Pheochromocytomas and Paragangliomas: New Developments with Regard to Classification, Genetics, and Cell of Origin


Abstract
:1. Introduction
2. Genetics
2.1. Cluster 1
2.1.1. VHL/EPAS1-Related
2.1.2. Tricarboxylic Acid TCA Cycle-Related
2.1.3. FH, MDH2, IDH1/IDH2
2.1.4. Recently Identified PCC/PGL Genes
2.2. Cluster 2
3. Histopathological Classification
3.1. Pheochromocytoma of the Adrenal Gland Scaled Score (PASS)
3.2. Grading of Adrenal Pheochromocytoma and Paraganglioma (GAPP)
4. Discussion/Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Lenders, J.W.M.; Duh, Q.; Eisenhofer, G.; Gimenez-Roqueplo, A.; Grebe, S.K.G.; Murad, M.H.; Naruse, M.; Pacak, K.; Young, W.F. Pheochromocytoma and Paraganglioma: An Endocrine Society Clinical Practice Guideline. J. Clin. Endocrinol. Metab. 2014, 99, 1915–1942. [Google Scholar] [CrossRef] [PubMed]
- Huber, K.; Kalcheim, C.; Unsicker, K. The Development of the Chromaffin Cell Lineage from the Neural Crest. Auton. Neurosci. 2009, 151, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Saito, D.; Takase, Y.; Murai, H.; Takahashi, Y. The Dorsal Aorta Initiates a Molecular Cascade that Instructs Sympatho-Adrenal Specification. Science 2012, 336, 1578–1581. [Google Scholar] [CrossRef] [PubMed]
- Chan, W.H.; Gonsalvez, D.G.; Young, H.M.; Southard-Smith, E.M.; Cane, K.N.; Anderson, C.R. Differences in CART Expression and Cell Cycle Behavior Discriminate Sympathetic Neuroblast from Chromaffin Cell Lineages in Mouse Sympathoadrenal Cells. Dev. Neurobiol. 2016, 76, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Ernsberger, U.; Esposito, L.; Partimo, S.; Huber, K.; Franke, A.; Bixby, J.L.; Kalcheim, C.; Unsicker, K. Expression of Neuronal Markers Suggests Heterogeneity of Chick Sympathoadrenal Cells Prior to Invasion of the Adrenal Anlagen. Cell Tissue Res. 2005, 319, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Furlan, A.; Dyachuk, V.; Kastriti, M.E.; Calvo-Enrique, L.; Abdo, H.; Hadjab, S.; Chontorotzea, T.; Akkuratova, N.; Usoskin, D.; Kamenev, D.; et al. Multipotent Peripheral Glial Cells Generate Neuroendocrine Cells of the Adrenal Medulla. Science 2017, 357. [Google Scholar] [CrossRef]
- Lumb, R.; Tata, M.; Xu, X.; Joyce, A.; Marchant, C.; Harvey, N.; Ruhrberg, C.; Schwarz, Q. Neuropilins Guide Preganglionic Sympathetic Axons and Chromaffin Cell Precursors to Establish the Adrenal Medulla. Development 2018, 145. [Google Scholar] [CrossRef]
- Kastriti, M.E.; Kameneva, P.; Kamenev, D.; Dyachuk, V.; Furlan, A.; Hampl, M.; Memic, F.; Marklund, U.; Lallemend, F.; Hadjab, S.; et al. Schwann Cell Precursors Generate the Majority of Chromaffin Cells in Zuckerkandl Organ and some Sympathetic Neurons in Paraganglia. Front. Mol. Neurosci. 2019, 12, 6. [Google Scholar] [CrossRef]
- Fishbein, L.; Nathanson, K.L. Pheochromocytoma and Paraganglioma: Understanding the Complexities of the Genetic Background. Cancer Genet. 2012, 205, 1–11. [Google Scholar] [CrossRef]
- Loyd, R.V.; Osamura, R.Y.; Kloppel, G.; Rosai, J. WHO Classification of Tumours of Endocrine System; International Agency for Research on Cancer (IARC): Lyon, France, 2017. [Google Scholar]
- Thompson, L.D. Pheochromocytoma of the Adrenal Gland Scaled Score (PASS) to Separate Benign from Malignant Neoplasms: A Clinicopathologic and Immunophenotypic Study of 100 Cases. Am. J. Surg. Pathol. 2002, 26, 551–566. [Google Scholar] [CrossRef]
- Kimura, N.; Takayanagi, R.; Takizawa, N.; Itagaki, E.; Katabami, T.; Kakoi, N.; Rakugi, H.; Ikeda, Y.; Tanabe, A.; Nigawara, T.; et al. Pathological Grading for Predicting Metastasis in Phaeochromocytoma and Paraganglioma. Endocr. Relat. Cancer 2014, 21, 405–414. [Google Scholar] [CrossRef]
- Dahia, P.L. Pheochromocytoma and Paraganglioma Pathogenesis: Learning from Genetic Heterogeneity. Nat. Rev. Cancer 2014, 14, 108–119. [Google Scholar] [CrossRef]
- Dahia, P.L.M. Pheochromocytomas and Paragangliomas, Genetically Diverse and Minimalist, all at Once! Cancer Cell 2017, 31, 159–161. [Google Scholar] [CrossRef]
- Zhikrivetskaya, S.O.; Snezhkina, A.V.; Zaretsky, A.R.; Alekseev, B.Y.; Pokrovsky, A.V.; Golovyuk, A.L.; Melnikova, N.V.; Stepanov, O.A.; Kalinin, D.V.; Moskalev, A.A.; et al. Molecular Markers of Paragangliomas/Pheochromocytomas. Oncotarget 2017, 8, 25756–25782. [Google Scholar] [CrossRef]
- Jochmanová, I.; Yang, C.; Zhuang, Z.; Pacak, K. Hypoxia-Inducible Factor Signaling in Pheochromocytoma: Turning the Rudder in the Right Direction. J. Natl. Cancer Inst. 2013, 105, 1270–1283. [Google Scholar] [CrossRef]
- Jochmanova, I.; Pacak, K. Genomic Landscape of Pheochromocytoma and Paraganglioma. Trends Cancer 2018, 4, 6–9. [Google Scholar] [CrossRef]
- Burnichon, N.; Buffet, A.; Gimenez-Roqueplo, A. Pheochromocytoma and Paraganglioma: Molecular Testing and Personalized Medicine. Curr. Opin. Oncol. 2016, 28, 5–10. [Google Scholar] [CrossRef]
- Kavinga Gunawardane, P.T.; Grossman, A. The Clinical Genetics of Phaeochromocytoma and Paraganglioma. Arch. Endocrinol. Metab. 2017, 61, 490–500. [Google Scholar] [CrossRef]
- Amorim-Pires, D.; Peixoto, J.; Lima, J. Hypoxia Pathway Mutations in Pheochromocytomas and Paragangliomas. Cytogenet. Genome Res. 2016, 150, 227–241. [Google Scholar] [CrossRef]
- Vicha, A.; Musil, Z.; Pacak, K. Genetics of Pheochromocytoma and Paraganglioma Syndromes: New Advances and Future Treatment Options. Curr. Opin. Endocrinol. Diabetes Obes. 2013, 20, 186–191. [Google Scholar] [CrossRef]
- Fishbein, L.; Leshchiner, I.; Walter, V.; Danilova, L.; Robertson, A.G.; Johnson, A.R.; Lichtenberg, T.M.; Murray, B.A.; Ghayee, H.K.; Else, T.; et al. Comprehensive Molecular Characterization of Pheochromocytoma and Paraganglioma. Cancer Cell 2017, 31, 181–193. [Google Scholar] [CrossRef]
- Crona, J.; Taïeb, D.; Pacak, K. New Perspectives on Pheochromocytoma and Paraganglioma: Toward a Molecular Classification. Endocr. Rev. 2017, 38, 489–515. [Google Scholar] [CrossRef]
- Kantorovich, V.; Pacak, K. New Insights on the Pathogenesis of Paraganglioma and Pheochromocytoma. F1000Res 2018, 7. [Google Scholar] [CrossRef]
- Alrezk, R.; Suarez, A.; Tena, I.; Pacak, K. Update of Pheochromocytoma Syndromes: Genetics, Biochemical Evaluation, and Imaging. Front. Endocrinol. 2018, 9, 515. [Google Scholar] [CrossRef]
- Comino-Méndez, I.; Tejera, Á.; .M.;Currás-Freixes, M.; Remacha, L.; Gonzalvo, P.; Tonda, R.; Letón, R.; Blasco, M.A.; Robledo, M.; Cascón, A. ATRX Driver Mutation in a Composite Malignant Pheochromocytoma. Cancer Genet. 2016, 209, 272–277. [Google Scholar]
- Jochmanova, I.; Pacak, K. Pheochromocytoma: The First Metabolic Endocrine Cancer. Clin. Cancer Res. 2016, 22, 5001–5011. [Google Scholar] [CrossRef] [Green Version]
- Maher, E.R.; Neumann, H.P.; Richard, S. Von Hippel-Lindau Disease: A Clinical and Scientific Review. Eur. J. Hum. Genet. 2011, 19, 617–623. [Google Scholar] [CrossRef]
- Lyssikatos, C.; Faucs, F.R.; Stratakis, C.A. Familial Endocrine Tumor Syndromes; Cambridge University Press: Cambridge, UK, 2016. [Google Scholar]
- Zhuang, Z.; Yang, C.; Lorenzo, F.; Merino, M.; Fojo, T.; Kebebew, E.; Popovic, V.; Stratakis, C.A.; Prchal, J.T.; Pacak, K. Somatic HIF2A Gain-of-Function Mutations in Paraganglioma with Polycythemia. N. Engl. J. Med. 2012, 367, 922–930. [Google Scholar] [CrossRef]
- Lorenzo, F.R.; Yang, C.; Ng Tang Fui, M.; Vankayalapati, H.; Zhuang, Z.; Huynh, T.; Grossmann, M.; Pacak, K.; Prchal, J.T. A Novel EPAS1/HIF2A Germline Mutation in a Congenital Polycythemia with Paraganglioma. J. Mol. Med. 2013, 91, 507–512. [Google Scholar] [CrossRef]
- Oudijk, L.; Gaal, J.; Koopman, K.; de Krijger, R.R. An Update on the Histology of Pheochromocytomas: How does it Relate to Genetics? Horm. Metab. Res. 2018. [Google Scholar] [CrossRef]
- Ladroue, C.; Carcenac, R.; Leporrier, M.; Gad, S.; Le Hello, C.; Galateau-Salle, F.; Feunteun, J.; Pouysségur, J.; Richard, S.; Gardie, B. PHD2 Mutation and Congenital Erythrocytosis with Paraganglioma. N. Engl. J. Med. 2008, 359, 2685–2692. [Google Scholar] [CrossRef]
- Yang, C.; Zhuang, Z.; Fliedner, S.M.J.; Shankavaram, U.; Sun, M.G.; Bullova, P.; Zhu, R.; Elkahloun, A.G.; Kourlas, P.J.; Merino, M.; et al. Germ-Line PHD1 and PHD2 Mutations Detected in Patients with Pheochromocytoma/Paraganglioma-Polycythemia. J. Mol. Med. 2015, 93, 93–104. [Google Scholar] [CrossRef]
- van Nederveen, F.H.; Korpershoek, E.; Lenders, J.W.M.; de Krijger, R.R.; Dinjens, W.N.M. Somatic SDHB Mutation in an Extraadrenal Pheochromocytoma. N. Engl. J. Med. 2007, 357, 306–308. [Google Scholar] [CrossRef]
- Turchini, J.; Cheung, V.K.Y.; Tischler, A.S.; De Krijger, R.R.; Gill, A.J. Pathology and Genetics of Phaeochromocytoma and Paraganglioma. Histopathology 2018, 72, 97–105. [Google Scholar] [CrossRef]
- Gill, A.J. Succinate Dehydrogenase (SDH) and Mitochondrial Driven Neoplasia. Pathology 2012, 44, 285–292. [Google Scholar] [CrossRef]
- Hoekstra, A.S.; Hensen, E.F.; Jordanova, E.S.; Korpershoek, E.; van der Horst-Schrivers, A.N.; Cornelisse, C.; Corssmit, E.P.M.; Hes, F.J.; Jansen, J.C.; Kunst, H.P.M.; et al. Loss of Maternal Chromosome 11 is a Signature Event in SDHAF2, SDHD, and VHL-Related Paragangliomas, but Less Significant in SDHB-Related Paragangliomas. Oncotarget 2017, 8, 14525–14536. [Google Scholar] [CrossRef]
- Hensen, E.F.; Jordanova, E.S.; van Minderhout, I.J.H.M.; Hogendoorn, P.C.W.; Taschner, P.E.M.; van der Mey Andel, G.L.; Devilee, P.; Cornelisse, C.J. Somatic Loss of Maternal Chromosome 11 Causes Parent-of-Origin-Dependent Inheritance in SDHD-Linked Paraganglioma and Phaeochromocytoma Families. Oncogene 2004, 23, 4076–4083. [Google Scholar] [CrossRef]
- van Nederveen, F.H.; Gaal, J.; Favier, J.; Korpershoek, E.; Oldenburg, R.A.; de Bruyn, E.M.; Sleddens, H.F.; Derkx, P.; Riviere, J.; Dannenberg, H.; et al. An Immunohistochemical Procedure to Detect Patients with Paraganglioma and Phaeochromocytoma with Germline SDHB, SDHC, Or SDHD Gene Mutations: A Retrospective and Prospective Analysis. Lancet Oncol. 2009, 10, 764–771. [Google Scholar] [CrossRef]
- Korpershoek, E.; Favier, J.; Gaal, J.; Burnichon, N.; van Gessel, B.; Oudijk, L.; Badoual, C.; Gadessaud, N.; Venisse, A.; Bayley, J.; et al. SDHA Immunohistochemistry Detects Germline SDHA Gene Mutations in Apparently Sporadic Paragangliomas and Pheochromocytomas. J. Clin. Endocrinol. Metab. 2011, 96, 1472. [Google Scholar] [CrossRef]
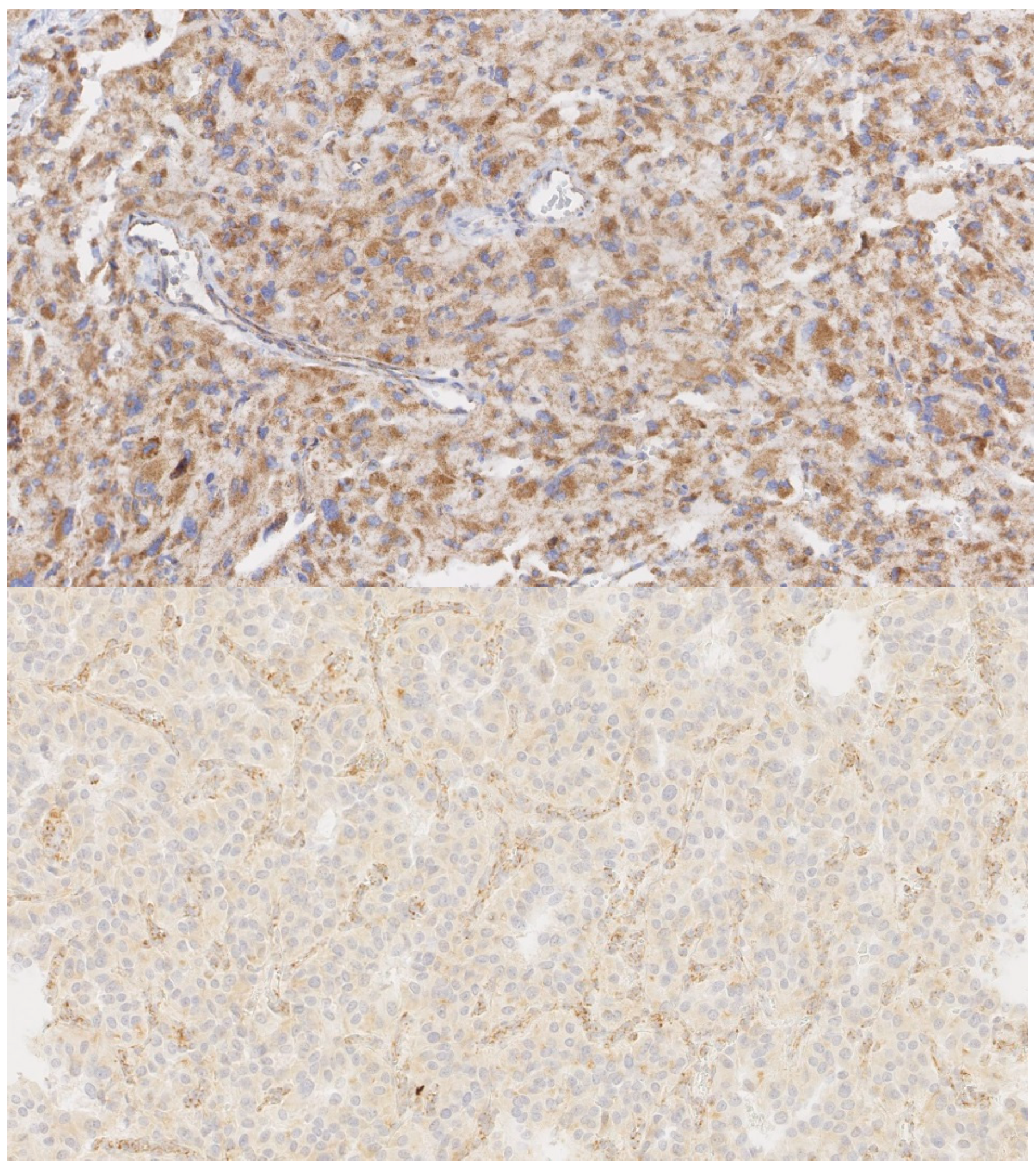
- Papathomas, T.G.; Oudijk, L.; Persu, A.; Gill, A.J.; van Nederveen, F.; Tischler, A.S.; Tissier, F.; Volante, M.; Matias-Guiu, X.; Smid, M.; et al. SDHB/SDHA Immunohistochemistry in Pheochromocytomas and Paragangliomas: A Multicenter Interobserver Variation Analysis using Virtual Microscopy: A Multinational Study of the European Network for the Study of Adrenal Tumors (ENS@T). Mod. Pathol. 2015, 28, 807–821. [Google Scholar] [CrossRef]
- Oudijk, L.; Gaal, J.; de Krijger, R.R. The Role of Immunohistochemistry and Molecular Analysis of Succinate Dehydrogenase in the Diagnosis of Endocrine and Non-Endocrine Tumors and Related Syndromes. Endocr Pathol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Tomlinson, I.P.M.; Alam, N.A.; Rowan, A.J.; Barclay, E.; Jaeger, E.E.M.; Kelsell, D.; Leigh, I.; Gorman, P.; Lamlum, H.; Rahman, S.; et al. Germline Mutations in FH Predispose to Dominantly Inherited Uterine Fibroids, Skin Leiomyomata and Papillary Renal Cell Cancer. Nat. Genet. 2002, 30, 406–410. [Google Scholar] [PubMed]
- Clark, G.R.; Sciacovelli, M.; Gaude, E.; Walsh, D.M.; Kirby, G.; Simpson, M.A.; Trembath, R.C.; Berg, J.N.; Woodward, E.R.; Kinning, E.; et al. Germline FH Mutations Presenting with Pheochromocytoma. J. Clin. Endocrinol. Metab. 2014, 99, 2046. [Google Scholar] [CrossRef] [PubMed]
- Cheung, V.K.Y.; Gill, A.J.; Chou, A. Old, New, and Emerging Immunohistochemical Markers in Pheochromocytoma and Paraganglioma. Endocr. Pathol. 2018, 29, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Castro-Vega, L.J.; Buffet, A.; De Cubas, A.A.; Cascón, A.; Menara, M.; Khalifa, E.; Amar, L.; Azriel, S.; Bourdeau, I.; Chabre, O.; et al. Germline Mutations in FH Confer Predisposition to Malignant Pheochromocytomas and Paragangliomas. Hum. Mol. Genet. 2014, 23, 2440–2446. [Google Scholar] [CrossRef]
- Remacha, L.; Comino-Méndez, I.; Richter, S.; Contreras, L.; Currás-Freixes, M.; Pita, G.; Letón, R.; Galarreta, A.; Torres-Pérez, R.; Honrado, E.; et al. Targeted Exome Sequencing of Krebs Cycle Genes Reveals Candidate Cancer-Predisposing Mutations in Pheochromocytomas and Paragangliomas. Clin. Cancer Res. 2017, 23, 6315–6324. [Google Scholar] [CrossRef] [PubMed]
- Gaal, J.; Burnichon, N.; Korpershoek, E.; Roncelin, I.; Bertherat, J.; Plouin, P.; de Krijger, R.R.; Gimenez-Roqueplo, A.; Dinjens, W.N.M. Isocitrate Dehydrogenase Mutations are Rare in Pheochromocytomas and Paragangliomas. J. Clin. Endocrinol. Metab. 2010, 95, 1274–1278. [Google Scholar] [CrossRef]
- Buffet, A.; Morin, A.; Castro-Vega, L.J.; Habarou, F.; Lussey-Lepoutre, C.; Letouze, E.; Lefebvre, H.; Guilhem, I.; Haissaguerre, M.; Raingeard, I.; et al. Germline Mutations in the Mitochondrial 2-Oxoglutarate/Malate Carrier SLC25A11 Gene Confer a Predisposition to Metastatic Paragangliomas. Cancer Res. 2018, 78, 1914–1922. [Google Scholar] [CrossRef]
- Remacha, L.; Currás-Freixes, M.; Torres-Ruiz, R.; Schiavi, F.; Torres-Pérez, R.; Calsina, B.; Letón, R.; Comino-Méndez, I.; Roldán-Romero, J.M.; Montero-Conde, C.; et al. Gain-of-Function Mutations in DNMT3A in Patients with Paraganglioma. Genet. Med. 2018, 20, 1644–1651. [Google Scholar] [CrossRef]
- Remacha, L.; Pirman, D.; Mahoney, C.E.; Coloma, J.; Calsina, B.; Currás-Freixes, M.; Letón, R.; Torres-Pérez, R.; Richter, S.; Pita, G.; et al. Recurrent Germline DLST Mutations in Individuals with Multiple Pheochromocytomas and Paragangliomas. Am. J. Hum. Genet. 2019, 104, 1008–1010. [Google Scholar] [CrossRef]
- Raue, F.; Frank-Raue, K. Genotype-Phenotype Correlation in Multiple Endocrine Neoplasia Type 2. Clinics 2012, 67, 69–75. [Google Scholar] [CrossRef]
- Korpershoek, E.; Petri, B.; Post, E.; van Eijck, C.H.J.; Oldenburg, R.A.; Belt, E.J.T.; de Herder, W.W.; de Krijger, R.R.; Dinjens, W.N.M. Adrenal Medullary Hyperplasia is a Precursor Lesion for Pheochromocytoma in MEN2 Syndrome. Neoplasia 2014, 16, 868–873. [Google Scholar] [CrossRef]
- Kimura, N.; Watanabe, T.; Fukase, M.; Wakita, A.; Noshiro, T.; Kimura, I. Neurofibromin and NF1 Gene Analysis in Composite Pheochromocytoma and Tumors Associated with Von Recklinghausen’s Disease. Mod. Pathol. 2002, 15, 183–188. [Google Scholar] [CrossRef]
- Neumann, H.P.H.; Sullivan, M.; Winter, A.; Malinoc, A.; Hoffmann, M.M.; Boedeker, C.C.; Bertz, H.; Walz, M.K.; Moeller, L.C.; Schmid, K.W.; et al. Germline Mutations of the TMEM127 Gene in Patients with Paraganglioma of Head and Neck and Extraadrenal Abdominal Sites. J. Clin. Endocrinol. Metab. 2011, 96, 1279. [Google Scholar] [CrossRef]
- Yao, L.; Schiavi, F.; Cascon, A.; Qin, Y.; Inglada-Pérez, L.; King, E.E.; Toledo, R.A.; Ercolino, T.; Rapizzi, E.; Ricketts, C.J.; et al. Spectrum and Prevalence of FP/TMEM127 Gene Mutations in Pheochromocytomas and Paragangliomas. JAMA 2010, 304, 2611–2619. [Google Scholar] [CrossRef]
- Burnichon, N.; Cascon, A.; Schiavi, F.; Morales, N.P.; Comino-Mendez, I.; Abermil, N.; Inglada-Perez, L.; de Cubas, A.A.; Amar, L.; Barontini, M.; et al. MAX Mutations Cause Hereditary and Sporadic Pheochromocytoma and Paraganglioma. Clin. Cancer Res. 2012, 18, 2828–2837. [Google Scholar] [CrossRef]
- Comino-Méndez, I.; Gracia-Aznárez, F.J.; Schiavi, F.; Landa, I.; Leandro-García, L.J.; Letón, R.; Honrado, E.; Ramos-Medina, R.; Caronia, D.; Pita, G.; et al. Exome Sequencing Identifies MAX Mutations as a Cause of Hereditary Pheochromocytoma. Nat. Genet. 2011, 43, 663–667. [Google Scholar] [CrossRef]
- Crona, J.; Delgado Verdugo, A.; Maharjan, R.; Stålberg, P.; Granberg, D.; Hellman, P.; Björklund, P. Somatic Mutations in H-RAS in Sporadic Pheochromocytoma and Paraganglioma Identified by Exome Sequencing. J. Clin. Endocrinol. Metab. 2013, 98, 1266. [Google Scholar] [CrossRef]
- Oudijk, L.; de Krijger, R.R.; Rapa, I.; Beuschlein, F.; de Cubas, A.A.; Dei Tos, A.P.; Dinjens, W.N.M.; Korpershoek, E.; Mancikova, V.; Mannelli, M.; et al. H-RAS Mutations are Restricted to Sporadic Pheochromocytomas Lacking Specific Clinical Or Pathological Features: Data from a Multi-Institutional Series. J. Clin. Endocrinol. Metab. 2014, 99, 1376. [Google Scholar] [CrossRef]
- Evenepoel, L.; Helaers, R.; Vroonen, L.; Aydin, S.; Hamoir, M.; Maiter, D.; Vikkula, M.; Persu, A. KIF1B and NF1 are the most Frequently Mutated Genes in Paraganglioma and Pheochromocytoma Tumors. Endocr. Relat. Cancer 2017, 24, L61. [Google Scholar] [CrossRef]
- Yeh, I.; Lenci, R.E.; Qin, Y.; Buddavarapu, K.; Ligon, A.H.; Leteurtre, E.; Do Cao, C.; Cardot-Bauters, C.; Pigny, P.; Dahia, P.L.M. A Germline Mutation of the KIF1B Beta Gene on 1p36 in a Family with Neural and Nonneural Tumors. Hum. Genet. 2008, 124, 279–285. [Google Scholar] [CrossRef]
- Schussheim, D.H.; Skarulis, M.C.; Agarwal, S.K.; Simonds, W.F.; Burns, A.L.; Spiegel, A.M.; Marx, S.J. Multiple Endocrine Neoplasia Type 1: New Clinical and Basic Findings. Trends Endocrinol. Metab. 2001, 12, 173–178. [Google Scholar] [CrossRef]
- Carty, S.E.; Helm, A.K.; Amico, J.A.; Clarke, M.R.; Foley, T.P.; Watson, C.G.; Mulvihill, J.J. The Variable Penetrance and Spectrum of Manifestations of Multiple Endocrine Neoplasia Type 1. Surgery 1998, 124, 1114. [Google Scholar] [CrossRef]
- Khatami, F.; Tavangar, S.M. Multiple Endocrine Neoplasia Syndromes from Genetic and Epigenetic Perspectives. Biomark Insights 2018, 13. [Google Scholar] [CrossRef]
- Linnoila, R.I.; Keiser, H.R.; Steinberg, S.M.; Lack, E.E. Histopathology of Benign Versus Malignant Sympathoadrenal Paragangliomas: Clinicopathologic Study of 120 Cases Including Unusual Histologic Features. Hum. Pathol. 1990, 21, 1168–1180. [Google Scholar] [CrossRef]
- Wu, D.; Tischler, A.S.; Lloyd, R.V.; DeLellis, R.A.; de Krijger, R.; van Nederveen, F.; Nosé, V. Observer Variation in the Application of the Pheochromocytoma of the Adrenal Gland Scaled Score. Am. J. Surg. Pathol. 2009, 33, 599–608. [Google Scholar] [CrossRef]
- Stenman, A.; Zedenius, J.; Juhlin, C.C. The Value of Histological Algorithms to Predict the Malignancy Potential of Pheochromocytomas and Abdominal Paragangliomas-A Meta-Analysis and Systematic Review of the Literature. Cancers 2019, 11, 225. [Google Scholar] [CrossRef]
- Feng, C.; Li, H.; Yan, W.; Gao, J.; Xu, W.; Luo, Y.; Cao, J. [The Significance of Ki-67 Antigen Expression in the Distinction between Benign and Malignant Pheochromocytomas]. Zhonghua Wai Ke Za Zhi 2007, 45, 1697–1700. [Google Scholar]
- Welander, J.; Söderkvist, P.; Gimm, O. Genetics and Clinical Characteristics of Hereditary Pheochromocytomas and Paragangliomas. Endocr. Relat. Cancer 2011, 18, 253. [Google Scholar]
- Stenman, A.; Zedenius, J.; Juhlin, C.C. Over-Diagnosis of Potential Malignant Behavior in MEN 2A-Associated Pheochromocytomas using the PASS and GAPP Algorithms. Langenbecks Arch. Surg. 2018, 403, 785–790. [Google Scholar] [CrossRef]
- Gimenez-Roqueplo, A.; Favier, J.; Rustin, P.; Rieubland, C.; Crespin, M.; Nau, V.; Khau Van Kien, P.; Corvol, P.; Plouin, P.; Jeunemaitre, X. Mutations in the SDHB Gene are Associated with Extra-Adrenal and/Or Malignant Phaeochromocytomas. Cancer Res. 2003, 63, 5615–5621. [Google Scholar]
- Geli, J.; Kiss, N.; Karimi, M.; Lee, J.; Bäckdahl, M.; Ekström, T.J.; Larsson, C. Global and Regional CpG Methylation in Pheochromocytomas and Abdominal Paragangliomas: Association to Malignant Behavior. Clin. Cancer Res. 2008, 14, 2551–2559. [Google Scholar] [CrossRef]
- Koh, J.; Ahn, S.H.; Kim, H.; Kim, B.; Sung, T.; Kim, Y.H.; Hong, S.J.; Song, D.E.; Lee, S.H. Validation of Pathological Grading Systems for Predicting Metastatic Potential in Pheochromocytoma and Paraganglioma. PLoS ONE 2017, 12, e0187398. [Google Scholar] [CrossRef]


{kind=link}
| GENE | Germline % | Somatic % | Tumor Type | Metastatic Risk % | Syndrome/Other Tumors |
|---|---|---|---|---|---|
| Cluster 1-VHL/EPAS1 | |||||
| VHL | 5–10 | 10 | PCC>>PGL | 5 | VHL syndrome |
| EPAS1/HIF2 alpha | <1 | 5–10 | PCC/ATPGL | 29 | Polycythemia, somatostatinoma |
| EGLN1/EGLN2 | <1 | - | PCC/ATPGL | ? | Polycythemia |
| Cluster 1-TCA | |||||
| SDHx | 20–30 | <1 | |||
| SDHA | <5 | PGL | Low | GIST, RCC, PA | |
| SDHB | 5–10 | ATPGL>>HNPGL/PCC | 30-70 | GIST, RCC, PA | |
| SDHC | <5 | HNPGL | Low | GIST, RCC | |
| SDHD | 5–10 | PGL>PCC | <5 | GIST, RCC, PA | |
| SDHAF2 | <1 | HNPGL | Low | ||
| FH | <5 | - | PCC/PGL | >50 | Leiomyoma, RCC |
| MDH2 | <1 | - | ATPGL | ? | |
| IDH1/IDH2 | - | <1 | PGL | ? | Low grade glioma |
| Recently identified genes | |||||
| SLC25A11 | <1 | - | PGL | High? | |
| IDH3B | <1 | - | HNPGL | ? | AML |
| GOT2 | <1 | - | ATPGL | High? | |
| DNMT3A | <1 | - | HNPGL | ? | AML |
| DLST | <1 | - | PCC/PGL | ? | |
| Cluster 2 | |||||
| RET | 5 | 5 | PCC | <5 | MEN2 syndrome |
| NF1 | <5 | 20–40 | PCC | 12 | NF1 syndrome |
| TMEM127 | <5 | - | PCC>PGL | Low | RCC |
| MAX | <5 | <5 | PCC/PGL | 10 | Renal oncocytoma |
| H-RAS | - | 5–10 | PCC | Low | |
| KIF1B | <1 | 20 | PCC | ? | Neuroblastoma |
| MEN1 | <1 | - | PCC/HNPGL | ? | MEN1 syndrome |
| PASS | GAPP | ||
|---|---|---|---|
| Pheochromocytoma | yes | Pheochromocytoma | yes |
| Paraganglioma | no | Paranganglioma | yes |
| Parameters | Score | Parameters | Score |
| Nuclear hyperchromasia | 1 | Histological Pattern | |
| Zellballen | 0 | ||
| Large and irregular cell nest | 1 | ||
| Pseudorosette (even focal) | 1 | ||
| Profound nuclear pleomorphism | 1 | Cellularity | |
| Low (less than 150 cells/U *) | 0 | ||
| Moderate (150–250 cells/U *) | 1 | ||
| High (more than 250 cells/U *) | 2 | ||
| Capsular invasion | 1 | Comedo Necrosis | |
| Absence | 0 | ||
| Presence | 2 | ||
| Vascular invasion | 1 | Vascular or Capsular Invasion | |
| Absence | 0 | ||
| Presence | 1 | ||
| Extension into periadrenal adipose tissue | 2 | Ki67 Labelling Index | |
| <1% | 0 | ||
| 1–3% | 1 | ||
| >3% | 2 | ||
| Atypical mitotic figures | 2 | Catecholamine Type | |
| Epinephrine type (E **, or E + NE ***) | 0 | ||
| Norepinephrine type (NE, or NE + D ****) | 1 | ||
| Non-functioning type | 1 | ||
| >3 mitotic figures/10 high-power field | 2 | Total | 10 |
| Tumour cell spindling | 2 | ||
| Cellular monotony | 2 | ||
| High cellularity | 2 | ||
| Central or confluent tumour necrosis | 2 | ||
| Large nests or diffuse growth (>10% of tumour volume) | 2 | ||
| Total | 20 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koopman, K.; Gaal, J.; de Krijger, R.R. Pheochromocytomas and Paragangliomas: New Developments with Regard to Classification, Genetics, and Cell of Origin. Cancers 2019, 11, 1070. https://doi.org/10.3390/cancers11081070
Koopman K, Gaal J, de Krijger RR. Pheochromocytomas and Paragangliomas: New Developments with Regard to Classification, Genetics, and Cell of Origin. Cancers. 2019; 11(8):1070. https://doi.org/10.3390/cancers11081070
Chicago/Turabian StyleKoopman, Karen, Jose Gaal, and Ronald R. de Krijger. 2019. "Pheochromocytomas and Paragangliomas: New Developments with Regard to Classification, Genetics, and Cell of Origin" Cancers 11, no. 8: 1070. https://doi.org/10.3390/cancers11081070