LAG3: The Biological Processes That Motivate Targeting This Immune Checkpoint Molecule in Human Cancer


Abstract
:1. Introduction
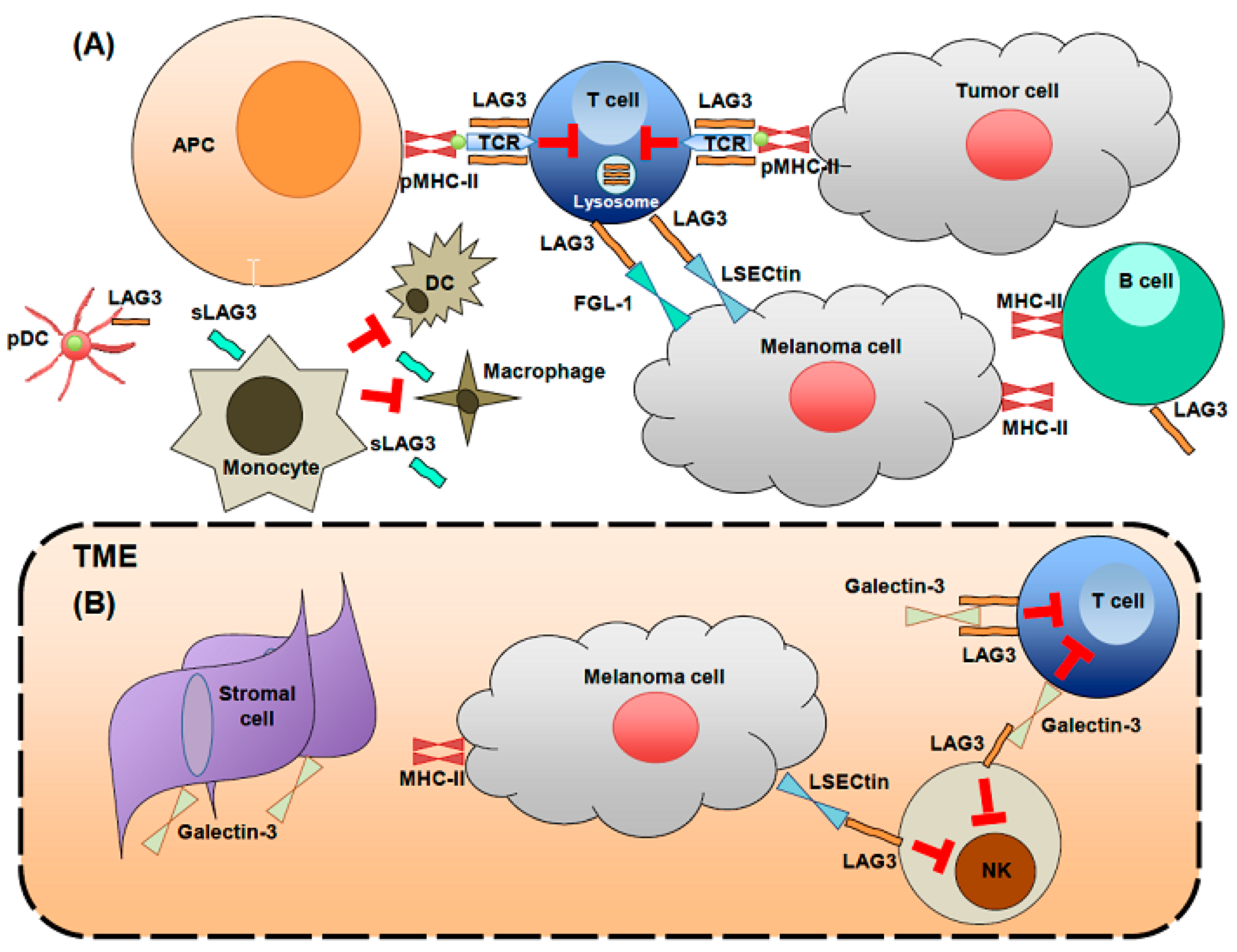
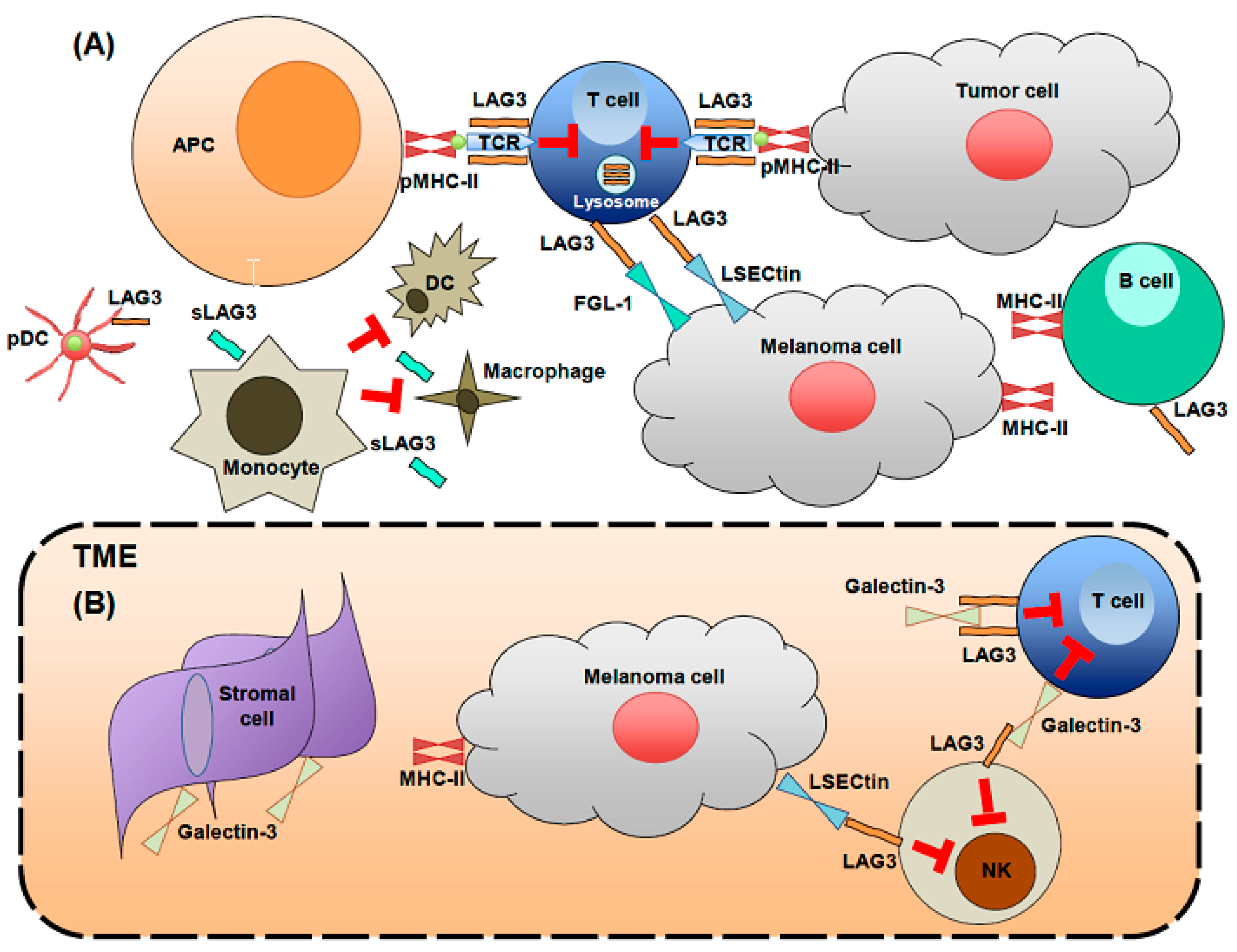
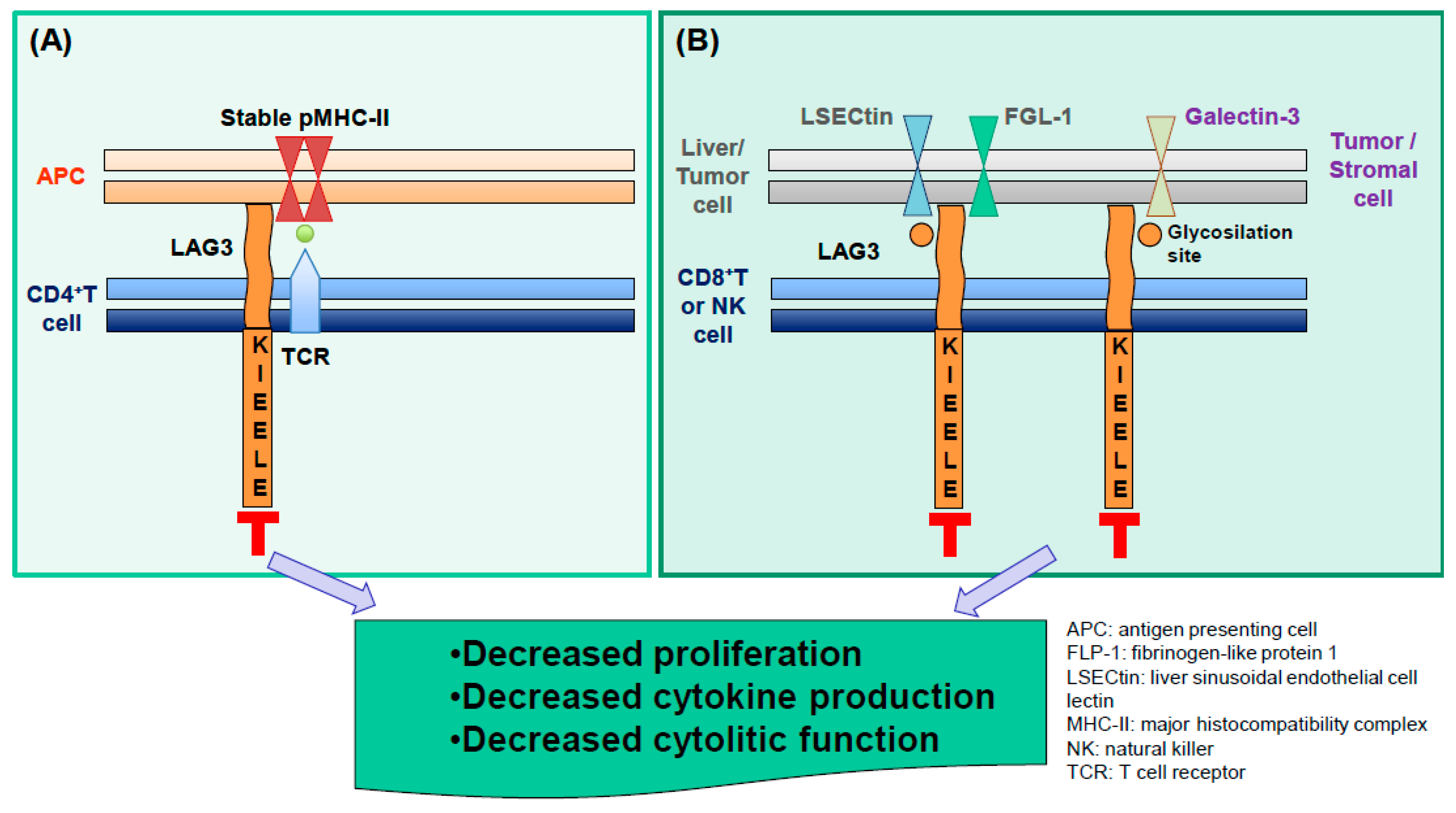
2. LAG3 Biological Activities and Expression
3. LAG3 Expression at Sites Characterized by Chronic Immune Activation
4. LAG3 in Human Breast Cancer
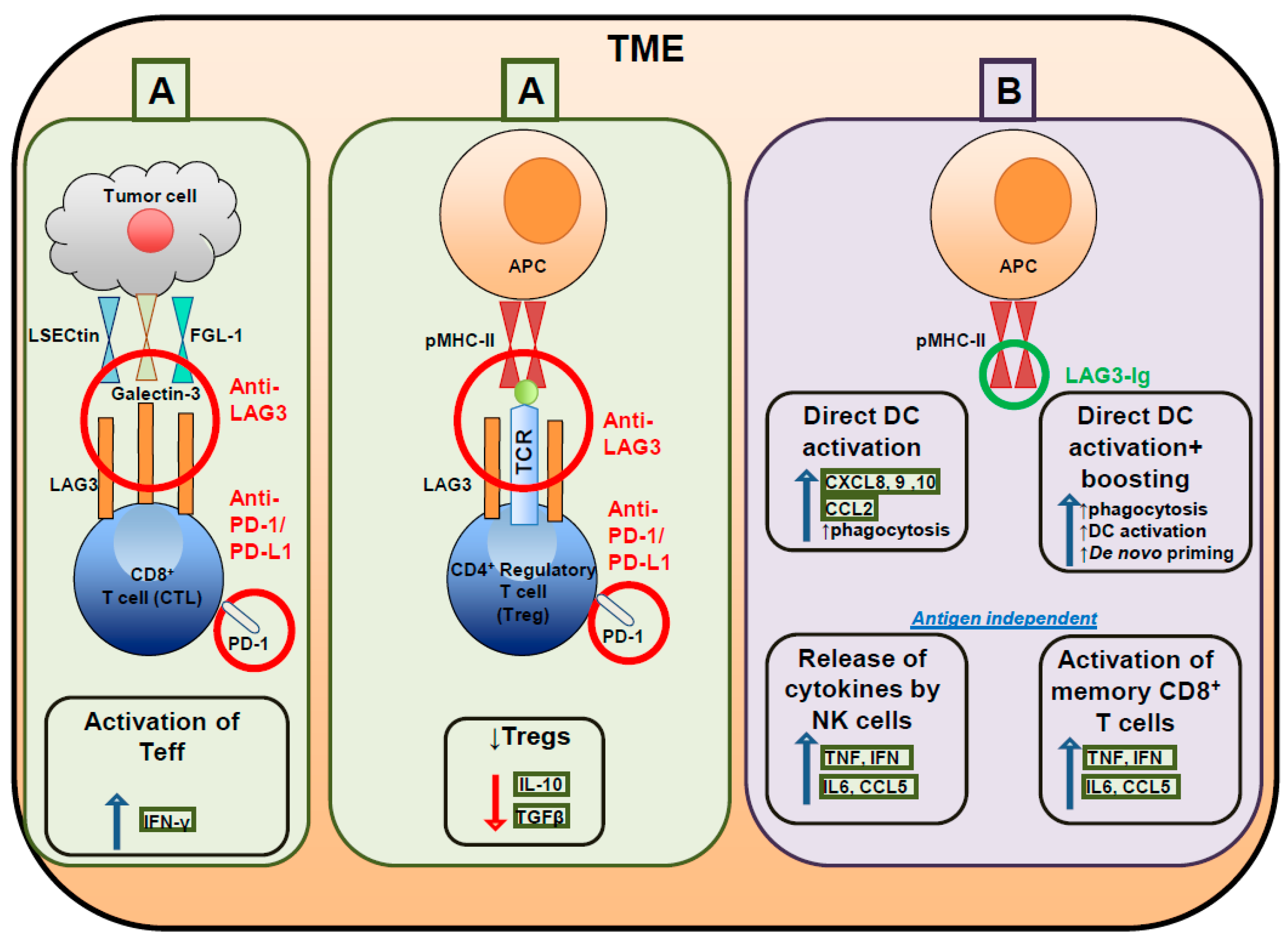
5. LAG3 Blockade in Cancer and Clinical Trials Testing LAG3 Targeting Compounds
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Khair, D.O.; Bax, H.J.; Mele, S.; Crescioli, S.; Pellizzari, G.; Khiabany, A.; Nakamura, M.; Harris, R.J.; French, E.; Hoffmann, R.M.; et al. Combining Immune Checkpoint Inhibitors: Established and Emerging Targets and Strategies to Improve Outcomes in Melanoma. Front. Immunol. 2019, 10, 453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pacheco, J.M.; Camidge, D.R.; Doebele, R.C.; Schenk, E. A Changing of the Guard: Immune Checkpoint Inhibitors with and Without Chemotherapy as First Line Treatment for Metastatic Non-small Cell Lung Cancer. Front. Oncol. 2019, 9, 195. [Google Scholar] [CrossRef] [PubMed]
- Kammerer-Jacquet, S.F.; Deleuze, A.; Saout, J.; Mathieu, R.; Laguerre, B.; Verhoest, G.; Dugay, F.; Belaud-Rotureau, M.A.; Bensalah, K.; Rioux-Leclercq, N. Targeting the PD-1/PD-L1 Pathway in Renal Cell Carcinoma. Int. J. Mol. Sci. 2019, 20, 1692. [Google Scholar] [CrossRef] [PubMed]
- Von der Grun, J.; Rodel, F.; Brandts, C.; Fokas, E.; Guckenberger, M.; Rodel, C.; Balermpas, P. Targeted Therapies and Immune-Checkpoint Inhibition in Head and Neck Squamous Cell Carcinoma: Where Do We Stand Today and Where to Go? Cancers 2019, 11, 472. [Google Scholar] [CrossRef] [PubMed]
- Stahl, M.; Goldberg, A.D. Immune Checkpoint Inhibitors in Acute Myeloid Leukemia: Novel Combinations and Therapeutic Targets. Curr. Oncol. Rep. 2019, 21, 37. [Google Scholar] [CrossRef]
- Esteva, F.J.; Hubbard-Lucey, V.M.; Tang, J.; Pusztai, L. Immunotherapy and targeted therapy combinations in metastatic breast cancer. Lancet Oncol. 2019, 20, e175–e186. [Google Scholar] [CrossRef]
- Doroshow, D.B.; Sanmamed, M.F.; Hastings, K.; Politi, K.; Rimm, D.L.; Chen, L.; Melero, I.; Schalper, K.A.; Herbst, R.S. Immunotherapy in Non-Small Cell Lung Cancer: Facts and Hopes. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2019. [Google Scholar] [CrossRef]
- Caccese, M.; Indraccolo, S.; Zagonel, V.; Lombardi, G. PD-1/PD-L1 immune-checkpoint inhibitors in glioblastoma: A concise review. Crit. Rev. Oncol. Hematol. 2019, 135, 128–134. [Google Scholar] [CrossRef]
- Lattanzi, M.; Balar, A.V. Current Status and Future Direction of Immunotherapy in Urothelial Carcinoma. Curr. Oncol. Rep. 2019, 21, 24. [Google Scholar] [CrossRef]
- Alhalabi, O.; Shah, A.Y.; Lemke, E.A.; Gao, J. Current and Future Landscape of Immune Checkpoint Inhibitors in Urothelial Cancer. Oncology 2019, 33, 11–18. [Google Scholar]
- Franssen, L.E.; Mutis, T.; Lokhorst, H.M.; van de Donk, N. Immunotherapy in myeloma: How far have we come? Ther. Adv. Hematol. 2019, 10, 2040620718822660. [Google Scholar] [CrossRef] [PubMed]
- Sandigursky, S.; Mor, A. Immune-Related Adverse Events in Cancer Patients Treated With Immune Checkpoint Inhibitors. Curr. Rheumatol. Rep. 2018, 20, 65. [Google Scholar] [CrossRef] [PubMed]
- Bhandari, S.; Gill, A.S.; Perez, C.A.; Jain, D. Management of immunotherapy toxicities in older adults. Semin. Oncol. 2018, 45, 226–231. [Google Scholar] [CrossRef] [PubMed]
- Porcu, M.; De Silva, P.; Solinas, C.; Battaglia, A.; Schena, M.; Scartozzi, M.; Bron, D.; Suri, J.S.; Willard-Gallo, K.; Sangiolo, D.; et al. Immunotherapy Associated Pulmonary Toxicity: Biology Behind Clinical and Radiological Features. Cancers 2019, 11, 305. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Saini, S.; Prabhakar, B.S. Cancer immunotherapy with check point inhibitor can cause autoimmune adverse events due to loss of Treg homeostasis. Semin. Cancer Biol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Ruggeri, R.M.; Campenni, A.; Giuffrida, G.; Trimboli, P.; Giovanella, L.; Trimarchi, F.; Cannavo, S. Endocrine and metabolic adverse effects of immune checkpoint inhibitors: An overview (what endocrinologists should know). J. Endocrinol. Investig. 2018. [Google Scholar] [CrossRef] [PubMed]
- Solinas, C.; Porcu, M.; De Silva, P.; Musi, M.; Aspeslagh, S.; Scartozzi, M.; Willard-Gallo, K.; Mariotti, S.; Saba, L. Cancer immunotherapy-associated hypophysitis. Semin. Oncol. 2018, 45, 181–186. [Google Scholar] [CrossRef]
- Solinas, C.; De Silva, P.; Bron, D.; Willard-Gallo, K.D.S. Significance of TIM3 expression in cancer: From biology to the clinic. Under review.
- Sek, K.; Molck, C.; Stewart, G.D.; Kats, L.; Darcy, P.K.; Beavis, P.A. Targeting Adenosine Receptor Signaling in Cancer Immunotherapy. Int. J. Mol. Sci. 2018, 19, 3837. [Google Scholar] [CrossRef]
- Zhu, M.M.T.; Dancsok, A.R.; Nielsen, T.O. Indoleamine Dioxygenase Inhibitors: Clinical Rationale and Current Development. Curr. Oncol. Rep. 2019, 21, 2. [Google Scholar] [CrossRef]
- Solomon, B.L.; Garrido-Laguna, I. TIGIT: A novel immunotherapy target moving from bench to bedside. Cancer Immunol. Immunother. CII 2018, 67, 1659–1667. [Google Scholar] [CrossRef]
- Dougall, W.C.; Kurtulus, S.; Smyth, M.J.; Anderson, A.C. TIGIT and CD96: New checkpoint receptor targets for cancer immunotherapy. Immunol. Rev. 2017, 276, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Hahn, A.W.; Gill, D.M.; Pal, S.K.; Agarwal, N. The future of immune checkpoint cancer therapy after PD-1 and CTLA-4. Immunotherapy 2017, 9, 681–692. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Chikina, M.; Szymczak-Workman, A.L.; Horne, W.; Kolls, J.K.; Vignali, K.M.; Normolle, D.; Bettini, M.; Workman, C.J.; Vignali, D.A.A. LAG3 limits regulatory T cell proliferation and function in autoimmune diabetes. Sci. Immunol. 2017, 2. [Google Scholar] [CrossRef] [PubMed]
- Kadowaki, A.; Miyake, S.; Saga, R.; Chiba, A.; Mochizuki, H.; Yamamura, T. Gut environment-induced intraepithelial autoreactive CD4(+) T cells suppress central nervous system autoimmunity via LAG-3. Nat. Commun. 2016, 7, 11639. [Google Scholar] [CrossRef] [PubMed]
- Van der Zwan, A.; Bi, K.; Norwitz, E.R.; Crespo, A.C.; Claas, F.H.J.; Strominger, J.L.; Tilburgs, T. Mixed signature of activation and dysfunction allows human decidual CD8(+) T cells to provide both tolerance and immunity. Proc. Natl. Acad. Sci. USA 2018, 115, 385–390. [Google Scholar] [CrossRef]
- Gameiro, S.F.; Ghasemi, F.; Barrett, J.W.; Koropatnick, J.; Nichols, A.C.; Mymryk, J.S.; Maleki Vareki, S. Treatment-naive HPV+ head and neck cancers display a T-cell-inflamed phenotype distinct from their HPV- counterparts that has implications for immunotherapy. Oncoimmunology 2018, 7, e1498439. [Google Scholar] [CrossRef] [PubMed]
- Everett, K.L.; Kraman, M.; Wollerton, F.P.G.; Zimarino, C.; Kmiecik, K.; Gaspar, M.; Pechouckova, S.; Allen, N.L.; Doody, J.F.; Tuna, M. Generation of Fcabs targeting human and murine LAG-3 as building blocks for novel bispecific antibody therapeutics. Methods 2018. [Google Scholar] [CrossRef]
- Workman, C.J.; Rice, D.S.; Dugger, K.J.; Kurschner, C.; Vignali, D.A. Phenotypic analysis of the murine CD4-related glycoprotein, CD223 (LAG-3). Eur. J. Immunol. 2002, 32, 2255–2263. [Google Scholar] [CrossRef]
- Workman, C.J.; Wang, Y.; El Kasmi, K.C.; Pardoll, D.M.; Murray, P.J.; Drake, C.G.; Vignali, D.A. LAG-3 regulates plasmacytoid dendritic cell homeostasis. J. Immunol. 2009, 182, 1885–1891. [Google Scholar] [CrossRef]
- Kisielow, M.; Kisielow, J.; Capoferri-Sollami, G.; Karjalainen, K. Expression of lymphocyte activation gene 3 (LAG-3) on B cells is induced by T cells. Eur. J. Immunol. 2005, 35, 2081–2088. [Google Scholar] [CrossRef]
- Kotaskova, J.; Tichy, B.; Trbusek, M.; Francova, H.S.; Kabathova, J.; Malcikova, J.; Doubek, M.; Brychtova, Y.; Mayer, J.; Pospisilova, S. High expression of lymphocyte-activation gene 3 (LAG3) in chronic lymphocytic leukemia cells is associated with unmutated immunoglobulin variable heavy chain region (IGHV) gene and reduced treatment-free survival. J. Mol. Diagn. Jmd 2010, 12, 328–334. [Google Scholar] [CrossRef] [PubMed]
- Lino, A.C.; Dang, V.D.; Lampropoulou, V.; Welle, A.; Joedicke, J.; Pohar, J.; Simon, Q.; Thalmensi, J.; Baures, A.; Fluhler, V.; et al. LAG-3 Inhibitory Receptor Expression Identifies Immunosuppressive Natural Regulatory Plasma Cells. Immunity 2018, 49, 120–133 e9. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.; Ou, M.T.; Karuppagounder, S.S.; Kam, T.I.; Yin, X.; Xiong, Y.; Ge, P.; Umanah, G.E.; Brahmachari, S.; Shin, J.H.; et al. Pathological alpha-synuclein transmission initiated by binding lymphocyte-activation gene 3. Science 2016, 353. [Google Scholar] [CrossRef] [PubMed]
- Bae, J.; Lee, S.J.; Park, C.G.; Lee, Y.S.; Chun, T. Trafficking of LAG-3 to the surface on activated T cells via its cytoplasmic domain and protein kinase C signaling. J. Immunol. 2014, 193, 3101–3112. [Google Scholar] [CrossRef]
- Salgado, R.; Denkert, C.; Demaria, S.; Sirtaine, N.; Klauschen, F.; Pruneri, G.; Wienert, S.; Eynden, G.V.d.; Baehner, F.L.; Penault-Llorca, F.; et al. The evaluation of tumor-infiltrating lymphocytes (TILs) in breast cancer: Recommendations by an International TILs Working Group 2014. Ann. Oncol. 2014, 26, 259–271. [Google Scholar] [CrossRef] [PubMed]
- Baixeras, E.; Huard, B.; Miossec, C.; Jitsukawa, S.; Martin, M.; Hercend, T.; Auffray, C.; Triebel, F.; Piatier-Tonneau, D. Characterization of the lymphocyte activation gene 3-encoded protein. A new ligand for human leukocyte antigen class II antigens. J. Exp. Med. 1992, 176, 327–337. [Google Scholar] [CrossRef]
- Maruhashi, T.; Okazaki, I.M.; Sugiura, D.; Takahashi, S.; Maeda, T.K.; Shimizu, K.; Okazaki, T. LAG-3 inhibits the activation of CD4(+) T cells that recognize stable pMHCII through its conformation-dependent recognition of pMHCII. Nat. Immunol. 2018, 19, 1415–1426. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.T.; Workman, C.J.; Flies, D.; Pan, X.; Marson, A.L.; Zhou, G.; Hipkiss, E.L.; Ravi, S.; Kowalski, J.; Levitsky, H.I.; et al. Role of LAG-3 in regulatory T cells. Immunity 2004, 21, 503–513. [Google Scholar] [CrossRef]
- Previte, D.M.; Martins, C.P.; O’Connor, E.C.; Marre, M.L.; Coudriet, G.M.; Beck, N.W.; Menk, A.V.; Wright, R.H.; Tse, H.M.; Delgoffe, G.M.; et al. Lymphocyte Activation Gene-3 Maintains Mitochondrial and Metabolic Quiescence in Naive CD4(+) T Cells. Cell Rep. 2019, 27, 129–141 e4. [Google Scholar] [CrossRef]
- Gagliani, N.; Magnani, C.F.; Huber, S.; Gianolini, M.E.; Pala, M.; Licona-Limon, P.; Guo, B.; Herbert, D.R.; Bulfone, A.; Trentini, F.; et al. Coexpression of CD49b and LAG-3 identifies human and mouse T regulatory type 1 cells. Nat. Med. 2013, 19, 739–746. [Google Scholar] [CrossRef]
- Liang, B.; Workman, C.; Lee, J.; Chew, C.; Dale, B.M.; Colonna, L.; Flores, M.; Li, N.; Schweighoffer, E.; Greenberg, S.; et al. Regulatory T cells inhibit dendritic cells by lymphocyte activation gene-3 engagement of MHC class II. J. Immunol. 2008, 180, 5916–5926. [Google Scholar] [CrossRef] [PubMed]
- Huard, B.; Tournier, M.; Triebel, F. LAG-3 does not define a specific mode of natural killing in human. Immunol. Lett. 1998, 61, 109–112. [Google Scholar] [CrossRef]
- Graydon, C.G.; Balasko, A.L.; Fowke, K.R. Roles, function and relevance of LAG3 in HIV infection. PLoS Pathog. 2019, 15, e1007429. [Google Scholar] [CrossRef] [PubMed]
- Kurachi, M. CD8(+) T cell exhaustion. Semin. Immunopathol. 2019, 41, 327–337. [Google Scholar] [CrossRef] [PubMed]
- McLane, L.M.; Abdel-Hakeem, M.S.; Wherry, E.J. CD8 T Cell Exhaustion During Chronic Viral Infection and Cancer. Annu. Rev. Immunol. 2019, 37, 457–495. [Google Scholar] [CrossRef] [PubMed]
- Edwards, J.; Wilmott, J.S.; Madore, J.; Gide, T.N.; Quek, C.; Tasker, A.; Ferguson, A.; Chen, J.; Hewavisenti, R.; Hersey, P.; et al. CD103(+) Tumor-Resident CD8(+) T Cells Are Associated with Improved Survival in Immunotherapy-Naive Melanoma Patients and Expand Significantly During Anti-PD-1 Treatment. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2018, 24, 3036–3045. [Google Scholar] [CrossRef] [PubMed]
- Martinet, L.; Smyth, M.J. Balancing natural killer cell activation through paired receptors. Nat. Rev. Immunol. 2015, 15, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Jurewicz, M.M.; Stern, L.J. Class II MHC antigen processing in immune tolerance and inflammation. Immunogenetics 2019, 71, 171–187. [Google Scholar] [CrossRef] [PubMed]
- Axelrod, M.L.; Cook, R.S.; Johnson, D.B.; Balko, J.M. Biological Consequences of MHC-II Expression by Tumor Cells in Cancer. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2019, 25, 2392–2402. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Liu, J.; Liu, D.; Liu, B.; Wang, M.; Hu, Z.; Du, X.; Tang, L.; He, F. LSECtin expressed on melanoma cells promotes tumor progression by inhibiting antitumor T-cell responses. Cancer Res. 2014, 74, 3418–3428. [Google Scholar] [CrossRef] [PubMed]
- Fei, F.; Joo, E.J.; Tarighat, S.S.; Schiffer, I.; Paz, H.; Fabbri, M.; Abdel-Azim, H.; Groffen, J.; Heisterkamp, N. B-cell precursor acute lymphoblastic leukemia and stromal cells communicate through Galectin-3. Oncotarget 2015, 6, 11378–11394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruvolo, P.P. Galectin 3 as a guardian of the tumor microenvironment. Biochim. Et Biophys. Acta 2016, 1863, 427–437. [Google Scholar] [CrossRef] [PubMed]
- Kaur, M.; Kumar, D.; Butty, V.; Singh, S.; Esteban, A.; Fink, G.R.; Ploegh, H.L.; Sehrawat, S. Galectin-3 Regulates gamma-Herpesvirus Specific CD8 T Cell Immunity. iScience 2018, 9, 101–119. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Sanmamed, M.F.; Datar, I.; Su, T.T.; Ji, L.; Sun, J.; Chen, L.; Chen, Y.; Zhu, G.; Yin, W.; et al. Fibrinogen-like Protein 1 Is a Major Immune Inhibitory Ligand of LAG-3. Cell 2019, 176, 334–347. [Google Scholar] [CrossRef] [PubMed]
- Lienhardt, C.; Azzurri, A.; Amedei, A.; Fielding, K.; Sillah, J.; Sow, O.Y.; Bah, B.; Benagiano, M.; Diallo, A.; Manetti, R.; et al. Active tuberculosis in Africa is associated with reduced Th1 and increased Th2 activity in vivo. Eur. J. Immunol. 2002, 32, 1605–1613. [Google Scholar] [CrossRef] [Green Version]
- Triebel, F.; Hacene, K.; Pichon, M.F. A soluble lymphocyte activation gene-3 (sLAG-3) protein as a prognostic factor in human breast cancer expressing estrogen or progesterone receptors. Cancer Lett. 2006, 235, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Jilisihan, B.; Wang, W.; Tang, Y.; Keyoumu, S. Soluble LAG3 acts as a potential prognostic marker of gastric cancer and its positive correlation with CD8+T cell frequency and secretion of IL-12 and INF-gamma in peripheral blood. Cancer Biomark. Sect. A Dis. Markers 2018, 23, 341–351. [Google Scholar] [CrossRef]
- Anderson, A.C.; Joller, N.; Kuchroo, V.K. Lag-3, Tim-3, and TIGIT: Co-inhibitory Receptors with Specialized Functions in Immune Regulation. Immunity 2016, 44, 989–1004. [Google Scholar] [CrossRef] [Green Version]
- Okazaki, T.; Okazaki, I.M.; Wang, J.; Sugiura, D.; Nakaki, F.; Yoshida, T.; Kato, Y.; Fagarasan, S.; Muramatsu, M.; Eto, T.; et al. PD-1 and LAG-3 inhibitory co-receptors act synergistically to prevent autoimmunity in mice. J. Exp. Med. 2011, 208, 395–407. [Google Scholar] [CrossRef] [Green Version]
- Blackburn, S.D.; Shin, H.; Haining, W.N.; Zou, T.; Workman, C.J.; Polley, A.; Betts, M.R.; Freeman, G.J.; Vignali, D.A.; Wherry, E.J. Coregulation of CD8+ T cell exhaustion by multiple inhibitory receptors during chronic viral infection. Nat. Immunol. 2009, 10, 29–37. [Google Scholar] [CrossRef]
- Butler, N.S.; Moebius, J.; Pewe, L.L.; Traore, B.; Doumbo, O.K.; Tygrett, L.T.; Waldschmidt, T.J.; Crompton, P.D.; Harty, J.T. Therapeutic blockade of PD-L1 and LAG-3 rapidly clears established blood-stage Plasmodium infection. Nat. Immunol. 2011, 13, 188–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuzaki, J.; Gnjatic, S.; Mhawech-Fauceglia, P.; Beck, A.; Miller, A.; Tsuji, T.; Eppolito, C.; Qian, F.; Lele, S.; Shrikant, P.; et al. Tumor-infiltrating NY-ESO-1-specific CD8+ T cells are negatively regulated by LAG-3 and PD-1 in human ovarian cancer. Proc. Natl. Acad. Sci. USA 2010, 107, 7875–7880. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.Y.; Francois, A.; McGray, A.R.; Miliotto, A.; Odunsi, K. Compensatory upregulation of PD-1, LAG-3, and CTLA-4 limits the efficacy of single-agent checkpoint blockade in metastatic ovarian cancer. Oncoimmunology 2017, 6, e1249561. [Google Scholar] [CrossRef] [PubMed]
- Llosa, N.J.; Cruise, M.; Tam, A.; Wicks, E.C.; Hechenbleikner, E.M.; Taube, J.M.; Blosser, R.L.; Fan, H.; Wang, H.; Luber, B.S.; et al. The vigorous immune microenvironment of microsatellite instable colon cancer is balanced by multiple counter-inhibitory checkpoints. Cancer Discov. 2015, 5, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Camisaschi, C.; Casati, C.; Rini, F.; Perego, M.; De Filippo, A.; Triebel, F.; Parmiani, G.; Belli, F.; Rivoltini, L.; Castelli, C. LAG-3 expression defines a subset of CD4(+)CD25(high)Foxp3(+) regulatory T cells that are expanded at tumor sites. J. Immunol. 2010, 184, 6545–6551. [Google Scholar] [CrossRef] [PubMed]
- Jie, H.B.; Gildener-Leapman, N.; Li, J.; Srivastava, R.M.; Gibson, S.P.; Whiteside, T.L.; Ferris, R.L. Intratumoral regulatory T cells upregulate immunosuppressive molecules in head and neck cancer patients. Br. J. Cancer 2013, 109, 2629–2635. [Google Scholar] [CrossRef] [PubMed]
- Wei, T.; Zhang, J.; Qin, Y.; Wu, Y.; Zhu, L.; Lu, L.; Tang, G.; Shen, Q. Increased expression of immunosuppressive molecules on intratumoral and circulating regulatory T cells in non-small-cell lung cancer patients. Am. J. Cancer Res. 2015, 5, 2190–2201. [Google Scholar] [PubMed]
- Solinas, C.; Garaud, S.; De Silva, P.; Boisson, A.; Van den Eynden, G.; de Wind, A.; Risso, P.; Rodrigues Vitória, J.; Richard, F.; Migliori, E.; et al. Immune Checkpoint Molecules on Tumor-Infiltrating Lymphocytes and Their Association with Tertiary Lymphoid Structures in Human Breast Cancer. Front. Immunol. 2017, 7. [Google Scholar] [CrossRef]
- Burugu, S.; Gao, D.; Leung, S.; Chia, S.K.; Nielsen, T.O. LAG-3+ tumor infiltrating lymphocytes in breast cancer: Clinical correlates and association with PD-1/PD-L1+ tumors. Ann. Oncol. Off. J. Eur. Soc. Med Oncol. 2017, 28, 2977–2984. [Google Scholar] [CrossRef]
- Wang, Y.; Dong, T.; Xuan, Q.; Zhao, H.; Qin, L.; Zhang, Q. Lymphocyte-Activation Gene-3 Expression and Prognostic Value in Neoadjuvant-Treated Triple-Negative Breast Cancer. J. Breast Cancer 2018, 21, 124–133. [Google Scholar] [CrossRef]
- Bottai, G.; Raschioni, C.; Losurdo, A.; Di Tommaso, L.; Tinterri, C.; Torrisi, R.; Reis-Filho, J.S.; Roncalli, M.; Sotiriou, C.; Santoro, A.; et al. An immune stratification reveals a subset of PD-1/LAG-3 double-positive triple-negative breast cancers. Breast Cancer Res. BCR 2016, 18, 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, D.B.; Nixon, M.J.; Wang, Y.; Wang, D.Y.; Castellanos, E.; Estrada, M.V.; Ericsson-Gonzalez, P.I.; Cote, C.H.; Salgado, R.; Sanchez, V.; et al. Tumor-specific MHC-II expression drives a unique pattern of resistance to immunotherapy via LAG-3/FCRL6 engagement. JCI Insight 2018, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasidharan Nair, V.; El Salhat, H.; Taha, R.Z.; John, A.; Ali, B.R.; Elkord, E. DNA methylation and repressive H3K9 and H3K27 trimethylation in the promoter regions of PD-1, CTLA-4, TIM-3, LAG-3, TIGIT, and PD-L1 genes in human primary breast cancer. Clin. Epigenetics 2018, 10, 78. [Google Scholar] [CrossRef] [PubMed]
- Gil Del Alcazar, C.R.; Huh, S.J.; Ekram, M.B.; Trinh, A.; Liu, L.L.; Beca, F.; Zi, X.; Kwak, M.; Bergholtz, H.; Su, Y.; et al. Immune Escape in Breast Cancer During In Situ to Invasive Carcinoma Transition. Cancer Discov. 2017, 7, 1098–1115. [Google Scholar] [CrossRef] [PubMed]
- Elashi, A.A.; Sasidharan Nair, V.; Taha, R.Z.; Shaath, H.; Elkord, E. DNA methylation of immune checkpoints in the peripheral blood of breast and colorectal cancer patients. Oncoimmunology 2019, 8, e1542918. [Google Scholar] [CrossRef]
- Lui, Y.; Davis, S.J. LAG-3: A very singular immune checkpoint. Nat. Immunol. 2018, 19, 1278–1279. [Google Scholar] [CrossRef] [PubMed]
- Ascierto, P.A.; Melero, I.; Bhatia, S.; Bono, P.; Sanborn, R.E.; Lipson, E.J.; Callahan, M.K.; Gajewski, T.; Gomez-Roca, C.A.; Hodi, F.S.; et al. Initial efficacy of anti-lymphocyte activation gene-3 (anti–LAG-3; BMS-986016) in combination with nivolumab (nivo) in pts with melanoma (MEL) previously treated with anti–PD-1/PD-L1 therapy. J. Clin. Oncol. 2017, 35, 9520. [Google Scholar] [CrossRef]
- Hodi, F.S.; Chiarion-Sileni, V.; Gonzalez, R.; Grob, J.J.; Rutkowski, P.; Cowey, C.L.; Lao, C.D.; Schadendorf, D.; Wagstaff, J.; Dummer, R.; et al. Nivolumab plus ipilimumab or nivolumab alone versus ipilimumab alone in advanced melanoma (CheckMate 067): 4-year outcomes of a multicentre, randomised, phase 3 trial. Lancet Oncol. 2018, 19, 1480–1492. [Google Scholar] [CrossRef]
- Prigent, P.; El Mir, S.; Dreano, M.; Triebel, F. Lymphocyte activation gene-3 induces tumor regression and antitumor immune responses. Eur. J. Immunol. 1999, 29, 3867–3876. [Google Scholar] [CrossRef]
- Romano, E.; Michielin, O.; Voelter, V.; Laurent, J.; Bichat, H.; Stravodimou, A.; Romero, P.; Speiser, D.E.; Triebel, F.; Leyvraz, S.; et al. MART-1 peptide vaccination plus IMP321 (LAG-3Ig fusion protein) in patients receiving autologous PBMCs after lymphodepletion: Results of a Phase I trial. J. Transl. Med. 2014, 12, 97. [Google Scholar] [CrossRef]
- Avice, M.N.; Sarfati, M.; Triebel, F.; Delespesse, G.; Demeure, C.E. Lymphocyte activation gene-3, a MHC class II ligand expressed on activated T cells, stimulates TNF-alpha and IL-12 production by monocytes and dendritic cells. J. Immunol. 1999, 162, 2748–2753. [Google Scholar] [PubMed]
- Brignone, C.; Escudier, B.; Grygar, C.; Marcu, M.; Triebel, F. A phase I pharmacokinetic and biological correlative study of IMP321, a novel MHC class II agonist, in patients with advanced renal cell carcinoma. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2009, 15, 6225–6231. [Google Scholar] [CrossRef] [PubMed]
- Wang-Gillam, A.; Plambeck-Suess, S.; Goedegebuure, P.; Simon, P.O.; Mitchem, J.B.; Hornick, J.R.; Sorscher, S.; Picus, J.; Suresh, R.; Lockhart, A.C.; et al. A phase I study of IMP321 and gemcitabine as the front-line therapy in patients with advanced pancreatic adenocarcinoma. Investig. New Drugs 2013, 31, 707–713. [Google Scholar] [CrossRef] [PubMed]
- Ascierto, P.A.; Bono, P.; Bhatia, S.; Melero, I.; Nyakas, M.S.; Svane, I.-M.; Larkin, J.; Gomez-Roca, C.; Schadendorf, D.; Dummer, R.; et al. LBA18. Efficacy of BMS-986016, a monoclonal antibody that targets lymphocyte activation gene-3 (LAG-3), in combination with nivolumab in pts with melanoma who progressed during prior anti–PD-1/PD-L1 therapy (mel prior IO) in all-comer and biomarker-enriched populations. Ann. Oncol. 2017, 28, mdx440.011. [Google Scholar] [CrossRef]
- Woo, S.R.; Turnis, M.E.; Goldberg, M.V.; Bankoti, J.; Selby, M.; Nirschl, C.J.; Bettini, M.L.; Gravano, D.M.; Vogel, P.; Liu, C.L.; et al. Immune inhibitory molecules LAG-3 and PD-1 synergistically regulate T-cell function to promote tumoral immune escape. Cancer Res. 2012, 72, 917–927. [Google Scholar] [CrossRef] [PubMed]
- Ko, E.C.; Raben, D.; Formenti, S.C. The Integration of Radiotherapy with Immunotherapy for the Treatment of Non-Small Cell Lung Cancer. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2018. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Reference | Drug(s) | Phase | Tumor Type | Main Objectives | Status |
|---|---|---|---|---|---|
| NCT 02676869 | IMP321 (LAG3 Ig fusion protein-eftilagimod alpha) + pembrolizumab (anti-PD-1) | I | Stage IV and stage III melanoma | Safety, tolerability and recommended phase 2 dose | Active, not yet recruiting |
| NCT 02614833 | IMP321 (LAG3 Ig fusion protein-eftilagimod alpha) + paclitaxel | IIb, randomized | Hormone receptor-positive stage IV breast cancer | Recommended phase II dose, safety and efficacy (survival and objective response rate) | Recruiting |
| NCT 03252938 | IMP321 (LAG3 Ig fusion protein-eftilagimod alpha) | I | Metastatic solid tumors; peritoneal carcinomatosis | Feasibility and safety of intra-tumoral and intra-peritoneal (new routes of administration), and subcutaneous injections | Recruiting |
| NCT 03625323 | IMP321 (LAG3 Ig fusion protein-eftilagimod alpha) + pembrolizumab (anti-PD-1) | II | Advanced or metastatic non-small cell lung carcinoma and head and neck squamous cell carcinoma | Safety and efficacy (objective response rate) | Recruiting |
| NCT 03642067 | Relatlimab (anti-LAG3 monoclonal antibody BMS-986016) + nivolumab (anti-PD-1) | II | Microsatellite stable (MSS) colorectal carcinomas Colorectal carcinoma | Response (objective response rate) and biomarkers | Recruiting |
| NCT 03623854 | Relatlimab (anti-LAG3 monoclonal antibody BMS-986016) + nivolumab (anti-PD-1) | II | Advanced chordomas | Clinical benefits (objective response rate, progression free survival) and safety | Not yet recruiting |
| NCT 03743766 | Relatlimab (anti-LAG3 monoclonal antibody BMS-986016) +/- nivolumab (anti-PD-1) | II | Stage III or stage IV melanoma | Change in LAG3 and PD-1 expression; change in tumor size and overall response rate | Recruiting |
| NCT 03607890 | Relatlimab (anti-LAG3 monoclonal antibody BMS-986016) + nivolumab (anti-PD-1) | II | Refractory microsatellite unstable high (MSI-H) solid tumors prior of PD-(L) 1 therapy MSI-H tumors | Objective response rate, toxicity, survival, disease control rate, best overall response, duration of response, duration of clinical benefit, time to objective response | Recruiting |
| NCT 03724968 | Relatlimab (anti-LAG3 monoclonal antibody BMS-986016) +/- nivolumab (anti-PD-1); nivolumab + ipilimumab (anti-CTLA-4) | II | Stage III and stage IV melanoma, stratified by MHC-II expression | Efficacy, measured by change in activated GZMB+ CD8+ T-cell density intratumorally; response rate, median progression free survival, overall survival, and safety and tolerability of nivolumab plus relatlimab in patients with MHC-II (+) melanoma, and of nivolumab plus ipilimumab in patients with MHC-II (-) melanoma; explore potential associations of biomarkers with clinical efficacy and/or incidence of adverse events due to study drug by analyzing biomarker measures within the peripheral blood and tumor microenvironment | Recruiting |
| NCT 03470922 | Nivolumab (anti-PD-1) +/- relatlimab (anti-LAG3 monoclonal antibody BMS-986016) | II/III | Previously untreated stage III or stage IV melanoma | Survival (progression free survival, overall survival), responses (objective response rate), duration of response, toxicity | Active, not yet recruiting |
| NCT 03044613 | Nivolumab (anti-PD-1) or Nivolumab plus relatlimab (anti-LAG3 monoclonal antibody BMS-986016); chemoradiation | Ib | Stage II/III gastric cancer, esophageal cancer, gastroesophageal cancer | Toxicity, feasibility, pathologic complete response rate; approximate quantitation of infused nivolumab bound to PD-1 receptors on the surface of T cells in the peripheral blood and within the resected tumor and lymph node specimens; changes in expression of selected immune markers; survival (overall and recurrence free survival) | Recruiting |
| NCT 03459222 | Relatlimab (anti-LAG3 monoclonal antibody BMS-986016) + nivolumab (anti-PD-1) or + BMS-986205 (IDO1 inhibitor) or relatlimab + nivolumab and ipilimumab (anti-CTLA-4) | I/II | Advanced malignant tumors | Toxicity, safety, objective response rate, disease control rate, median duration of response | Recruiting |
| NCT 03867799 | Relatlimab (anti-LAG3 monoclonal antibody BMS-986016) + nivolumab (anti-PD-1) | II | Metastatic colorectal cancer | Disease control rate, toxicity, duration of disease control, best objective response rate, progression free survival, overall survival | Not yet recruiting |
| NCT 02519322 | Nivolumab (anti-PD-1); nivolumab + relatlimab (anti-LAG3 monoclonal antibody BMS-986016); nivolumab + ipilimumab (anti-CTLA-4) | II | Melanoma | Pathologic response; immunological response (changes in T cell infiltrate); objective response; survival (recurrence free and overall survival); adverse events | Recruiting |
| NCT 03704077 | Relatlimab (anti-LAG3 monoclonal antibody BMS-986016) + nivolumab (anti-PD-1) + paclitaxel; nivolumab + paclitaxel; ramucirumab + paclitaxel; relatlimab + nivolumab; nivolumab | II | Recurrent, locally advanced, or metastatic gastric cancer (GC) or gastroesophageal junction (GEJ) adenocarcinoma | Overall response rate; toxicity; duration of response; survival (progression free survival and overall survival) | Not yet recruiting |
| NCT 01968109 | Relatlimab (anti-LAG3 monoclonal antibody BMS-986016) + nivolumab (anti-PD-1) | I/II | Advanced solid tumors | Safety (adverse events), tolerability and efficacy (objective response rate, disease control rate, duration of response) | Recruiting |
| NCT 03662659 | Relatlimab (anti-LAG3 monoclonal antibody BMS-986016) + investigator’s choice chemotherapy; nivolumab (anti-PD-1) + investigator’s choice chemotherapy | II | Unresectable, untreated, locally advanced or metastatic gastric or gastroesophageal junction cancer | Objective response rate; toxicity; duration of response; survival (overall survival and progression free survival) | Recruiting |
| NCT 03610711 | Nivolumab (anti-PD-1) +/- relatlimab (anti-LAG3 monoclonal antibody BMS-986016) + stereotactic radiotherapy | Ib/II | Recurrent or limited metastatic gastroesophageal cancer | Change in the infiltrating CD8+ T cell density; safety; efficacy | Recruiting |
| NCT 02966548 | Relatlimab (anti-LAG3 monoclonal antibody BMS-986016) +/- nivolumab (anti-PD-1) | I | Advanced solid tumors | Safety, tolerability, and efficacy | Recruiting |
| NCT 02996110 | Relatlimab (anti-LAG3 monoclonal antibody BMS-986016) + nivolumab (anti-PD-1); nivolumab + BMS-986205 (IDO-inhibitor); nivolumab + ipilimumab (anti-CTLA-4) | II | Advanced renal cell carcinoma | Objective response rate; duration of response; progression free survival rate; safety; tolerability | Recruiting |
| NCT 02935634 | Relatlimab (anti-LAG3 monoclonal antibody BMS-986016) + nivolumab (anti-PD-1); nivolumab + ipilimumab (anti-CTLA-4); nivolumab and BMS-986205 (IDO-inhibitor) | II | Advanced gastric cancer | Objective response rate; duration of response; progression free survival rate; adverse events | Recruiting |
| NCT 02488759 | Nivolumab (anti-PD-1); nivolumab + ipilimumab (anti-CTLA-4); nivolumab + relatlimab (anti-LAG3 monoclonal antibody BMS-986016); nivolumab + daratumumab (anti-CD38) | I/II | Virus associated tumors (anal canal cancer; cervical cancer; Epstein Barr Virus (EBV) positive gastric cancer; HPV positive and negative squamous cell cancer of the head and neck (SCCHN); Merkel cell cancer; nasopharyngeal cancer; penile cancer; vaginal and vulvar cancer | Safety, tolerability; objective response rate; survival (progression free survival; overall survival); duration of response | Recruiting |
| NCT 02750514 | Nivolumab (anti-PD-1); nivolumab + dasatinib (src, c-Kit, ephri receptor inhibitor); nivolumab + relatlimab (anti-LAG3 monoclonal antibody BMS-986016); nivolumab + ipilimumab (anti-CTLA-4); nivolumab + BMS-986205 (IDO-inhibitor) | II | Advanced non-small cell lung cancer | Objective response; duration of response; progression free survival rate; safety and tolerability | Recruiting |
| NCT 03335540 | Nivolumab (anti-PD-1) + relatlimab (anti-LAG3 monoclonal antibody BMS-986016); nivolumab + lirilumab (anti-KIR monoclonal antibody); nivolumab + cabiralizumab (anti-CSF1R); nivolumab + ipilimumab (anti-CTLA-4); nivolumab + anti-GITR; nivolumab + BMS-986205 (IDO-inhibitor); nivolumab + radiation therapy | I | Solid tumors | Number of patients with biopsy specimens; change from baseline in histopatologic and biomarker expression patterns; number of adverse events; | Recruiting |
| NCT 03219268 | MGD013 (Anti-PD-1, anti-LAG-3 bispecific DART protein) | I | Unresectable, locally advanced or metastatic solid tumors of any histology | Adverse events; pharmacokinetics; pharmacodynamics; immunogenicity, and preliminary antitumor activity | Recruiting |
| NCT 03365791 | LAG525 (anti-LAG3 IgG4 antibody) + PDR001 (anti-PD-1 IgG4 antibody) | II | Advanced solid and hematological malignancies | Clinical benefit rate; progression free survival; overall response rate; time to response; safety and tolerability; duration of response; time to progression | Active, not recruiting |
| NCT 03499899 | LAG525 (anti-LAG3 IgG4 antibody) + PDR001 (anti-PD-1 IgG4 antibody); LAG525 + PDR001 + carboplatin; LAG525 + carboplatin | II | Advanced triple negative breast cancer (first or second line) | Overall response rate; duration of response; overall survival; pharmacokinetic parameters; time to response; progression free survival; clinical benefit rate; anti-drug antibodies prevalence at baseline and incidence on treatment | Recruiting |
| NCT 02460224 | LAG525 (anti-LAG3 IgG4 antibody) +/- PDR001 (anti-PD-1 IgG4 antibody) | I/II | Advances malignancies | Dose limiting toxicities; overall response rate; area under the curve; concentration of anti-LAG525 and anti-PDR001 antibodies; correlation of PD-L1 and LAG-3 expression; overall response rate; expression of IFN-γ immune-related genes by mRNA profiling; safety; tolerability; progression free survival; duration of response; disease control rate | Active, not recruiting |
| NCT 03742349 | LAG525 (anti-LAG3 IgG4 antibody) + PDR001 (anti-PD-1 IgG4 antibody) + NIR178 (adenosine A2A receptor antagonist); LAG525 + PDR001 + capmatinib (c-Met inhibitor); LAG525 + PDR001 + MCS110 (anti-M-CSF antibody); LAG525 + PDR001 + canakinumab (anti-interleukin-1 beta) | Ib | Adult patients with triple negative breast cancer | Safety; dose limiting toxicities; best overall response; progression free survival; presence of anti-PDR001. of anti-LAG525, of anti-MCS110, of anti- canakinumab antibodies; pharmacokinetic and pharmacodynamic parameters | Recruiting |
| NCT 03484923 | LAG525 (anti-LAG3 IgG4 antibody) + PDR001 (anti-PD-1 IgG4 antibody); PDR001 + capmatinib (c-Met inhibitor); PDR001 + canakinumab (anti-interleukin-1 beta) | II | Previously treated stage III and stage IV melanoma | Overall response rate; duration of response; survival (overall survival and progression free survival); disease control rate; prevalence and incidence of anti-drug antibodies; frequency of patients with a favorable biomarker profile | Recruiting |
| NCT 03005782 | REGN3767 (anti-LAG3 monoclonal antibody) +/- REGN2810 (anti-PD-1 monoclonal antibody) | I | Advanced tumors | Safety; tolerability; activity and pharmacokinetics | Recruiting |
| NCT 03250832 | TSR-033 (humanized monoclonal anti-LAG3 IgG4) +/- anti-PD-1 (IgG4 antibody) | I | Advanced solid tumors | Adverse events; responses; pharmacokinetic parameters; duration of response; disease control rate; survival (progression free survival; overall survival) | Recruiting |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Solinas, C.; Migliori, E.; De Silva, P.; Willard-Gallo, K. LAG3: The Biological Processes That Motivate Targeting This Immune Checkpoint Molecule in Human Cancer. Cancers 2019, 11, 1213. https://doi.org/10.3390/cancers11081213
Solinas C, Migliori E, De Silva P, Willard-Gallo K. LAG3: The Biological Processes That Motivate Targeting This Immune Checkpoint Molecule in Human Cancer. Cancers. 2019; 11(8):1213. https://doi.org/10.3390/cancers11081213
Chicago/Turabian StyleSolinas, Cinzia, Edoardo Migliori, Pushpamali De Silva, and Karen Willard-Gallo. 2019. "LAG3: The Biological Processes That Motivate Targeting This Immune Checkpoint Molecule in Human Cancer" Cancers 11, no. 8: 1213. https://doi.org/10.3390/cancers11081213
APA StyleSolinas, C., Migliori, E., De Silva, P., & Willard-Gallo, K. (2019). LAG3: The Biological Processes That Motivate Targeting This Immune Checkpoint Molecule in Human Cancer. Cancers, 11(8), 1213. https://doi.org/10.3390/cancers11081213