Gliosarcoma Is Driven by Alterations in PI3K/Akt, RAS/MAPK Pathways and Characterized by Collagen Gene Expression Signature

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Results
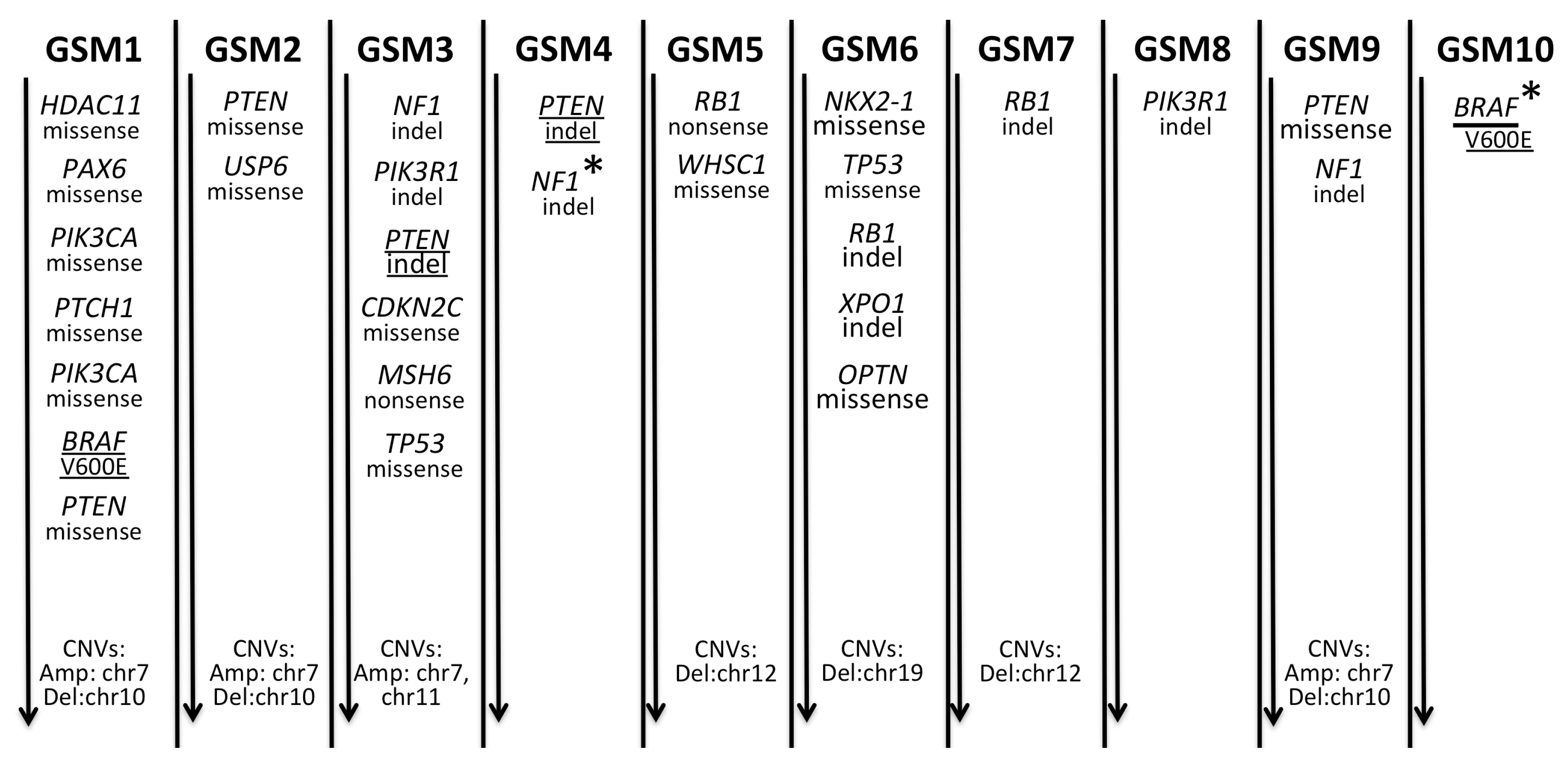
2.1. A Spectrum of Somatic Mutations and Indels in Gliosarcomas
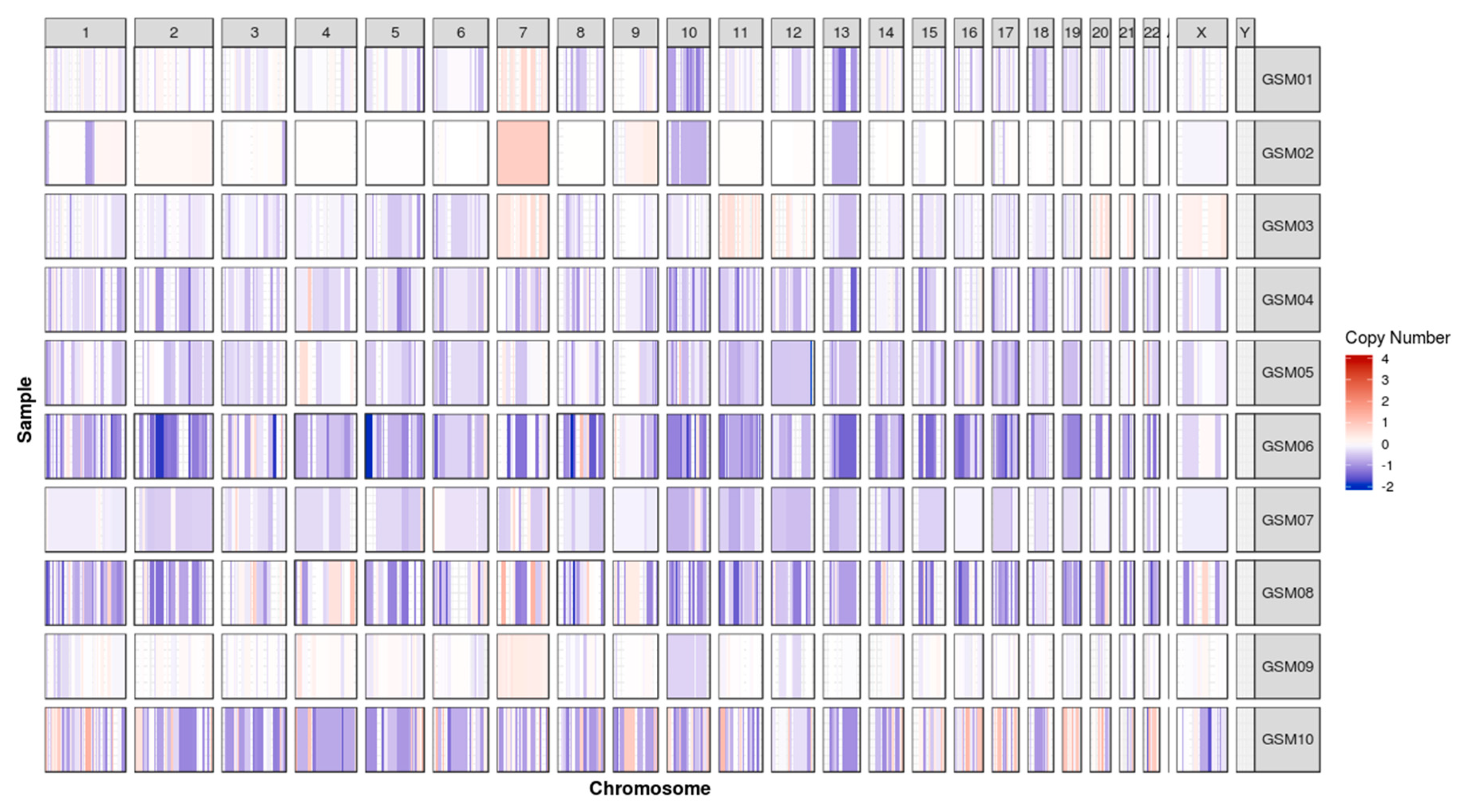
2.2. Chromosomal Gain/Loss in Gliosarcomas
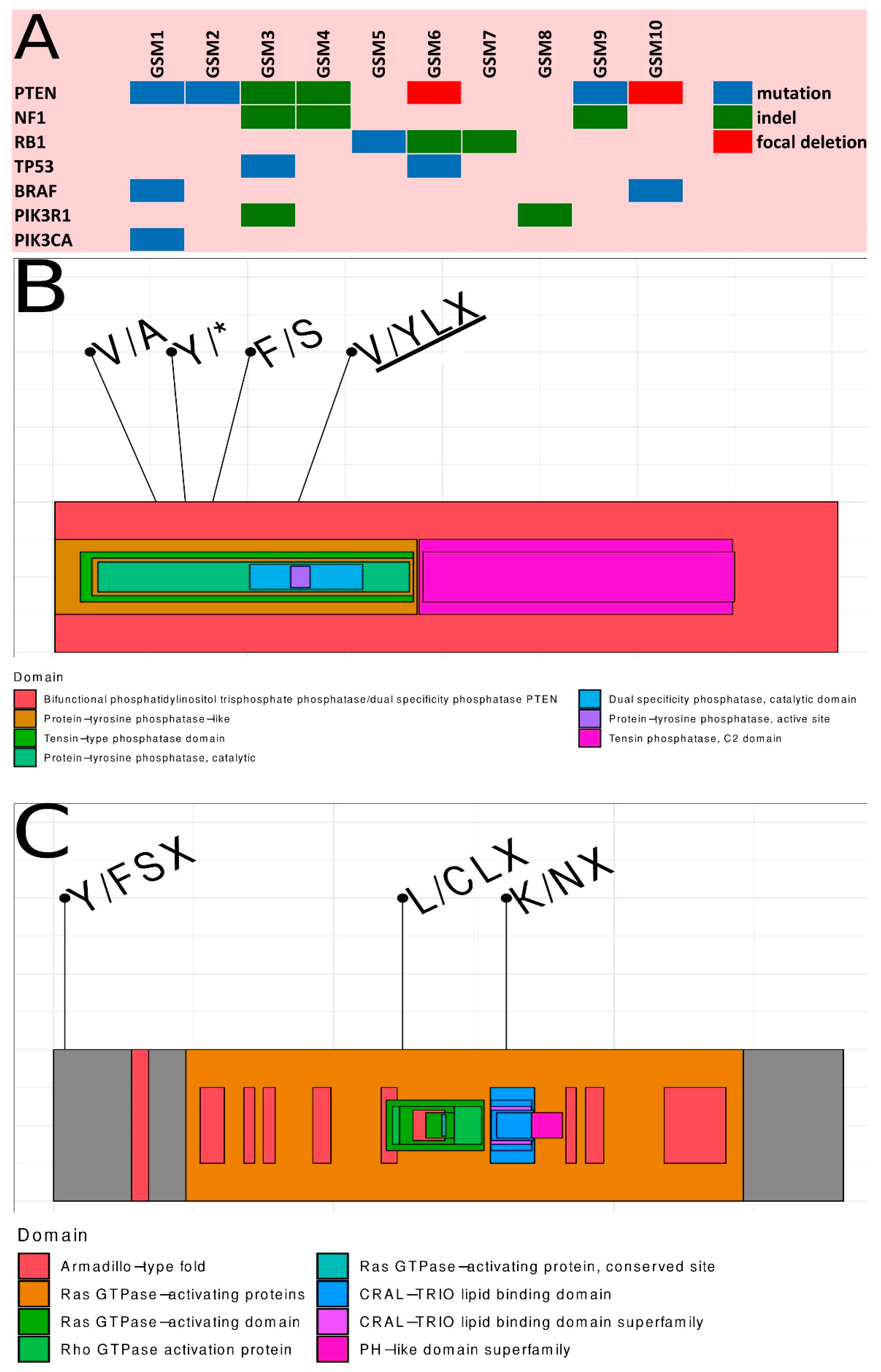
2.3. Summary of Somatic Alterations in Gliosarcoma
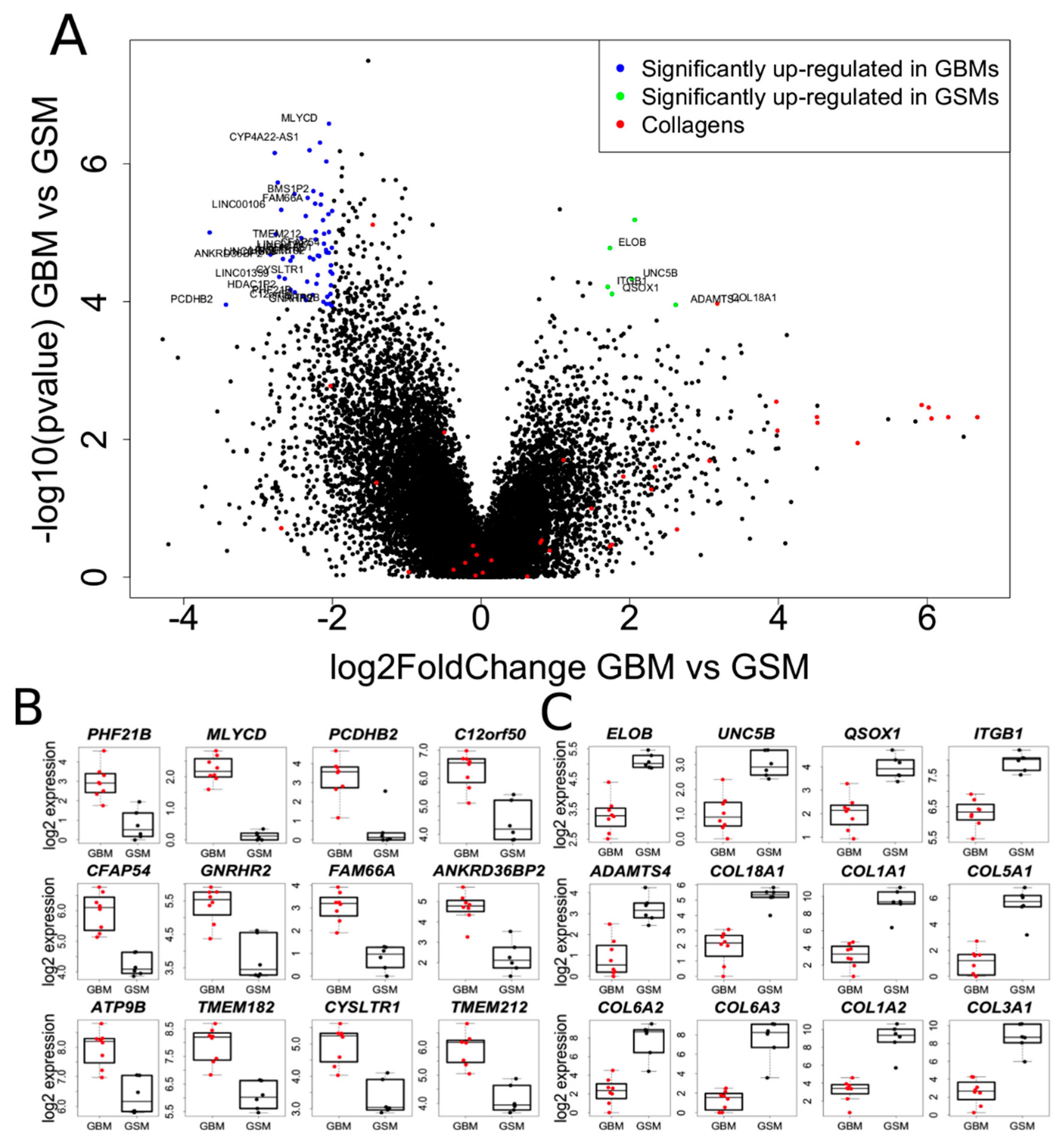
2.4. Transcriptome Analysis of Gliosarcoma and Comparison to GBM
2.5. Translocations in Gliosarcomas and Their Biological Consequences
2.6. Collagens Expression in Gliomatous and Sacromatous Compartments
3. Discussion
4. Materials and Methods
4.1. Tumor Samples
4.2. DNA/RNA Isolation
4.3. Design of Targeted Cancer-Related Gene Enrichment Panel
4.4. DNA and RNA Sequencing
4.5. Data Analysis
4.5.1. Sequence Alignment Pipeline
4.5.2. Somatic Mutation Calling
4.5.3. Somatic Indel Calling
4.5.4. Copy Number Variation (CNV) Calling
4.5.5. Tumor Purity
4.5.6. RNA Expression and Functional Analyses
4.5.7. Translocations
4.6. Immunohistochemical Staining
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Kozak, K.R.; Mahadevan, A.; Moody, J.S. Adult gliosarcoma: Epidemiology, natural history, and factors associated with outcome. Neuro-Oncology 2009, 11, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.; Alexe, D.-M.; Boeris, D.; Pereira, E. Primary gliosarcoma: Epidemiology, clinical presentation, management and survival. J. Neurosurg. Sci. 2017. [Google Scholar] [CrossRef]
- Beaumont, T.L.; Kupsky, W.J.; Barger, G.R.; Sloan, A.E. Gliosarcoma with multiple extracranial metastases: Case report and review of the literature. J. Neuro-Oncol. 2007, 83, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.R.; Wu, C.-C.; Saadatmand, H.J.; Isaacson, S.R.; Cheng, S.K.; Sisti, M.B.; Bruce, J.N.; Sheth, S.A.; Lassman, A.B.; Iwamoto, F.M.; et al. Clinical and molecular characteristics of gliosarcoma and modern prognostic significance relative to conventional glioblastoma. J. Neurooncol. 2018, 137, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Ortega, A.; Nuño, M.; Walia, S.; Mukherjee, D.; Black, K.L.; Patil, C.G. Treatment and survival of patients harboring histological variants of glioblastoma. J. Clin. Neurosci. 2014, 21, 1709–1713. [Google Scholar] [CrossRef] [PubMed]
- Cachia, D.; Kamiya-Matsuoka, C.; Mandel, J.J.; Olar, A.; Cykowski, M.D.; Armstrong, T.S.; Fuller, G.N.; Gilbert, M.R.; De Groot, J.F. Primary and secondary gliosarcomas: Clinical, molecular and survival characteristics. J. Neurooncol. 2015, 125, 401–410. [Google Scholar] [CrossRef] [PubMed]
- Castelli, J.; Feuvret, L.; Haoming, Q.C.; Biau, J.; Jouglar, E.; Berger, A.; Truc, G.; Gutierrez, F.L.; Morandi, X.; Le Reste, P.J.; et al. Prognostic and therapeutic factors of gliosarcoma from a multi-institutional series. J. Neurooncol. 2016, 129, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Cho, Y.H.; Kang, S.Y.; Yoon, N.; Sung, C.O.; Suh, Y.-L. BRAF V600E mutations are frequent in dysembryoplastic neuroepithelial tumors and subependymal giant cell astrocytomas. J. Surg. Oncol. 2015, 111, 359–364. [Google Scholar] [CrossRef] [PubMed]
- Meis, J.M.; Ho, K.L.; Nelson, J.S. Gliosarcoma: A histologic and immunohistochemical reaffirmation. Mod. Pathol. 1990, 3, 19–24. [Google Scholar] [PubMed]
- Han, S.J.; Yang, I.; Tihan, T.; Prados, M.D.; Parsa, A.T. Primary gliosarcoma: Key clinical and pathologic distinctions from glioblastoma with implications as a unique oncologic entity. J. Neurooncol. 2010, 96, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.-Y.; Park, C.; Na, D.; Han, J.Y.; Lee, J.; Park, O.-K.; Zhang, C.; Sung, C.O.; Moon, H.E.; Kim, Y.; et al. High prevalence of TP53 mutations is associated with poor survival and an EMT signature in gliosarcoma patients. Exp. Mol. Med. 2017, 49, e317. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Kang, S.Y.; Suh, Y.-L.; Jeong, J.Y.; Lee, J.-I.; Nam, D.-H. Clinicopathologic and genomic features of gliosarcomas. J. Neurooncol. 2012, 107, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.E.; Ohta, T.; Nonoguchi, N.; Satomi, K.; Capper, D.; Pierscianek, D.; Sure, U.; Vital, A.; Paulus, W.; Mittelbronn, M.; et al. Genetic Alterations in Gliosarcoma and Giant Cell Glioblastoma. Brain Pathol. 2016, 26, 517–522. [Google Scholar] [CrossRef] [PubMed]
- Reis, R.M.; Könü-Lebleblicioglu, D.; Lopes, J.M.; Kleihues, P.; Ohgaki, H. Genetic Profile of Gliosarcomas. Am. J. Pathol. 2000, 156, 425–432. [Google Scholar] [CrossRef]
- Biernat, W.; Aguzzi, A.; Sure, U.; Grant, J.W.; Kleihues, P.; Hegi, M.E. Identical mutations of the p53 tumor suppressor gene in the gliomatous and the sarcomatous components of gliosarcomas suggest a common origin from glial cells. J. Neuropathol. Exp. Neurol. 1995, 54, 651–656. [Google Scholar] [CrossRef] [PubMed]
- Boerman, R.H.; Anderl, K.; Herath, J.; Borell, T.; Johnson, N.; Schaeffer-Klein, J.; Kirchhof, A.; Raap, A.K.; Scheithauer, B.W.; Jenkins, R.B. The glial and mesenchymal elements of gliosarcomas share similar genetic alterations. J. Neuropathol. Exp. Neurol. 1996, 55, 973–981. [Google Scholar] [CrossRef] [PubMed]
- Actor, B.; Cobbers, J.M.J.L.; Büschges, R.; Wolter, M.; Knobbe, C.B.; Lichter, P.; Reifenberger, G.; Weber, R.G. Comprehensive analysis of genomic alterations in gliosarcoma and its two tissue components. Genes Chromosomes Cancer 2002, 34, 416–427. [Google Scholar] [CrossRef] [PubMed]
- Hyman, D.M.; Puzanov, I.; Subbiah, V.; Faris, J.E.; Chau, I.; Blay, J.-Y.; Wolf, J.; Raje, N.S.; Diamond, E.L.; Hollebecque, A.; et al. Vemurafenib in Multiple Nonmelanoma Cancers with BRAF V600 Mutations. N. Engl. J. Med. 2015, 373, 726–736. [Google Scholar] [CrossRef] [PubMed]
- Brennan, C.W.; Verhaak, R.G.W.; McKenna, A.; Campos, B.; Noushmehr, H.; Salama, S.R.; Zheng, S.; Chakravarty, D.; Sanborn, J.Z.; Berman, S.H.; et al. The somatic genomic landscape of glioblastoma. Cell 2013, 155, 462–477. [Google Scholar] [CrossRef] [PubMed]
- Thiel, G.; Marczinek, K.; Neumann, R.; Witkowski, R.; Marchuk, D.A.; Nurnberg, P. Somatic mutations in the neurofibromatosis 1 gene in gliomas and primitive neuroectodermal tumours. Anticancer Res. 1995, 15, 2495–2499. [Google Scholar] [PubMed]
- Fahsold, R.; Hoffmeyer, S.; Mischung, C.; Gille, C.; Ehlers, C.; Kücükceylan, N.; Abdel-Nour, M.; Gewies, A.; Peters, H.; Kaufmann, D.; et al. Minor lesion mutational spectrum of the entire NF1 gene does not explain its high mutability but points to a functional domain upstream of the GAP-related domain. Am. J. Hum. Genet. 2000, 66, 790–818. [Google Scholar] [CrossRef] [PubMed]
- Upadhyaya, M.; Osborn, M.J.; Maynard, J.; Kim, M.R.; Tamanoi, F.; Cooper, D.N. Mutational and functional analysis of the neurofibromatosis type 1 (NF1) gene. Hum. Genet. 1997, 99, 88–92. [Google Scholar] [CrossRef] [PubMed]
- Akino, T.; Han, X.; Nakayama, H.; McNeish, B.; Zurakowski, D.; Mammoto, A.; Klagsbrun, M.; Smith, E. Netrin-1 promotes medulloblastoma cell invasiveness and angiogenesis, and demonstrates elevated expression in tumor tissue and urine of patients with pediatric medulloblastoma. Cancer Res. 2014, 74, 3716–3726. [Google Scholar] [CrossRef] [PubMed]
- Barr, F.; Lambright, D.G. Rab GEFs and GAPs. Curr. Opin. Cell Biol. 2010, 22, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Lu, A.; Chen, X.; Wei, L.; Ding, J. RABEX-5 is upregulated and plays an oncogenic role in gastric cancer development by activating the VEGF signaling pathway. PLoS ONE 2014, 9, e113891. [Google Scholar] [CrossRef] [PubMed]
- Stricker, T.P.; Brown, C.D.; Bandlamudi, C.; McNerney, M.; Kittler, R.; Montoya, V.; Peterson, A.; Grossman, R.; White, K.P. Robust stratification of breast cancer subtypes using differential patterns of transcript isoform expression. PLoS Genet. 2017, 13, e1006589. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Hua, L.; Yan, D.; Zhao, F.; Liu, J.; Zhou, H.; Liu, J.; Wu, M.; Zhang, C.; Chen, Y.; et al. Overexpression of PCBP2 contributes to poor prognosis and enhanced cell growth in human hepatocellular carcinoma. Oncol. Rep. 2016, 36, 3456–3464. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.R.; DeClue, J.E.; Felzmann, S.; Vass, W.C.; Xu, G.; White, R.; Lowy, D.R. Neurofibromin can inhibit Ras-dependent growth by a mechanism independent of its GTPase-accelerating function. Mol. Cell. Biol. 1994, 14, 641–645. [Google Scholar] [CrossRef] [PubMed]
- Maehama, T.; Dixon, J.E. The Tumor Suppressor, PTEN/MMAC1, Dephosphorylates the Lipid Second Messenger, Phosphatidylinositol 3,4,5-Trisphosphate. J. Biol. Chem. 1998, 273, 13375–13378. [Google Scholar] [CrossRef] [PubMed]
- Samuels, Y.; Wang, Z.; Bardelli, A.; Silliman, N.; Ptak, J.; Szabo, S.; Yan, H.; Gazdar, A.; Powell, S.M.; Riggins, G.J.; et al. High frequency of mutations of the PIK3CA gene in human cancers. Science 2004, 304, 554. [Google Scholar] [CrossRef] [PubMed]
- Schwetye, K.E.; Joseph, N.M.; Al-Kateb, H.; Rich, K.M.; Schmidt, R.E.; Perry, A.; Gutmann, D.H.; Dahiya, S. Gliosarcomas lack BRAF(V600E) mutation, but a subset exhibit β-catenin nuclear localization. Neuropathology 2016, 36, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Sun, J.; Li, Z.; Chen, L.; Fu, Y.; Zhao, L.; Liu, L.; Wei, Y.; Teng, L.; Lu, D. Gliosarcomas with the BRAF V600E mutation: A report of two cases and review of the literature. J. Clin. Pathol. 2017, 70, 1079–1083. [Google Scholar] [CrossRef] [PubMed]
- Schindler, G.; Capper, D.; Meyer, J.; Janzarik, W.; Omran, H.; Herold-Mende, C.; Schmieder, K.; Wesseling, P.; Mawrin, C.; Hasselblatt, M.; et al. Analysis of BRAF V600E mutation in 1,320 nervous system tumors reveals high mutation frequencies in pleomorphic xanthoastrocytoma, ganglioglioma and extra-cerebellar pilocytic astrocytoma. Acta Neuropathol. 2011, 121, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Chu, W.; Song, X.; Yang, X.; Ma, L.; Zhu, J.; He, M.; Wang, Z.; Wu, Y. Neuropilin-1 promotes epithelial-to-mesenchymal transition by stimulating nuclear factor-kappa B and is associated with poor prognosis in human oral squamous cell carcinoma. PLoS ONE 2014, 9, e101931. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.Y.-J.; Kuay, K.T.; Tan, T.Z.; Asad, M.; Tang, H.M.; Ng, A.H.C.; Ye, J.; Chung, V.Y.; Thiery, J.P. Functional relevance of a six mesenchymal gene signature in epithelial-mesenchymal transition (EMT) reversal by the triple angiokinase inhibitor, nintedanib (BIBF1120). Oncotarget 2015, 6, 22098–22113. [Google Scholar] [CrossRef] [PubMed]
- Yeh, C.-M.; Chen, P.-C.; Hsieh, H.-Y.; Jou, Y.-C.; Lin, C.-T.; Tsai, M.-H.; Huang, W.-Y.; Wang, Y.-T.; Lin, R.-I.; Ch, S.-S.; et al. Methylomics analysis identifies ZNF671 as an epigenetically repressed novel tumor suppressor and a potential non-invasive biomarker for the detection of urothelial carcinoma. Oncotarget 2015, 6, 29555–29572. [Google Scholar] [CrossRef] [PubMed]
- Sripathy, S.P.; Stevens, J.; Schultz, D.C. The KAP1 corepressor functions to coordinate the assembly of de novo HP1-demarcated microenvironments of heterochromatin required for KRAB zinc finger protein-mediated transcriptional repression. Mol. Cell. Biol. 2006, 26, 8623–8638. [Google Scholar] [CrossRef] [PubMed]
- Urrutia, R. KRAB-containing zinc-finger repressor proteins. Genome Biol. 2003, 4, 231. [Google Scholar] [CrossRef] [PubMed]
- Dawson, M.A.; Kouzarides, T. Cancer epigenetics: From mechanism to therapy. Cell 2012, 150, 12–27. [Google Scholar] [CrossRef] [PubMed]
- Maleszewska, M.; Kaminska, B. Is glioblastoma an epigenetic malignancy? Cancers 2013, 5, 1120–1139. [Google Scholar] [CrossRef] [PubMed]
- Tessarz, P.; Kouzarides, T. Histone core modifications regulating nucleosome structure and dynamics. Nat. Rev. Mol. Cell Biol. 2014, 15, 703–708. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatic 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatic 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [PubMed]
- Koboldt, D.C.; Zhang, Q.; Larson, D.E.; Shen, D.; McLellan, M.D.; Lin, L.; Miller, C.A.; Mardis, E.R.; Ding, L.; Wilson, R.K. VarScan 2: Somatic mutation and copy number alteration discovery in cancer by exome sequencing. Genome Res. 2012, 22, 568–576. [Google Scholar] [CrossRef] [PubMed]
- Yates, A.; Akanni, W.; Amode, M.R.; Barrell, D.; Billis, K.; Carvalho-Silva, D.; Cummins, C.; Clapham, P.; Fitzgerald, S.; Gil, L.; et al. Ensembl 2016. Nucleic Acids Res. 2016, 44, D710–D716. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Henikoff, S.; Ng, P.C. Predicting the effects of coding non-synonymous variants on protein function using the SIFT algorithm. Nat. Protoc. 2009, 4, 1073–1081. [Google Scholar] [CrossRef] [PubMed]
- Adzhubei, I.A.; Schmidt, S.; Peshkin, L.; Ramensky, V.E.; Gerasimova, A.; Bork, P.; Kondrashov, A.S.; Sunyaev, S.R. A method and server for predicting damaging missense mutations. Nat. Methods 2010, 7, 248–249. [Google Scholar] [CrossRef] [PubMed]
- Olshen, A.B.; Bengtsson, H.; Neuvial, P.; Spellman, P.T.; Olshen, R.A.; Seshan, V.E. Parent-specific copy number in paired tumor-normal studies using circular binary segmentation. Bioinformatic 2011, 27, 2038–2046. [Google Scholar] [CrossRef] [PubMed]
- Skidmore, Z.L.; Wagner, A.H.; Lesurf, R.; Campbell, K.M.; Kunisaki, J.; Griffith, O.L.; Griffith, M. GenVisR: Genomic Visualizations in R. Bioinformatic 2016, 32, 3012–3014. [Google Scholar] [CrossRef] [PubMed]
- Bao, L.; Pu, M.; Messer, K. AbsCN-seq: A statistical method to estimate tumor purity, ploidy and absolute copy numbers from next-generation sequencing data. Bioinformatic 2014, 30, 1056–1063. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. TopHat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013, 14, R36. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed]
- Reimand, J.; Arak, T.; Vilo, J. g: Profiler—A web server for functional interpretation of gene lists (2011 update). Nucleic Acids Res. 2011, 39, W307–W315. [Google Scholar] [CrossRef] [PubMed]
- Schwint, O.A.; Labraga, M.; Cervino, C.O.; Haffar, M.; Sequeiros, P.H.; Marcos, H.J.A. A modification of the staining technique of reticular fibres for image analysis of the cardiac collagen network. Cardiovasc. Pathol. 2004, 13, 213–220. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
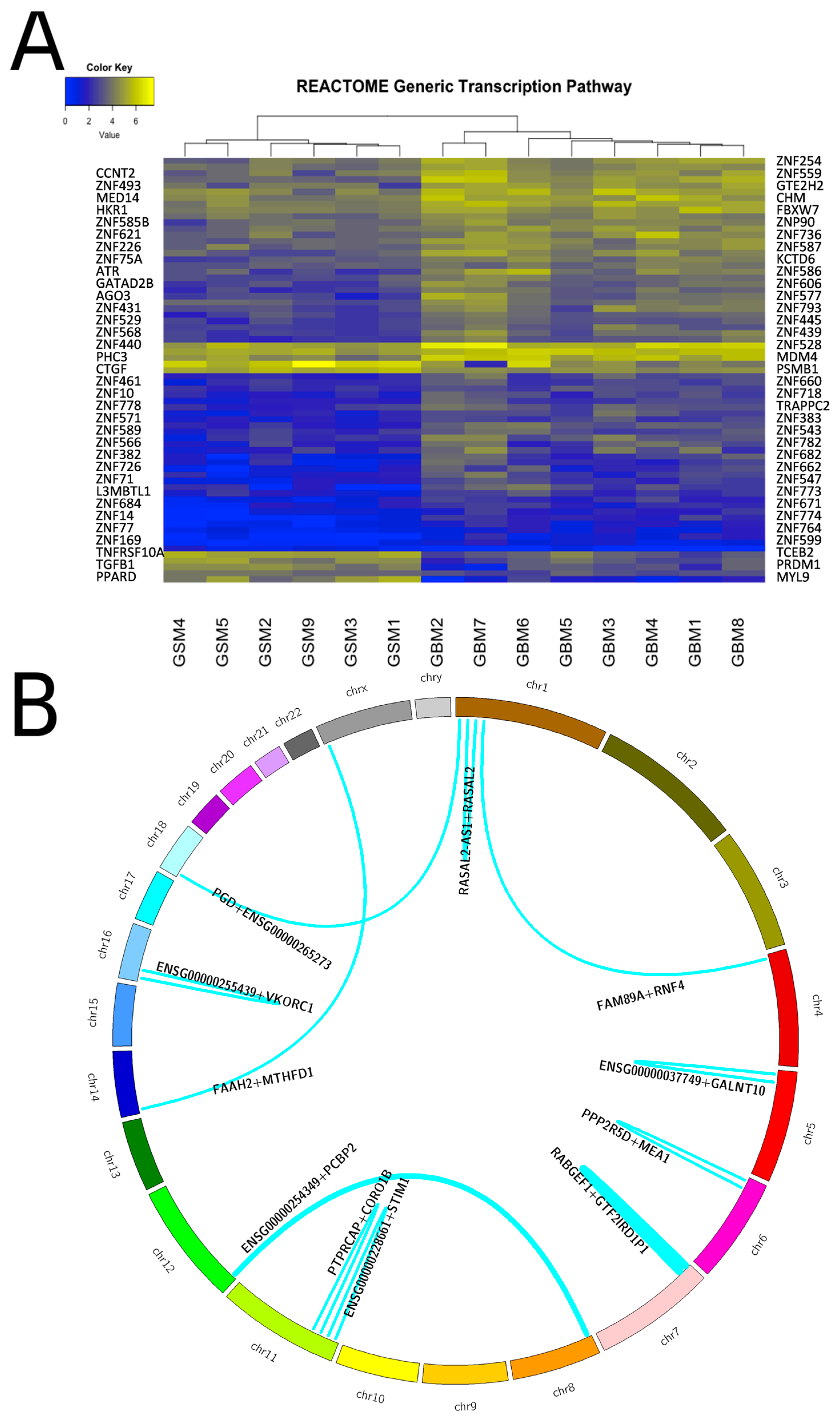{kind=link}
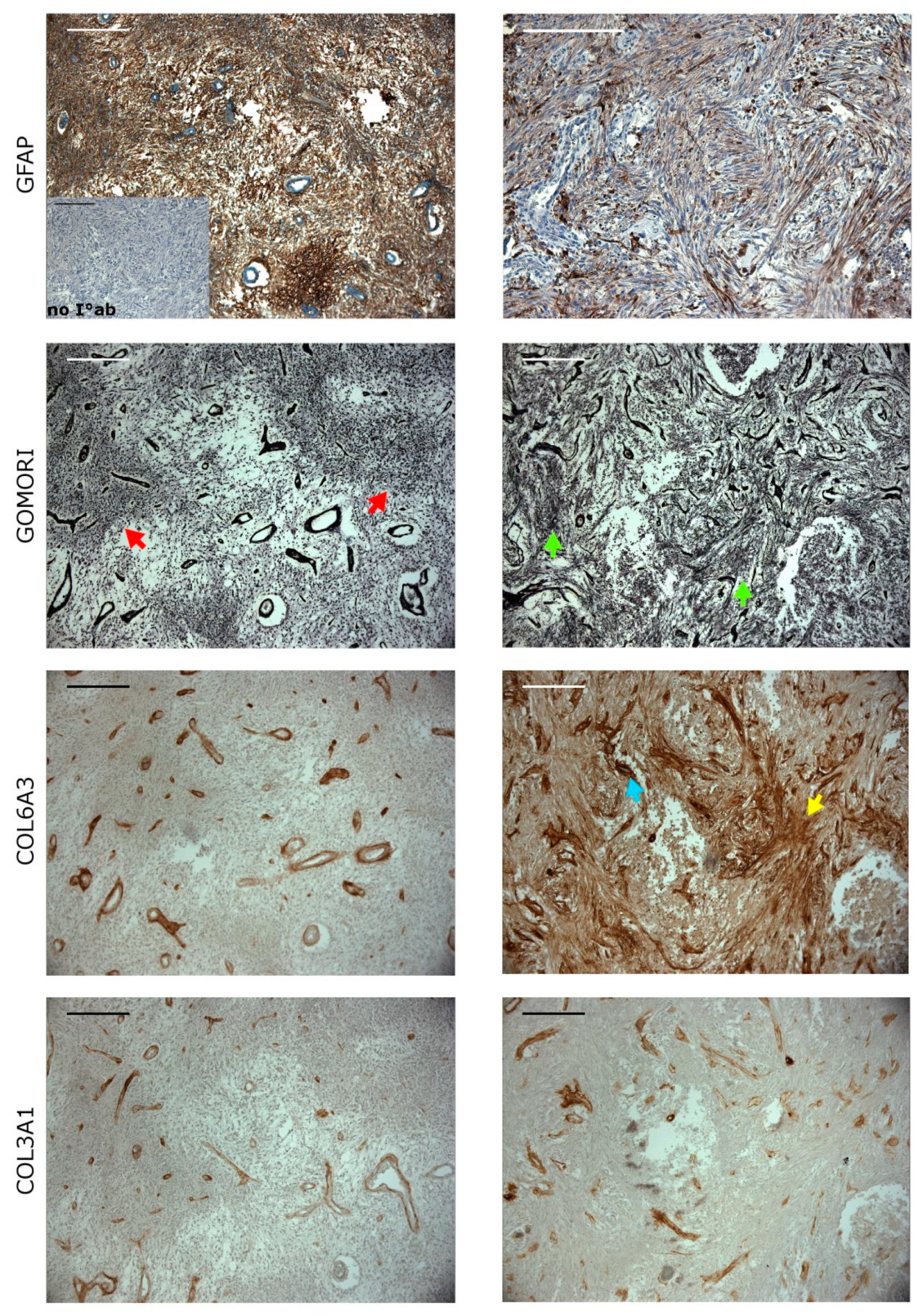{kind=link}
| Gene_Name | Chrom | Position | Reference | Variant | VAF | p Value | Sample | Sanger |
|---|---|---|---|---|---|---|---|---|
| HDAC11 | chr3 | 13504480 | C | T | 39.13% | 7.15 × 10−7 | GSM1 | validated |
| PAX6 | chr11 | 31802832 | G | A | 38.66% | 5.65 × 10−4 | GSM1 | not tested |
| PIK3CA | chr3 | 179198940 | G | A | 37.56% | 8.01 × 10−9 | GSM1 | validated |
| PTCH1 | chr9 | 95447237 | C | T | 36.81% | 6.74 × 10−4 | GSM1 | not tested |
| BRAF | chr7 | 140753336 | A | T | 26.68% | 9.73 × 10−5 | GSM1 | validated |
| PTEN | chr10 | 87894103 | T | C | 21.70% | 6.02 × 10−4 | GSM1 | not validated |
| PTEN | chr10 | 87925552 | C | A | 55.32% | 6.09 × 10−6 | GSM2 | validated |
| USP6 | chr17 | 5170636 | G | C | 33.99% | 1.19 × 10−4 | GSM2 | not tested |
| CDKN2C | chr1 | 50970430 | T | G | 60.38% | 1.02 × 10−14 | GSM3 | validated |
| TP53 | chr17 | 7673821 | G | A | 55.38% | 1.53 × 10−5 | GSM3 | not tested |
| MSH6 | chr2 | 47798725 | C | T | 53.85% | 1.04 × 10−12 | GSM3 | validated |
| RB1 | chr13 | 48342630 | G | A | 25.27% | 1.65 × 10−5 | GSM5 | validated |
| WHSC1 | chr4 | 1959574 | C | T | 22.05% | 1.67 × 10−3 | GSM5 | not tested |
| NKX2-1 | chr14 | 36519297 | C | A | 50% | 1.25 × 10−3 | GSM6 | not tested |
| TP53 | chr17 | 7674947 | A | G | 42.86% | 2.45 × 10−4 | GSM6 | not tested |
| OPTN | chr10 | 13127868 | G | T | 27.27% | 1.02 × 10−2 | GSM6 | not tested |
| PTEN | chr10 | 87960910 | T | C | 30.25% | 2.38 × 10−2 | GSM9 | validated |
| BRAF * | chr7 | 140753336 | A | T | 6.00% | ND | GSM10 | not tested |
| Gene_Name | Chrom | Position | Ref | Variant | VAF | p Value | Sample | Sanger |
|---|---|---|---|---|---|---|---|---|
| NF1 | chr17 | 31325895 | G | -C | 55.04% | 6.9 × 10−15 | GSM3 | validated |
| PIK3R1 | chr5 | 68280682 | A | -CT | 51.72% | 8.2 × 10−10 | GSM3 | not tested |
| PTEN | chr10 | 87961041 | G | -TACT | 50% | 2.4 × 10−11 | GSM3 | validated |
| PTEN | chr10 | 87961041 | G | -TACT | 19.23% | 1.1 × 10−4 | GSM4 | validated |
| NF1 * | chr17 | 31235638 | C | -TGTT | 14% | 8.4 × 10−3 | GSM4 | validated |
| RB1 | chr13 | 48456342 | T | -A | 34.88% | 3.5 × 10−5 | GSM6 | not tested |
| RB1 | chr13 | 48307352 | C | -AG | 17.28% | 2.0 × 10−2 | GSM7 | not tested |
| PIK3R1 | chr5 | 68293757 | C | -ATGAAT | 29.41% | 1.6 × 10−3 | GSM8 | not tested |
| NF1 | chr17 | 31159044 | A | -TCTC | 18.01% | 1.6 × 10−3 | GSM9 | validated |
| p Value | Term.id | Domain | Term.Name |
|---|---|---|---|
| 2.69 × 10−10 | REAC:212436 | re | Generic Transcription Pathway |
| 1.48 × 10−7 | GO:0003676 | MF | nucleic acid binding |
| 4.31 × 10−7 | GO:0046872 | MF | metal ion binding |
| 2.54 × 10−6 | GO:0043169 | MF | cation binding |
| 0.000116 | GO:0003677 | MF | DNA binding |
| 0.000167 | REAC:74160 | re | Gene Expression |
| 0.000202 | GO:0044260 | BP | cellular macromolecule metabolic process |
| 0.000492 | GO:0090304 | BP | nucleic acid metabolic process |
| 0.000769 | GO:0016070 | BP | RNA metabolic process |
| 0.00198 | GO:0043170 | BP | macromolecule metabolic process |
| 0.00417 | GO:0005925 | CC | focal adhesion |
| 0.0058 | GO:0005924 | CC | cell-substrate adherens junction |
| 0.00701 | GO:0006139 | BP | nucleobase-containing compound metabolic process |
| 0.00751 | GO:0030055 | CC | cell-substrate junction |
| 0.00796 | GO:0005634 | CC | nucleus |
| 0.0227 | GO:0043227 | CC | membrane-bounded organelle |
| 0.0234 | CORUM:2853 | co | ITGA5-ITGB1-CAL4A3 complex |
| 0.0267 | GO:1901363 | MF | heterocyclic compound binding |
| 0.0339 | GO:0009059 | BP | macromolecule biosynthetic process |
| 0.0346 | GO:0010467 | BP | gene expression |
| 0.0366 | GO:0043231 | CC | intracellular membrane-bounded organelle |
| 0.0391 | GO:0051252 | BP | regulation of RNA metabolic process |
| 0.0463 | CORUM:3104 | co | ITGB1-NRP1 complex |
| 0.05 | CORUM:5658 | co | Nrp1-PlexinD1 complex |
| 0.05 | HP:0002693 | hp | Abnormality of the skull base |
| 0.05 | KEGG:00511 | ke | Other glycan degradation |
| Sample | gene1 | Chr_gene1 | Position_gene1 | gene2 | Chr_gene2 | Position_gene2 |
|---|---|---|---|---|---|---|
| GSM1 | PTPRCAP | chr11 | 67204279 | CORO1B | chr11 | 67206320 |
| GSM1 | RABGEF1 * | chr7 | 66273872 | GTF2IRD1P1 | chr7 | 66275833 |
| GSM2 | MTHFD1 | chr14 | 64909104 | FAAH2 | chrX | 57419899 |
| GSM3 | ENSG00000037749 | chr5 | 153569748 | GALNT10 | chr5 | 153674375 |
| GSM3 | ENSG00000228661 | chr11 | 3876644 | STIM1 | chr11 | 3988780 |
| GSM3 | FAM89A | chr1 | 231157568 | RNF4 | chr4 | 2514808 |
| GSM3 | RABGEF1 * | chr7 | 66273872 | GTF2IRD1P1 | chr7 | 66275833 |
| GSM5 | ENSG00000254349 | chr8 | 75515899 | PCBP2 | chr12 | 53858635 |
| GSM5 | PGD | chr1 | 10464318 | ENSG00000265273 | chr18 | 29542460 |
| GSM5 | RASAL2-AS1 | chr1 | 178063003 | RASAL2 | chr1 | 178063501 |
| GSM9 | ENSG00000254349 | chr8 | 75515899 | PCBP2 | chr12 | 53858635 |
| GSM9 | ENSG00000255439 | chr16 | 31102095 | VKORC1 | chr16 | 31102662 |
| GSM9 | RABGEF1 * | chr7 | 66273872 | GTF2IRD1P1 | chr7 | 66275833 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wojtas, B.; Gielniewski, B.; Wojnicki, K.; Maleszewska, M.; Mondal, S.S.; Nauman, P.; Grajkowska, W.; Glass, R.; Schüller, U.; Herold-Mende, C.; et al. Gliosarcoma Is Driven by Alterations in PI3K/Akt, RAS/MAPK Pathways and Characterized by Collagen Gene Expression Signature. Cancers 2019, 11, 284. https://doi.org/10.3390/cancers11030284
Wojtas B, Gielniewski B, Wojnicki K, Maleszewska M, Mondal SS, Nauman P, Grajkowska W, Glass R, Schüller U, Herold-Mende C, et al. Gliosarcoma Is Driven by Alterations in PI3K/Akt, RAS/MAPK Pathways and Characterized by Collagen Gene Expression Signature. Cancers. 2019; 11(3):284. https://doi.org/10.3390/cancers11030284
Chicago/Turabian StyleWojtas, Bartosz, Bartlomiej Gielniewski, Kamil Wojnicki, Marta Maleszewska, Shamba S. Mondal, Pawel Nauman, Wieslawa Grajkowska, Rainer Glass, Ulrich Schüller, Christel Herold-Mende, and et al. 2019. "Gliosarcoma Is Driven by Alterations in PI3K/Akt, RAS/MAPK Pathways and Characterized by Collagen Gene Expression Signature" Cancers 11, no. 3: 284. https://doi.org/10.3390/cancers11030284
APA StyleWojtas, B., Gielniewski, B., Wojnicki, K., Maleszewska, M., Mondal, S. S., Nauman, P., Grajkowska, W., Glass, R., Schüller, U., Herold-Mende, C., & Kaminska, B. (2019). Gliosarcoma Is Driven by Alterations in PI3K/Akt, RAS/MAPK Pathways and Characterized by Collagen Gene Expression Signature. Cancers, 11(3), 284. https://doi.org/10.3390/cancers11030284