Rab GTPases: Emerging Oncogenes and Tumor Suppressive Regulators for the Editing of Survival Pathways in Cancer

 , and
, and
Abstract
:1. Introduction
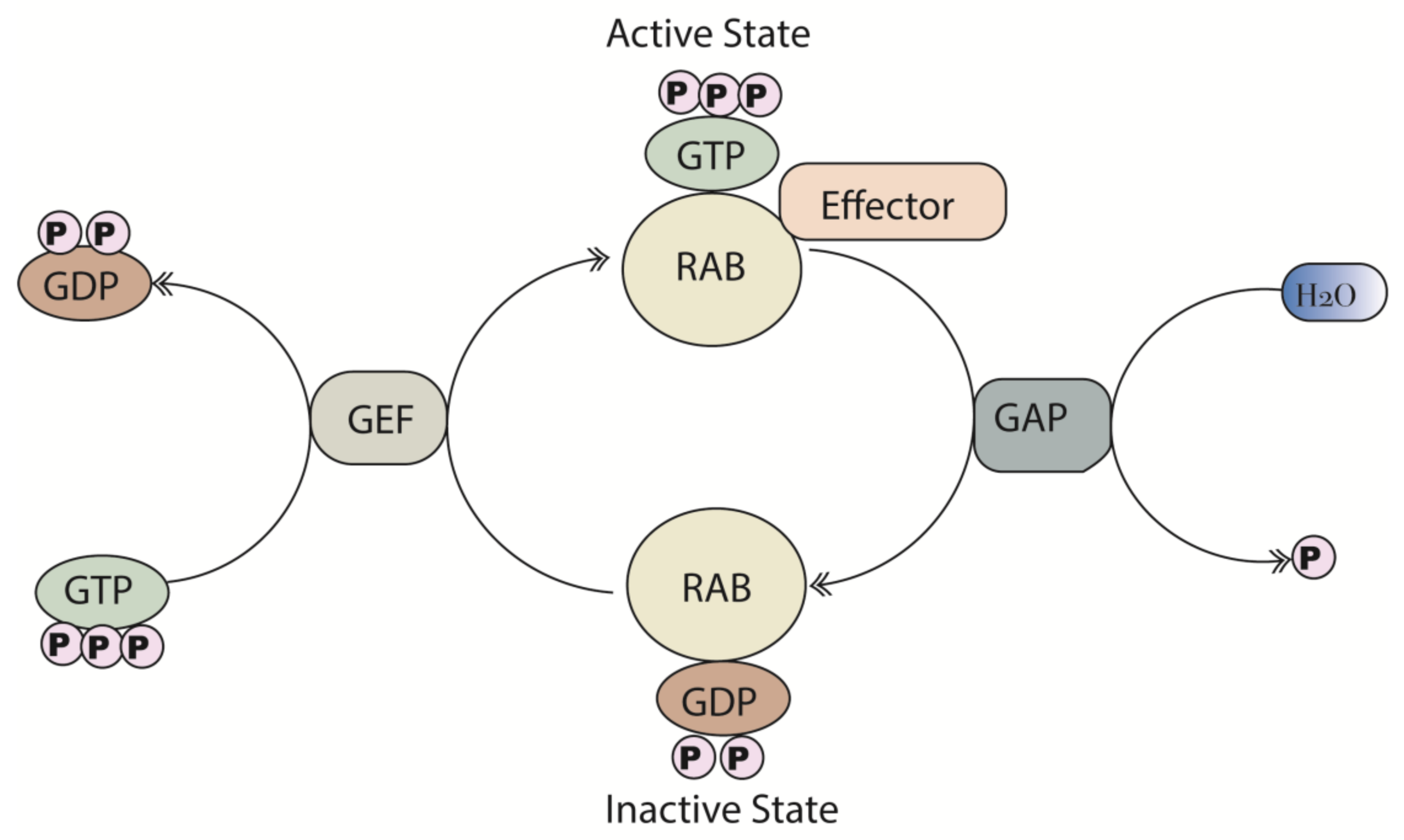
2. Structure and Function of Rab Proteins
3. Dysregulated Rab Expression in Cancer and other Genetic Diseases
4. Regulation of Apoptosis by Rab Proteins
4.1. Overview of Apoptotic Signaling Pathways
4.2. Rabs Involved in the Regulation of Apoptotic Proteins via Signaling Pathways
4.2.1. Rab25
4.2.2. Rab31
4.2.3. Rab35
4.2.4. Rab14
4.2.5. Rab2b
4.3. Rabs that Regulate Mitochondrial Membrane Potential
4.4. Regulation of Apoptosis through Production of Intracellular Stress
4.5. Rabs Regulating Cell Cycle Progression
5. Therapeutic Strategies to Modulate Apoptosis
5.1. Treatment Approaches Targeting the Anti-Apoptotic BCL-2 Family of Proteins
5.2. Treatment Approaches Based on Targeting Caspases
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Tsuruta, F.; Masuyama, N.; Gotoh, Y. The phosphatidylinositol 3-kinase (PI3K)-Akt pathway suppresses Bax translocation to mitochondria. J. Biol. Chem. 2002, 277, 14040–14047. [Google Scholar] [CrossRef] [Green Version]
- Pereira-Leal, J.B.; Seabra, M.C. The mammalian Rab family of small GTPases: Definition of family and subfamily sequence motifs suggests a mechanism for functional specificity in the Ras superfamily. J. Mol. Biol. 2000, 301, 1077–1087. [Google Scholar] [CrossRef] [Green Version]
- Li, G. Rab GTPases, membrane trafficking and diseases. Curr. Drug Targets 2011, 12, 1188–1193. [Google Scholar] [CrossRef] [Green Version]
- Ménasché, G.; Pastural, E.; Feldmann, J.; Certain, S.; Ersoy, F.; Dupuis, S.; Wulffraat, N.; Bianchi, D.; Fischer, A.; Le Deist, F.; et al. Mutations in RAB27A cause griscelli syndrome associated with haemophagocytic syndrome. Nat. Genet. 2000, 25, 173–176. [Google Scholar] [CrossRef]
- Wasmeier, C.; Romao, M.; Plowright, L.; Bennett, D.C.; Raposo, G.; Seabra, M.C. Rab38 and Rab32 control post-Golgi trafficking of melanogenic enzymes. J. Cell Biol. 2006, 175, 271–281. [Google Scholar] [CrossRef] [Green Version]
- Giannandrea, M.; Bianchi, V.; Mignogna, M.L.; Sirri, A.; Carrabino, S.; D’Elia, E.; Vecellio, M.; Russo, S.; Cogliati, F.; Larizza, L.; et al. Mutations in the small GTPase gene RAB39B are responsible for X-linked mental retardation associated with autism, epilepsy, and macrocephaly. Am. J. Hum. Genet. 2010, 86, 185–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colicelli, J. Human RAS superfamily proteins and related GTPases. Sci. STKE 2004, 2004, re13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira-Leal, J.B.; Seabra, M.C. Evolution of the rab family of small GTP-binding proteins. J. Mol. Biol. 2001, 313, 889–901. [Google Scholar] [CrossRef] [Green Version]
- Segev, N. Ypt/rab gtpases: Regulators of protein trafficking. Sci. STKE 2001, 2001, re11. [Google Scholar] [CrossRef]
- Hutagalung, H.A.; Novick, P.J. Role of rab GTPases in membrane traffic and cell physiology. Physiol. Rev. 2011, 91, 119–149. [Google Scholar] [CrossRef] [Green Version]
- Shinde, R.S.; Maddika, S. Post translational modifications of Rab GTPases. Small GTPases 2018, 9, 49–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griscelli, C.; Durandy, A.; Guy-Grand, D.; Daguillard, F.; Herzog, C.; Prunieras, M. A syndrome associating partial albinism and immunodeficiency. Am. J. Med. 1978, 65, 691–702. [Google Scholar] [CrossRef]
- Jenkins, D.; Seelow, D.; Jehee, F.S.; Perlyn, C.A.; Alonso, L.G.; Bueno, D.F.; Donnai, D.; Josifiova, D.; Mathijssen, I.M.; Morton, J.E.; et al. RAB23 mutations in carpenter syndrome imply an unexpected role for hedgehog signaling in cranial-suture development and obesity. Am. J. Hum. Genet. 2007, 80, 1162–1170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ginsberg, S.D.; Alldred, M.J.; Counts, S.E.; Cataldo, A.M.; Neve, R.L.; Jiang, Y.; Wuu, J.; Chao, M.V.; Mufson, E.J.; Nixon, R.A.; et al. Microarray analysis of hippocampal CA1 neurons implicates early endosomal dysfunction during Alzheimer’s disease progression. Biol. Psychiatry 2010, 68, 885–893. [Google Scholar] [CrossRef] [Green Version]
- Inoue, J.; Ninomiya, M.; Umetsu, T.; Nakamura, T.; Kogure, T.; Kakazu, E.; Iwata, T.; Takai, S.; Sano, A.; Fukuda, M.; et al. siRNA screening for the small GTPase rab proteins identifies that Rab5B is a major regulator of hepatitis B virus production. J. Virol. 2019, JVI.00621. [Google Scholar] [CrossRef] [Green Version]
- Thomas, J.D.; Zhang, Y.J.; Wei, Y.H.; Cho, J.H.; Morris, L.E.; Wang, H.Y.; Zheng, X.S. Rab1A is an mTORC1 activator and a colorectal oncogene. Cancer Cell 2014, 26, 754–769. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Yoo, Y.; Fan, H.; Kim, E.; Guan, K.L.; Guan, J.L. Regulation of Integrin β 1 recycling to lipid rafts by Rab1a to promote cell migration. J. Biol. Chem. 2010, 285, 29398–29405. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Liu, W.; Lu, X.A.; Fu, Y.; Li, L.; Luo, Y. High expression of small GTPase Rab3D promotes cancer progression and metastasis. Oncotarget 2015, 6, 11125–11138. [Google Scholar] [CrossRef] [Green Version]
- Wheeler, D.B.; Zoncu, R.; Root, D.E.; Sabatini, D.M.; Sawyers, C.L. Identification of an oncogenic RAB protein. Science 2015, 350, 211–217. [Google Scholar] [CrossRef] [Green Version]
- Silva, P.; Mendoza, P.; Rivas, S.; Díaz, J.; Moraga, C.; Quest, A.F.; Torres, V.A. Hypoxia promotes Rab5 activation, leading to tumor cell migration, invasion and metastasis. Oncotarget 2016, 7, 29548–29562. [Google Scholar] [CrossRef] [Green Version]
- Cheng, K.W.; Lahad, J.P.; Gray, J.W.; Mills, G.B. Emerging role of RAB GTPases in cancer and human disease. Cancer Res. 2005, 65, 2516–2519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villagomez, F.R.; Medina-Contreras, O.; Cerna-Cortes, J.F.; Patino-Lopez, G. The role of the oncogenic Rab35 in cancer invasion, metastasis, and immune evasion, especially in leukemia. Small GTPases 2018, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caswell, P.T.; Spence, H.J.; Parsons, M.; White, D.P.; Clark, K.; Cheng, K.W.; Mills, G.B.; Humphries, M.J.; Messent, A.J.; Anderson, K.I.; et al. Rab25 associates with α5β1 integrin to promote invasive migration in 3D microenvironments. Dev. Cell 2007, 13, 496–510. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Mao, Z.; Liu, L.; Zhang, R.; Liang, Q.; Xiong, Y.; Yuan, W.; Wei, L. Rab17 inhibits the tumorigenic properties of hepatocellular carcinoma via the Erk pathway. Tumour Biol. 2015, 36, 5815–5824. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.Y.; Tseng, R.C.; Hsu, H.S.; Wang, Y.C.; Hsu, M.T. Frequent down-regulation of hRAB37 in metastatic tumor by genetic and epigenetic mechanisms in lung cancer. Lung Cancer 2009, 63, 360–367. [Google Scholar] [CrossRef]
- Tsai, C.H.; Cheng, H.C.; Wang, Y.S.; Lin, P.; Jen, J.; Kuo, I.Y.; Chang, Y.H.; Liao, P.C.; Chen, R.H.; Yuan, W.C.; et al. Small GTPase Rab37 targets tissue inhibitor of metalloproteinase 1 for exocytosis and thus suppresses tumour metastasis. Nat. Commun. 2014, 5, 4804. [Google Scholar] [CrossRef] [Green Version]
- Cheng, J.M.; Volk, L.; Janaki, D.K.; Vyakaranam, S.; Ran, S.; Rao, K.A. Tumor suppressor function of Rab25 in triple-negative breast cancer. Int. J. Cancer 2010, 126, 2799–2812. [Google Scholar] [CrossRef]
- Nam, K.T.; Lee, H.J.; Smith, J.J.; Lapierre, L.A.; Kamath, V.P.; Chen, X.; Aronow, B.J.; Yeatman, T.J.; Bhartur, S.G.; Calhoun, B.C.; et al. Loss of Rab25 promotes the development of intestinal neoplasia in mice and is associated with human colorectal adenocarcinomas. J. Clin. Investig. 2010, 120, 840–849. [Google Scholar] [CrossRef] [Green Version]
- Tong, M.; Chan, K.W.; Bao, J.Y.; Wong, K.Y.; Chen, J.N.; Kwan, P.S.; Tang, K.H.; Fu, L.; Qin, Y.R.; Lok, S.; et al. Rab25 is a tumor suppressor gene with antiangiogenic and anti-invasive activities in esophageal squamous cell carcinoma. Cancer Res. 2012, 72, 6024–6035. [Google Scholar] [CrossRef] [Green Version]
- Ichim, G.; Tait, S.W.G. A fate worse than death: Apoptosis as an oncogenic process. Nat. Rev. Cancer 2016, 16, 539. [Google Scholar] [CrossRef] [Green Version]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Carrington, E.M.; Zhan, Y.; Brady, J.L.; Zhang, J.G.; Sutherland, R.M.; Anstee, N.S.; Schenk, R.L.; Vikstrom, I.B.; Delconte, R.B.; Segal, D.; et al. Anti-apoptotic proteins BCL-2, MCL-1 and A1 summate collectively to maintain survival of immune cell populations both in vitro and in vivo. Cell Death Differ. 2017, 24, 878–888. [Google Scholar] [CrossRef] [Green Version]
- Saelens, X.; Festjens, N.; Walle, L.V.; Van Gurp, M.; van Loo, G.; Vandenabeele, P. Toxic proteins released from mitochondria in cell death. Oncogene 2004, 23, 2861–2874. [Google Scholar] [CrossRef] [Green Version]
- Hsu, H.; Xiong, J.; Goeddel, D.V. The TNF receptor 1-associated protein TRADD signals cell death and NF-κB activation. Cell 1995, 81, 495–504. [Google Scholar] [CrossRef] [Green Version]
- Ashkenazi, A.; Dixit, V.M. Death receptors: Signaling and modulation. Science 1998, 281, 1305–1308. [Google Scholar] [CrossRef] [Green Version]
- Cohen, G.M. Caspases: The executioners of apoptosis. Biochem. J. 1997, 326, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Saraste, A.; Pulkki, K. Morphologic and biochemical hallmarks of apoptosis. Cardiovasc. Res. 2000, 45, 528–537. [Google Scholar] [CrossRef]
- Kaushal, V.; Herzog, C.; Haun, R.S.; Kaushal, G.P. Caspase protocols in mice. Methods Mol. Biol. 2014, 1133, 141–154. [Google Scholar]
- Logue, S.E.; Martin, S.J. Caspase activation cascades in apoptosis. Biochem. Soc. Trans. 2008, 36, 1–9. [Google Scholar] [CrossRef]
- Jason, S.L.; Cui, W. Proliferation, survival and metabolism: The role of PI3K/AKT/mTOR signalling in pluripotency and cell fate determination. Development 2016, 143, 3050–3060. [Google Scholar]
- Janku, F.; Yap, T.A.; Meric-Bernstam, F. Targeting the PI3K pathway in cancer: Are we making headway? Nat. Rev. Clin. Oncol. 2018, 15, 273. [Google Scholar] [CrossRef]
- Chang, F.; Lee, J.T.; Navolanic, P.M.; Steelman, L.S.; Shelton, J.G.; Blalock, W.L.; Franklin, R.A.; McCubrey, J.A. Involvement of PI3K/Akt pathway in cell cycle progression, apoptosis, and neoplastic transformation: A target for cancer chemotherapy. Leukemia 2003, 17, 590–603. [Google Scholar] [CrossRef] [Green Version]
- Sangawa, A.; Shintani, M.; Yamao, N.; Kamoshida, S. Phosphorylation status of Akt and caspase-9 in gastric and colorectal carcinomas. Int. J. Clin. Exp. Pathol. 2014, 7, 3312–3317. [Google Scholar]
- Zhou, H.; Li, X.M.; Meinkoth, J.; Pittman, R.N. Akt regulates cell survival and apoptosis at a postmitochondrial level. J. Cell Biol. 2000, 151, 483–494. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Zhang, H.; Shi, L.; Zhang, W.; Yuan, J.; Chen, X.; Liu, J.; Zhang, Y.; Wang, Z. Inhibition of Rac1 activity induces G1/S phase arrest through the GSK3/cyclin D1 pathway in human cancer cells. Oncol. Rep. 2014, 32, 1395–1400. [Google Scholar] [CrossRef] [Green Version]
- Abukhdeir, A.M.; Park, B.H. P21 and p27: Roles in carcinogenesis and drug resistance. Expert Rev. Mol. Med. 2008, 10. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Hu, C.; Wu, F.; He, S. Rab25 GTPase: Functional roles in cancer. Oncotarget 2017, 8, 64591–64599. [Google Scholar] [CrossRef] [Green Version]
- Casanova, J.E.; Wang, X.; Kumar, R.; Bhartur, S.G.; Navarre, J.; Woodrum, J.E.; Altschuler, Y.; Ray, G.S.; Goldenring, J.R. Association of Rab25 and Rab11a with the apical recycling system of polarized Madin-Darby canine kidney cells. Mol. Biol. Cell 1999, 10, 47–61. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, R.; Jurisica, I.; Mills, G.B.; Cheng, K.W. The emerging role of the RAB25 small GTPase in cancer. Traffic 2009, 10, 1561–1568. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Kumar, R.; Navarre, J.; Casanova, J.E.; Goldenring, J.R. Regulation of vesicle trafficking in madin-darby canine kidney cells by Rab11a and Rab25. J. Biol. Chem. 2000, 275, 29138–29146. [Google Scholar] [CrossRef] [Green Version]
- Hoekstra, D.; Tyteca, D.; van IJzendoorn, S.C. The subapical compartment: A traffic center in membrane polarity development. J. Cell Sci. 2004, 117, 2183–2192. [Google Scholar] [CrossRef] [Green Version]
- Cheng, K.W.; Agarwal, R.; Mitra, S.; Lee, J.S.; Carey, M.; Gray, J.W.; Mills, G.B. Rab25 increases cellular ATP and glycogen stores protecting cancer cells from bioenergetic stress. EMBO Mol. Med. 2012, 4, 125–141. [Google Scholar] [CrossRef]
- Cheng, K.W.; Lahad, J.P.; Kuo, W.L.; Lapuk, A.; Yamada, K.; Auersperg, N.; Liu, J.; Smith-McCune, K.; Lu, K.H.; Fishman, D.; et al. The RAB25 small GTPase determines aggressiveness of ovarian and breast cancers. Nat. Med. 2004, 10, 1251–1256. [Google Scholar] [CrossRef]
- Kotzsch, M.; Sieuwerts, A.M.; Grosser, M.; Meye, A.; Fuessel, S.; Meijer-van Gelder, M.E.; Smid, M.; Schmitt, M.; Baretton, G.; Luther, T.; et al. Urokinase receptor splice variant uPAR-del4/5-associated gene expression in breast cancer: Identification of rab31 as an independent prognostic factor. Breast Cancer Res. Treat. 2007, 111, 229–240. [Google Scholar] [CrossRef]
- Grismayer, B.; Sölch, S.; Seubert, B.; Kirchner, T.; Schäfer, S.; Baretton, G.; Schmitt, M.; Luther, T.; Krüger, A.; Kotzsch, M.; et al. Rab31 expression levels modulate tumor-relevant characteristics of breast cancer cells. Mol. Cancer 2012, 11, 62. [Google Scholar] [CrossRef] [Green Version]
- Jin, C.; Rajabi, H.; Pitroda, S.; Li, A.; Kharbanda, A.; Weichselbaum, R.; Kufe, D. Cooperative interaction between the MUC1-C oncoprotein and the Rab31 GTPase in estrogen receptor-positive breast cancer cells. PLoS ONE 2012, 7, e39432. [Google Scholar] [CrossRef] [Green Version]
- Sui, Y.; Zheng, X.; Zhao, D. Rab31 promoted hepatocellular carcinoma (HCC) progression via inhibition of cell apoptosis induced by PI3K/AKT/Bcl-2/BAX pathway. Tumor Biol. 2015, 36, 8661–8670. [Google Scholar] [CrossRef]
- Pan, Y.; Zhang, Y.; Chen, L.; Liu, Y.; Feng, Y.; Yan, J. The critical role of Rab31 in cell proliferation and apoptosis in cancer progression. Mol. Neurobiol. 2016, 53, 4431–4437. [Google Scholar] [CrossRef]
- Tang, C.T.; Liang, Q.; Yang, L.; Lin, X.L.; Wu, S.; Chen, Y.; Zhang, X.T.; Gao, Y.J.; Ge, Z.Z. RAB31 targeted by MiR-30c-2–3p regulates the GLI1 signaling pathway, affecting gastric cancer cell proliferation and apoptosis. Front. Oncol. 2018, 8, 554. [Google Scholar] [CrossRef]
- Davey, J.R.; Humphrey, S.J.; Junutula, J.R.; Mishra, A.K.; Lambright, D.G.; James, D.E.; Stöckli, J. TBC1D13 is a RAB35 specific GAP that plays an important role in GLUT4 trafficking in adipocytes. Traffic 2012, 13, 1429–1441. [Google Scholar] [CrossRef] [Green Version]
- Shaughnessy, R.; Echard, A. Rab35 GTPase and cancer: Linking membrane trafficking to tumorigenesis. Traffic 2018, 19, 247–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Proikas-Cezanne, T.; Gaugel, A.; Frickey, T.; Nordheim, A. Rab14 is part of the early endosomal clathrin-coated TGN microdomain. FEBS Lett. 2006, 580, 5241–5246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reed, S.E.; Hodgson, L.R.; Song, S.; May, M.T.; Kelly, E.E.; McCaffrey, M.W.; Mastick, C.C.; Verkade, P.; Tavaré, J.M. A role for Rab14 in the endocytic trafficking of GLUT4 in 3T3-L1 adipocytes. J. Cell Sci. 2013, 126, 1931–1941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, B.; Wang, W.; Zhao, Z.; Li, Q.; Zhou, K.; Zhao, L.; Wang, L.; Yang, J.; Huang, C. Rab14 act as oncogene and induce proliferation of gastric cancer cells via AKT signaling pathway. PLoS ONE 2017, 12, e0170620. [Google Scholar] [CrossRef]
- Ni, X.; Ma, Y.; Cheng, H.; Jiang, M.; Guo, L.; Ji, C.; Gu, S.; Cao, Y.; Xie, Y.; Mao, Y. Molecular cloning and characterization of a novel human Rab (Rab2B) gene. J. Hum. Genet. 2002, 47, 548–551. [Google Scholar] [CrossRef] [Green Version]
- Jin, J.; Wu, Y.; Zhou, D.; Sun, Q.; Wang, W. miR-448 targets Rab2B and is pivotal in the suppression of pancreatic cancer. Oncol. Rep. 2018, 40, 1379–1389. [Google Scholar] [CrossRef]
- Wu, Q.; Sun, X.; Yue, W.; Lu, T.; Ruan, Y.; Chen, T.; Zhang, D. RAB18, a protein associated with Warburg Micro syndrome, controls neuronal migration in the developing cerebral cortex. Mol. Brain 2016, 9, 19. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.; Li, Y.; Wu, L.; Li, Y.; Zhao, D.; Yu, J.; Huang, T.; Ferguson, C.; Parton, R.G.; Yang, H.; et al. Rab18 promotes lipid droplet (LD) growth by tethering the ER to LDs through SNARE and NRZ interactions. J. Cell Biol. 2018, 217, 975–995. [Google Scholar] [CrossRef] [Green Version]
- Wu, B.; Qi, R.; Liu, X.; Qian, L.; Wu, Z. Rab18 overexpression promotes proliferation and chemoresistance through regulation of mitochondrial function in human gastric cancer. OncoTargets Ther. 2018, 11, 7805–7820. [Google Scholar] [CrossRef] [Green Version]
- Oshita, H.; Nishino, R.; Takano, A.; Fujitomo, T.; Aragaki, M.; Kato, T.; Akiyama, H.; Tsuchiya, E.; Kohno, N.; Nakamura, Y.; et al. RASEF is a novel diagnostic biomarker and a therapeutic target for lung cancer. Mol. Cancer Res. 2013, 11, 937–951. [Google Scholar] [CrossRef] [Green Version]
- Shintani, M.; Tada, M.; Kobayashi, T.; Kajiho, H.; Kontani, K.; Katada, T. Characterization of Rab45/RASEF containing EF-hand domain and a coiled-coil motif as a self-associating GTPase. Biochem. Biophys. Res. Commun. 2007, 357, 661–667. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, S.; Takemura, T.; Tan, L.; Nagata, Y.; Yokota, D.; Hirano, I.; Shigeno, K.; Shibata, K.; Fujie, M.; Fujisawa, S.; et al. Small GTPase RAB45-mediated p38 activation in apoptosis of chronic myeloid leukemia progenitor cells. Carcinogenesis 2011, 32, 1758–1772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, G.; Yussman, M.G.; Barrett, T.J.; Hahn, H.S.; Osinska, H.; Hilliard, G.M.; Wang, X.; Toyokawa, T.; Yatani, A.; Lynch, R.A.; et al. Increased myocardial Rab GTPase expression: A consequence and cause of cardiomyopathy. Circ. Res. 2001, 89, 1130–1137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winslow, A.R.; Chen, C.W.; Corrochano, S.; Acevedo-Arozena, A.; Gordon, D.E.; Peden, A.A.; Lichtenberg, M.; Menzies, F.M.; Ravikumar, B.; Imarisio, S.; et al. α-Synuclein impairs macroautophagy: Implications for Parkinson’s disease. J. Cell Biol. 2010, 190, 1023–1037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.Z.; Li, X.X.; Zhang, Y.J.; Rodriguez-Rodriguez, L.; Xiang, M.Q.; Wang, H.Y.; Zheng, X.S. Rab1 in cell signaling, cancer and other diseases. Oncogene 2016, 35, 5699–5704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saraste, J.; Lahtinen, U.; Goud, B. Localization of the small GTP-binding protein rab1p to early compartments of the secretory pathway. J. Cell Sci. 1995, 108, 1541. [Google Scholar]
- Yang, Y.; Hou, N.; Wang, X.; Wang, L.; Chang, S.E.; He, K.; Zhao, Z.; Zhao, X.; Song, T.; Huang, C. miR-15b-5p induces endoplasmic reticulum stress and apoptosis in human hepatocellular carcinoma, both in vitro and in vivo, by suppressing Rab1A. Oncotarget 2015, 6, 16227–16238. [Google Scholar] [CrossRef] [Green Version]
- Pucci, B.; Kasten, M.; Giordano, A. Cell cycle and apoptosis. Neoplasia 2000, 2, 291–299. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.A.; Kang, O.H.; Kwon, D.Y. Cryptotanshinone induces cell cycle arrest and apoptosis of NSCLC cells through the PI3K/Akt/GSK-3β pathway. Int. J. Mol. Sci. 2018, 19, 2739. [Google Scholar] [CrossRef] [Green Version]
- Diao, B.; Huang, X.; Yang, C.; Guo, S.; Fei, L.; Chen, Y.; Wu, Y. Rab13 silencing causes inhibition of growth and induction of apoptosis in human glioma cells. Int. J. Clin. Exp. Pathol. 2016, 9, 3007–3014. [Google Scholar]
- Ge, J.; Chen, Q.; Liu, B.; Wang, L.; Zhang, S.; Ji, B. Knockdown of Rab21 inhibits proliferation and induces apoptosis in human glioma cells. Cell. Mol. Biol. Lett. 2017, 22, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Jin, Q.; Huang, F.; Tang, Z.; Huang, J. Effects of Rab27A and Rab27B on invasion, proliferation, apoptosis, and chemoresistance in human pancreatic cancer cells. Pancreas 2017, 46, 1173–1179. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, X.; Zhang, Z.; Xiao, B.; An, B.; Zhang, J. The overexpression of Rab9 promotes tumor progression regulated by XBP1 in breast cancer. OncoTargets Ther. 2019, 12, 1815–1824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Jia, W.D.; Hu, B.; Pan, Y.Y. RAB10 overexpression promotes tumor growth and indicates poor prognosis of hepatocellular carcinoma. Oncotarget 2017, 8, 26434–26447. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Wang, W.; Li, Z.; Chen, Z.; Zhi, X.; Xu, J.; Li, Q.; Wang, L.; Huang, X.; Wang, L.; et al. MicroRNA-148a-3p enhances cisplatin cytotoxicity in gastric cancer through mitochondrial fission induction and cyto-protective autophagy suppression. Cancer Lett. 2017, 410, 212–227. [Google Scholar] [CrossRef]
- Liu, Y.; Zeng, C.; Bao, N.; Zhao, J.; Hu, Y.; Li, C.; Chi, S. Effect of Rab23 on the proliferation and apoptosis in breast cancer. Oncol. Rep. 2015, 34, 1835–1844. [Google Scholar] [CrossRef]
- Ganley, I.G.; Carroll, K.; Bittova, L.; Pfeffer, S. Rab9 GTPase regulates late endosome size and requires effector interaction for its stability. Mol. Biol. Cell 2004, 15, 5420–5430. [Google Scholar] [CrossRef] [Green Version]
- Schuldt, A. ER trailblazing by RAB10. Nat. Rev. Mol. Cell Biol. 2013, 14, 63. [Google Scholar] [CrossRef]
- Matsui, T.; Fukuda, M. Rab12 regulates mTORC1 activity and autophagy through controlling the degradation of amino-acid transporter PAT4. EMBO Rep. 2013, 14, 450–457. [Google Scholar] [CrossRef] [Green Version]
- Gerondopoulos, A.; Bastos, R.N.; Yoshimura, S.I.; Anderson, R.; Carpanini, S.; Aligianis, I.; Handley, M.T.; Barr, F.A. Rab18 and a Rab18 GEF complex are required for normal ER structure. J. Cell Biol. 2014, 205, 707. [Google Scholar] [CrossRef] [Green Version]
- Pellinen, T.; Arjonen, A.; Vuoriluoto, K.; Kallio, K.; Fransen, J.A.; Ivaska, J. Small GTPase Rab21 regulates cell adhesion and controls endosomal traffic of β1-integrins. J. Cell Biol. 2006, 173, 767–780. [Google Scholar] [CrossRef] [PubMed]
- Evans, T.M.; Ferguson, C.; Wainwright, B.J.; Parton, R.G.; Wicking, C. Rab23, a negative regulator of hedgehog signaling, localizes to the plasma membrane and the endocytic pathway. Traffic 2003, 4, 869–884. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Fang, R.; Fang, J.; He, S.; Liu, T. Functional implications of Rab27 GTPases in cancer. Cell Commun. Signal. 2018, 16, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Zhang, Y.; Yang, J.; Li, S.; Chen, J. Upregulation of miR-582–5p inhibits cell proliferation, cell cycle progression and invasion by targeting Rab27a in human colorectal carcinoma. Cancer Gene Ther. 2015, 22, 475–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez-Gabin, A.G.; Yin, X.; Si, Q.; Larocca, J.N. Transport of mannose-6-phosphate receptors from the trans-Golgi network to endosomes requires Rab31. Exp. Cell Res. 2009, 315, 2215–2230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joung, J.Y.; Lee, H.Y.; Park, J.; Lee, J.Y.; Chang, B.H.; No, K.T.; Nam, K.Y.; Hwang, J.S. Identification of novel rab27a/melanophilin blockers by pharmacophore-based virtual screening. Appl. Biochem. Biotechnol. 2014, 172, 1882–1897. [Google Scholar] [CrossRef] [PubMed]
- Sorolla, A.; Wang, E.; Golden, E.; Duffy, C.; Henriques, S.T.; Redfern, A.D.; Blancafort, P. Precision medicine by designer interference peptides: Applications in oncology and molecular therapeutics. Oncogene 2019. [Google Scholar] [CrossRef] [Green Version]
- Wong, R.S.Y. Apoptosis in cancer: From pathogenesis to treatment. J. Exp. Clin. Cancer Res. CR 2011, 30, 87. [Google Scholar] [CrossRef] [Green Version]
- Fischer, U.; Schulze-Osthoff, K. Apoptosis-based therapies and drug targets. Cell Death Differ. 2005, 12, 942–961. [Google Scholar] [CrossRef]
- Kang, M.H.; Reynolds, C.P. Bcl-2 inhibitors: Targeting mitochondrial apoptotic pathways in cancer therapy. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2009, 15, 1126–1132. [Google Scholar] [CrossRef] [Green Version]
- Mérino, D.; Khaw, S.L.; Glaser, S.P.; Anderson, D.J.; Belmont, L.D.; Wong, C.; Yue, P.; Robati, M.; Phipson, B.; Fairlie, W.D.; et al. Bcl-2, Bcl-x(L), and Bcl-w are not equivalent targets of ABT-737 and navitoclax (ABT-263) in lymphoid and leukemic cells. Blood 2012, 119, 5807–5816. [Google Scholar] [CrossRef] [PubMed]
- Bedikian, A.Y.; Millward, M.; Pehamberger, H.; Conry, R.; Gore, M.; Trefzer, U.; Pavlick, A.C.; DeConti, R.; Hersh, E.M.; Hersey, P.; et al. Bcl-2 antisense (oblimersen sodium) plus dacarbazine in patients with advanced melanoma: The oblimersen melanoma study group. J. Clin. Oncol. 2006, 24, 4738–4745. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, S.; Moore, J.O.; Boyd, T.E.; Larratt, L.M.; Skotnicki, A.; Koziner, B.; Chanan-Khan, A.A.; Seymour, J.F.; Bociek, R.G.; Pavletic, S.; et al. Randomized phase III trial of fludarabine plus cyclophosphamide with or without oblimersen sodium (Bcl-2 antisense) in patients with relapsed or refractory chronic lymphocytic leukemia. J. Clin. Oncol. 2007, 25, 1114–1120. [Google Scholar] [CrossRef] [PubMed]
- Moore, V.D.; Brown, J.R.; Certo, M.; Love, T.M.; Novina, C.D.; Letai, A. Chronic lymphocytic leukemia requires BCL2 to sequester prodeath BIM, explaining sensitivity to BCL2 antagonist ABT-737. J. Clin. Investig. 2007, 117, 112–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oltersdorf, T.; Elmore, S.W.; Shoemaker, A.R.; Armstrong, R.C.; Augeri, D.J.; Belli, B.A.; Bruncko, M.; Deckwerth, T.L.; Dinges, J.; Hajduk, P.J.; et al. An inhibitor of Bcl-2 family proteins induces regression of solid tumours. Nature 2005, 435, 677–681. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, D.; Velankar, M.; Brahmandam, M.; Hideshima, T.; Podar, K.; Richardson, P.; Schlossman, R.; Ghobrial, I.; Raje, N.; Munshi, N.; et al. A novel Bcl-2/Bcl-XL/Bcl-w inhibitor ABT-737 as therapy in multiple myeloma. Oncogene 2007, 26, 2374–2380. [Google Scholar] [CrossRef] [Green Version]
- Konopleva, M.; Contractor, R.; Tsao, T.; Samudio, I.; Ruvolo, P.P.; Kitada, S.; Deng, X.; Zhai, D.; Shi, Y.X.; Sneed, T.; et al. Mechanisms of apoptosis sensitivity and resistance to the BH3 mimetic ABT-737 in acute myeloid leukemia. Cancer Cell 2006, 10, 375–388. [Google Scholar] [CrossRef] [Green Version]
- Van Delft, M.F.; Wei, A.H.; Mason, K.D.; Vandenberg, C.J.; Chen, L.; Czabotar, P.E.; Willis, S.N.; Scott, C.L.; Day, C.L.; Cory, S.; et al. The BH3 mimetic ABT-737 targets selective Bcl-2 proteins and efficiently induces apoptosis via Bak/Bax if Mcl-1 is neutralized. Cancer Cell 2006, 10, 389–399. [Google Scholar] [CrossRef] [Green Version]
- Baggstrom, M.Q.; Qi, Y.; Koczywas, M.; Argiris, A.; Johnson, E.A.; Millward, M.J.; Murphy, S.C.; Erlichman, C.; Rudin, C.M.; Govindan, R.; et al. A phase II study of AT-101 (Gossypol) in chemotherapy-sensitive recurrent extensive-stage small cell lung cancer. J. Thorac. Oncol. Off. Publ. Int. Assoc. Study Lung Cancer 2011, 6, 1757–1760. [Google Scholar] [CrossRef] [Green Version]
- MacVicar, G.R.; Kuzel, T.M.; Curti, B.D.; Poiesz, B.; Somer, B.; Greco, F.A.; Gressler, V.; Brill, K.; Leopold, L. An open-label, multicenter, phase I/II study of AT-101 in combination with docetaxel (D) and prednisone (P) in men with hormone refractory prostate cancer (HRPC). J. Clin. Oncol. 2008, 26, 16043. [Google Scholar] [CrossRef]
- Tse, C.; Shoemaker, A.R.; Adickes, J.; Anderson, M.G.; Chen, J.; Jin, S.; Johnson, E.F.; Marsh, K.C.; Mitten, M.J.; Nimmer, P.; et al. ABT-263: A potent and orally bioavailable Bcl-2 family inhibitor. Cancer Res. 2008, 68, 3421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, A.; Gandhi, L.; O’Connor, O.A.; Rudin, C.M.; Khaira, D.; Xiong, H.; Chiu, Y.; Greco, R.; Krivoshik, A.P.; Wilson, W.H. Reduction in platelet counts as a mechanistic biomarker and guide for adaptive dose-escalation in phase I studies of the Bcl-2 family inhibitor ABT-263. J. Clin. Oncol. 2008, 26, 3542. [Google Scholar] [CrossRef]
- Wilson, W.H.; O’Connor, O.A.; Czuczman, M.S.; LaCasce, A.S.; Gerecitano, J.F.; Leonard, J.P.; Tulpule, A.; Dunleavy, K.; Xiong, H.; Chiu, Y.L.; et al. Navitoclax, a targeted high-affinity inhibitor of BCL-2, in lymphoid malignancies: A phase 1 dose-escalation study of safety, pharmacokinetics, pharmacodynamics, and antitumour activity. Lancet Oncol. 2010, 11, 1149–1159. [Google Scholar] [CrossRef] [Green Version]
- Mason, K.D.; Carpinelli, M.R.; Fletcher, J.I.; Collinge, J.E.; Hilton, A.A.; Ellis, S.; Kelly, P.N.; Ekert, P.G.; Metcalf, D.; Roberts, A.W.; et al. Programmed anuclear cell death delimits platelet life span. Cell 2007, 128, 1173–1186. [Google Scholar] [CrossRef] [Green Version]
- Cang, S.; Iragavarapu, C.; Savooji, J.; Song, Y.; Liu, D. ABT-199 (venetoclax) and BCL-2 inhibitors in clinical development. J. Hematol. Oncol. 2015, 8, 129. [Google Scholar] [CrossRef]
- Xiang, W.; Yang, C.-Y.; Bai, L. MCL-1 inhibition in cancer treatment. OncoTargets Ther. 2018, 11, 7301–7314. [Google Scholar] [CrossRef] [Green Version]
- Ocker, M.; Neureiter, D.; Lueders, M.; Zopf, S.; Ganslmayer, M.; Hahn, E.G.; Herold, C.; Schuppan, D. Variants of bcl-2 specific siRNA for silencing antiapoptotic bcl-2 in pancreatic cancer. Gut 2005, 54, 1298–1308. [Google Scholar] [CrossRef]
- Philchenkov, A.; Zavelevich, M.; Kroczak, T.J.; Los, M.J. Caspases and cancer: Mechanisms of inactivation and new treatment modalities. Exp. Oncol. 2004, 2, 82–97. [Google Scholar]
- Arap, W.; Pasqualini, R.; Ruoslahti, E. Cancer treatment by targeted drug delivery to tumor vasculature in a mouse model. Science 1998, 279, 377–380. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Fan, R.; Zou, X.; Gao, L.; Jin, H.; Du, R.; Xia, L.; Fan, D. Inhibitory effect of recombinant adenovirus carrying immunocaspase-3 on hepatocellular carcinoma. Biochem. Biophys. Res. Commun. 2007, 358, 489–494. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rabs | * Genetic Alterations | Localization | Cancer Type | Molecular Function | Ref(s) |
|---|---|---|---|---|---|
| Rab1a | Amplification | ER, Golgi | Liver | Inhibition of Rab1a (shRab1a) induces apoptosis in HCC | [76,77] |
| Rab2b | Amplification | Golgi | Pancreas | Inhibition of Rab2b (by miR-448) induces apoptosis in pancreatic cancer cells | [65,66] |
| Rab9 | Deletion/ Amplification | Late endosomes | Breast | Inhibition of Rab9 (siRab9) induces apoptosis | [83,87] |
| Rab10 | Amplification | ER | Liver | Inhibition of Rab10 (shRab10) induces apoptosis in HCC | [84,88] |
| Rab12 | Amplification | Golgi | Gastric | Inhibition of Rab12 (siRab12) promotes apoptosis | [85,89] |
| Rab13 | Amplification | Brain | Inhibition of Rab13 induces apoptosis in glioma cells | [80] | |
| Rab14 | Amplification | Golgi/TGN/Early endosomes | Gastric | Inhibition of Rab14 (shRab14) induces apoptosis in gastric cancer cells | [62,64] |
| Rab18 | Amplification | ER | Gastric | Overexpression of Rab18 cDNA inhibits apoptosis through the intrinsic pathway in gastric cancer cells | [69,90] |
| Rab21 | Amplification | Early endosomes | Brain | Inhibition of Rab21 (siRab21) induces apoptosis in glioma cells | [81,91] |
| Rab23 | Amplification | Plasma membrane | Breast | Rab23 elevates breast cancer cell apoptosis | [86,92] |
| Rab25 | Amplification | TGN, Apical recycling endosomes, | Breast | Overexpression of Rab25 cDNA inhibits apoptosis | [21,48] |
| Rab27a/b | Deletion/ Missense | Endosomal exocytic vesicles | Pancreas, Colorectal | Inhibition of Rab27a/b (siRab27a/b) induces apoptosis in pancreatic cells | [82,93,94] |
| Rab31 | Amplification | Late endosomes, Trans Golgi, TGN | Gastric Liver Brain/ Cervix | Overexpression of Rab31 cDNA inhibits apoptosis in gastric cancer cells Inhibition of Rab31 (siRab31) induces apoptosis in HCC Overexpression of Rab31 cDNA inhibits apoptosis in glioblastoma and cervical cancer cells | [57,58,59,95] |
| Rab35 | Amplification/ Missense mutation | Plasma membrane, Endosomes | Kidney | Overexpression of Rab35 cDNA expression suppresses apoptosis | [19] |
| Rab45 | Missense mutation | Perinuclear region | Leukemia | Overexpression of Rab45 cDNA induces apoptosis | [71,72] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gopal Krishnan, P.D.; Golden, E.; Woodward, E.A.; Pavlos, N.J.; Blancafort, P. Rab GTPases: Emerging Oncogenes and Tumor Suppressive Regulators for the Editing of Survival Pathways in Cancer. Cancers 2020, 12, 259. https://doi.org/10.3390/cancers12020259
Gopal Krishnan PD, Golden E, Woodward EA, Pavlos NJ, Blancafort P. Rab GTPases: Emerging Oncogenes and Tumor Suppressive Regulators for the Editing of Survival Pathways in Cancer. Cancers. 2020; 12(2):259. https://doi.org/10.3390/cancers12020259
Chicago/Turabian StyleGopal Krishnan, Priya D., Emily Golden, Eleanor A. Woodward, Nathan J. Pavlos, and Pilar Blancafort. 2020. "Rab GTPases: Emerging Oncogenes and Tumor Suppressive Regulators for the Editing of Survival Pathways in Cancer" Cancers 12, no. 2: 259. https://doi.org/10.3390/cancers12020259