Thioredoxin Interacting Protein (TXNIP) Is Differentially Expressed in Human Tumor Samples but Is Absent in Human Tumor Cell Line Xenografts: Implications for Its Use as an Immunosurveillance Marker




Simple Summary
Abstract
1. Introduction
2. Results
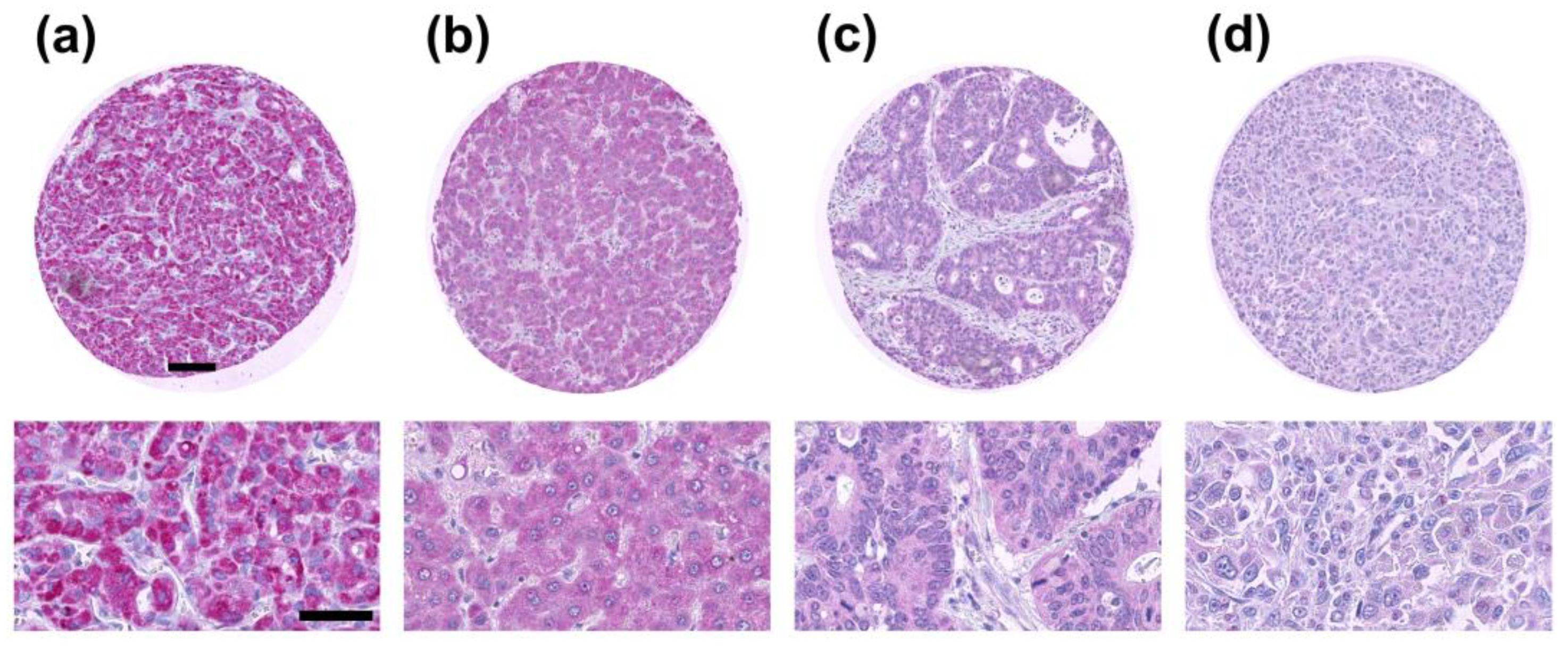
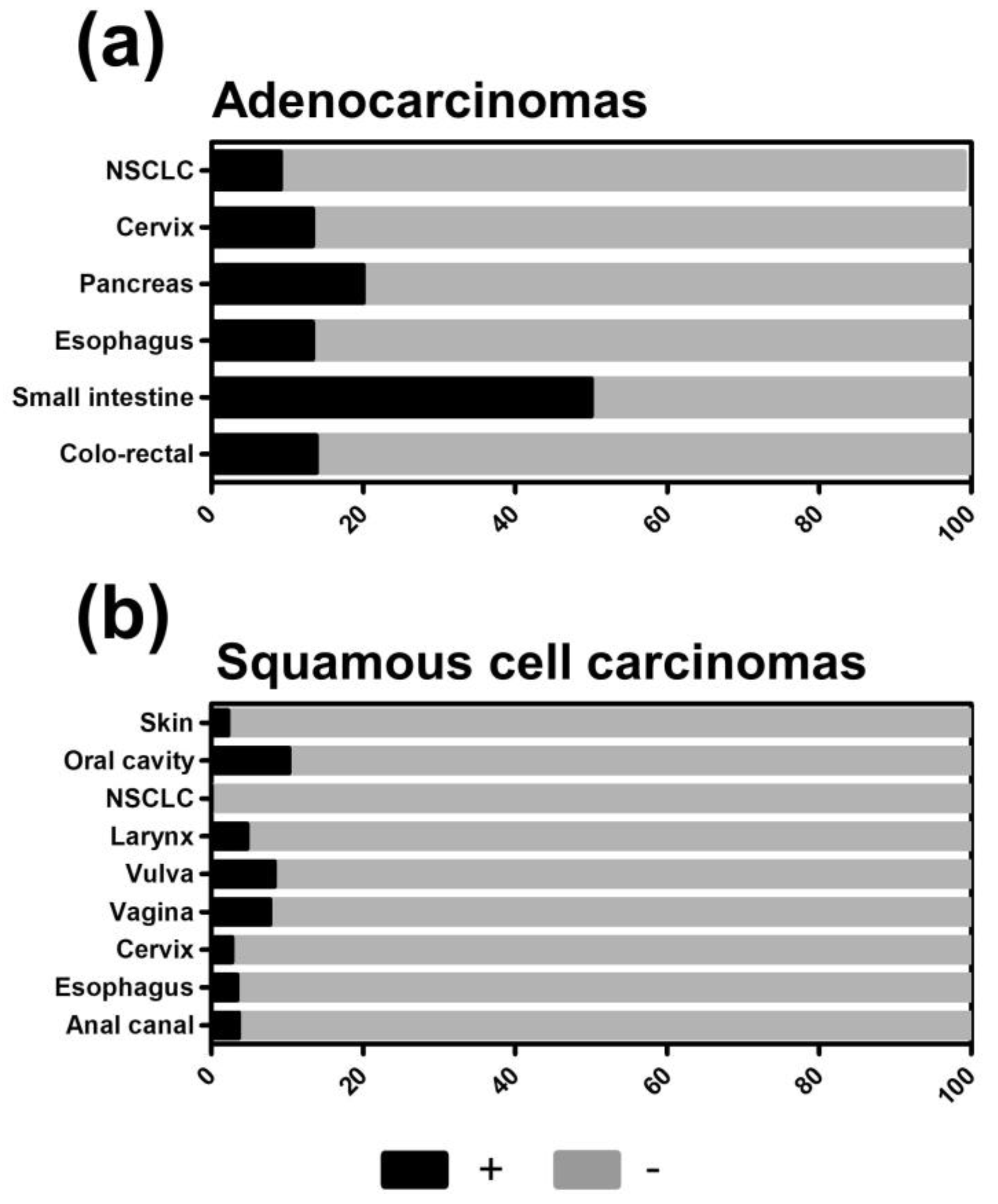
2.1. TXNIP Expression in Primary Human Tumor Samples
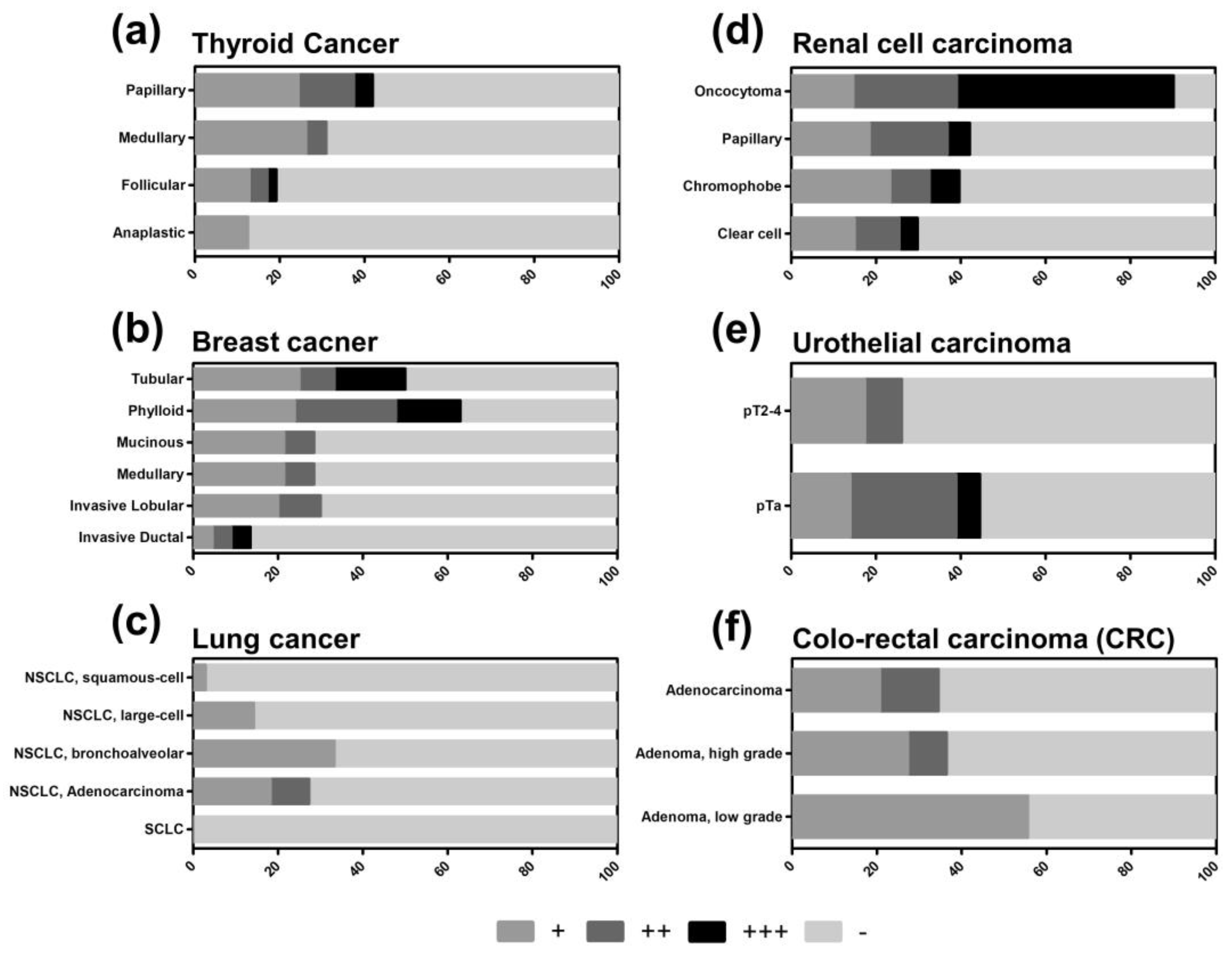
2.2. TXNIP is Differently Expressed in Tumor Subtypes
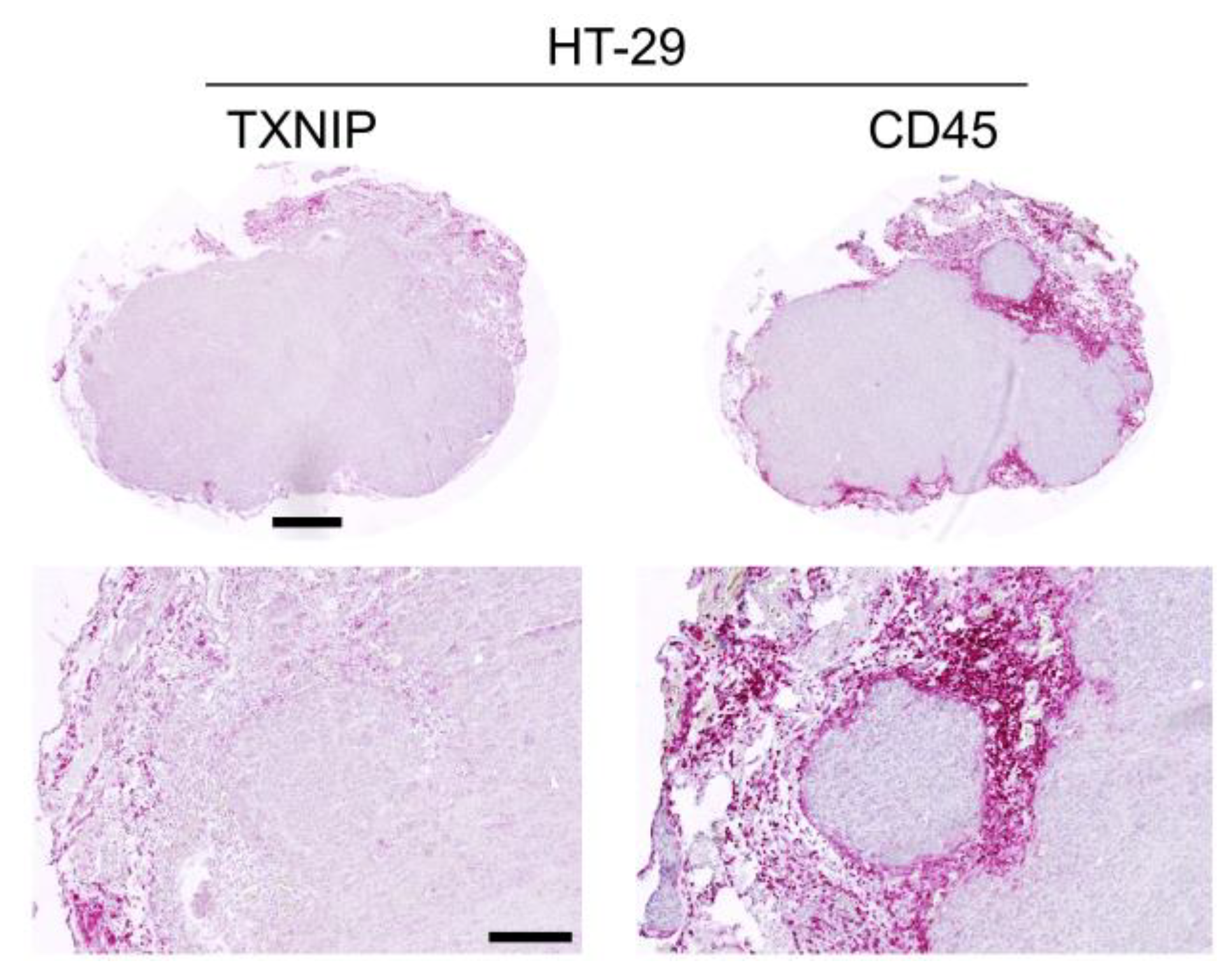
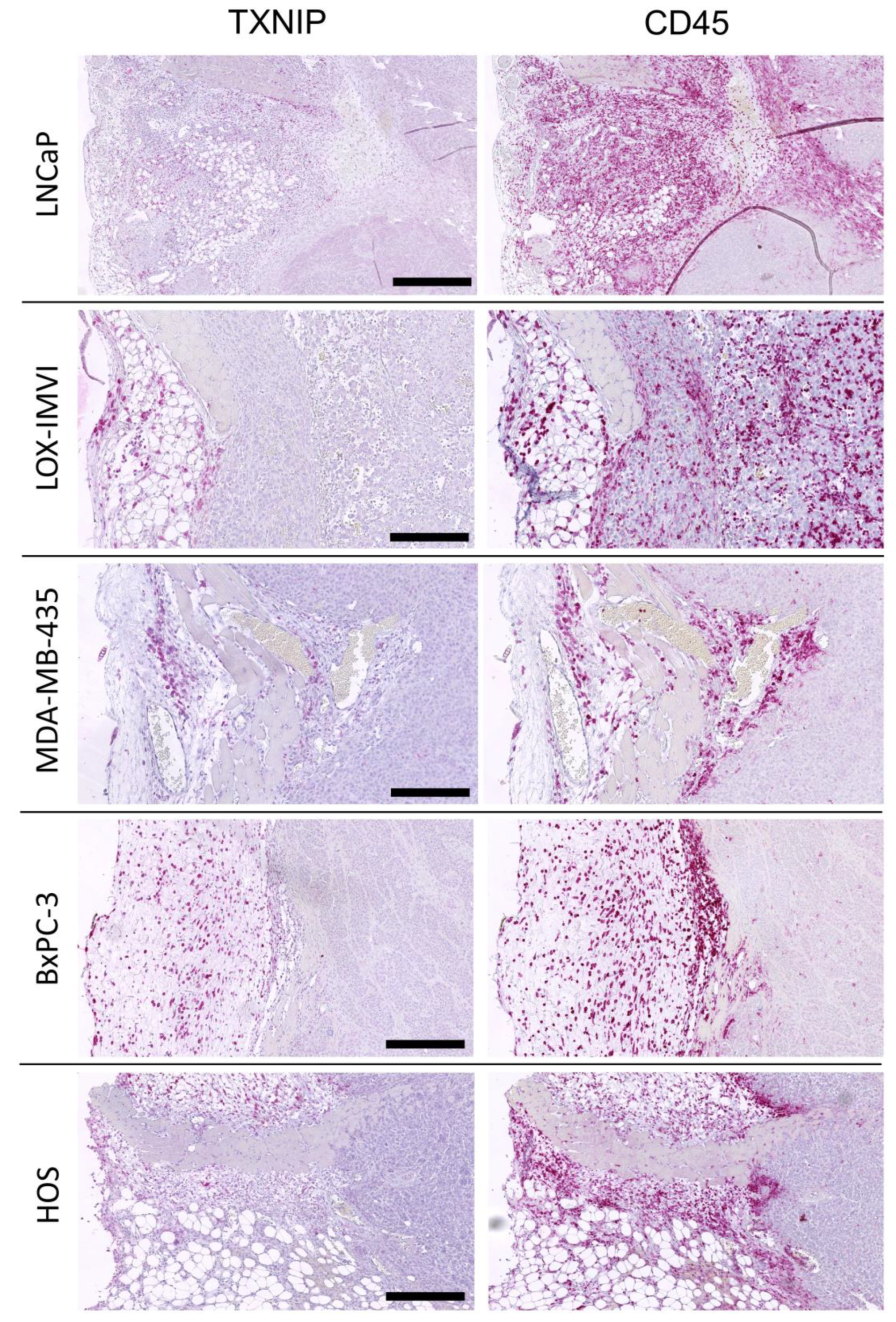
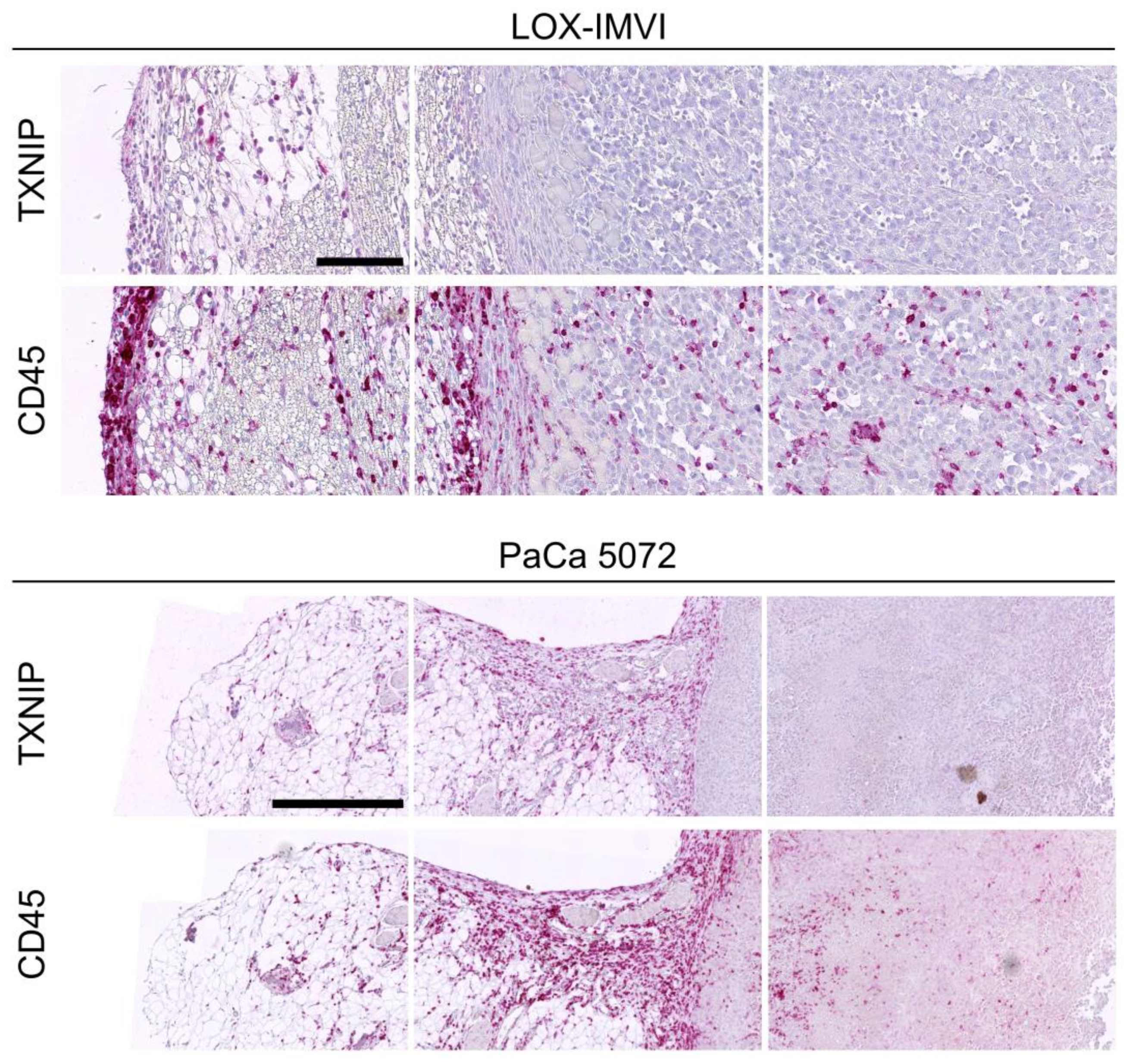
2.3. TXNIP Expression in Mouse Xenograft Tumors
2.4. TXNIP Expression in Tumor Infiltrating Leukocytes
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Cell Culture
4.2. Mouse Xenograft Tumors of Human Cancer Cell Lines
4.3. Multi-Tumor Tissue Microarray (TMA)
4.4. Fixation, Embedding and Sectioning of Cancer Cells and Xenograft Tumors
4.5. Immunohistochemistry (IHC)
4.6. Microscopy and Image Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hayes, J.D.; Dinkova-Kostova, A.T.; Tew, K.D. Oxidative Stress in Cancer. Cancer Cell 2020, 38, 167–197. [Google Scholar] [CrossRef]
- Cao, X.; He, W.; Pang, Y.; Cao, Y.; Qin, A. Redox-dependent and independent effects of thioredoxin interacting protein. Biol. Chem. 2020, 1. [Google Scholar] [CrossRef]
- Jia, J.-J.; Geng, W.-S.; Wang, Z.-Q.; Chen, L.; Zeng, X.-S. The role of thioredoxin system in cancer: Strategy for cancer therapy. Cancer Chemother. Pharmacol. 2019, 84, 453–470. [Google Scholar] [CrossRef]
- Chen, K.-S.; DeLuca, H.F. Isolation and characterization of a novel cDNA from HL-60 cells treated with 1,25-dihydroxyvitamin D-3. Biochim. Biophys. Acta BBA - Gene Struct. Expr. 1994, 1219, 26–32. [Google Scholar] [CrossRef]
- Ludwig, D.L.; Kotanides, H.; Le, T.; Chavkin, D.; Bohlen, P.; Witte, L. Cloning, genetic characterization, and chromosomal mapping of the mouse VDUP1 gene. Gene 2001, 269, 103–112. [Google Scholar] [CrossRef]
- Patwari, P.; Higgins, L.J.; Chutkow, W.A.; Yoshioka, J.; Lee, R.T. The Interaction of Thioredoxin with Txnip EVIDENCE FOR FORMATION OF A MIXED DISULFIDE BY DISULFIDE EXCHANGE. J. Biol. Chem. 2006, 281, 21884–21891. [Google Scholar] [CrossRef]
- Patwari, P.; Chutkow, W.A.; Cummings, K.; Verstraeten, V.L.R.M.; Lammerding, J.; Schreiter, E.R.; Lee, R.T. Thioredoxin-independent Regulation of Metabolism by the α-Arrestin Proteins. J. Biol. Chem. 2009, 284, 24996–25003. [Google Scholar] [CrossRef] [PubMed]
- Nishiyama, A.; Matsui, M.; Iwata, S.; Hirota, K.; Masutani, H.; Nakamura, H.; Takagi, Y.; Sono, H.; Gon, Y.; Yodoi, J. Identification of Thioredoxin-binding Protein-2/Vitamin D3 Up-regulated Protein 1 as a Negative Regulator of Thioredoxin Function and Expression. J. Biol. Chem. 1999, 274, 21645–21650. [Google Scholar] [CrossRef] [PubMed]
- Junn, E.; Han, S.H.; Im, J.Y.; Yang, Y.; Cho, E.W.; Um, H.D.; Kim, D.K.; Lee, K.W.; Han, P.L.; Rhee, S.G.; et al. Vitamin D3 Up-Regulated Protein 1 Mediates Oxidative Stress Via Suppressing the Thioredoxin Function. J. Immunol. 2000, 164, 6287–6295. [Google Scholar] [CrossRef]
- Han, S.H.; Jeon, J.H.; Ju, H.R.; Jung, U.; Kim, K.Y.; Yoo, H.S.; Lee, Y.H.; Song, K.S.; Hwang, H.M.; Na, Y.S.; et al. VDUP1 upregulated by TGF- β 1 and 1,25-dihydorxyvitamin D 3 inhibits tumor cell growth by blocking cell-cycle progression. Oncogene 2003, 22, 4035–4046. [Google Scholar] [CrossRef]
- Tobiume, K.; Matsuzawa, A.; Takahashi, T.; Nishitoh, H.; Morita, K.; Takeda, K.; Minowa, O.; Miyazono, K.; Noda, T.; Ichijo, H. ASK1 is required for sustained activations of JNK/p38 MAP kinases and apoptosis. EMBO Rep. 2001, 2, 222–228. [Google Scholar] [CrossRef] [PubMed]
- Saxena, G.; Chen, J.; Shalev, A. Intracellular Shuttling and Mitochondrial Function of Thioredoxin-interacting Protein. J. Biol. Chem. 2010, 285, 3997–4005. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.; Kim, M.J.; Kim, D.O.; Kim, W.S.; Yoon, S.-J.; Park, Y.-J.; Yoon, S.R.; Kim, T.-D.; Suh, H.-W.; Yun, S.; et al. TXNIP Maintains the Hematopoietic Cell Pool by Switching the Function of p53 under Oxidative Stress. Cell Metab. 2013, 18, 75–85. [Google Scholar] [CrossRef]
- Kwon, H.-J.; Won, Y.-S.; Suh, H.-W.; Jeon, J.-H.; Shao, Y.; Yoon, S.-R.; Chung, J.-W.; Kim, T.-D.; Kim, H.-M.; Nam, K.-H.; et al. Vitamin D3 Upregulated Protein 1 Suppresses TNF-α–Induced NF-κB Activation in Hepatocarcinogenesis. J. Immunol. 2010, 185, 3980–3989. [Google Scholar] [CrossRef]
- Yamaguchi, F.; Takata, M.; Kamitori, K.; Nonaka, M.; Dong, Y.; Sui, L.; Tokuda, M. Rare sugar D-allose induces specific up-regulation of TXNIP and subsequent G1 cell cycle arrest in hepatocellular carcinoma cells by stabilization of p27kip1. Int. J. Oncol. 2008, 32, 377–385. [Google Scholar] [CrossRef] [PubMed]
- Nishizawa, K.; Nishiyama, H.; Matsui, Y.; Kobayashi, T.; Saito, R.; Kotani, H.; Masutani, H.; Oishi, S.; Toda, Y.; Fujii, N.; et al. Thioredoxin-interacting protein suppresses bladder carcinogenesis. Carcinogenesis 2011, 32, 1459–1466. [Google Scholar] [CrossRef] [PubMed]
- Ungerstedt, J.S.; Sowa, Y.; Xu, W.-S.; Shao, Y.; Dokmanovic, M.; Perez, G.; Ngo, L.; Holmgren, A.; Jiang, X.; Marks, P.A. Role of thioredoxin in the response of normal and transformed cells to histone deacetylase inhibitors. Proc. Natl. Acad. Sci. 2005, 102, 673–678. [Google Scholar] [CrossRef]
- Lee, K.N.; Kang, H.-S.; Jeon, J.-H.; Kim, E.-M.; Yoon, S.-R.; Song, H.; Lyu, C.-Y.; Piao, Z.-H.; Kim, S.-U.; Han, Y.-H.; et al. VDUP1 Is Required for the Development of Natural Killer Cells. Immunity 2005, 22, 195–208. [Google Scholar] [CrossRef]
- Levring, T.B.; Kongsbak-Wismann, M.; Rode, A.K.O.; Al-Jaberi, F.A.H.; Lopez, D.V.; Met, Ö.; Woetmann, A.; Bonefeld, C.M.; Ødum, N.; Geisler, C. Tumor necrosis factor induces rapid down-regulation of TXNIP in human T cells. Sci. Rep. 2019, 9, 16725. [Google Scholar] [CrossRef]
- Zhou, R.; Tardivel, A.; Thorens, B.; Choi, I.; Tschopp, J. Thioredoxin-interacting protein links oxidative stress to inflammasome activation. Nat. Immunol. 2010, 11, 136–140. [Google Scholar] [CrossRef]
- Szpigel, A.; Hainault, I.; Carlier, A.; Venteclef, N.; Batto, A.-F.; Hajduch, E.; Bernard, C.; Ktorza, A.; Gautier, J.-F.; Ferré, P.; et al. Lipid environment induces ER stress, TXNIP expression and inflammation in immune cells of individuals with type 2 diabetes. Diabetologia 2018, 61, 399–412. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-K.; Choe, J.-Y.; Park, K.-Y. TXNIP-mediated nuclear factor-κB signaling pathway and intracellular shifting of TXNIP in uric acid-induced NLRP3 inflammasome. Biochem. Biophys. Res. Commun. 2019, 511, 725–731. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Chng, W.-J. Roles of thioredoxin binding protein (TXNIP) in oxidative stress, apoptosis and cancer. Mitochondrion 2013, 13, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Sherman, S.I.; Angelos, P.; Ball, D.W.; Byrd, D.; Clark, O.H.; Daniels, G.H.; Dilawari, R.A.; Ehya, H.; Farrar, W.B.; Gagel, R.F.; et al. Thyroid carcinoma. J. Natl. Compr. Cancer Netw. JNCCN 2007, 5, 568–621. [Google Scholar] [CrossRef]
- Chutkow, W.A.; Patwari, P.; Yoshioka, J.; Lee, R.T. Thioredoxin-interacting Protein (Txnip) Is a Critical Regulator of Hepatic Glucose Production. J. Biol. Chem. 2008, 283, 2397–2406. [Google Scholar] [CrossRef]
- Parikh, H.; Carlsson, E.; Chutkow, W.A.; Johansson, L.E.; Storgaard, H.; Poulsen, P.; Saxena, R.; Ladd, C.; Schulze, P.C.; Mazzini, M.J.; et al. TXNIP Regulates Peripheral Glucose Metabolism in Humans. PLOS Med. 2007, 4, e158. [Google Scholar] [CrossRef]
- Zhang, Y.; Huang, J.; Yang, X.; Sun, X.; Xu, Q.; Wang, B.; Zhong, P.; Wei, Z. Altered Expression of TXNIP in the peripheral leukocytes of patients with coronary atherosclerotic heart disease. Medicine (Baltimore) 2017, 96. [Google Scholar] [CrossRef]
- Gao, Y.; Qi, J.-C.; Li, X.; Sun, J.-P.; Ji, H.; Li, Q.-H. Decreased expression of TXNIP predicts poor prognosis in patients with clear cell renal cell carcinoma. Oncol. Lett. 2020, 19, 763–770. [Google Scholar] [CrossRef]
- Lim, J.Y.; Yoon, S.O.; Hong, S.W.; Kim, J.W.; Choi, S.H.; Cho, J.Y. Thioredoxin and thioredoxin-interacting protein as prognostic markers for gastric cancer recurrence. World J. Gastroenterol. WJG 2012, 18, 5581–5588. [Google Scholar] [CrossRef]
- Cadenas, C.; Franckenstein, D.; Schmidt, M.; Gehrmann, M.; Hermes, M.; Geppert, B.; Schormann, W.; Maccoux, L.J.; Schug, M.; Schumann, A.; et al. Role of thioredoxin reductase 1 and thioredoxin interacting protein in prognosis of breast cancer. Breast Cancer Res. 2010, 12, R44. [Google Scholar] [CrossRef]
- Zhou, J.; Yu, Q.; Chng, W.-J. TXNIP (VDUP-1, TBP-2): A major redox regulator commonly suppressed in cancer by epigenetic mechanisms. Int. J. Biochem. Cell Biol. 2011, 43, 1668–1673. [Google Scholar] [CrossRef]
- Yan, G.-R.; Xu, S.-H.; Tan, Z.-L.; Liu, L.; He, Q.-Y. Global identification of miR-373-regulated genes in breast cancer by quantitative proteomics. PROTEOMICS 2011, 11, 912–920. [Google Scholar] [CrossRef] [PubMed]
- Elgort, M.G.; O’Shea, J.M.; Jiang, Y.; Ayer, D.E. Transcriptional and Translational Downregulation of Thioredoxin Interacting Protein Is Required for Metabolic Reprogramming during G1. Genes Cancer 2010, 1, 893–907. [Google Scholar] [CrossRef] [PubMed]
- Yoshihara, E.; Masaki, S.; Matsuo, Y.; Chen, Z.; Tian, H.; Yodoi, J. Thioredoxin/Txnip: Redoxisome, as a Redox Switch for the Pathogenesis of Diseases. Front. Immunol. 2014, 4. [Google Scholar] [CrossRef] [PubMed]
- Lincoln, D.T.; Ali Emadi, E.M.; Tonissen, K.F.; Clarke, F.M. The thioredoxin-thioredoxin reductase system: Over-expression in human cancer. Anticancer Res. 2003, 23, 2425–2433. [Google Scholar]
- Lin, F.; Zhang, P.; Zuo, Z.; Wang, F.; Bi, R.; Shang, W.; Wu, A.; Ye, J.; Li, S.; Sun, X.; et al. Thioredoxin-1 promotes colorectal cancer invasion and metastasis through crosstalk with S100P. Cancer Lett. 2017, 401, 1–10. [Google Scholar] [CrossRef]
- Yoshioka, J.; Imahashi, K.; Gabel, S.A.; Chutkow, W.A.; Burds, A.A.; Gannon, J.; Schulze, P.C.; MacGillivray, C.; London, R.E.; Murphy, E. Targeted Deletion of Thioredoxin-Interacting Protein Regulates Cardiac Dysfunction in Response to Pressure Overload. Circ. Res. 2007, 101, 1328–1338. [Google Scholar] [CrossRef]
- Heiden, M.G.V.; Cantley, L.C.; Thompson, C.B. Understanding the Warburg Effect: The Metabolic Requirements of Cell Proliferation. Science 2009, 324, 1029–1033. [Google Scholar] [CrossRef]
- Brodbeck, T.; Nehmann, N.; Bethge, A.; Wedemann, G.; Schumacher, U. Perforin-dependent direct cytotoxicity in natural killer cells induces considerable knockdown of spontaneous lung metastases and computer modelling-proven tumor cell dormancy in a HT29 human colon cancer xenograft mouse model. Mol. Cancer 2014, 13, 244. [Google Scholar] [CrossRef]
- Sodeur, S.; Ullrich, S.; Gustke, H.; Zangemeister-Wittke, U.; Schumacher, U. Increased numbers of spontaneous SCLC metastasis in absence of NK cells after subcutaneous inoculation of different SCLC cell lines into pfp/rag2 double knock out mice. Cancer Lett. 2009, 282, 146–151. [Google Scholar] [CrossRef]
- Schumacher, U.; Adam, E. Lectin histochemical HPA-binding pattern of human breast and colon cancers is associated with metastases formation in severe combined immunodeficient mice. Histochem. J. 1997, 29, 677–684. [Google Scholar] [CrossRef] [PubMed]
- Ghulam, J.; Stuerken, C.; Wicklein, D.; Pries, R.; Wollenberg, B.; Schumacher, U. Immunohistochemical Analysis of Transcription Factors and Markers of Epithelial–Mesenchymal Transition (EMT) in Human Tumors. Anticancer Res. 2019, 39, 5437–5448. [Google Scholar] [CrossRef] [PubMed]
- Valentiner, U.; Hall, D.M.S.; Brooks, S.A.; Schumacher, U. HPA binding and metastasis formation of human breast cancer cell lines transplanted into severe combined immunodeficient (scid) mice. Cancer Lett. 2005, 219, 233–242. [Google Scholar] [CrossRef]
- Schröder, C.; Schumacher, U.; Müller, V.; Wirtz, R.M.; Streichert, T.; Richter, U.; Wicklein, D.; Milde-Langosch, K. The transcription factor Fra-2 promotes mammary tumour progression by changing the adhesive properties of breast cancer cells. Eur. J. Cancer 2010, 46, 1650–1660. [Google Scholar] [CrossRef] [PubMed]
- Thies, A.; Mauer, S.; Fodstad, O.; Schumacher, U. Clinically proven markers of metastasis predict metastatic spread of human melanoma cells engrafted in scid mice. Br. J. Cancer 2007, 96, 609–616. [Google Scholar] [CrossRef] [PubMed]
- Valentiner, U.; Valentiner, F.-U.; Schumacher, U. Expression of CD44 Is Associated with a Metastatic Pattern of Human Neuroblastoma Cells in a SCID Mouse Xenograft Model. Tumor Biol. 2008, 29, 152–160. [Google Scholar] [CrossRef] [PubMed]
- Gebauer, F.; Wicklein, D.; Stübke, K.; Nehmann, N.; Schmidt, A.; Salamon, J.; Peldschus, K.; Nentwich, M.F.; Adam, G.; Tolstonog, G.; et al. Selectin binding is essential for peritoneal carcinomatosis in a xenograft model of human pancreatic adenocarcinoma in pfp−−/rag2−− mice. Gut 2013, 62, 741–750. [Google Scholar] [CrossRef]
- Lange, T.; Ullrich, S.; Müller, I.; Nentwich, M.F.; Stübke, K.; Feldhaus, S.; Knies, C.; Hellwinkel, O.J.C.; Vessella, R.L.; Abramjuk, C.; et al. Human Prostate Cancer in a Clinically Relevant Xenograft Mouse Model: Identification of β(1,6)-Branched Oligosaccharides as a Marker of Tumor Progression. Clin. Cancer Res. 2012, 18, 1364–1373. [Google Scholar] [CrossRef]
- Böckelmann, L.; Starzonek, C.; Niehoff, A.-C.; Karst, U.; Thomale, J.; Schlüter, H.; Bokemeyer, C.; Aigner, A.; Schumacher, U. Detection of doxorubicin, cisplatin and therapeutic antibodies in formalin-fixed paraffin-embedded human cancer cells. Histochem. Cell Biol. 2020. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organ System | Tissue | Specific Tumor Entity | n | Negative % | Weak % | Medium % | High % | Leukocytes TXNIP Positive % |
|---|---|---|---|---|---|---|---|---|
| Bone | Chondrosarcoma | 14 | 86 | 7 | 7 | 0 | 7 | |
| Osteosarcoma | 20 | 80 | 20 | 0 | 0 | 15 | ||
| Endocrine | Adrenal gland | Adenoma | 43 | 47 | 35 | 14 | 5 | 0 |
| Carcinoma | 14 | 57 | 43 | 0 | 0 | 0 | ||
| Pheochromocytoma | 30 | 13 | 20 | 13 | 53 | 0 | ||
| Thyroid | Adenoma | 47 | 81 | 11 | 6 | 2 | 4 | |
| Anaplastic carcinoma | 24 | 88 | 13 | 0 | 0 | 13 | ||
| Follicular carcinoma | 47 | 81 | 13 | 4 | 2 | 2 | ||
| Medullary carcinoma | 42 | 69 | 26 | 5 | 0 | 10 | ||
| Papillary carcinoma | 45 | 58 | 24 | 13 | 4 | 2 | ||
| Neuroendocrine tumor | 27 | 41 | 33 | 7 | 19 | 30 | ||
| Gastrointestinal | Anal canal | Squamous cell carcinoma | 28 | 79 | 18 | 4 | 0 | 43 |
| Large intestine | Adenoma, low grade | 18 | 44 | 57 | 0 | 0 | 33 | |
| Adenoma, high grade | 22 | 64 | 27 | 9 | 0 | 64 | ||
| Adenocarcinoma | 29 | 66 | 21 | 14 | 0 | 52 | ||
| Small intestine | Adenocarcinoma | 6 | 33 | 17 | 50 | 0 | 33 | |
| Liver | Cholangiocellular carcinoma | 29 | 55 | 24 | 17 | 3 | 10 | |
| Hepatocellular carcinoma | 41 | 32 | 42 | 22 | 5 | 0 | ||
| Stomach | Carcinoma, diffuse Type | 34 | 94 | 6 | 0 | 0 | 47 | |
| Carcinoma, intestinal Type | 31 | 77 | 19 | 3 | 0 | 52 | ||
| Esophagus | Adenocarcinoma | 30 | 67 | 20 | 13 | 0 | 53 | |
| Squamous cell carcinoma | 29 | 93 | 3 | 3 | 0 | 69 | ||
| Pancreas | Adenocarcinoma, ductal | 36 | 61 | 11 | 17 | 11 | 67 | |
| Adenocarcinoma, papillary | 18 | 78 | 11 | 11 | 0 | 67 | ||
| Neuroendocrine Tumor | 29 | 52 | 21 | 21 | 7 | 45 | ||
| Gastrointestinal Stroma tumor | 37 | 92 | 5 | 3 | 0 | 16 | ||
| Gynecologic | Cervix | Adenocarcinoma | 45 | 73 | 13 | 11 | 2 | 42 |
| Squamous cell carcinoma | 37 | 92 | 5 | 0 | 3 | 51 | ||
| Endometrium | Carcinoma, endometroid | 46 | 74 | 15 | 7 | 4 | 30 | |
| Carcinoma, serous | 40 | 78 | 20 | 3 | 0 | 0 | ||
| Breast | Carcinoma, ductal (NST) | 22 | 87 | 5 | 5 | 5 | 14 | |
| Carcinoma, lobular | 30 | 70 | 20 | 10 | 0 | 43 | ||
| Carcinoma, medullary | 14 | 71 | 21 | 7 | 0 | 50 | ||
| Carcinoma, mucinous | 14 | 71 | 21 | 7 | 0 | 21 | ||
| Carcinoma, phyllodes | 46 | 37 | 24 | 24 | 15 | 30 | ||
| Carcinoma, tubular | 12 | 50 | 25 | 8 | 17 | 50 | ||
| Ovaries | Brenner tumor | 6 | 50 | 50 | 0 | 0 | 17 | |
| Carcinoma, endometroid | 28 | 61 | 21 | 7 | 11 | 11 | ||
| Carcinoma, mucinous | 24 | 54 | 33 | 13 | 0 | 13 | ||
| Carcinoma, serous | 41 | 66 | 20 | 10 | 5 | 22 | ||
| Uterus | Carcinosarcoma | 47 | 81 | 13 | 6 | 0 | 13 | |
| Stroma sarcoma | 12 | 100 | 0 | 0 | 0 | 25 | ||
| Vagina | Squamous cell carcinoma | 26 | 92 | 0 | 8 | 0 | 38 | |
| Vulva | Squamous cell carcinoma | 36 | 78 | 14 | 6 | 3 | 67 | |
| Hematologic | Hodgkin-Lymphoma | 41 | 98 | 2 | 0 | 0 | 27 | |
| Non-Hodgkin-Lymphoma | 43 | 100 | 0 | 0 | 0 | 40 | ||
| Thymoma | 26 | 77 | 19 | 4 | 0 | 46 | ||
| Head, Chest, and Respiratory tract | Larynx | Squamous cell carcinoma | 43 | 81 | 14 | 5 | 0 | 65 |
| Lung (Non-small cell) | Adenocarcinoma | 33 | 73 | 18 | 9 | 0 | 52 | |
| Bronchoalveolar carcinoma | 6 | 67 | 33 | 0 | 0 | 67 | ||
| Bronchial carcinoma, large cell | 21 | 86 | 14 | 0 | 0 | 43 | ||
| Squamous cell carcinoma | 34 | 97 | 3 | 0 | 0 | 50 | ||
| Lung (Small cell) | Carcinoma | 12 | 100 | 0 | 0 | 0 | 42 | |
| Metastasis small cell carcinoma | 2 | 100 | 0 | 0 | 0 | 0 | ||
| Oral cavity | Squamous cell carcinoma | 49 | 67 | 22 | 4 | 6 | 57 | |
| Pleura | Mesothelioma | 24 | 65 | 13 | 19 | 3 | 17 | |
| Salivary glands | Basal cell adenoma | 14 | 43 | 0 | 43 | 14 | 7 | |
| Parotis, pleomorphic adenoma | 47 | 89 | 9 | 0 | 2 | 4 | ||
| Parotis, Warthin tumor | 47 | 9 | 21 | 30 | 41 | 19 | ||
| Skin | Basalioma | 43 | 95 | 2 | 2 | 0 | 0 | |
| Benign Naevus | 27 | 93 | 4 | 4 | 0 | 41 | ||
| Melanoma | 44 | 82 | 7 | 11 | 0 | 18 | ||
| Merkel cell carcinoma | 38 | 100 | 0 | 0 | 0 | 13 | ||
| Pilomatrixoma | 26 | 100 | 0 | 0 | 0 | 35 | ||
| Squamous cell carcinoma | 45 | 84 | 11 | 4 | 0 | 60 | ||
| Soft tissue | Angiosarcoma | 27 | 93 | 0 | 7 | 0 | 63 | |
| Granular cell tumor | 21 | 100 | 0 | 0 | 0 | 76 | ||
| Giant cell tumor of tendon sheath | 38 | 82 | 18 | 0 | 0 | 42 | ||
| Leiomyoma | 50 | 98 | 2 | 0 | 0 | 40 | ||
| Leiomyosarcoma | 48 | 94 | 6 | 0 | 0 | 52 | ||
| Liposarcoma | 35 | 94 | 6 | 0 | 0 | 46 | ||
| Stromal cell | Brain | 20 | 85 | 15 | 0 | 0 | 5 | |
| Heart | 19 | 84 | 16 | 0 | 0 | 0 | ||
| Kidney | 16 | 13 | 0 | 50 | 38 | 0 | ||
| Large intestine | 8 | 63 | 25 | 13 | 0 | 0 | ||
| Liver | 13 | 33 | 8 | 8 | 50 | 0 | ||
| Lymph nodes | 9 | 100 | 0 | 0 | 0 | 22 | ||
| Pancreas | 15 | 0 | 0 | 13 | 87 | 0 | ||
| Prostate | 19 | 58 | 21 | 21 | 0 | 21 | ||
| Skin | 3 | 100 | 0 | 0 | 0 | 33 | ||
| Thyroid | 17 | 94 | 6 | 0 | 0 | 6 | ||
| Urogenital | Kidney | Carcinoma, clear cell | 47 | 70 | 15 | 11 | 4 | 32 |
| Carcinoma, chromophobe | 43 | 61 | 23 | 9 | 7 | 2 | ||
| Carcinoma, papillary | 38 | 58 | 18 | 18 | 5 | 13 | ||
| Oncocytoma | 41 | 10 | 15 | 24 | 51 | 5 | ||
| Prostate | Adenocarcinoma | 45 | 82 | 18 | 0 | 0 | 80 | |
| Small cell | 11 | 100 | 0 | 0 | 0 | 27 | ||
| Germ cell tumor | Carcinoma, embryonal | 49 | 100 | 0 | 0 | 0 | 4 | |
| Seminoma | 48 | 100 | 0 | 0 | 0 | 4 | ||
| Teratoma | 33 | 70 | 18 | 6 | 6 | 55 | ||
| Yolk sac tumor | 45 | 100 | 0 | 0 | 0 | 9 | ||
| Bladder | Carcinoma, small cell | 16 | 88 | 13 | 0 | 0 | 31 | |
| Urothelial | Carcinoma, pTa | 36 | 56 | 14 | 25 | 6 | 19 | |
| Carcinoma, T2-4 | 46 | 74 | 17 | 9 | 0 | 41 |
| Tumor | Cell Line | n | Mean Area of Positive Cells in % | Cells in Agar | Tumor Cells | ||
|---|---|---|---|---|---|---|---|
| Surrounding Tissues | |||||||
| CD45 | TXNIP | TXNIP/CD45 | TXNIP | TXNIP | |||
| Bronchial Carcinoma | H69AR1 | 4 | 6.4 | 1.7 | 26.6 | − | − |
| H69AR3 | 3 | 8.7 | 5.6 | 64.4 | − | − | |
| NCI-H69 | 5 | 4.6 | 1.7 | 37.0 | − | − | |
| NCI-H82 | 4 | 12.1 | 5.5 | 45.5 | − | ||
| SW2 | 4 | 5.1 | 3.1 | 60.8 | − | − | |
| Colon Carcinoma | HT-29 | 5 | 12.5 | 2.1 | 16.8 | ++ | − |
| HCT 116 | + | − | |||||
| Mamma Carcinoma | DU4475 | 2 | 9.6 | 3.6 | 37.5 | + | − |
| MCF-7 | 5 | 7 | 0.7 | 10.0 | − | − | |
| MDA-MB-231 | 5 | 19.6 | 3.8 | 19.4 | − | − | |
| T-47D | 2 | 5.8 | 2.6 | 44.8 | − | ||
| Melanoma | FEMX-I | 5 | 6.3 | 3.6 | 57.1 | − | − |
| LOX-IMVI | 5 | 5.2 | 1.3 | 25.0 | − | − | |
| MDA-MB-435 | 5 | 5.3 | 2.4 | 45.3 | − | ||
| MeWo | 5 | 4.5 | 2.9 | 64.4 | − | − | |
| MV3 | 2 | 4.6 | 1.5 | 32.6 | − | − | |
| Neuroblastoma | LA-N-1 | 7 | 6.5 | 6.5 | 100.0 | − | − |
| LS | 4 | 8.3 | 7 | 84.3 | + | − | |
| Osteosarcoma | U2OS | 3 | 12.4 | 3.5 | 28.2 | − | − |
| HOS | 4 | 8.1 | 3.9 | 48.1 | − | − | |
| Pancreas Carcinoma | PaCa 5072 | 4 | 8.9 | 2.8 | 31.5 | + | − |
| PaCa 5061 | 4 | 13.9 | 3.5 | 25.2 | + | − | |
| BxPC-3 | 5 | 29.9 | 6.3 | 21.1 | − | − | |
| PANC-1 | 5 | 12 | 4 | 33.3 | − | − | |
| Prostate Carcinoma | DU145 | 4 | 7 | 1.9 | 27.1 | − | |
| LNCaP | 4 | 9.9 | 1.9 | 19.2 | − | − | |
| LuCaP 23.1 | 3 | 9.3 | 2.7 | 29.0 | − | ||
| PC-3 | 3 | 9.8 | 2 | 20.4 | − | − | |
| VCaP | 3 | 13.1 | 1.2 | 9.2 | − | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schröder, J.; Schumacher, U.; Böckelmann, L.C. Thioredoxin Interacting Protein (TXNIP) Is Differentially Expressed in Human Tumor Samples but Is Absent in Human Tumor Cell Line Xenografts: Implications for Its Use as an Immunosurveillance Marker. Cancers 2020, 12, 3028. https://doi.org/10.3390/cancers12103028
Schröder J, Schumacher U, Böckelmann LC. Thioredoxin Interacting Protein (TXNIP) Is Differentially Expressed in Human Tumor Samples but Is Absent in Human Tumor Cell Line Xenografts: Implications for Its Use as an Immunosurveillance Marker. Cancers. 2020; 12(10):3028. https://doi.org/10.3390/cancers12103028
Chicago/Turabian StyleSchröder, Joana, Udo Schumacher, and Lukas Clemens Böckelmann. 2020. "Thioredoxin Interacting Protein (TXNIP) Is Differentially Expressed in Human Tumor Samples but Is Absent in Human Tumor Cell Line Xenografts: Implications for Its Use as an Immunosurveillance Marker" Cancers 12, no. 10: 3028. https://doi.org/10.3390/cancers12103028
APA StyleSchröder, J., Schumacher, U., & Böckelmann, L. C. (2020). Thioredoxin Interacting Protein (TXNIP) Is Differentially Expressed in Human Tumor Samples but Is Absent in Human Tumor Cell Line Xenografts: Implications for Its Use as an Immunosurveillance Marker. Cancers, 12(10), 3028. https://doi.org/10.3390/cancers12103028