ACLY (ATP Citrate Lyase) Mediates Radioresistance in Head and Neck Squamous Cell Carcinomas and is a Novel Predictive Radiotherapy Biomarker

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
2.1. Multiple Metabolic Genes Associate with Radioresistance and –Sensitivity
2.2. Inhibition of ACLY Affects DNA Damage Repair and Radiosensitizes HNSCC Cells
2.3. High ACLY Levels are Associated with Poor Prognosis in HNSCC Patients Receiving Radiotherapy
2.4. Nuclear ACLY Localisation is Associated with Locoregional Control in HNSCC Patients
3. Discussion
4. Materials and Methods
4.1. Cell Lines, Reagents, and Irradiation
4.2. Homologous Recombination Assay
4.3. Single Molecule Molecular Inversion Probe Sequencing
4.4. Colony Forming Assays
4.5. Radiosensitivity Analysis and Linear Quadratic Fitting
4.6. siRNA Interference and Real Time Quantitative PCR (RT-qPCR)
4.7. Immunofluorescence
4.8. TCGA Data Extraction and Selection Criteria
4.9. TCGA Data Analysis
4.10. Validation Dataset
4.11. Immunohistochemistry
4.12. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Torre, L.A.; Bray, F.; Siegel, R.L.; Ferlay, J.; Lortet-Tieulent, J.; Jemal, A. Global cancer statistics, 2012. CA Cancer J. Clin. 2015, 65, 87–108. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Zhang, J.; Shi, C.; Liu, L.; Wei, Y. Survival, recurrence and toxicity of HNSCC in comparison of a radiotherapy combination with cisplatin versus cetuximab: A meta-analysis. BMC Cancer 2016, 16. [Google Scholar] [CrossRef] [Green Version]
- D’Souza, G.; Westra, W.H.; Wang, S.J.; van Zante, A.; Wentz, A.; Kluz, N.; Rettig, E.; Ryan, W.R.; Ha, P.K.; Kang, H.; et al. Differences in the Prevalence of Human Papillomavirus (HPV) in Head and Neck Squamous Cell Cancers by Sex, Race, Anatomic Tumor Site, and HPV Detection Method. JAMA Oncol. 2017, 3, 169–177. [Google Scholar] [CrossRef]
- Rieckmann, T.; Tribius, S.; Grob, T.J.; Meyer, F.; Busch, C.J.; Petersen, C.; Dikomey, E.; Kriegs, M. HNSCC cell lines positive for HPV and p16 possess higher cellular radiosensitivity due to an impaired DSB repair capacity. Radiother. Oncol. 2013, 107, 242–246. [Google Scholar] [CrossRef]
- Göttgens, E.L.; Bussink, J.; Leszczynska, K.B.; Peters, H.; Span, P.N.; Hammond, E.M. Inhibition of CDK4/CDK6 Enhances Radiosensitivity of HPV Negative Head and Neck Squamous Cell Carcinomas. Int. J. Radiat. Oncol. Biol. Phys. 2019, 105, 548–558. [Google Scholar] [CrossRef]
- Ang, K.K.; Harris, J.; Wheeler, R.; Weber, R.; Rosenthal, D.I.; Nguyen-Tân, P.F.; Westra, W.H.; Chung, C.H.; Jordan, R.C.; Lu, C.; et al. Human papillomavirus and survival of patients with oropharyngeal cancer. N. Engl. J. Med. 2010, 363, 24–35. [Google Scholar] [CrossRef] [Green Version]
- Fakhry, C.; Westra, W.H.; Li, S.; Cmelak, A.; Ridge, J.A.; Pinto, H.; Forastiere, A.; Gillison, M.L. Improved survival of patients with human papillomavirus-positive head and neck squamous cell carcinoma in a prospective clinical trial. J. Natl. Cancer Inst. 2008, 100, 261–269. [Google Scholar] [CrossRef] [Green Version]
- Kimple, R.J.; Smith, M.A.; Blitzer, G.C.; Torres, A.D.; Martin, J.A.; Yang, R.Z.; Peet, C.R.; Lorenz, L.D.; Nickel, K.P.; Klingelhutz, A.J.; et al. Enhanced radiation sensitivity in HPV-positive head and neck cancer. Cancer Res. 2013, 73, 4791–4800. [Google Scholar] [CrossRef] [Green Version]
- Tang, L.; Wei, F.; Wu, Y.; He, Y.; Shi, L.; Xiong, F.; Gong, Z.; Guo, C.; Li, X.; Deng, H.; et al. Role of metabolism in cancer cell radioresistance and radiosensitization methods. J. Exp. Clin. Cancer Res. 2018, 37. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, P.; Kamarajan, P.; Somashekar, B.S.; MacKinnon, N.; Chinnaiyan, A.M.; Kapila, Y.L.; Rajendiran, T.M.; Ramamoorthy, A. Delineating metabolic signatures of head and neck squamous cell carcinoma: Phospholipase A2, a potential therapeutic target. Int. J. Biochem. Cell Biol. 2012, 44, 1852–1861. [Google Scholar] [CrossRef] [Green Version]
- Mims, J.; Bansal, N.; Bharadwaj, M.S.; Chen, X.; Molina, A.J.; Tsang, A.W.; Furdui, C.M. Energy metabolism in a matched model of radiation resistance for head and neck squamous cell cancer. Radiat. Res. 2015, 183, 291–304. [Google Scholar] [CrossRef] [Green Version]
- Sandulache, V.C.; Ow, T.J.; Pickering, C.R.; Frederick, M.J.; Zhou, G.; Fokt, I.; Davis-Malesevich, M.; Priebe, W.; Myers, J.N. Glucose, not glutamine, is the dominant energy source required for proliferation and survival of head and neck squamous carcinoma cells. Cancer 2011, 117, 2926–2938. [Google Scholar] [CrossRef]
- Meijer, T.W.; Kaanders, J.H.; Span, P.N.; Bussink, J. Targeting hypoxia, HIF-1, and tumor glucose metabolism to improve radiotherapy efficacy. Clin. Cancer Res. 2012, 18, 5585–5594. [Google Scholar] [CrossRef] [Green Version]
- Sivanand, S.; Rhoades, S.; Jiang, Q.; Lee, J.V.; Benci, J.; Zhang, J.; Yuan, S.; Viney, I.; Zhao, S.; Carrer, A.; et al. Nuclear Acetyl-CoA Production by ACLY Promotes Homologous Recombination. Mol. Cell 2017, 67, 252–265. [Google Scholar] [CrossRef] [Green Version]
- de Bitter, T.; van de Water, C.; van den Heuvel, C.; Zeelen, C.; Eijkelenboom, A.; Tops, B.; Oosterwijk, E.; Kolev, D.; Mulders, P.; Laan, M.t.; et al. Profiling of the metabolic transcriptome via single molecule molecular inversion probes. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef]
- van den Heuvel, C.; van Ewijk, A.; Zeelen, C.; de Bitter, T.; Huynen, M.; Mulders, P.; Oosterwijk, E.; Leenders, W.P.J. Molecular Profiling of Druggable Targets in Clear Cell Renal Cell Carcinoma Through Targeted RNA Sequencing. Front. Oncol. 2019, 9. [Google Scholar] [CrossRef] [Green Version]
- Xie, A.; Hartlerode, A.; Stucki, M.; Odate, S.; Puget, N.; Kwok, A.; Nagaraju, G.; Yan, C.; Alt, F.W.; Chen, J.; et al. Distinct roles of chromatin-associated proteins MDC1 and 53BP1 in mammalian double-strand break repair. Mol. Cell 2007, 28, 1045–1057. [Google Scholar] [CrossRef] [Green Version]
- Pierce, A.J.; Johnson, R.D.; Thompson, L.H.; Jasin, M. XRCC3 promotes homology-directed repair of DNA damage in mammalian cells. Genes Dev. 1999, 13, 2633–2638. [Google Scholar] [CrossRef] [Green Version]
- Budczies, J.; Klauschen, F.; Sinn, B.V.; Győrffy, B.; Schmitt, W.D.; Darb-Esfahani, S.; Denkert, C. Cutoff Finder: A comprehensive and straightforward Web application enabling rapid biomarker cutoff optimization. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [Green Version]
- van der Heijden, M.; de Jong, M.C.; Verhagen, C.V.M.; de Roest, R.H.; Sanduleanu, S.; Hoebers, F.; Leemans, C.R.; Brakenhoff, R.H.; Vens, C.; Verheij, M.; et al. Acute Hypoxia Profile is a Stronger Prognostic Factor than Chronic Hypoxia in Advanced Stage Head and Neck Cancer Patients. Cancers 2019, 11, 583. [Google Scholar] [CrossRef] [Green Version]
- Liskamp, C.P.; Janssens, G.O.; Bussink, J.; Melchers, W.J.; Kaanders, J.H.; Verhoef, C.G. Adverse effect of smoking on prognosis in human papillomavirus-associated oropharyngeal carcinoma. Head Neck 2016, 38, 1780–1787. [Google Scholar] [CrossRef]
- Zhi, X.; Lamperska, K.; Golusinski, P.; Schork, N.J.; Luczewski, L.; Kolenda, T.; Golusinski, W.; Masternak, M.M. Gene expression analysis of head and neck squamous cell carcinoma survival and recurrence. Oncotarget 2015, 6, 547–555. [Google Scholar] [CrossRef]
- Garcia-Fuentes, E.; Santiago-Fernández, C.; Gutiérrez-Repiso, C.; Mayas, M.D.; Oliva-Olivera, W.; Coín-Aragüez, L.; Alcaide, J.; Ocaña-Wilhelmi, L.; Vendrell, J.; Tinahones, F.J.; et al. Hypoxia is associated with a lower expression of genes involved in lipogenesis in visceral adipose tissue. J. Transl. Med. 2015, 13. [Google Scholar] [CrossRef] [Green Version]
- Currie, E.; Schulze, A.; Zechner, R.; Walther, T.C.; Farese, R.V., Jr. Cellular fatty acid metabolism and cancer. Cell Metab. 2013, 18. [Google Scholar] [CrossRef] [Green Version]
- Rae, C.; Haberkorn, U.; Babich, J.W.; Mairs, R.J. Inhibition of Fatty Acid Synthase Sensitizes Prostate Cancer Cells to Radiotherapy. Radiat. Res. 2015, 184, 482–493. [Google Scholar] [CrossRef] [Green Version]
- Zhan, N.; Li, B.; Xu, X.; Xu, J.; Hu, S. Inhibition of FASN expression enhances radiosensitivity in human non-small cell lung cancer. Oncol. Lett. 2018, 15, 4578–4584. [Google Scholar] [CrossRef] [Green Version]
- Sivanand, S.; Viney, I.; Wellen, K.E. Spatiotemporal Control. of Acetyl-CoA Metabolism in Chromatin Regulation. Trends Biochem. Sci. 2018, 43, 61–74. [Google Scholar] [CrossRef]
- Arts, P.; van der Raadt, J.; van Gestel, S.H.C.; Steehouwer, M.; Shendure, J.; Hoischen, A.; Albers, C.A. Quantification of differential gene expression by multiplexed targeted resequencing of cDNA. Nat. Commun. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Boyle, E.A.; O’Roak, B.J.; Martin, B.K.; Kumar, A.; Shendure, J. MIPgen: Optimized modeling and design of molecular inversion probes for targeted resequencing. Bioinformatics 2014, 30, 2670–2672. [Google Scholar] [CrossRef]
- Cancer Genome Atlas Network. Comprehensive genomic characterization of head and neck squamous cell carcinomas. Nature 2015, 517, 576–582. [Google Scholar] [CrossRef] [Green Version]
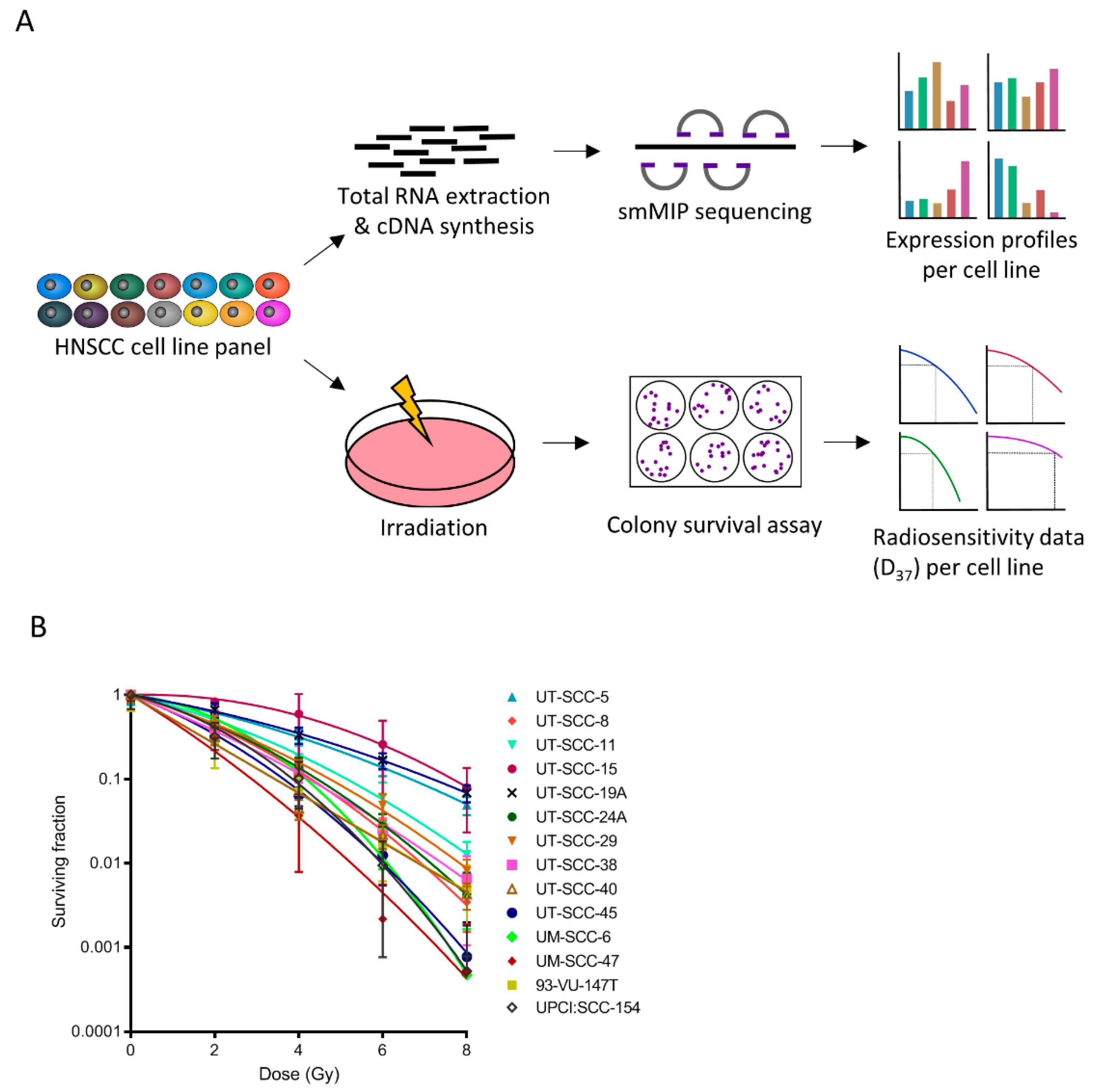
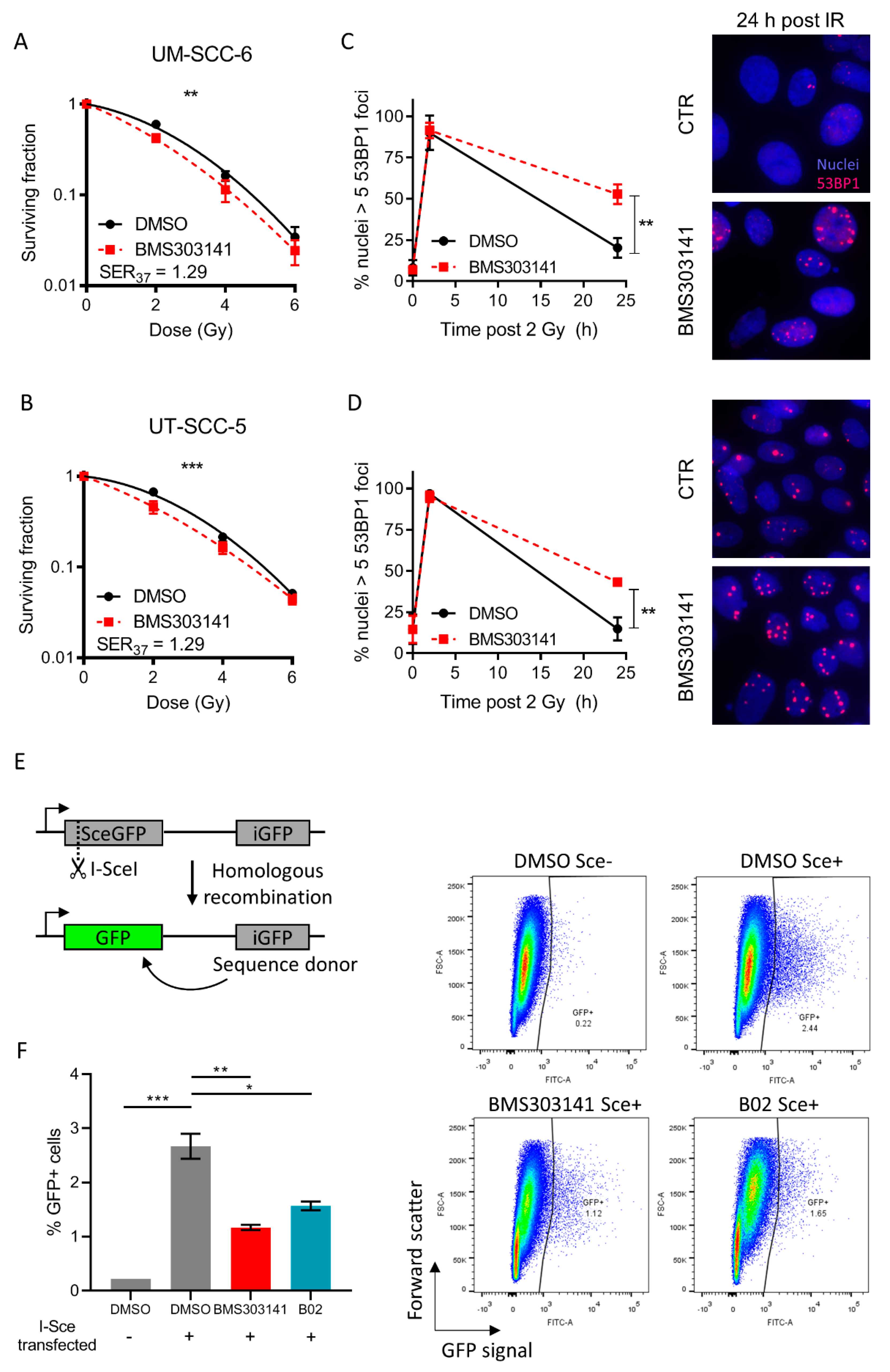



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Encoded Protein | Spearman’s Rho | p-Value | High Expression Associated with Radioresistance/Radiosensitivity |
|---|---|---|---|---|
| ACACA | Acetyl-CoA carboxylase alpha | 0.596 | 0.025 | Resistance |
| ACLY | ATP citrate lyase | 0.670 | 0.009 | Resistance |
| ALDOA | Aldolase, fructose-bisphosphate A | −0.538 | 0.047 | Sensitivity |
| BRAF | B-Raf | 0.657 | 0.011 | Resistance |
| EGF | Epidermal growth factor | 0.573 | 0.032 | Resistance |
| GCLC | Glutamate cysteine ligase | 0.534 | 0.049 | Resistance |
| GLDC | Glycine dehydrogenase | 0.650 | 0.012 | Resistance |
| GLUL | Glutamine synthetase | 0.666 | 0.009 | Resistance |
| GOT1 | Glutamate oxaloacetate transaminase | 0.684 | 0.007 | Resistance |
| HPV | E2, E6, E7 (all transcripts) | −0.547 | 0.043 | Sensitivity |
| IDH3A | Isocitrate dehydrogenase 3, mitochondrial, alpha | 0.578 | 0.030 | Resistance |
| KDR | Vascular endothelial growth factor receptor 2 | 0.602 | 0.023 | Resistance |
| L2HGDH | L-2-hydroxyglutarate dehydrogenase | 0.604 | 0.022 | Resistance |
| MYC | V-myc avian myelocytomatosis viral oncogene homolog | 0.692 | 0.006 | Resistance |
| PFKM | Phosphofructokinase 1 | 0.640 | 0.014 | Resistance |
| RPIA | Ribose 5-phosphate isomerase A | 0.710 | 0.004 | Resistance |
| SDHD | Succinate dehydrogenase complex, subunit D | 0.604 | 0.022 | Resistance |
| Variable | Low ACLY | High ACLY | p-Value | RT− | RT+ | p-Value | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| N | % | N | % | N | % | N | % | ||||
| Gender | Female | 98 | 27.1 | 18 | 21.7 | 0.313 | 55 | 35.0 | 61 | 21.2 | 0.001 * |
| Male | 264 | 72.9 | 65 | 78.3 | 102 | 65.0 | 227 | 78.8 | |||
| Clinical T status | T1 + T2 | 136 | 37.6 | 18 | 21.7 | 0.141 | 84 | 53.5 | 70 | 24.3 | <0.001 ** |
| T3 + T4 | 215 | 59.4 | 63 | 75.9 | 69 | 44.0 | 209 | 72.6 | |||
| Tx or missing | 11 | 3.0 | 2 | 2.4 | 4 | 2.5 | 9 | 3.1 | |||
| Clinical N status | N0 | 168 | 46.4 | 36 | 43.4 | 0.975 | 98 | 62.4 | 106 | 36.8 | <0.001 ** |
| N1-3 | 178 | 49.2 | 45 | 54.2 | 54 | 34.4 | 169 | 58.7 | |||
| Nx or missing | 16 | 4.4 | 2 | 2.4 | 5 | 3.2 | 13 | 4.5 | |||
| Clinical M status | M0 | 343 | 94.8 | 80 | 96.4 | 0.667 | 152 | 96.8 | 271 | 94.1 | <0.001 ** |
| Mx or missing | 19 | 5.2 | 3 | 3.6 | 5 | 3.2 | 17 | 5.9 | |||
| Clinical stage | Stage I + II | 82 | 22.7 | 14 | 16.9 | 0.677 | 66 | 42.0 | 30 | 10.4 | <0.001 ** |
| Stage III + IV | 271 | 74.8 | 67 | 80.7 | 87 | 55.4 | 251 | 87.2 | |||
| Missing | 9 | 2.5 | 2 | 2.4 | 4 | 2.5 | 7 | 2.4 | |||
| Received radiotherapy | Yes | 238 | 65.7 | 50 | 60.2 | 0.344 | 124 | 79.0 | 238 | 82.6 | 0.344 |
| No | 124 | 34.3 | 33 | 39.8 | 33 | 21.0 | 50 | 17.4 | |||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Göttgens, E.-L.; van den Heuvel, C.N.; de Jong, M.C.; Kaanders, J.H.; Leenders, W.P.; Ansems, M.; Bussink, J.; Span, P.N. ACLY (ATP Citrate Lyase) Mediates Radioresistance in Head and Neck Squamous Cell Carcinomas and is a Novel Predictive Radiotherapy Biomarker. Cancers 2019, 11, 1971. https://doi.org/10.3390/cancers11121971
Göttgens E-L, van den Heuvel CN, de Jong MC, Kaanders JH, Leenders WP, Ansems M, Bussink J, Span PN. ACLY (ATP Citrate Lyase) Mediates Radioresistance in Head and Neck Squamous Cell Carcinomas and is a Novel Predictive Radiotherapy Biomarker. Cancers. 2019; 11(12):1971. https://doi.org/10.3390/cancers11121971
Chicago/Turabian StyleGöttgens, Eva-Leonne, Corina NAM van den Heuvel, Monique C de Jong, Johannes HAM Kaanders, William PJ Leenders, Marleen Ansems, Johan Bussink, and Paul N Span. 2019. "ACLY (ATP Citrate Lyase) Mediates Radioresistance in Head and Neck Squamous Cell Carcinomas and is a Novel Predictive Radiotherapy Biomarker" Cancers 11, no. 12: 1971. https://doi.org/10.3390/cancers11121971