IL3RA-Targeting Antibody–Drug Conjugate BAY-943 with a Kinesin Spindle Protein Inhibitor Payload Shows Efficacy in Preclinical Models of Hematologic Malignancies

 , , and
, , and
Abstract
Simple Summary
Abstract
1. Introduction
2. Results
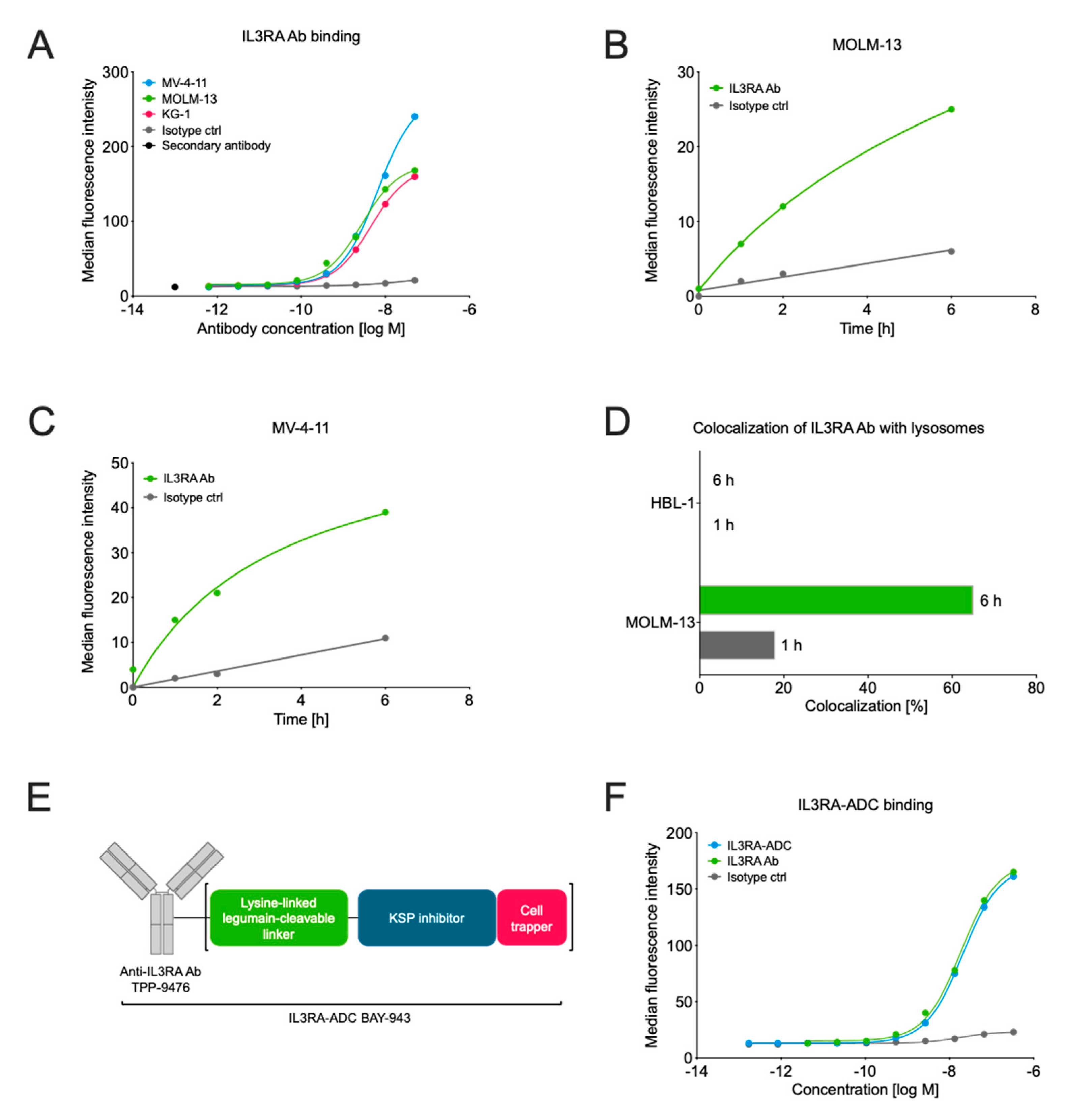
2.1. Characterization of the IL3RA-Targeting Antibody TPP-9476 and IL3RA-ADC BAY-943
2.2. IL3RA-ADC Shows Potent and Selective Efficacy In Vitro
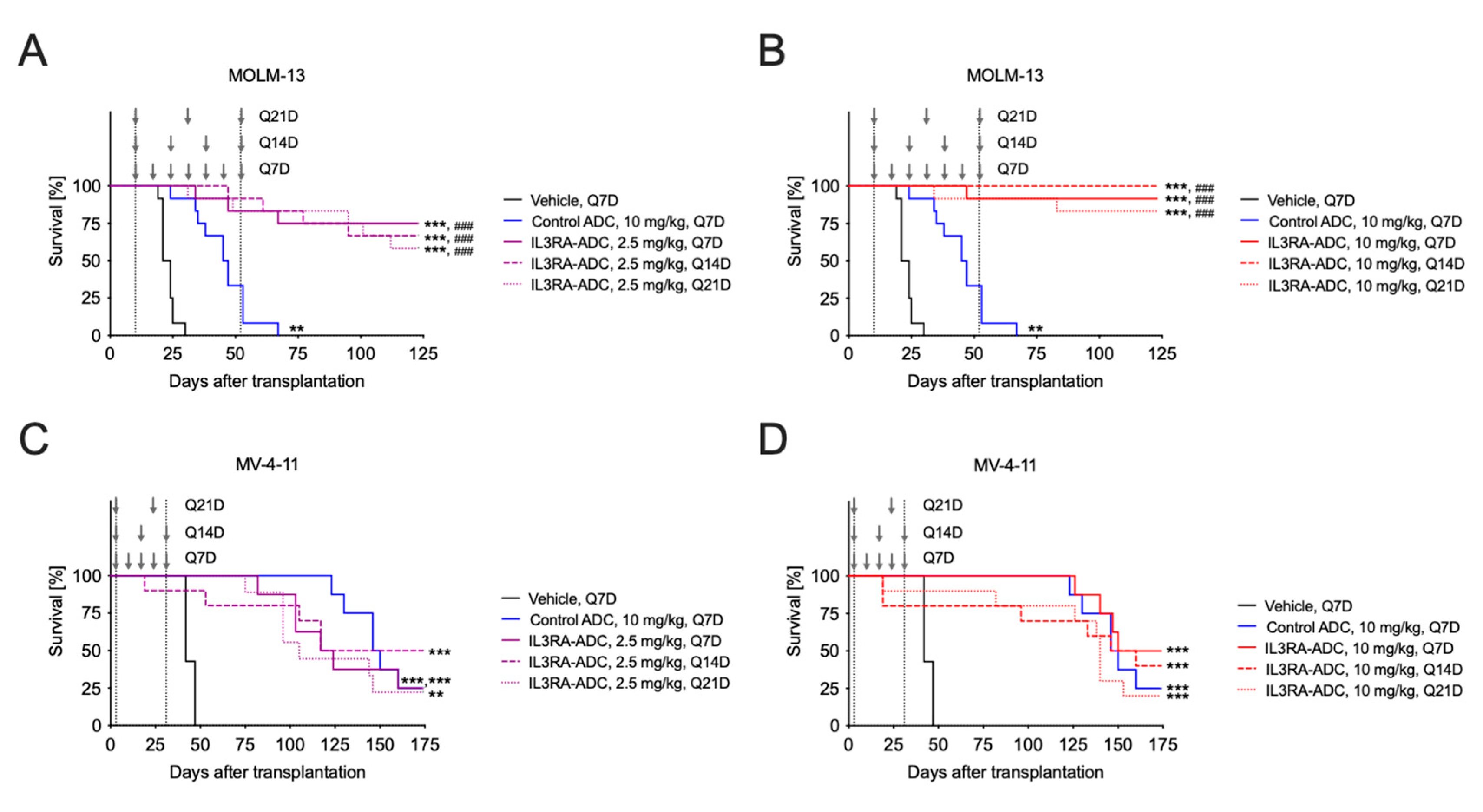
2.3. IL3RA-ADC Improves Survival in the MOLM-13 and MV-4-11 Xenograft Models
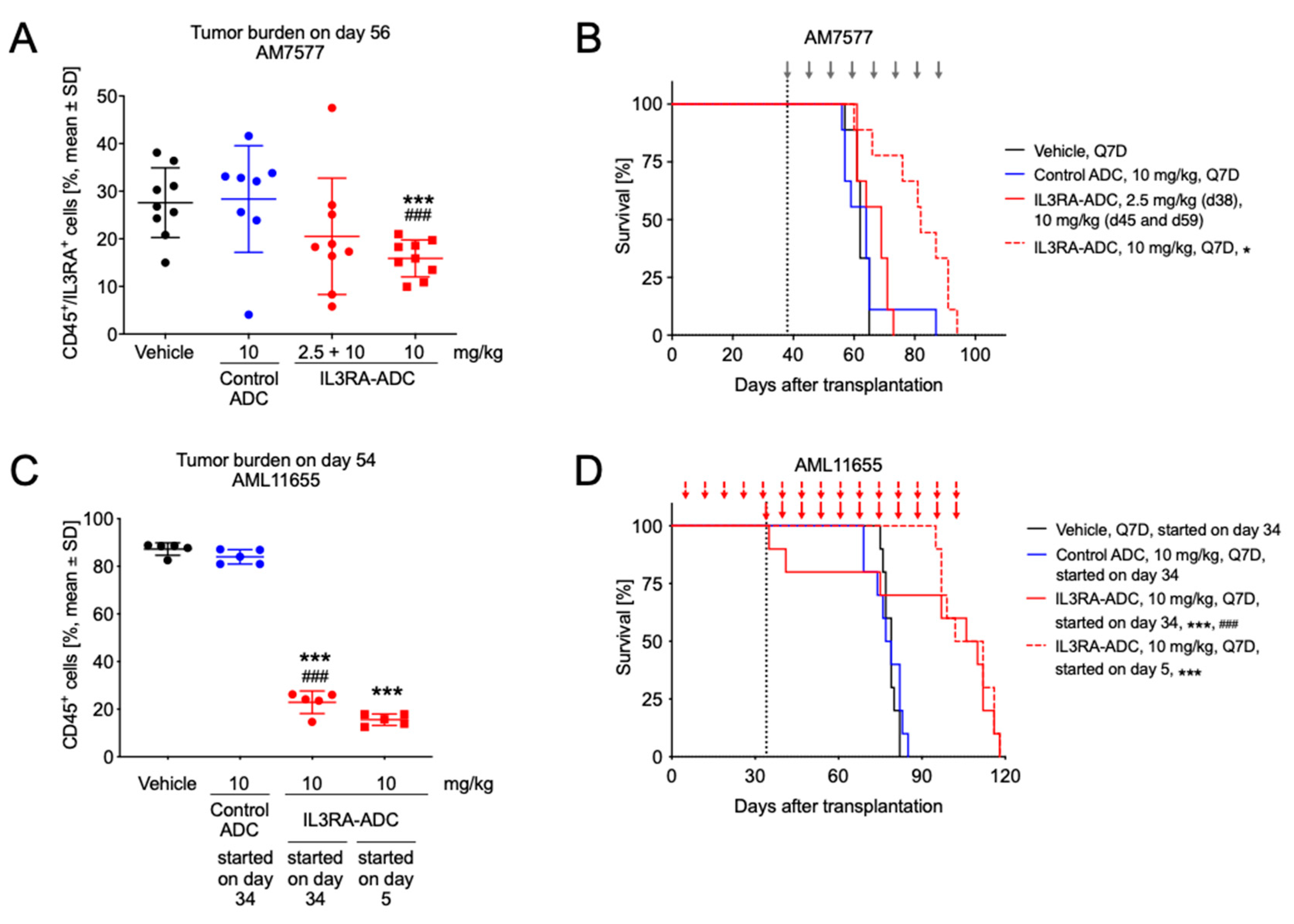
2.4. IL3RA-ADC Suppresses Tumor Burden and Improves Survival in Systemic AM7577 and AML11655 PDX Models
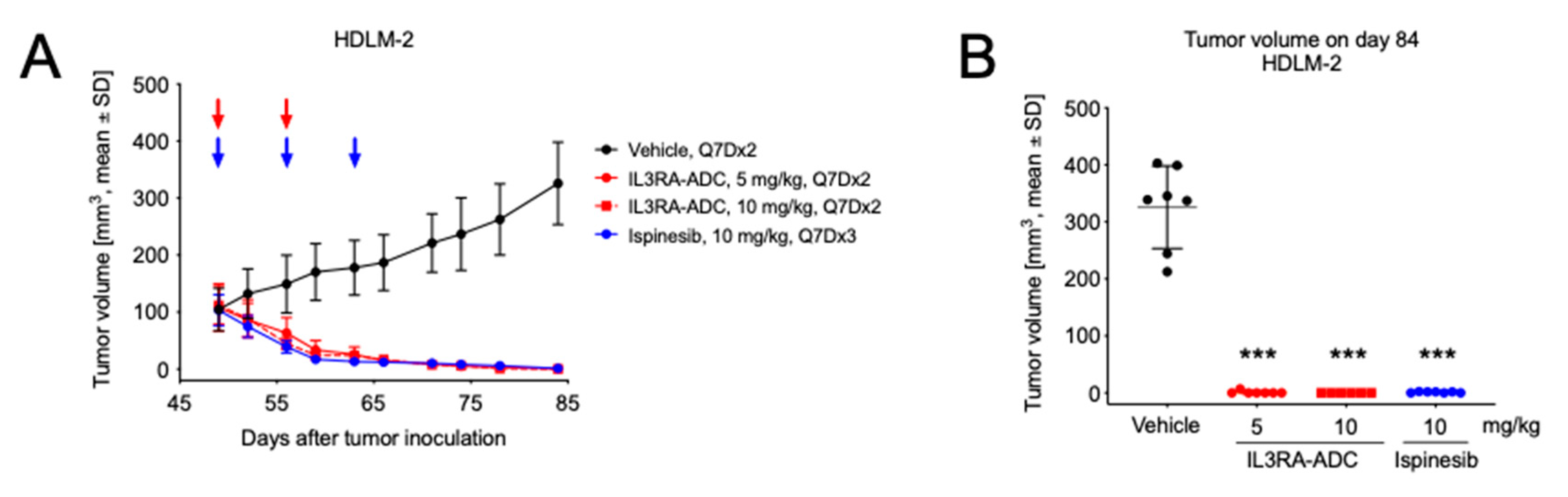
2.5. IL3RA-ADC Demonstrates Antitumor Efficacy in Subcutaneous HDLM-2 Hodgkin Lymphoma Xenograft Model
2.6. IL3RA-ADC Is Well-Tolerated
3. Discussion
4. Materials and Methods
4.1. Cell Lines
4.2. Compounds
4.3. Internalization and Lysosomal Colocalization of IL3RA-Ab
4.4. In Vitro Cytotoxicity of IL3RA-ADC
4.5. In Vivo Studies
4.6. Statsitical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| Ab | antibody |
| ABC | antibodies bound per cell |
| ADC | antibody–drug conjugate |
| AML | acute myeloid leukemia |
| BPDCN | blastic plasmacytoid dendritic cell neoplasm |
| CAR-T | chimeric antigen receptor T cell |
| cHL | classical Hodgkin lymphoma |
| DAR | drug-to-antibody ratio |
| DLBCL | diffuse large B-cell lymphoma |
| EC50 | half-maximal effective concentration |
| FACS | fluorescence-activated cell sorting |
| FCS | fetal calf serum |
| hCD45 | human CD45 |
| hIL3RA | human IL3RA |
| HL | Hodgkin lymphoma |
| IC50 | half-maximal inhibitory concentration |
| IGN | indolinobenzodiazepine pseudodimer |
| IL-3 | interleukin 3 |
| IL3RA | interleukin 3 receptor subunit alpha |
| IL3RA-Ab | IL3RA-targeting antibody |
| i.p. | intraperitoneal(ly) |
| i.v. | intravenous(ly) |
| KD | dissociation constant |
| KSP | kinesin spindle protein |
| KSPi | kinesin spindle protein inhibitor |
| MDS | myelodysplastic syndrome |
| MFI | median fluorescence intensity |
| MST | median survival time |
| Papp | apparent permeability |
| PBD | pyrrolobenzodiazepine |
| PBS | phosphate-buffered saline |
| pDCs | plasmacytoid dendritic cells |
| PDX | patient-derived xenograft |
| P-gP | P-glycoprotein |
| Q7D | once weekly |
| Q14D | every two weeks |
| Q21D | every three weeks |
| rrMM | relapsed refractory multiple myeloma |
| s.c. | subcutaneous(ly) |
| siRNA | small interfering RNA |
| SPR | surface plasmon resonance |
References
- Ehninger, A.; Kramer, M.; Röllig, C.; Thiede, C.; Bornhäuser, M.; von Bonin, M.; Wermke, M.; Feldmann, A.; Bachmann, M.; Ehninger, G.; et al. Distribution and levels of cell surface expression of CD33 and CD123 in acute myeloid leukemia. Blood Cancer J. 2014, 4, e218. [Google Scholar] [CrossRef]
- Muñoz, L.; Nomdedéu, J.F.; Lopez, O.; Carnicer, M.J.; Bellido, M.; Aventín, A.; Brunet, S.; Sierra, J. Interleukin-3 receptor alpha chain (CD123) is widely expressed in hematologic malignancies. Haematologica 2001, 86, 1261–1269. [Google Scholar]
- Testa, U.; Pelosi, E.; Frankel, A. CD 123 is a membrane biomarker and a therapeutic target in hematologic malignancies. Biomark. Res. 2014, 2, 4. [Google Scholar] [CrossRef]
- Lopez, A.F.; Hercus, T.R.; Ekert, P.; Littler, D.R.; Guthridge, M.; Thomas, D.; Ramshaw, H.S.; Stomski, F.; Perugini, M.; D’Andrea, R.; et al. Molecular basis of cytokine receptor activation. Int. Union Biochem. Mol. Biol. Life 2010, 62, 509–518. [Google Scholar] [CrossRef] [PubMed]
- Bras, A.E.; de Haas, V.; van Stigt, A.; Jongen-Lavrencic, M.; Beverloo, H.B.; Te Marvelde, J.G.; Zwaan, C.M.; van Dongen, J.J.M.; Leusen, J.H.W.; van der Velden, V.H.J. CD123 expression levels in 846 acute leukemia patients based on standardized immunophenotyping. Cytom. Part B Clin. Cytom. 2019, 96, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Aldinucci, D.; Poletto, D.; Gloghini, A.; Nanni, P.; Degan, M.; Perin, T.; Ceolin, P.; Rossi, F.M.; Gattei, V.; Carbone, A.; et al. Expression of functional interleukin-3 receptors on Hodgkin and Reed-Sternberg cells. Am. J. Pathol. 2002, 160, 585–596. [Google Scholar] [CrossRef]
- Fromm, J.R. Flow cytometric analysis of CD123 is useful for immunophenotyping classical Hodgkin lymphoma. Cytom. Part B Clin. Cytom. 2011, 80, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Ruella, M.; Klichinsky, M.; Kenderian, S.S.; Shestova, O.; Ziober, A.; Kraft, D.O.; Feldman, M.; Wasik, M.A.; June, C.H.; Gill, S. Overcoming the immunosuppressive tumor microenvironment of Hodgkin lymphoma using chimeric antigen receptor T cells. Cancer Discov. 2017, 7, 1154–1167. [Google Scholar] [CrossRef] [PubMed]
- Kerr, D., II; Zhang, L.; Sokol, L. Blastic plasmacytoid dendritic cell neoplasm. Curr. Treat. Options Oncol. 2019, 20, 9. [Google Scholar] [CrossRef]
- Testa, U.; Pelosi, E.; Castelli, G. CD123 as a therapeutic target in the treatment of hematological malignancies. Cancers 2019, 11, 1358. [Google Scholar] [CrossRef]
- De Smet, D.; Trullemans, F.; Jochmans, K.; Renmans, W.; Smet, L.; Heylen, O.; Bael, A.M.; Schots, R.; Leus, B.; De Waele, M. Diagnostic potential of CD34+ cell antigen expression in myelodysplastic syndromes. Am. J. Clin. Pathol. 2012, 138, 732–743. [Google Scholar] [CrossRef] [PubMed]
- Li, L.J.; Tao, J.L.; Fu, R.; Wang, H.Q.; Jiang, H.J.; Yue, L.Z.; Zhang, W.; Liu, H.; Shao, Z.H. Increased CD34+CD38−CD123+ cells in myelodysplastic syndrome displaying malignant features similar to those in AML. Int. J. Hematol. 2014, 100, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Shastri, A.; Will, B.; Steidl, U.; Verma, A. Stem and progenitor cell alterations in myelodysplastic syndromes. Blood 2017, 129, 1586–1594. [Google Scholar] [CrossRef] [PubMed]
- Yue, L.Z.; Fu, R.; Wang, H.Q.; Li, L.J.; Hu, H.R.; Fu, L.; Shao, Z.H. Expression of CD123 and CD114 on the bone marrow cells of patients with myelodysplastic syndrome. Chin. Med. J. (UK) 2010, 123, 2034–2037. [Google Scholar]
- Testa, U.; Riccioni, R.; Militi, S.; Coccia, E.; Stellacci, E.; Samoggia, P.; Latagliata, R.; Mariani, G.; Rossini, A.; Battistini, A.; et al. Elevated expression of IL-3Ralpha in acute myelogenous leukemia is associated with enhanced blast proliferation, increased cellularity, and poor prognosis. Blood 2002, 100, 2980–2988. [Google Scholar] [CrossRef]
- Lantz, C.S.; Boesiger, J.; Song, C.H.; Mach, N.; Kobayashi, T.; Mulligan, R.C.; Nawa, Y.; Dranoff, G.; Galli, S.J. Role for interleukin-3 in mast-cell and basophil development and in immunity to parasites. Nature 1998, 392, 90–93. [Google Scholar] [CrossRef]
- Broughton, S.E.; Dhagat, U.; Hercus, T.R.; Nero, T.L.; Grimbaldeston, M.A.; Bonder, C.S.; Lopez, A.F.; Parker, M.W. The GM-CSF/IL-3/IL-5 cytokine receptor family: From ligand recognition to initiation of signaling. Immunol. Rev. 2012, 250, 277–302. [Google Scholar] [CrossRef]
- Jin, L.; Lee, E.M.; Ramshaw, H.S.; Busfield, S.J.; Peoppl, A.G.; Wilkinson, L.; Guthridge, M.A.; Thomas, D.; Barry, E.F.; Boyd, A.; et al. Monoclonal antibody-mediated targeting of CD123, IL-3 receptor alpha chain, eliminates human acute myeloid leukemic stem cells. Cell Stem Cell 2009, 5, 31–42. [Google Scholar] [CrossRef]
- Jordan, C.T.; Upchurch, D.; Szilvassy, S.J.; Guzman, M.L.; Howard, D.S.; Pettigrew, A.L.; Meyerrose, T.; Rossi, R.; Grimes, B.; Rizzieri, D.A.; et al. The interleukin-3 receptor alpha chain is a unique marker for human acute myelogenous leukemia stem cells. Leukemia 2000, 14, 1777–1784. [Google Scholar] [CrossRef]
- Coustan-Smith, E.; Song, G.; Clark, C.; Key, L.; Liu, P.; Mehrpooya, M.; Stow, P.; Su, X.; Shurtleff, S.; Pui, C.H.; et al. New markers for minimal residual disease detection in acute lymphoblastic leukemia. Blood 2011, 117, 6267–6276. [Google Scholar] [CrossRef]
- Han, L.; Jorgensen, J.L.; Wang, S.A.; Huang, X.; Nogueras González, G.M.; Brooks, C.; Rowinsky, E.; Levis, M.; Zhou, J.; Ciurea, S.O.; et al. Leukemia stem cell marker CD123 (IL-3R alpha) predicts minimal residual disease and relapse, providing a valid target for SL-101 in acute myeloid leukemia with FLT3-ITD mutations. Blood 2013, 122, 359. [Google Scholar] [CrossRef]
- Lerchen, H.G.; Rebstock, A.S.; Cancho-Grande, Y.; Wittrock, S.; Stelte-Ludwig, B.; Kirchhoff, D.; Mahlert, C.; Greven, S.; Märsch, S. Specific Antibody-Drug-Conjugates (ADCs) with KSP Inhibitors and Anti-CD123-Antibodies. Patent WO 2017/216028, 21 December 2017. [Google Scholar]
- Lerchen, H.G.; Stelte-Ludwig, B.; Berndt, S.; Sommer, A.; Dietz, L.; Rebstock, A.S.; Johannes, S.; Marx, L.; Jorissen, H.; Mahlert, C.; et al. Antibody-prodrug conjugates with KSP inhibitors and legumain-mediated metabolite formation. Chemistry 2019, 25, 8208–8213. [Google Scholar] [CrossRef] [PubMed]
- Lerchen, H.G.; Stelte-Ludwig, B.; Sommer, A.; Berndt, M.C.; Rebstock, A.-S.; Johannes, S.; Mahlert, C.; Greven, S.; Dietz, L.; Jörissen, H. Tailored linker chemistries for the efficient and selective activation of ADCs with KSPi payloads. Bioconjugate Chem. 2020, 31, 1893–1898. [Google Scholar] [CrossRef] [PubMed]
- Lerchen, H.G.; Wittrock, S.; Stelte-Ludwig, B.; Sommer, A.; Berndt, S.; Griebenow, N.; Rebstock, A.S.; Johannes, S.; Cancho-Grande, Y.; Mahlert, C.; et al. Antibody-drug conjugates with pyrrole-based KSP inhibitors as the payload class. Angew. Chem. Int. Ed. 2018, 57, 15243–15247. [Google Scholar] [CrossRef] [PubMed]
- Myers, S.M.; Collins, I. Recent findings and future directions for interpolar mitotic kinesin inhibitors in cancer therapy. Future Med. Chem. 2016, 8, 463–489. [Google Scholar] [CrossRef] [PubMed]
- Carter, B.Z.; Mak, D.H.; Woessner, R.; Gross, S.; Schober, W.D.; Estrov, Z.; Kantarjian, H.; Andreeff, M. Inhibition of KSP by ARRY-520 induces cell cycle block and cell death via the mitochondrial pathway in AML cells. Leukemia 2009, 23, 1755–1762. [Google Scholar] [CrossRef] [PubMed]
- El-Nassan, H.B. Advances in the discovery of kinesin spindle protein (Eg5) inhibitors as antitumor agents. Eur. J. Med. Chem. 2013, 62, 614–631. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.; Xing, N.; Lu, J.; Zhang, H.; Nishizawa, K.; Liu, S.; Yuan, X.; Qin, Y.; Liu, Y.; Ogawa, O.; et al. Overexpression of Eg5 predicts unfavorable prognosis in non-muscle invasive bladder urothelial carcinoma. Int. J. Urol. 2011, 18, 432–438. [Google Scholar] [CrossRef] [PubMed]
- Mayer, T.U.; Kapoor, T.M.; Haggarty, S.J.; King, R.W.; Schreiber, S.L.; Mitchison, T.J. Small molecule inhibitor of mitotic spindle bipolarity identified in a phenotype-based screen. Science 1999, 286, 971–974. [Google Scholar] [CrossRef] [PubMed]
- Pelosi, E.; Castelli, G.; Testa, U. Targeting LSCs through membrane antigens selectively or preferentially expressed on these cells. Blood Cells Mol. Dis. 2015, 55, 336–346. [Google Scholar] [CrossRef] [PubMed]
- Rath, O.; Kozielski, F. Kinesins and cancer. Nat. Rev. Cancer 2012, 12, 527–539. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Woodcock, J.M.; Rapoport, A.; Stomski, F.C.; Korpelainen, E.I.; Bagley, C.J.; Goodall, G.J.; Smith, W.B.; Gamble, J.R.; Vadas, M.A.; et al. Monoclonal antibody 7G3 recognizes the N-terminal domain of the human interleukin-3 (IL-3) receptor alpha-chain and functions as a specific IL-3 receptor antagonist. Blood 1996, 87, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Quentmeier, H.; Reinhardt, J.; Zaborski, M.; Drexler, H.G. FLT3 mutations in acute myeloid leukemia cell lines. Leukemia 2003, 17, 120–124. [Google Scholar] [CrossRef] [PubMed]
- Wakui, H.; Yamamoto, N.; Kitazono, S.; Mizugaki, H.; Nakamichi, S.; Fujiwara, Y.; Nokihara, H.; Yamada, Y.; Suzuki, K.; Kanda, H.; et al. A phase 1 and dose-finding study of LY2523355 (litronesib), an Eg5 inhibitor, in Japanese patients with advanced solid tumors. Cancer Chemother. Pharmacol. 2014, 74, 15–23. [Google Scholar] [CrossRef]
- Angelova, E.; Audette, C.; Kovtun, Y.; Daver, N.; Wang, S.A.; Pierce, S.; Konoplev, S.N.; Khogeer, H.; Jorgensen, J.L.; Konopleva, M.; et al. CD123 expression patterns and selective targeting with a CD123-targeted antibody-drug conjugate (IMGN632) in acute lymphoblastic leukemia. Haematologica 2019, 104, 749–755. [Google Scholar] [CrossRef]
- Kovtun, Y.; Jones, G.E.; Adams, S.; Harvey, L.; Audette, C.A.; Wilhelm, A.; Bai, C.; Rui, L.; Laleau, R.; Liu, F.; et al. A CD123-targeting antibody-drug conjugate, IMGN632, designed to eradicate AML while sparing normal bone marrow cells. Blood Adv. 2018, 2, 848–858. [Google Scholar] [CrossRef]
- Li, F.; Sutherland, M.K.; Yu, C.; Walter, R.B.; Westendorf, L.; Valliere-Douglass, J.; Pan, L.; Cronkite, A.; Sussman, D.; Klussman, K.; et al. Characterization of SGN-CD123A, a potent CD123-directed antibody-drug conjugate for acute myeloid leukemia. Mol. Cancer Ther. 2018, 17, 554–564. [Google Scholar] [CrossRef]
- Sutherland, M.S.K.; Yu, C.; Walter, R.B.; Westendorf, L.; Valliere-Douglass, J.; Pan, L.; Sussman, D.; Anderson, M.; Zeng, W.; Stone, I.; et al. SGN-CD123A, a pyrrolobenzodiazepine dimer linked anti-CD123 antibody drug conjugate, demonstrates effective anti-leukemic activity in multiple preclinical models of AML. Blood 2015, 126, 330. [Google Scholar] [CrossRef]
- Zhu, C.; Zhao, J.; Bibikova, M.; Leverson, J.D.; Bossy-Wetzel, E.; Fan, J.B.; Abraham, R.T.; Jiang, W. Functional analysis of human microtubule-based motor proteins, the kinesins and dyneins, in mitosis/cytokinesis using RNA interference. Mol. Biol. Cell 2005, 16, 3187–3199. [Google Scholar] [CrossRef]
- Burris, H.A., III; Jones, S.F.; Williams, D.D.; Kathman, S.J.; Hodge, J.P.; Pandite, L.; Ho, P.T.; Boerner, S.A.; Lorusso, P. A phase I study of ispinesib, a kinesin spindle protein inhibitor, administered weekly for three consecutive weeks of a 28-day cycle in patients with solid tumors. Investig. New Drugs 2011, 29, 467–472. [Google Scholar] [CrossRef]
- Gomez, H.L.; Philco, M.; Pimentel, P.; Kiyan, M.; Monsalvo, M.L.; Conlan, M.G.; Saikali, K.G.; Chen, M.M.; Seroogy, J.J.; Wolff, A.A.; et al. Phase I dose-escalation and pharmacokinetic study of ispinesib, a kinesin spindle protein inhibitor, administered on days 1 and 15 of a 28-day schedule in patients with no prior treatment for advanced breast cancer. Anticancer Drugs 2012, 23, 335–341. [Google Scholar] [CrossRef] [PubMed]
- Khoury, H.J.; Garcia-Manero, G.; Borthakur, G.; Kadia, T.; Foudray, M.C.; Arellano, M.; Langston, A.; Bethelmie-Bryan, B.; Rush, S.; Litwiler, K.; et al. A phase 1 dose-escalation study of ARRY-520, a kinesin spindle protein inhibitor, in patients with advanced myeloid leukemias. Cancer 2012, 118, 3556–3564. [Google Scholar] [CrossRef] [PubMed]
- Shah, J.J.; Kaufman, J.L.; Zonder, J.A.; Cohen, A.D.; Bensinger, W.I.; Hilder, B.W.; Rush, S.A.; Walker, D.H.; Tunquist, B.J.; Litwiler, K.S.; et al. A phase 1 and 2 study of filanesib alone and in combination with low-dose dexamethasone in relapsed/refractory multiple myeloma. Cancer 2017, 123, 4617–4630. [Google Scholar] [CrossRef] [PubMed]
- Beck, A.; Goetsch, L.; Dumontet, C.; Corvaia, N. Strategies and challenges for the next generation of antibody-drug conjugates. Nat. Rev. Drug Discov. 2017, 16, 315–337. [Google Scholar] [CrossRef]
- Birrer, M.J.; Moore, K.N.; Betella, I.; Bates, R.C. Antibody-drug conjugate-based therapeutics: State of the science. J. Natl. Cancer Inst. 2019, 111, 538–549. [Google Scholar] [CrossRef]
- Tolcher, A.W. The evolution of antibody-drug conjugates: A positive inflexion point. Am. Soc. Clin. Oncol. Educ. Book 2020, 40, 1–8. [Google Scholar] [CrossRef]
- Donaghy, H. Effects of antibody, drug and linker on the preclinical and clinical toxicities of antibody-drug conjugates. MAbs 2016, 8, 659–671. [Google Scholar] [CrossRef]
- Masters, J.C.; Nickens, D.J.; Xuan, D.; Shazer, R.L.; Amantea, M. Clinical toxicity of antibody drug conjugates: A meta-analysis of payloads. Investig. New Drugs 2018, 36, 121–135. [Google Scholar] [CrossRef]
- Lo, K.M.; Leger, O.; Hock, B. Antibody engineering. Microbiol. Spectr. 2014, 2, AID-0007-2012. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. Available online: https://www.R-project.org/ (accessed on 18 November 2020).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | Provider (Catalog No.) | Date Obtained | Date Authenticated | Origin | Anti-IL3RA ABC | IL3RA-ADC IC50 (M) |
|---|---|---|---|---|---|---|
| MV-4-11 | ATCC (CRL 9591) | 5/5/2008 | 02/05/2019 | Biphenotypic B myelomonocytic leukemia | ≈26,700 | 1.58 × 10−10 |
| MOLM-13 | DSMZ (ACC 554) | 5/2/2008 | 02/05/2019 | Acute myeloid leukemia | ≈15,100 a | 6.37 × 10−10 |
| HDLM-2 | DSMZ (ACC 17) | 19/02/2015 | 06/05/2015 | Pleural effusion of Hodgkin lymphoma | ≈74,300 | 1.97 × 10−9 |
| L-428 | origin unknown | 1996 | 17/04/2013 | Pleural effusion of Hodgkin lymphoma | ≈111,300 | 3.97 × 10−10 |
| THP-1 | ATCC (TIB 202) | 15/02/2006 | 19/03/2014 | Acute monocytic leukemia | ≈21,100 | 2.92 × 10−9 |
| KG-1 | ATCC (CCL 246) | 28/10/2010 | 24/03/2011 | Acute myelogenous leukemia | ≈7200 | 8.34 × 10−9 |
| HT | DSMZ (ACC567) | 12/09/2013 | 19/03/2014 | Diffuse mixed lymphoma | ≈350 | >3.00 × 10−7 |
| NCI-H292 | ATCC (CRL 1848) | 13/08/2009 | 07/02/2012 | Non-small cell lung cancer | ≈500 | >3.00 × 10−7 |
| HBL-1 | Charité (Prof. Lenz) | 15/04/2011 | 03/11/2017 | Diffuse B cell lymphoma | n.d. | n.d. |
| Kasumi-3 | DSMZ (ACC 714) | 20/04/2017 | 05/09/2017 | Acute myeloid leukemia | 23,500 | 6.89 × 10−9 b |
| Rec-1 | ATCC (CRL-3004) | 24/02/2014 | 23/08/2018 | Mantle cell lymphoma | n.d. | 1.03 × 10−7 |
| OVCAR-8 | NCI (NCI-60 panel) | 20/10/2008 | 02/05/2019 | Ovarian cancer | n.d. | 1.47 × 10−7 |
| MDA-MB-231 | ATCC HTB-26 | 05/04/2006 | 15/10/2019 | Breast cancer | ≈890 | >3.00 × 10−7 |
| Ramos | ATCC CRL 1596 | 08/03/2011 | 06/0572015 | Burkitt’s lymphoma | n.d. a | n.d. |
| Cytotoxicity, IC50 (M) | ||||||
|---|---|---|---|---|---|---|
| Compound | DAR | MV-4-11 (IL3RA ≈26,700) | MOLM-13 (IL3RA ≈15,100) | HDLM-2 (IL3RA ≈74,300) | THP-1 (IL3RA ≈21,100) | NCI-H292 (IL3RA ≈500) |
| IL3RA-ADC | 6.3 | 1.58 × 10−10 | 6.37 × 10−10 | 1.29 × 10−9 | 2.92 × 10−9 | >3.00 × 10−7 |
| Control ADC | 7 | >3.00 × 10−7 | 2.18 × 10−9 | 1.52 × 10−7 | 1.48 × 10−8 | 2.12 × 10−7 |
| IL3RA-Ab | n.a. | >3.00 × 10−7 | >3.00 × 10−7 | >3.00 × 10−7 | >3.00 × 10−7 | >3.00 × 10−7 |
| KSPi SMOL | n.a. | 9.05 × 10−11 | 8.95 × 10−11 | 1.00 × 10−10 | 3.07 × 10−10 | 2.16 × 10−10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kirchhoff, D.; Stelte-Ludwig, B.; Lerchen, H.-G.; Wengner, A.M.; Ahsen, O.v.; Buchmann, P.; Märsch, S.; Mahlert, C.; Greven, S.; Dietz, L.; et al. IL3RA-Targeting Antibody–Drug Conjugate BAY-943 with a Kinesin Spindle Protein Inhibitor Payload Shows Efficacy in Preclinical Models of Hematologic Malignancies. Cancers 2020, 12, 3464. https://doi.org/10.3390/cancers12113464
Kirchhoff D, Stelte-Ludwig B, Lerchen H-G, Wengner AM, Ahsen Ov, Buchmann P, Märsch S, Mahlert C, Greven S, Dietz L, et al. IL3RA-Targeting Antibody–Drug Conjugate BAY-943 with a Kinesin Spindle Protein Inhibitor Payload Shows Efficacy in Preclinical Models of Hematologic Malignancies. Cancers. 2020; 12(11):3464. https://doi.org/10.3390/cancers12113464
Chicago/Turabian StyleKirchhoff, Dennis, Beatrix Stelte-Ludwig, Hans-Georg Lerchen, Antje Margret Wengner, Oliver von Ahsen, Pascale Buchmann, Stephan Märsch, Christoph Mahlert, Simone Greven, Lisa Dietz, and et al. 2020. "IL3RA-Targeting Antibody–Drug Conjugate BAY-943 with a Kinesin Spindle Protein Inhibitor Payload Shows Efficacy in Preclinical Models of Hematologic Malignancies" Cancers 12, no. 11: 3464. https://doi.org/10.3390/cancers12113464
APA StyleKirchhoff, D., Stelte-Ludwig, B., Lerchen, H.-G., Wengner, A. M., Ahsen, O. v., Buchmann, P., Märsch, S., Mahlert, C., Greven, S., Dietz, L., Erkelenz, M., Zierz, R., Johanssen, S., Mumberg, D., & Sommer, A. (2020). IL3RA-Targeting Antibody–Drug Conjugate BAY-943 with a Kinesin Spindle Protein Inhibitor Payload Shows Efficacy in Preclinical Models of Hematologic Malignancies. Cancers, 12(11), 3464. https://doi.org/10.3390/cancers12113464