Role of Macrophage Migration Inhibitory Factor (MIF) in Melanoma


{kind=link}
Abstract
1. Introduction
1.1. Melanoma
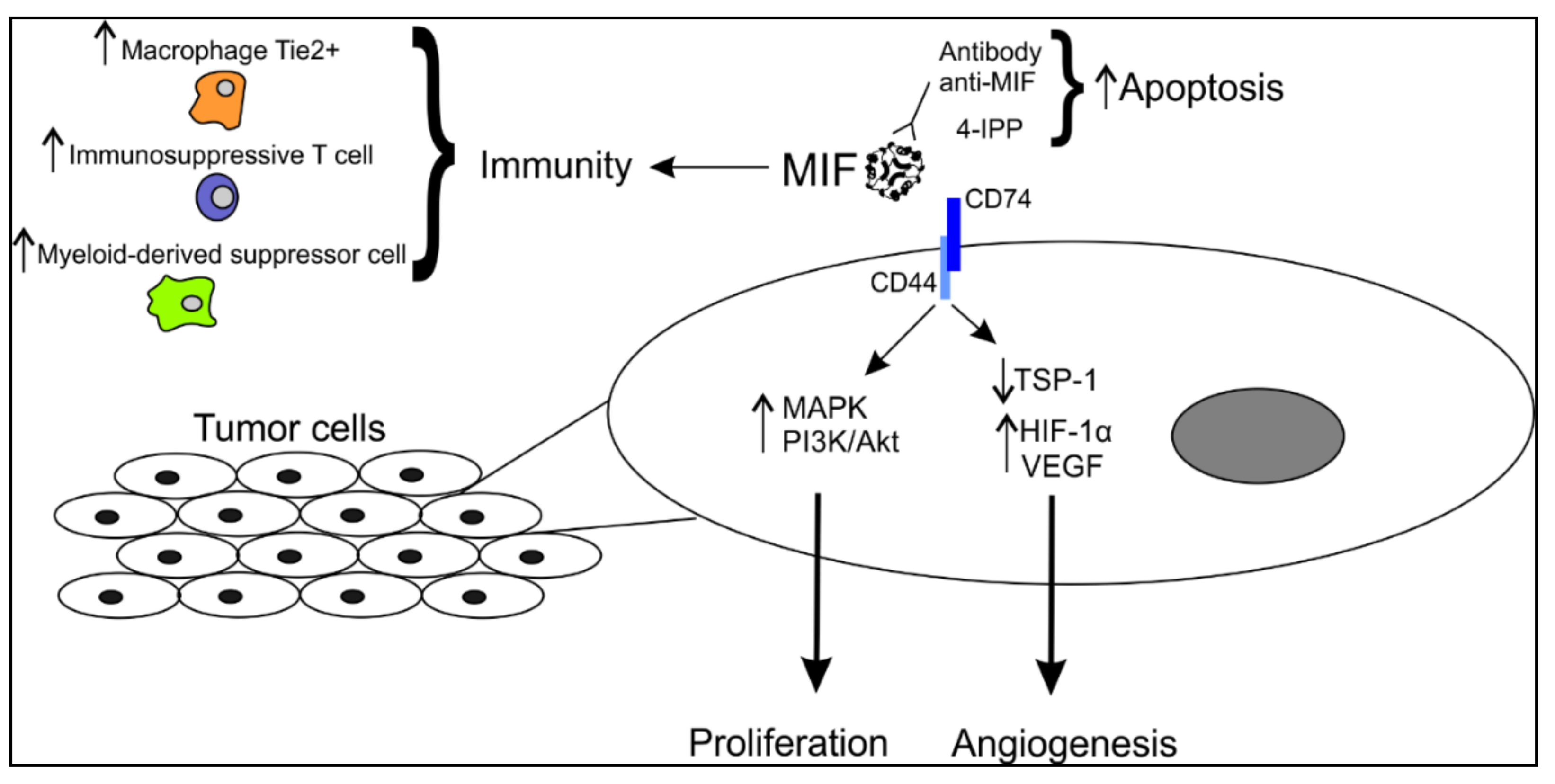
1.2. MIF and Cancer
1.3. Genomic Alteration of MIF in Melanoma
1.4. MIF mRNA and Protein Expression in Melanoma
1.5. MIF and Pigmentation
2. MIF Impact on Melanomagenesis
2.1. MIF and Melanoma Patient Outcome
2.2. Signaling Pathways
2.3. Angiogenesis
2.4. Immunity
2.5. Metastasis
3. Future Therapies against MIF in Melanoma
4. Conclusions
Funding
Conflicts of Interest
References
- Najem, A.; Krayem, M.; Perdrix, A.; Kerger, J.; Awada, A.; Journe, F.; Ghanem, G. New drug combination strategies in melanoma: Current status and future directions. Anticancer Res. 2017, 37, 5941–5953. [Google Scholar] [PubMed]
- Millet, A.; Martin, A.R.; Ronco, C.; Rocchi, S.; Benhida, R. Metastatic melanoma: Insights into the evolution of the treatments and future challenges. Med. Res. Rev. 2017, 37, 98–148. [Google Scholar] [CrossRef] [PubMed]
- Davey, R.J.; van der Westhuizen, A.; Bowden, N.A. Metastatic melanoma treatment: Combining old and new therapies. Crit. Rev. Oncol. Hematol. 2016, 98, 242–253. [Google Scholar] [CrossRef] [PubMed]
- Bloom, B.R.; Bennett, B. Mechanism of a reaction in vitro associated with delayed-type hypersensitivity. Science 1966, 153, 80–82. [Google Scholar] [CrossRef]
- Conroy, H.; Mawhinney, L.; Donnelly, S.C. Inflammation and cancer: Macrophage migration inhibitory factor (MIF)—The potential missing link. QJM Int. J. Med. 2010, 103, 831–836. [Google Scholar] [CrossRef]
- Santos, L.L.; Morand, E.F. Macrophage migration inhibitory factor: A key cytokine in RA, SLE and atherosclerosis. Clin. Chim. Acta Int. J. Clin. Chem. 2009, 399, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Zamora, Y.I.; Rodriguez-Sosa, M. The role of MIF in type 1 and type 2 diabetes mellitus. J. Diabetes Res. 2014, 2014, 804519. [Google Scholar] [CrossRef]
- Rendon, B.E.; Roger, T.; Teneng, I.; Zhao, M.; Al-Abed, Y.; Calandra, T.; Mitchell, R.A. Regulation of human lung adenocarcinoma cell migration and invasion by macrophage migration inhibitory factor. J. Biol. Chem. 2007, 282, 29910–29918. [Google Scholar] [CrossRef] [PubMed]
- He, X.-X.; Chen, K.; Yang, J.; Li, X.-Y.; Gan, H.-Y.; Liu, C.-Y.; Coleman, T.R.; Al-Abed, Y. Macrophage migration inhibitory factor promotes colorectal cancer. Mol. Med. 2009, 15, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Richard, V.; Kindt, N.; Decaestecker, C.; Gabius, H.-J.; Laurent, G.; Noël, J.-C.; Saussez, S. Involvement of macrophage migration inhibitory factor and its receptor (CD74) in human breast cancer. Oncol. Rep. 2014, 32, 523–529. [Google Scholar] [CrossRef] [PubMed]
- Meyer-Siegler, K.L.; Iczkowski, K.A.; Leng, L.; Bucala, R.; Vera, P.L. Inhibition of macrophage migration inhibitory factor or its receptor (CD74) attenuates growth and invasion of DU-145 prostate cancer cells. J. Immunol. 2006, 177, 8730–8739. [Google Scholar] [CrossRef] [PubMed]
- Kindt, N.; Preillon, J.; Kaltner, H.; Gabius, H.-J.; Chevalier, D.; Rodriguez, A.; Johnson, B.D.; Megalizzi, V.; Decaestecker, C.; Laurent, G.; et al. Macrophage migration inhibitory factor in head and neck squamous cell carcinoma: Clinical and experimental studies. J. Cancer Res. Clin. Oncol. 2013, 139, 727–737. [Google Scholar] [CrossRef] [PubMed]
- Kindt, N.; Lechien, J.R.; Nonclercq, D.; Laurent, G.; Saussez, S. Involvement of CD74 in head and neck squamous cell carcinomas. J. Cancer Res. Clin. Oncol. 2014, 140, 937–947. [Google Scholar] [CrossRef]
- Dessein, A.-F.; Stechly, L.; Jonckheere, N.; Dumont, P.; Monté, D.; Leteurtre, E.; Truant, S.; Pruvot, F.-R.; Figeac, M.; Hebbar, M.; et al. Autocrine induction of invasive and metastatic phenotypes by the MIF-CXCR4 axis in drug-resistant human colon cancer cells. Cancer Res. 2010, 70, 4644–4654. [Google Scholar] [CrossRef]
- Shimizu, T.; Abe, R.; Nakamura, H.; Ohkawara, A.; Suzuki, M.; Nishihira, J. High expression of macrophage migration inhibitory factor in human melanoma cells and its role in tumor cell growth and angiogenesis. Biochem. Biophys. Res. Commun. 1999, 264, 751–758. [Google Scholar] [CrossRef] [PubMed]
- Rumpler, G.; Becker, B.; Hafner, C.; McClelland, M.; Stolz, W.; Landthaler, M.; Schmitt, R.; Bosserhoff, A.; Vogt, T. Identification of differentially expressed genes in models of melanoma progression by cDNA array analysis: SPARC, MIF and a novel cathepsin protease characterize aggressive phenotypes. Exp. Dermatol. 2003, 12, 761–771. [Google Scholar] [CrossRef]
- Miracco, C.; De Nisi, M.C.; Arcuri, F.; Cosci, E.; Pacenti, L.; Toscano, M.; Lalinga, A.V.; Biagioli, M.; Rubegni, P.; Vatti, R.; et al. Macrophage migration inhibitory factor protein and mRNA expression in cutaneous melanocytic tumours. Int. J. Oncol. 2006, 28, 345–352. [Google Scholar] [CrossRef][Green Version]
- Yasasever, V.; Camlica, H.; Duranyildiz, D.; Oguz, H.; Tas, F.; Dalay, N. Macrophage migration inhibitory factor in cancer. Cancer Investig. 2007, 25, 715–719. [Google Scholar] [CrossRef] [PubMed]
- Schulz, R.; Marchenko, N.D.; Holembowski, L.; Fingerle-Rowson, G.; Pesic, M.; Zender, L.; Dobbelstein, M.; Moll, U.M. Inhibiting the HSP90 chaperone destabilizes macrophage migration inhibitory factor and thereby inhibits breast tumor progression. J. Exp. Med. 2012, 209, 275–289. [Google Scholar] [CrossRef] [PubMed]
- López, S.; Smith-Zubiaga, I.; García de Galdeano, A.; Boyano, M.D.; García, O.; Gardeazábal, J.; Martinez-Cadenas, C.; Izagirre, N.; de la Rúa, C.; Alonso, S.; et al. Comparison of the transcriptional profiles of melanocytes from dark and light skinned individuals under basal conditions and following ultraviolet-B irradiation. PLoS ONE 2015, 10, e0134911. [Google Scholar]
- Cournia, Z.; Leng, L.; Gandavadi, S.; Du, X.; Bucala, R.; Jorgensen, W.L. Discovery of human macrophage migration inhibitory factor (MIF)-CD74 antagonists via virtual screening. J. Med. Chem. 2009, 52, 416–424. [Google Scholar] [CrossRef]
- Guyonneau, L.; Murisier, F.; Rossier, A.; Moulin, A.; Beermann, F. Melanocytes and pigmentation are affected in dopachrome tautomerase knockout mice. Mol. Cell. Biol. 2004, 24, 3396–3403. [Google Scholar] [CrossRef]
- Oliveira, C.S.; de Bock, C.E.; Molloy, T.J.; Sadeqzadeh, E.; Geng, X.Y.; Hersey, P.; Zhang, X.; Thorne, R.F. Macrophage migration inhibitory factor engages PI3K/Akt signalling and is a prognostic factor in metastatic melanoma. BMC Cancer 2014, 14, 630. [Google Scholar] [CrossRef]
- Ekmekcioglu, S.; Davies, M.A.; Tanese, K.; Roszik, J.; Shin-Sim, M.; Bassett, R.L., Jr.; Milton, D.R.; Voidman, C.E.; Admitted, W.G.; Gerschenwald, J.E.; et al. Inflammatory marker testing identifies CD74 expression in melanoma tumor cells, and its expression associates with favorable survival for stage III melanoma. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2016, 22, 3016–3024. [Google Scholar] [CrossRef]
- Leng, L.; Metz, C.N.; Fang, Y.; Xu, J.; Donnelly, S.; Baugh, J.; Delohery, T.; Chen, Y.; Mitchell, R.A.; Bucala, R. MIF Signal transduction initiated by binding to CD74. J. Exp. Med. 2003, 197, 1467–1476. [Google Scholar] [CrossRef]
- Nguyen, M.T.; Lue, H.; Kleemann, R.; Thiele, M.; Tolle, G.; Finkelmeier, D.; Wagner, E.; Braun, A.; Bernhagen, J. The cytokine macrophage migration inhibitory factor reduces pro-oxidative stress-induced apoptosis. J. Immunol. 2003, 170, 3337–3347. [Google Scholar] [CrossRef]
- Amin, M.A.; Volpert, O.V.; Woods, J.M.; Kumar, P.; Harlow, L.A.; Koch, A.E. Migration inhibitory factor mediates angiogenesis via mitogen-activated protein kinase and phosphatidylinositol kinase. Circ. Res. 2003, 93, 321–329. [Google Scholar] [CrossRef]
- Culp, W.D.; Tsagozis, P.; Burgio, M.; Russell, P.; Pisa, P.; Garland, D. Interference of macrophage migration inhibitory factor expression in a mouse melanoma inhibits tumor establishment by up-regulating thrombospondin-1. Mol. Cancer Res. 2007, 5, 1225–1231. [Google Scholar] [CrossRef]
- Good, D.J.; Polverini, P.J.; Rastinejad, F.; Beau, M.M.L.; Lemons, R.S.; Frazier, W.A.; Bouck, N.P. A tumor suppressor-dependent inhibitor of angiogenesis is immunologically and functionally indistinguishable from a fragment of thrombospondin. Proc. Natl. Acad. Sci. USA 1990, 87, 6624–6628. [Google Scholar] [CrossRef]
- Girard, E.; Strathdee, C.; Trueblood, E.; Quéva, C. Macrophage migration inhibitory factor produced by the tumour stroma but not by tumour cells regulates angiogenesis in the B16-F10 melanoma model. Br. J. Cancer 2012, 107, 1498–1505. [Google Scholar] [CrossRef]
- Rendon, B.E.; Willer, S.S.; Zundel, W.; Mitchell, R.A. Mechanisms of macrophage migration inhibitory factor (MIF)-dependent tumor microenvironmental adaptation. Exp. Mol. Pathol. 2009, 86, 180–185. [Google Scholar] [CrossRef]
- Yaddanapudi, K.; Putty, K.; Rendon, B.E.; Lamont, G.J.; Faughn, J.D.; Satoskar, A.; Lasnik, A.; Eaton, J.W.; Mitchell, R.A. Control of tumor-associated macrophage alternative activation by macrophage migration inhibitory factor. J. Immunol. 2013, 190, 2984–2993. [Google Scholar] [CrossRef]
- Apte, R.S.; Mayhew, E.; Niederkorn, J.Y. Local inhibition of natural killer cell activity promotes the progressive growth of intraocular tumors. Investig. Ophthalmol. Vis. Sci. 1997, 38, 1277–1282. [Google Scholar] [CrossRef]
- Repp, A.C.; Mayhew, E.S.; Apte, S.; Niederkorn, J.Y. Human uveal melanoma cells produce macrophage migration-inhibitory factor to prevent lysis by NK cells. J. Immunol. 2000, 165, 710–715. [Google Scholar] [CrossRef]
- Schoenfeld, J.; Jinushi, M.; Nakazaki, Y.; Wiener, D.; Park, J.; Soiffer, R.; Neuberg, D.; Mihm, M.; Hodi, F.S.; Dranoff, G. Active immunotherapy induces antibody responses that target tumor angiogenesis. Cancer Res. 2010, 70, 10150–10160. [Google Scholar] [CrossRef]
- Yaddanapudi, K.; Rendon, B.E.; Lamont, G.; Kim, E.J.; Al Rayyan, N.; Richie, J.; Albeituni, S.; Waigel, S.; Wise, A.; Mitchell, R.A. MIF is necessary for late-stage melanoma patient MDSC immune suppression and differentiation. Cancer Immunol. Res. 2016, 4, 101–112. [Google Scholar] [CrossRef]
- Hoos, A.; Ibrahim, R.; Korman, A.; Abdallah, K.; Berman, D.; Shahabi, V.; Chin, K.; Canetta, R.; Humphrey, R. Development of ipilimumab: Contribution to a new paradigm for cancer immunotherapy. Semin. Oncol. 2010, 37, 533–546. [Google Scholar] [CrossRef]
- Chae, Y.K.; Arya, A.; Iams, W.; Cruz, M.R.; Chandra, S.; Choi, J.; Giles, F. Current landscape and future of dual anti-CTLA4 and PD-1/PD-L1 blockade immunotherapy in cancer; lessons learned from clinical trials with melanoma and non-small cell lung cancer (NSCLC). J. Immunother. Cancer 2018, 6, 39. [Google Scholar] [CrossRef]
- Mantovani, A.; Sica, A. Macrophages, innate immunity and cancer: Balance, tolerance, and diversity. Curr. Opin. Immunol. 2010, 22, 231–237. [Google Scholar] [CrossRef]
- Abe, R.; Peng, T.; Sailors, J.; Bucala, R.; Metz, C.N. Regulation of the CTL response by macrophage migration inhibitory factor. J. Immunol. 2001, 166, 747–753. [Google Scholar] [CrossRef]
- Tanese, K.; Hashimoto, Y.; Berkova, Z.; Wang, Y.; Samaniego, F.; Lee, J.E.; Ekmekcioglu, S.; Grimm, E.A. Cell surface CD74-MIF interactions drive melanoma survival in response to interferon-γ. J. Investig. Dermatol. 2015, 135, 2775–2784. [Google Scholar] [CrossRef]
- Clawson, G.A.; Kimchi, E.; Patrick, S.D.; Xin, P.; Harouaka, R.; Zheng, S.; Berg, A.; Schell, T.; Staveley-O’Carroll, K.F.; Neves, R.I.; et al. Circulating tumor cells in melanoma patients. PLoS ONE 2012, 7, e41052. [Google Scholar] [CrossRef] [PubMed]
- Clawson, G.A.; Matters, G.L.; Xin, P.; Imamura-Kawasawa, Y.; Du, Z.; Thiboutot, D.M.; Helm, K.F.; Neves, R.I.; Abraham, T. Macrophage-tumor cell fusions from peripheral blood of melanoma patients. PLoS ONE 2015, 10, e0134320. [Google Scholar] [CrossRef] [PubMed]
- Ioannou, K.; Cheng, K.F.; Crichlow, G.V.; Birmpilis, A.I.; Lolis, E.J.; Tsitsilonis, O.E.; Al-Abed, Y. ISO-66, a novel inhibitor of macrophage migration, shows efficacy in melanoma and colon cancer models. Int. J. Oncol. 2014, 45, 1457–1468. [Google Scholar] [PubMed]
- Hussain, F.; Freissmuth, M.; Völkel, D.; Thiele, M.; Douillard, P.; Antoine, G.; Thurner, P.; Ehrlich, H.; Black, H.-P.; Scheiflinger, F.; et al. Human anti macrophage migration inhibitory factor antibodies inhibit growth of human prostate cancer cells in vitro and in vivo. Mol. Cancer Ther. 2013, 12, 1223–1234. [Google Scholar] [CrossRef] [PubMed]
- Schulz, R.; Moll, U.M. Targeting the heat shock protein 90: A rational way to inhibit macrophage migration inhibitory factor function in cancer. Curr. Opin. Oncol. 2014, 26, 108–113. [Google Scholar] [CrossRef]
- XL888 + Vemurafenib + Cobimetinib for Unresectable BRAF Mutated Stage III/IV Melanoma—Full Text View—ClinicalTrials.gov. Available online: https://clinicaltrials.gov/ct2/show/NCT02721459 (accessed on 15 March 2019).
- Study of XL888 With Vemurafenib for Patients with Unresectable BRAF Mutated Stage III/IV Melanoma—No Study Results Posted—ClinicalTrials.gov. Available online: https://clinicaltrials.gov/ct2/show/results/NCT01657591 (accessed on 15 March 2019).
- Figueiredo, C.R.; Azevedo, R.A.; Mousdell, S.; Resende-Lara, P.T.; Ireland, L.; Santos, A.; Girola, N.; Cunha, R.L.O.R.; Schmid, M.C.; Polonelli, L.; et al. Blockade of MIF–CD74 signalling on macrophages and dendritic cells restores the antitumour immune response against metastatic melanoma. Front. Immunol. 2018, 9, 1132. [Google Scholar] [CrossRef] [PubMed]

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soumoy, L.; Kindt, N.; Ghanem, G.; Saussez, S.; Journe, F. Role of Macrophage Migration Inhibitory Factor (MIF) in Melanoma. Cancers 2019, 11, 529. https://doi.org/10.3390/cancers11040529
Soumoy L, Kindt N, Ghanem G, Saussez S, Journe F. Role of Macrophage Migration Inhibitory Factor (MIF) in Melanoma. Cancers. 2019; 11(4):529. https://doi.org/10.3390/cancers11040529
Chicago/Turabian StyleSoumoy, Laura, Nadège Kindt, Ghanem Ghanem, Sven Saussez, and Fabrice Journe. 2019. "Role of Macrophage Migration Inhibitory Factor (MIF) in Melanoma" Cancers 11, no. 4: 529. https://doi.org/10.3390/cancers11040529
APA StyleSoumoy, L., Kindt, N., Ghanem, G., Saussez, S., & Journe, F. (2019). Role of Macrophage Migration Inhibitory Factor (MIF) in Melanoma. Cancers, 11(4), 529. https://doi.org/10.3390/cancers11040529