Distinct Pattern of Endoplasmic Reticulum Protein Processing and Extracellular Matrix Proteins in Functioning and Silent Corticotroph Pituitary Adenomas

 ,
,
Abstract
:Simple Summary
Abstract

1. Introduction
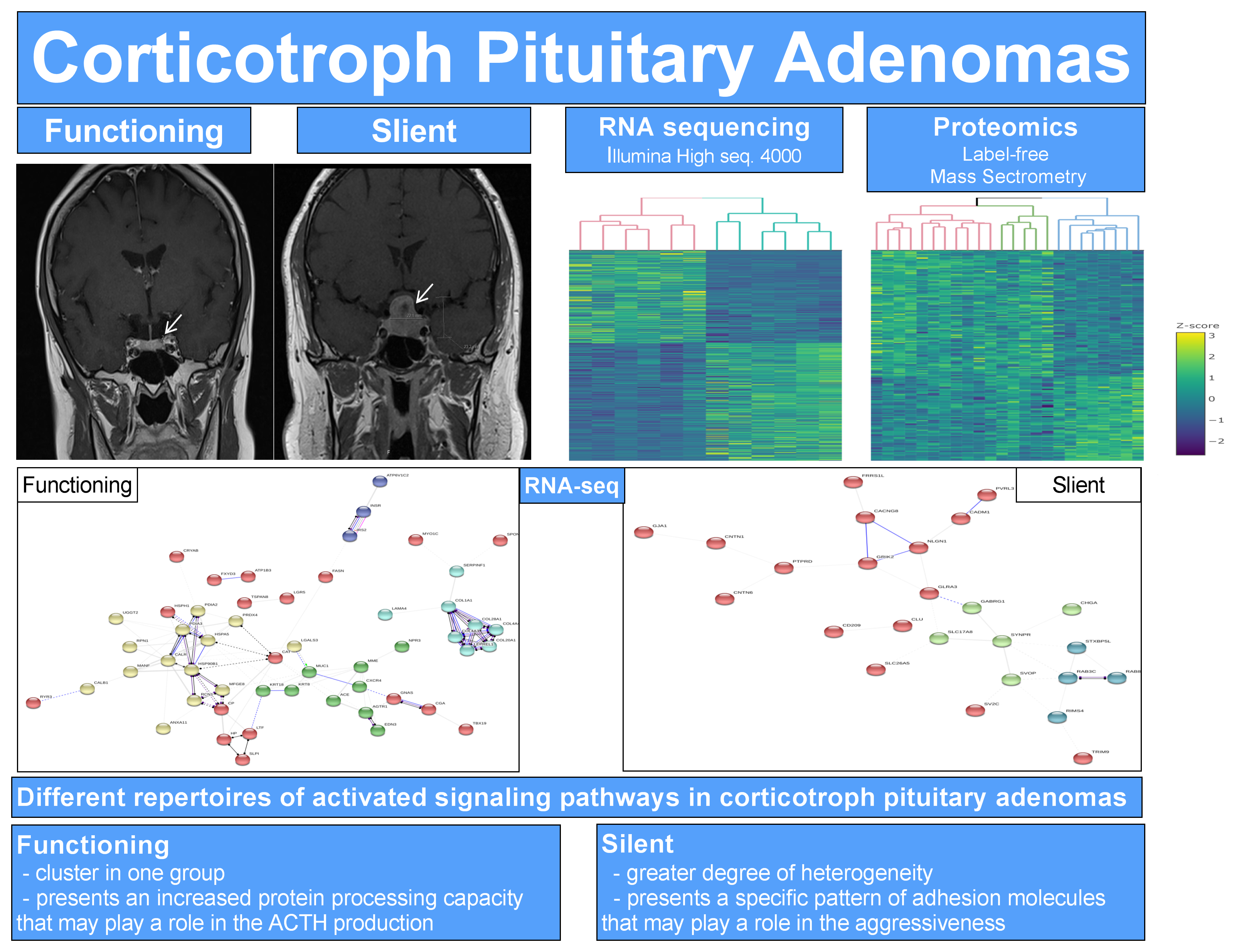
2. Results
2.1. Clinical Characteristics of Patients with FCA and SCA
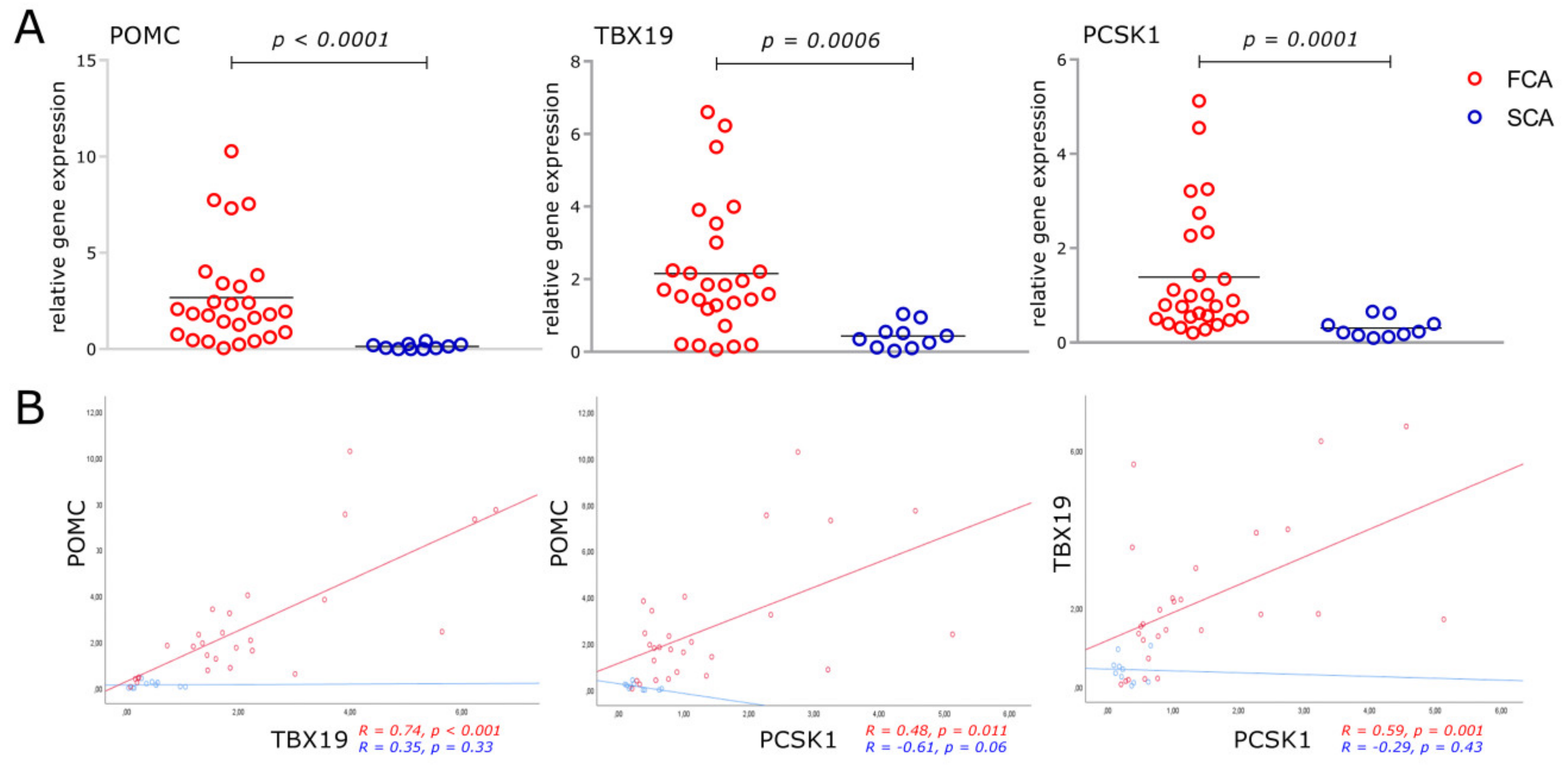
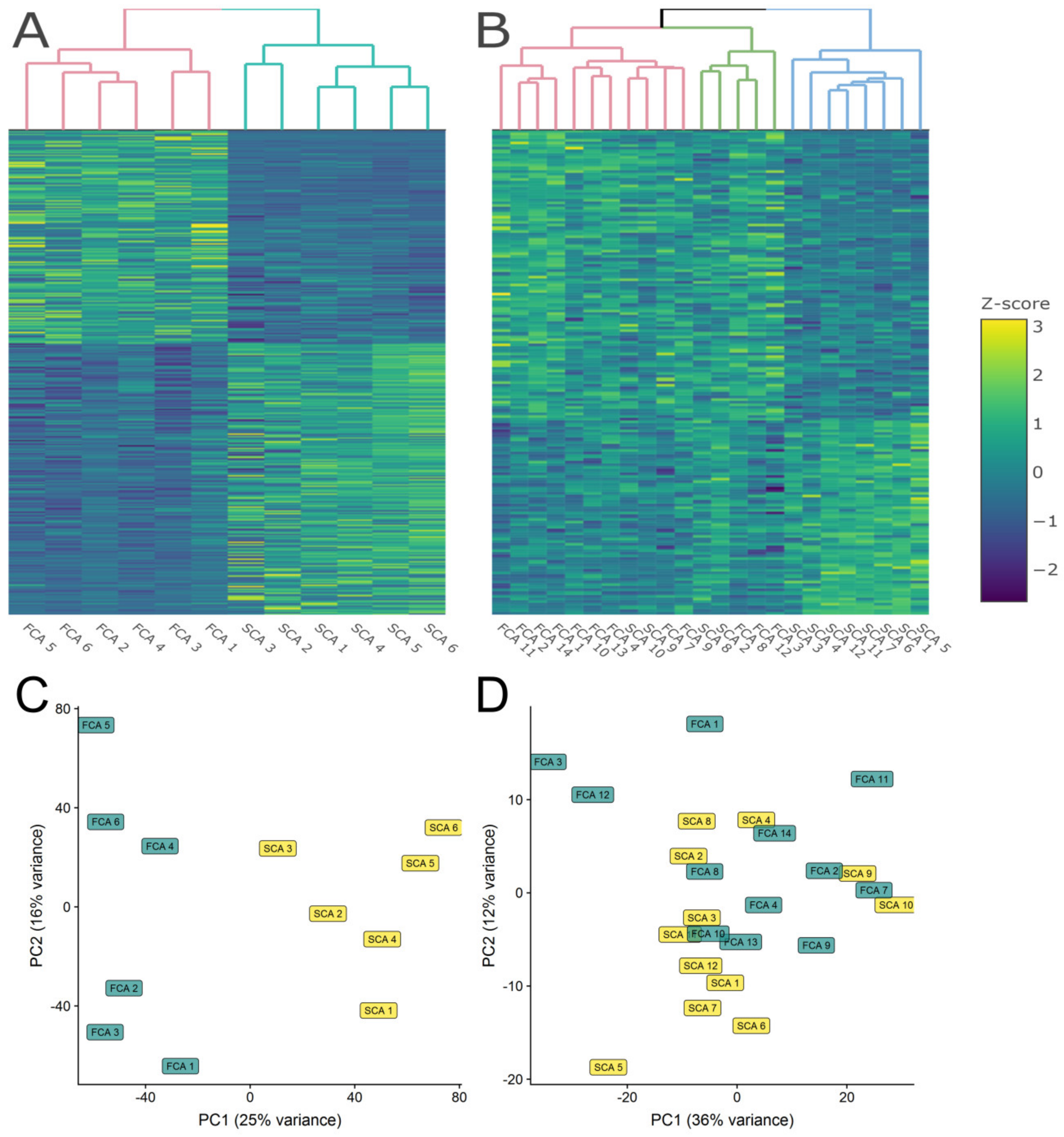
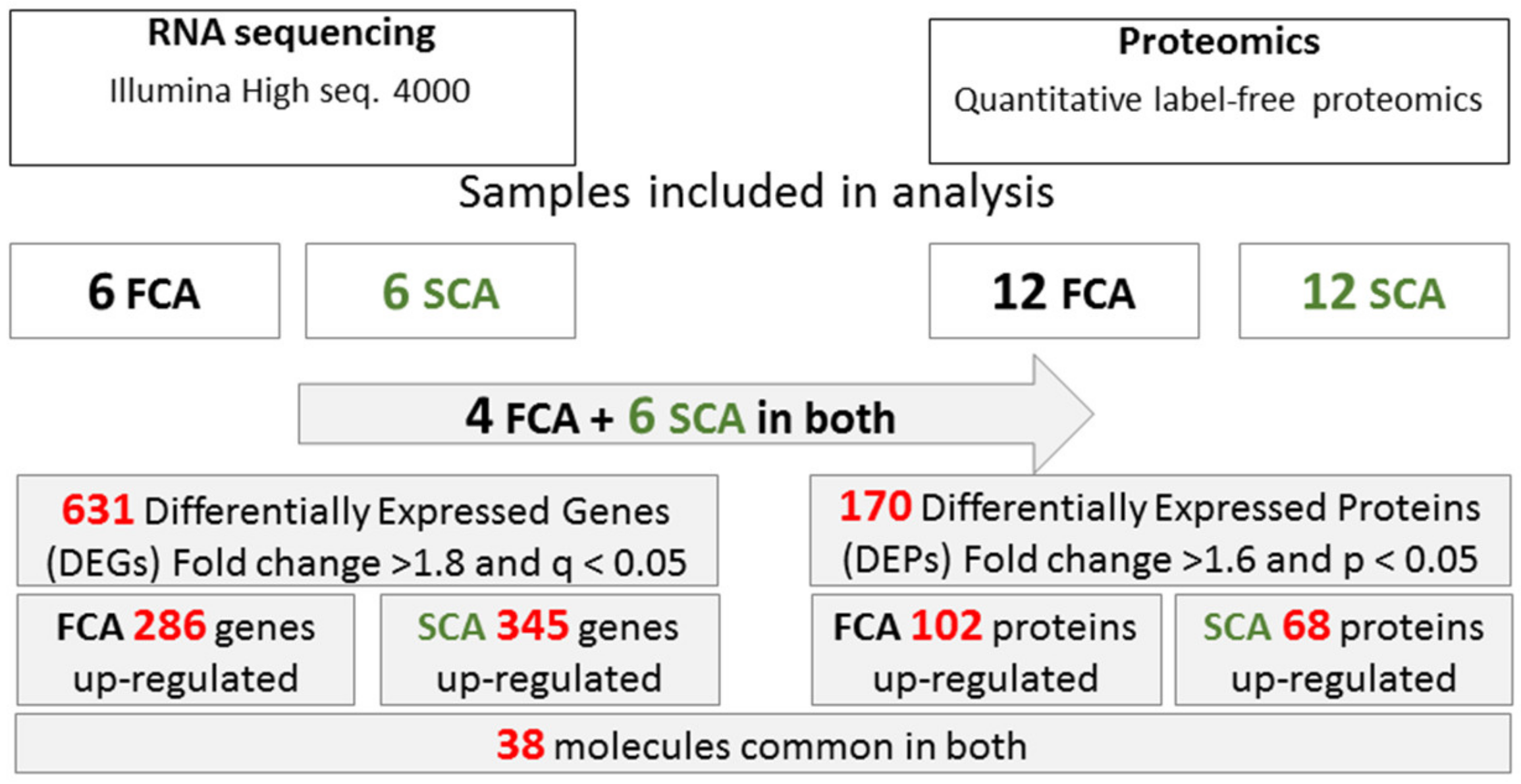
2.2. RNA-seq
2.3. Quantitative Label-Free Proteomics
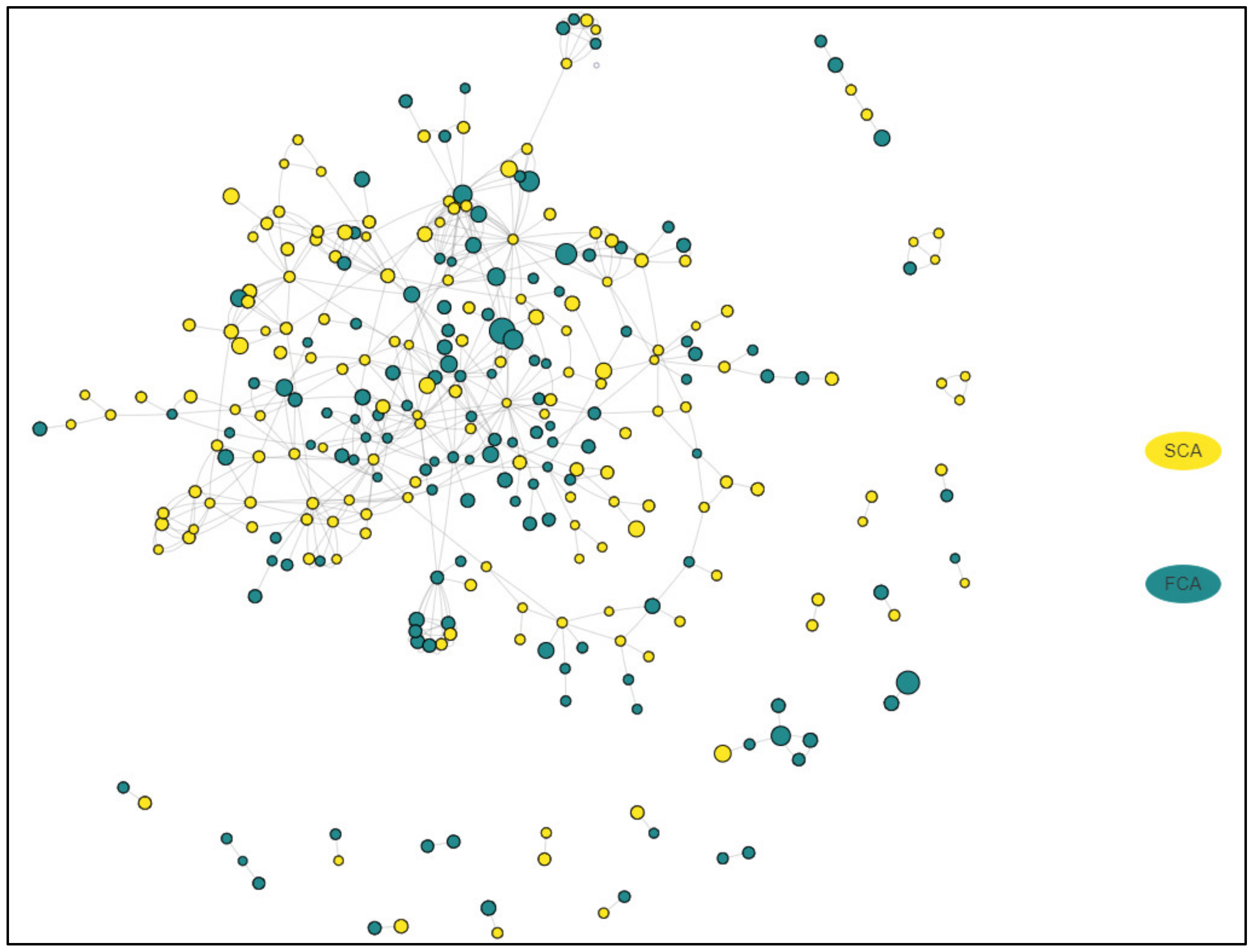
2.4. Common Molecules Presented in Transcriptomics and Proteomics Studies
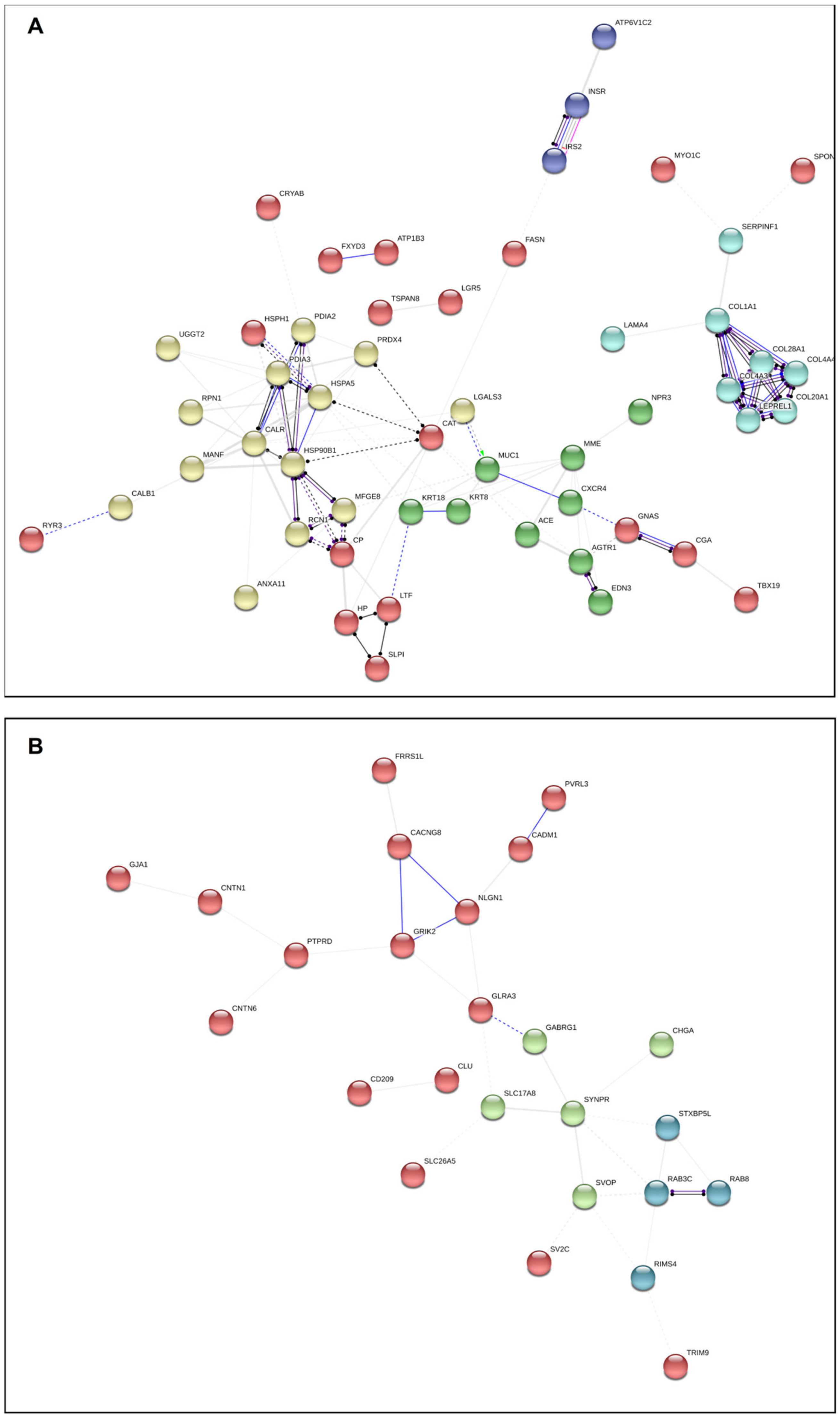
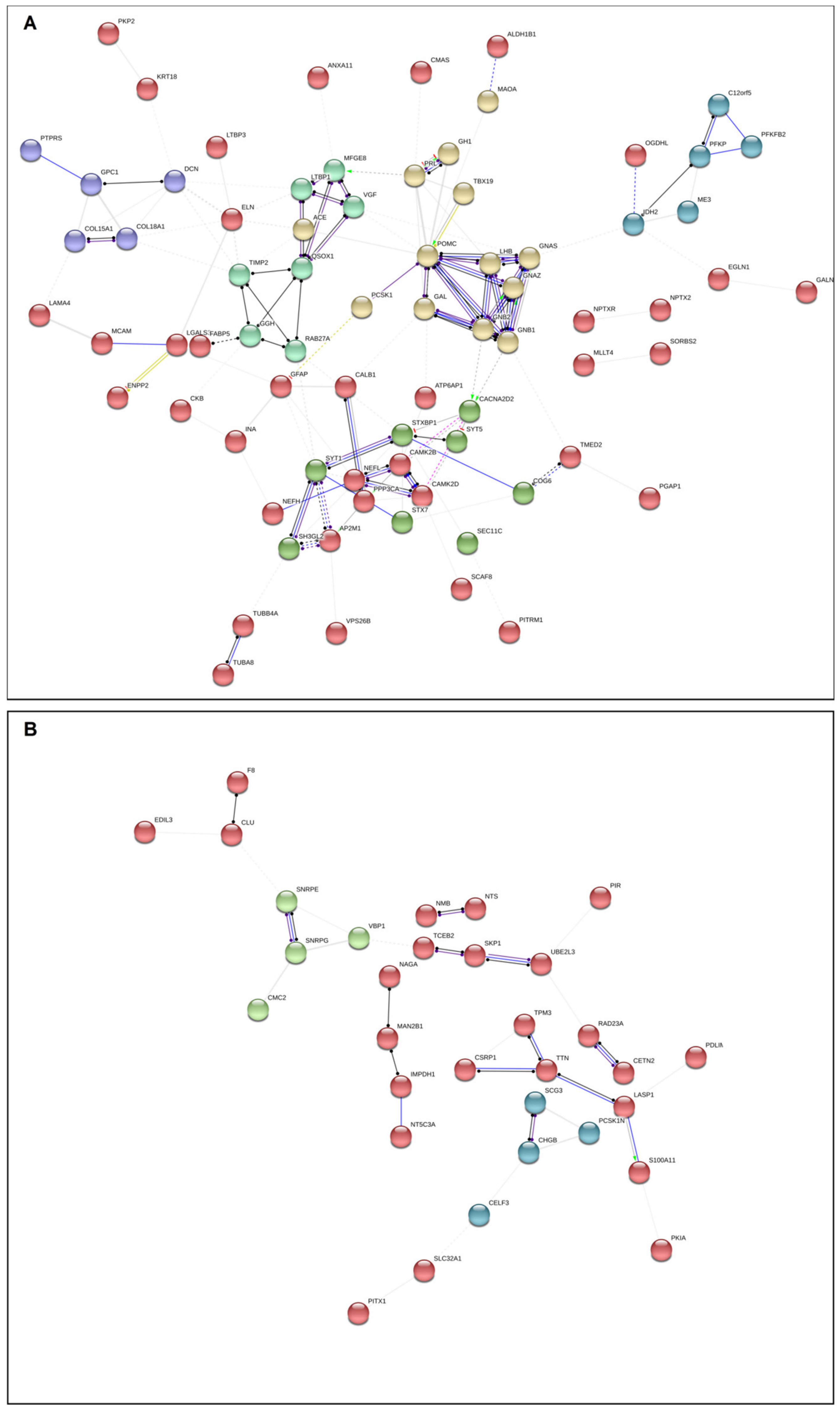
2.5. Network Analysis in FCA and SCA
3. Discussion
4. Material and Methods
4.1. Sample Selection
4.2. RNA Isolation and Reverse Transcription
4.3. RT-qPCR
4.4. RNA-Seq
4.5. Pre-Processing and Cleanup
4.6. Transcriptome Alignment
4.7. Quantitative Label-Free Mass Spectrometry-Based Proteomic Analysis
4.8. Pathways
4.9. Heatmap and PCA
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Agustsson, T.T.; Baldvinsdottir, T.; Jonasson, J.G.; Ólafsdóttir, E.; Steinthorsdottir, V.; Sigurdsson, G.; Thorsson, A.V.; Carroll, P.V.; Korbonits, M.; Benediktsson, R. The epidemiology of pituitary adenomas in Iceland, 1955–2012: A nationwide population-based study. Eur. J. Endocrinol. 2015, 173, 655–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tjörnstrand, A.; Gunnarsson, K.; Evert, M.; Holmberg, E.; Ragnarsson, O.; Rosén, T.; Nyström, H.F. The incidence rate of pituitary adenomas in western Sweden for the period 2001–2011. Eur. J. Endocrinol. 2014, 171, 519–526. [Google Scholar] [CrossRef] [Green Version]
- Cooper, O.; Melmed, S. Subclinical hyperfunctioning pituitary adenomas: The silent tumors. Best Pr. Res. Clin. Endocrinol. Metab. 2012, 26, 447–460. [Google Scholar] [CrossRef] [Green Version]
- Ntali, G.; Capatina, C.; Fazal-Sanderson, V.; Byrne, J.V.; Cudlip, S.; Grossman, A.; Wass, J.A.; Karavitaki, N. Mortality in patients with non-functioning pituitary adenoma is increased: Systematic analysis of 546 cases with long follow-up. Eur. J. Endocrinol. 2016, 174, 137–145. [Google Scholar] [CrossRef] [Green Version]
- Drummond, J.; Roncaroli, F.; Grossman, A.B.; Korbonits, M. Clinical and Pathological Aspects of Silent Pituitary Adenomas. J. Clin. Endocrinol. Metab. 2018, 104, 2473–2489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karavitaki, N. Prevalence and incidence of pituitary adenomas. Ann. d’Endocrinologie 2012, 73, 79–80. [Google Scholar] [CrossRef] [PubMed]
- Raappana, A.; Koivukangas, J.; Ebeling, T.; Pirilä, T. Incidence of Pituitary Adenomas in Northern Finland in 1992–2007. J. Clin. Endocrinol. Metab. 2010, 95, 4268–4275. [Google Scholar] [CrossRef]
- Cohen-Inbar, O.; Xu, Z.; Lee, C.-C.; Wu, C.-C.; Chytka, T.; Silva, D.; Sharma, M.; Radwan, H.; Grills, I.S.; Nguyen, B.; et al. Prognostic significance of corticotroph staining in radiosurgery for non-functioning pituitary adenomas: A multicenter study. J. Neuro-Oncology 2017, 135, 67–74. [Google Scholar] [CrossRef]
- Cooper, O.; Ben-Shlomo, A.; Bonert, V.; Bannykh, S.; Mirocha, J.; Melmed, S. Silent Corticogonadotroph Adenomas: Clinical and Cellular Characteristics and Long-Term Outcomes. Horm. Cancer 2010, 1, 80–92. [Google Scholar] [CrossRef] [Green Version]
- Jahangiri, A.; Wagner, J.R.; Pekmezci, M.; Hiniker, A.; Chang, E.F.; Kunwar, S.; Blevins, L.; Aghi, M.K. A Comprehensive Long-term Retrospective Analysis of Silent Corticotrophic Adenomas vs Hormone-Negative Adenomas. Neurosurg. 2013, 73, 8–18. [Google Scholar] [CrossRef]
- Langlois, F.; Lim, D.S.T.; Yedinak, C.G.; Cetas, I.; McCartney, S.; Cetas, J.; Dogan, A.; Fleseriu, M. Predictors of silent corticotroph adenoma recurrence; a large retrospective single center study and systematic literature review. Pituit. 2017, 21, 32–40. [Google Scholar] [CrossRef]
- Pawlikowski, M.; Kunert-Radek, J.; Radek, M. “Silent” corticotropinoma. Neuro Endocrinol. Lett. 2008, 29, 347–350. [Google Scholar] [PubMed]
- Raverot, G.; Wierinckx, A.; Jouanneau, E.; Auger, C.; Borson-Chazot, F.; Lachuer, J.; Pugeat, M.; Trouillas, J. Clinical, hormonal and molecular characterization of pituitary ACTH adenomas without (silent corticotroph adenomas) and with Cushing’s disease. Eur. J. Endocrinol. 2010, 163, 35–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tateno, T.; Izumiyama, H.; Doi, M.; Yoshimoto, T.; Shichiri, M.; Inoshita, N.; Oyama, K.; Yamada, S.; Hirata, Y. Differential gene expression in ACTH -secreting and non-functioning pituitary tumors. Eur. J. Endocrinol. 2007, 157, 717–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben-Shlomo, A.; Cooper, O. Silent corticotroph adenomas. Pituit. 2018, 21, 183–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kojima, Y.; Suzuki, S.; Yamamura, K.; Ohhashi, G.; Yamamoto, I. Comparison of ACTH secretion in Cushing’s adenoma and clinically silent corticotroph adenoma by cell immunoblot assay. Endocr J. 2002, 49, 285–292. [Google Scholar] [CrossRef] [Green Version]
- Tateno, T.; Kato, M.; Tani, Y.; Yoshimoto, T.; Oki, Y.; Hirata, Y. Processing of high-molecular-weight form adrenocorticotropin in human adrenocorticotropin-secreting tumor cell line (DMS-79) after transfection of prohormone convertase 1/3 gene. J. Endocrinol. Investig. 2009, 33, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Parvin, R.; Saito-Hakoda, A.; Shimada, H.; Shimizu, K.; Noro, E.; Iwasaki, Y.; Fujiwara, K.; Yokoyama, A.; Sugawara, A. Role of NeuroD1 on the negative regulation of Pomc expression by glucocorticoid. PLoS ONE 2017, 12, e0175435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harno, E.; Ramamoorthy, T.G.; Coll, A.P.; White, A. POMC: The Physiological Power of Hormone Processing. Physiol. Rev. 2018, 98, 2381–2430. [Google Scholar] [CrossRef] [PubMed]
- Tabuchi, Y.; Kitamura, T.; Fukuhara, A.; Mukai, K.; Onodera, T.; Miyata, Y.; Hamasaki, T.; Oshino, S.; Saitoh, Y.; Morii, E.; et al. Nur77 gene expression levels were involved in different ACTH-secretion autonomy between Cushing’s disease and subclinical Cushing’s disease. Endocr. J. 2016, 63, 545–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pendleton, K.E.; Chen, B.; Liu, K.; Hunter, O.V.; Xie, Y.; Tu, B.P.; Conrad, N.K. The U6 snRNA m 6 A Methyltransferase METTL16 Regulates SAM Synthetase Intron Retention. Cell 2017, 169, 824–835.e14. [Google Scholar] [CrossRef] [Green Version]
- Reyes, R.; Valladares, F.; Díaz-Flores, L.; Feria, L.; Alonso, R.; Tramu, G.; Bello, A.R.; Reyes, R. Immunohistochemical localization of hormones and peptides in the human pituitary cells in a case of hypercortisolism by ACTH secreting microadenoma. Histol. Histopathol. 2007, 22, 709–717. [Google Scholar] [PubMed]
- Olarescu, N.C.; Normann, K.R.; Sundaram, A.Y.; Oystese, K.A.B.; Eieland, A.; Lekva, T.; Bollerslev, J. Pathway enrichment analysis in functioning and silent corticotroph pituitary adenomas reveals mechanisms to explain their distinct clinical behaviour. Endocr. Abstr. 2019, 63 GP154. [Google Scholar] [CrossRef]
- Mertens, F.; Gremeaux, L.; Chen, J.; Fu, Q.; Willems, C.; Roose, H.; Govaere, O.; Roskams, T.; Cristina, C.; Becú-Villalobos, D.; et al. Pituitary tumors contain a side population with tumor stem cell-associated characteristics. Endocr. Relat. Cancer 2015, 22, 481–504. [Google Scholar] [CrossRef]
- Stout, K.A.; Dunn, A.R.; Hoffman, C.; Miller, G.W. The Synaptic Vesicle Glycoprotein 2: Structure, Function, and Disease Relevance. ACS Chem. Neurosci. 2019, 10, 3927–3938. [Google Scholar] [CrossRef]
- García-Martínez, A.; Cano, D.A.; Flores-Martínez, Á.; Gil, J.; Domingo, M.P.; Webb, S.M.; Soto-Moreno, A.; Pico, A.M.; Soto, A. Why don’t corticotroph tumors always produce Cushing’s disease? Eur. J. Endocrinol. 2019, 181, 351–361. [Google Scholar] [CrossRef] [PubMed]
- Pulichino, A.-M.; Vallette-Kasic, S.; Tsai, J.P.-Y.; Couture, C.; Gauthier, Y.; Drouin, J. Tpit determines alternate fates during pituitary cell differentiation. Genes Dev. 2003, 17, 738–747. [Google Scholar] [CrossRef] [Green Version]
- Fortenberry, Y.; Hwang, J.-R.; Apletalina, E.V.; Lindberg, I. Functional Characterization of ProSAAS. J. Biol. Chem. 2001, 277, 5175–5186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griss, J.; Viteri, G.; Sidiropoulos, K.; Nguyen, V.; Fabregat, A.; Hermjakob, H. ReactomeGSA–Efficient Multi-Omics Comparative Pathway Analysis. Mol. Cell. Proteom. 2020. [Google Scholar] [CrossRef] [PubMed]
- Cassarino, M.F.; Ambrogio, A.G.; Cassarino, A.; Terreni, M.R.; Gentilini, D.; Sesta, A.; Cavagnini, F.; Losa, M.; Giraldi, F.P. Gene expression profiling in human corticotroph tumours reveals distinct, neuroendocrine profiles. J. Neuroendocr. 2018, 30, e12628. [Google Scholar] [CrossRef]
- Khetchoumian, K.; Balsalobre, A.; Mayran, A.; Christian, H.; Chénard, V.; St-Pierre, J.; Drouin, J. Pituitary cell translation and secretory capacities are enhanced cell autonomously by the transcription factor Creb3l2. Nat. Commun. 2019, 10, 3960. [Google Scholar] [CrossRef] [Green Version]
- Cakir, I.; Nillni, E.A. Endoplasmic Reticulum Stress, the Hypothalamus, and Energy Balance. Trends Endocrinol. Metab. 2019, 30, 163–176. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, P.A.; Smiljanic, K.; Prévide, R.M.; Iben, J.R.; Li, T.; Rokic, M.B.; Sherman, A.; Coon, S.L.; Stojilkovic, S.S. Cell Type- and Sex-Dependent Transcriptome Profiles of Rat Anterior Pituitary Cells. Front. Endocrinol. 2019, 10, 623. [Google Scholar] [CrossRef] [PubMed]
- Cheung, L.Y.M.; George, A.S.; McGee, S.R.; Daly, A.Z.; Brinkmeier, M.L.; Ellsworth, B.S.; Camper, S.A. Single-Cell RNA Sequencing Reveals Novel Markers of Male Pituitary Stem Cells and Hormone-Producing Cell Types. Endocrinology 2018, 159, 3910–3924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mete, O.; Hayhurst, C.; Alahmadi, H.; Monsalves, E.; Gücer, H.; Gentili, F.; Ezzat, S.; Asa, S.L.; Zadeh, G. The Role of Mediators of Cell Invasiveness, Motility, and Migration in the Pathogenesis of Silent Corticotroph Adenomas. Endocr. Pathol. 2013, 24, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Di Ieva, A.; Davidson, J.M.; Syro, L.V.; Rotondo, F.; Montoya, J.F.; Horvath, E.; Cusimano, M.D.; Kovacs, K. Crooke’s Cell Tumors of the Pituitary. Neurosurg. 2015, 76, 616–622. [Google Scholar] [CrossRef]
- Kuchenbauer, F.; Hopfner, U.; Stalla, J.; Arzt, E.; Stalla, G.K.; Paez-Pereda, M. Extracellular matrix components regulate ACTH production and proliferation in corticotroph tumor cells. Mol. Cell. Endocrinol. 2001, 175, 141–148. [Google Scholar] [CrossRef]
- Øystese, K.A.; Berg, J.P.; Normann, K.R.; Zucknick, M.; Casar-Borota, O.; Bollerslev, J. The role of E and N-cadherin in the postoperative course of gonadotroph pituitary tumours. Endocr. 2018, 62, 351–360. [Google Scholar] [CrossRef]
- Falch, C.M.; Sundaram, A.Y.; Øystese, K.A.; Normann, K.R.; Lekva, T.; Silamikelis, I.; Eieland, A.K.; Anderson, M.; Bollerslev, J.; Olarescu, N.C. Gene expression profiling of fast- and slow-growing non-functioning gonadotroph pituitary adenomas. Eur. J. Endocrinol. 2018, 178, 295–307. [Google Scholar] [CrossRef] [Green Version]
- Lekva, T.; Berg, J.P.; Fougner, S.L.; Olstad, O.K.; Ueland, T.; Bollerslev, J. Gene Expression Profiling Identifies ESRP1 as a Potential Regulator of Epithelial Mesenchymal Transition in Somatotroph Adenomas from a Large Cohort of Patients with Acromegaly. J. Clin. Endocrinol. Metab. 2012, 97, E1506–E1514. [Google Scholar] [CrossRef] [Green Version]
- Evang, J.A.; Berg, J.P.; Casar-Borota, O.; Lekva, T.; Kringen, M.K.; Ramm-Pettersen, J.; Bollerslev, J. Reduced levels of E-cadherin correlate with progression of corticotroph pituitary tumours. Clin. Endocrinol. 2011, 75, 811–818. [Google Scholar] [CrossRef] [PubMed]
- Neou, M.; Villa, C.; Armignacco, R.; Jouinot, A.; Raffin-Sanson, M.-L.; Septier, A.; Letourneur, F.; Diry, S.; Diedisheim, M.; Izac, B.; et al. Pangenomic Classification of Pituitary Neuroendocrine Tumors. Cancer Cell 2020, 37, 123–134.e5. [Google Scholar] [CrossRef] [PubMed]
- Napoli, J.L. Cellular retinoid binding-proteins, CRBP, CRABP, FABP5: Effects on retinoid metabolism, function and related diseases. Pharmacol. Ther. 2017, 173, 19–33. [Google Scholar] [CrossRef] [Green Version]
- Nieto, L.; Fuertes, M.; Rosmino, J.; Senin, S.; Arzt, E. Crosstalk of BMP-4 and RA signaling pathways on Pomc gene regulation in corticotrophs. J. Mol. Endocrinol. 2019, 63, 161–174. [Google Scholar] [CrossRef] [PubMed]
- Theodoropoulou, M.; Reincke, M. Tumor-Directed Therapeutic Targets in Cushing Disease. J. Clin. Endocrinol. Metab. 2018, 104, 925–933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez-Castro, C.; Renner, U.; Haedo, M.R.; Stalla, G.K.; Arzt, E. Cellular and Molecular Specificity of Pituitary Gland Physiology. Physiol. Rev. 2012, 92, 1–38. [Google Scholar] [CrossRef] [Green Version]
- Murakami, I.; Takeuchi, S.; Kudo, T.; Sutou, S.; Takahashi, S. Corticotropin-releasing hormone or dexamethasone regulates rat proopiomelanocortin transcription through Tpit/Pitx-responsive element in its promoter. J. Endocrinol. 2007, 193, 279–290. [Google Scholar] [CrossRef] [Green Version]
- Caimari, F.; Korbonits, M. Novel Genetic Causes of Pituitary Adenomas. Clin. Cancer Res. 2016, 22, 5030–5042. [Google Scholar] [CrossRef] [Green Version]
- Jin, L.; Riss, D.; Ruebel, K.; Kajita, S.; Scheithauer, B.W.; Horvath, E.; Kovacs, K.; Lloyd, R.V. Galectin-3 Expression in Functioning and Silent ACTH-Producing Adenomas. Endocr. Pathol. 2005, 16, 107–114. [Google Scholar] [CrossRef]
- Thodou, E.; Argyrakos, T.; Kontogeorgos, G. Galectin-3 as a marker distinguishing functioning from silent corticotroph adenomas. Horm. 2007, 6, 227–232. [Google Scholar]
- Ponnusamy, M.P.; Seshacharyulu, P.; Lakshmanan, I.; Vaz, A.P.; Chugh, S.; Batra, S.K. Emerging role of mucins in epithelial to mesenchymal transition. Curr. Cancer Drug Targets 2013, 13, 945–956. [Google Scholar] [CrossRef] [Green Version]
- Mori, Y.; Akita, K.; Yashiro, M.; Sawada, T.; Hirakawa, K.; Murata, T.; Nakada, H. Binding of Galectin-3, a β-Galactoside-binding Lectin, to MUC1 Protein Enhances Phosphorylation of Extracellular Signal-regulated Kinase 1/2 (ERK1/2) and Akt, Promoting Tumor Cell Malignancy*. J. Boil. Chem. 2015, 290, 26125–26140. [Google Scholar] [CrossRef] [Green Version]
- Leung, B.; Lismaa, T.P.; Hort, Y.J.; Turner, J.; Sheehy, J.P.; Ho, K.K.Y. Galanin in human pituitary adenomas: Frequency and clinical significance. Clin. Endocrinol. 2002, 56, 397–403. [Google Scholar] [CrossRef]
- Grenbäck, E.; Bjellerup, P.; Wallerman, E.; Lundblad, L.; Anggård, A.; Ericson, K.; Aman, K.; Landry, M.; Schmidt, W.E.; Hökfelt, T.; et al. Galanin in pituitary adenomas. Regul. Pept. 2004, 117, 127–139. [Google Scholar] [CrossRef] [PubMed]
- Ibañez-Costa, A.; Rivero-Cortés, E.; Vázquez-Borrego, M.C.; Gahete, M.D.; Jiménez-Reina, L.; Venegas-Moreno, E.; De La Riva, A.; Arráez, M.Á.; González-Molero, I.; Schmid, H.A.; et al. Octreotide and pasireotide (dis)similarly inhibit pituitary tumor cells in vitro. J. Endocrinol. 2016, 231, 135–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, J.; Song, H.; Wang, X.; Shen, Y.; Chen, F.; Liu, Y.; Li, S.; Wang, Y.; Shou, X.; Zhang, Y.; et al. Gene expression profiling in human null cell pituitary adenoma tissue. Pituit. 2007, 10, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Hamzehzadeh, L.; Caraglia, M.; Atkin, S.L.; Sahebkar, A. Dickkopf homolog 3 (DKK3): A candidate for detection and treatment of cancers? J. Cell. Physiol. 2018, 233, 4595–4605. [Google Scholar] [CrossRef]
- Habuta, M.; Fujita, H.; Sato, K.; Bando, T.; Inoue, J.; Kondo, Y.; Miyaishi, S.; Kumon, H.; Ohuchi, H. Dickkopf3 (Dkk3) is required for maintaining the integrity of secretory vesicles in the mouse adrenal medulla. Cell and Tissue Res. 2019, 379, 157–167. [Google Scholar] [CrossRef]
- Sjöstedt, E.; Bollerslev, J.; Mulder, J.; Lindskog, C.; Pontén, F.; Casar-Borota, O. A specific antibody to detect transcription factor T-Pit: A reliable marker of corticotroph cell differentiation and a tool to improve the classification of pituitary neuroendocrine tumours. Acta Neuropathol. 2017, 134, 675–677. [Google Scholar] [CrossRef]
- Øystese, K.A.; Casar-Borota, O.; Normann, K.R.; Zucknick, M.; Berg, J.P.; Bollerslev, J. Estrogen Receptor α, a Sex-Dependent Predictor of Aggressiveness in Nonfunctioning Pituitary Adenomas: SSTR and Sex Hormone Receptor Distribution in NFPA. J. Clin. Endocrinol. Metab. 2017, 102, 3581–3590. [Google Scholar] [CrossRef] [Green Version]
- Kolnes, A.J.; Øystese, K.A.; Dahlberg, D.; Berg–Johnsen, J.; Niehusmann, P.; Pahnke, J.; Bollerslev, J.; Jørgensen, A.P. Cut-off values for sufficient cortisol response to low dose Short Synacthen Test after surgery for non-functioning pituitary adenoma. Acta Neurochir. 2020, 162, 845–852. [Google Scholar] [CrossRef]
- Fleige, S.; Pfaffl, M.W. RNA integrity and the effect on the real-time qRT-PCR performance. Mol. Asp. Med. 2006, 27, 126–139. [Google Scholar] [CrossRef]
- Normann, K.R.; Øystese, K.A.; Berg, J.P.; Lekva, T.; Berg-Johnsen, J.; Bollerslev, J.; Olarescu, N.C. Selection and validation of reliable reference genes for RT-qPCR analysis in a large cohort of pituitary adenomas. Mol. Cell. Endocrinol. 2016, 437, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinform. 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. TopHat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013, 14, R36-13. [Google Scholar] [CrossRef] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goff, L.A.; Trapnell, C.; Kelley, D. CummeRbund: Visualization and Exploration of Cufflinks High-throughput Sequencing Data. Available online: https://bioconductor.statistik.tu-dortmund.de/packages/3.1/bioc/vignettes/cummeRbund/inst/doc/cummeRbund-manual.pdf (accessed on 28 June 2018).
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein–protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2018, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, P.D.; Campbell, M.J.; Kejariwal, A.; Mi, H.; Karlak, B.; Daverman, R.; Diemer, K.; Muruganujan, A.; Narechania, A. PANTHER: A Library of Protein Families and Subfamilies Indexed by Function. Genome Res. 2003, 13, 2129–2141. [Google Scholar] [CrossRef] [Green Version]
- Galili, T.; O’Callaghan, A.; Sidi, J.; Sievert, C. heatmaply: An R package for creating interactive cluster heatmaps for online publishing. Bioinform. 2017, 34, 1600–1602. [Google Scholar] [CrossRef]
- Sievert, C. Interactive Web-Based Data Visualization with R., Plotly, and Shiny; Chapman and Hall/CRC: Boca Raton, FL, USA, 2019; ISBN 9781138331457. Available online: https://plotly-r.com/ (accessed on 12 July 2019).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Short Name | Gene Description | Cell Adhesion | RNA-seq Fold Change | Proteomics Fold Change |
|---|---|---|---|---|
| GH1 | growth hormone 1 | 1982.1 | 18.1 | |
| PRL | prolactin | 176.1 | 19.8 | |
| POMC | proopiomelanocortin | 33.3 | 9.7 | |
| LGALS3 | galectin 3 | X | 22.8 | 7.7 |
| ELN | elastin | 14.3 | 1.9 | |
| KRT18 | keratin 18 | X | 13.1 | 9.6 |
| CACNA2D2 | calcium voltage-gated channel auxiliary subunit alpha2delta 2 | 8.7 | 3.5 | |
| ENPP2 | ectonucleotide pyrophosphatase/phosphodiesterase 2 | X | 6.0 | 5.2 |
| DKK3 | dickkopf WNT signaling pathway inhibitor 3 | 5.4 | 7.6 | |
| ANXA11 | annexin A11 | 5.1 | 3.3 | |
| LAMA4 | laminin subunit alpha 4 | X | 4.9 | 2.6 |
| MAOA | monoamine oxidase A | 4.5 | 2.9 | |
| MFGE8 | milk fat globule-EGF factor 8 protein | 4.5 | 9.0 | |
| ACE | angiotensin I converting enzyme | 4.4 | 2.7 | |
| DCN | decorin | X | 4.2 | 6.6 |
| CALB1 | calbindin 1 | 3.9 | 10.2 | |
| TBX19 | T-box 19 | 3.8 | 4.9 | |
| SORBS2 | sorbin and SH3 domain containing 2 | 3.3 | 3.4 | |
| KCTD12 | potassium channel tetramerization domain containing 12 | 2.7 | 2.5 | |
| APMAP | adipocyte plasma membrane associated protein | 2.6 | 1.9 | |
| LTBP3 | latent transforming growth factor beta binding protein 3 | 2.5 | 4.1 | |
| GNAS | GNAS complex locus | 2.3 | 1.9 | |
| MAT2A * | methionine adenosyltransferase 2A | -2.5 | 1.7 | |
| LASP1 | LIM and SH3 protein 1 | X | -2.5 | −2.4 |
| CAB39L | calcium binding protein 39 like | -2.7 | −3.1 | |
| CSRP1 | cysteine and glycine rich protein 1 | -3.0 | −2.9 | |
| QPCT | glutaminyl-peptide cyclotransferase | -3.1 | −3.5 | |
| CLU | clusterin | X | -3.9 | −3.5 |
| S100A11 | S100 calcium binding protein A11 | X | -4.2 | −5.7 |
| DBI | diazepam binding inhibitor, acyl-CoA binding protein | -4.5 | −4.2 | |
| PDLIM1 | PDZ and LIM domain 1 | X | -5.2 | −5.3 |
| FAM107B | family with sequence similarity 107 member B | -5.9 | −2.8 | |
| SLC32A1 | solute carrier family 32 member 1 | -7.2 | −4.5 | |
| GPC4 | glypican 4 | X | -8.6 | −3.0 |
| EDIL3 | EGF like repeats and discoidin domains 3 | X | -8.7 | −6.7 |
| CRABP2 | cellular retinoic acid binding protein 2 | -9.9 | −8.7 | |
| LAMC2 | laminin subunit gamma 2 | X | -12.9 | −9.6 |
| NTS | neurotensin | -45.6 | −46.6 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eieland, A.K.; Normann, K.R.; Sundaram, A.Y.M.; Nyman, T.A.; Øystese, K.A.B.; Lekva, T.; Berg, J.P.; Bollerslev, J.; Olarescu, N.C. Distinct Pattern of Endoplasmic Reticulum Protein Processing and Extracellular Matrix Proteins in Functioning and Silent Corticotroph Pituitary Adenomas. Cancers 2020, 12, 2980. https://doi.org/10.3390/cancers12102980
Eieland AK, Normann KR, Sundaram AYM, Nyman TA, Øystese KAB, Lekva T, Berg JP, Bollerslev J, Olarescu NC. Distinct Pattern of Endoplasmic Reticulum Protein Processing and Extracellular Matrix Proteins in Functioning and Silent Corticotroph Pituitary Adenomas. Cancers. 2020; 12(10):2980. https://doi.org/10.3390/cancers12102980
Chicago/Turabian StyleEieland, Alexander K., Kjersti R. Normann, Arvind Y. M. Sundaram, Tuula A. Nyman, Kristin A. B. Øystese, Tove Lekva, Jens P. Berg, Jens Bollerslev, and Nicoleta C. Olarescu. 2020. "Distinct Pattern of Endoplasmic Reticulum Protein Processing and Extracellular Matrix Proteins in Functioning and Silent Corticotroph Pituitary Adenomas" Cancers 12, no. 10: 2980. https://doi.org/10.3390/cancers12102980
APA StyleEieland, A. K., Normann, K. R., Sundaram, A. Y. M., Nyman, T. A., Øystese, K. A. B., Lekva, T., Berg, J. P., Bollerslev, J., & Olarescu, N. C. (2020). Distinct Pattern of Endoplasmic Reticulum Protein Processing and Extracellular Matrix Proteins in Functioning and Silent Corticotroph Pituitary Adenomas. Cancers, 12(10), 2980. https://doi.org/10.3390/cancers12102980