelen, R.J.S.; de Keijzer, M.J.; Weijer, R.; Loukachov, V.V.; Wiggers, J.K.; Mul, F.P.J.; van Wijk, A.; Fong, Y.; Heger, M.; van Gulik, T.M. In vitro detection of cholangiocarcinoma cells using a fluorescent protein-expressing oncolytic herpes virus. Cancer Gene Ther. 2017, 24, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Pugalenthi, A.; Mojica, K.; Ady, J.W.; Johnsen, C.; Love, D.; Chen, N.G.; Aguilar, R.J.; Szalay, A.A.; Fong, Y. Recombinant vaccinia virus GLV-1h68 is a promising oncolytic vector in the treatment of cholangiocarcinoma. Cancer Gene Ther. 2015, 22, 591–596. [Google Scholar] [CrossRef]
- Lange, S.; Lampe, J.; Bossow, S.; Zimmermann, M.; Neubert, W.; Bitzer, M.; Lauer, U.M. A novel armed oncolytic measles vaccine virus for the treatment of cholangiocarcinoma. Hum. Gene Ther. 2013, 24, 554–564. [Google Scholar] [CrossRef]
- Hao, G.; Xu, Z.P.; Li, L. Manipulating extracellular tumour pH: An effective target for cancer therapy. RSC Adv. 2018, 8, 22182–22192. [Google Scholar] [CrossRef]
- Breitbach, C.J.; Arulanandam, R.; De Silva, N.; Thorne, S.H.; Patt, R.; Daneshmand, M.; Moon, A.; Ilkow, C.; Burke, J.; Hwang, T.H.; et al. Oncolytic vaccinia virus disrupts tumor-associated vasculature in humans. Cancer Res. 2013, 73, 1265–1275. [Google Scholar] [CrossRef]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoo, S.Y.; Badrinath, N.; Lee, H.L.; Heo, J.; Kang, D.-H. A Cancer-Favoring, Engineered Vaccinia Virus for Cholangiocarcinoma. Cancers 2019, 11, 1667. https://doi.org/10.3390/cancers11111667
Yoo SY, Badrinath N, Lee HL, Heo J, Kang D-H. A Cancer-Favoring, Engineered Vaccinia Virus for Cholangiocarcinoma. Cancers. 2019; 11(11):1667. https://doi.org/10.3390/cancers11111667
Chicago/Turabian StyleYoo, So Young, Narayanasamy Badrinath, Hye Lim Lee, Jeong Heo, and Dae-Hwan Kang. 2019. "A Cancer-Favoring, Engineered Vaccinia Virus for Cholangiocarcinoma" Cancers 11, no. 11: 1667. https://doi.org/10.3390/cancers11111667
APA StyleYoo, S. Y., Badrinath, N., Lee, H. L., Heo, J., & Kang, D.-H. (2019). A Cancer-Favoring, Engineered Vaccinia Virus for Cholangiocarcinoma. Cancers, 11(11), 1667. https://doi.org/10.3390/cancers11111667