Nuclear Lamins and Emerin Are Differentially Expressed in Osteosarcoma Cells and Scale with Tumor Aggressiveness

 , ,
, ,
Abstract
:1. Introduction
2. Results
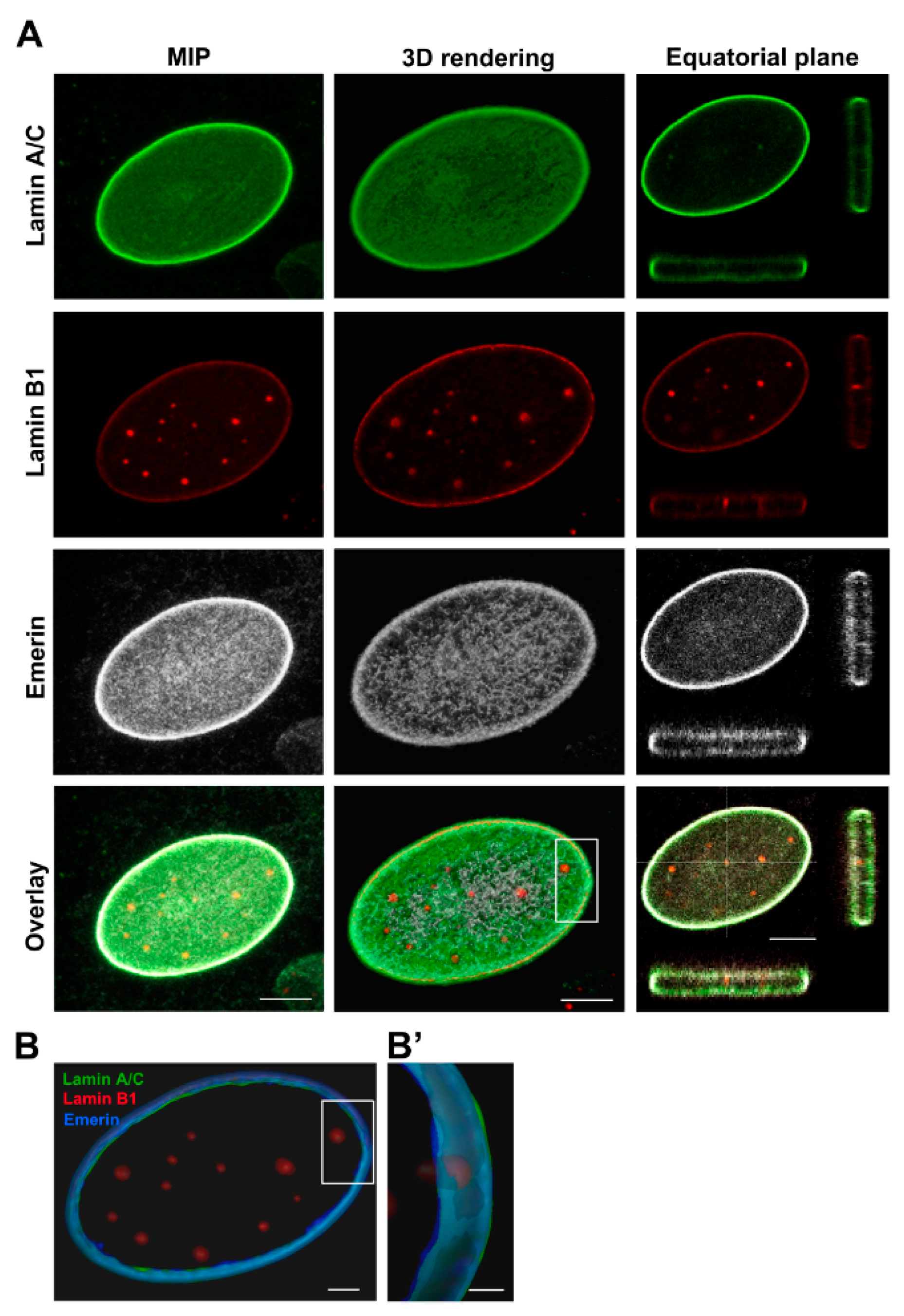
2.1. Osteosarcoma Cells Exhibit Laminopathic Nuclear Phenotypes
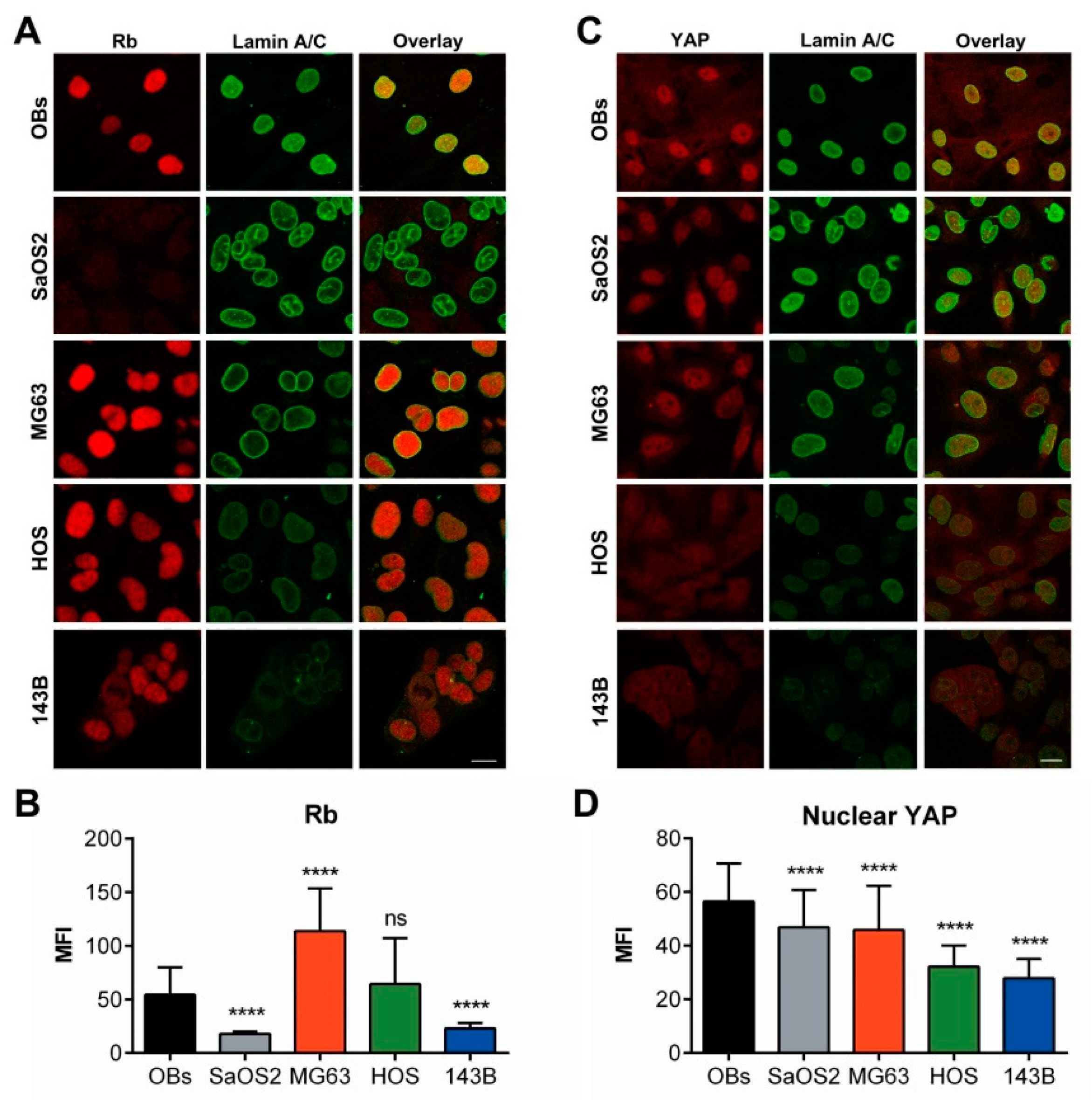
2.2. Differential Expression of A/C and B1 Lamins and Emerin in Osteosarcoma Cell Lines Correlates with Their Nucleoskeleton Architectures
2.2.1. Osteoblasts (OBs)
2.2.2. SaOS2
2.2.3. MG63
2.2.4. HOS
2.2.5. 143B
2.3. Functional Effects Mediated by Lamin A/C Modulation on Cell Aggressiveness
2.4. Prognostic Relevance of Lamins and Emerin Expression on a TMA
2.5. Linking Nucleoskeleton to Cytoskeleton: The Altered Nuclear Shuttling of MKL1 Follows the Lamin A/C-Emerin Behavior and the Polymerization Status of Actin
2.6. Functional Effects of Differential Expression of A-Type Lamin in Osteosarcoma Cells
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Culture
4.2. Immunofluorescence
4.3. Confocal Microscopy and Image Analysis
4.4. Western Blotting
4.5. RNA Interference Knockdown and Plasmid Over-Expression
4.6. XTT Proliferation Assay
4.7. Osteosarcoma Patient’s Samples on a Tissue MicroArray (TMA)
4.8. RNA Isolation, Reverse Transcription (RT) PCR Analysis
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Gerace, L.; Tapia, O. Messages from the voices within: Regulation of signaling by proteins of the nuclear lamina. Curr. Opin. Cell Boil. 2018, 52, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Gruenbaum, Y.; Foisner, R. Lamins: Nuclear Intermediate Filament Proteins with Fundamental Functions in Nuclear Mechanics and Genome Regulation. Annu. Rev. Biochem. 2015, 84, 131–164. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.; Worman, H.J. Structural organization of the human gene encoding nuclear lamin A and nuclear lamin C. J. Boil. Chem. 1993, 268, 16321–16326. [Google Scholar]
- Biamonti, G.; Giacca, M.; Perini, G.; Contreas, G.; Zentilin, L.; Weighardt, F.; Guerra, M.; Della Valle, G.; Saccone, S.; Riva, S. The gene for a novel human lamin maps at a highly transcribed locus of chromosome 19 which replicates at the onset of S-phase. Mol. Cell. Boil. 1992, 12, 3499–3506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furukawa, K.; Hotta, Y. cDNA cloning of a germ cell specific lamin B3 from mouse spermatocytes and analysis of its function by ectopic expression in somatic cells. EMBO J. 1993, 12, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.; Worman, H.J. Structural Organization of the Human Gene (LMNB1) Encoding Nuclear Lamin B1. Genomics 1995, 27, 230–236. [Google Scholar] [CrossRef]
- Constantinescu, D.; Gray, H.L.; Sammak, P.J.; Schatten, G.P.; Csoka, A.B. Lamin A/C expression is a marker of mouse and human embryonic stem cell differentiation. Stem Cells 2006, 24, 177–185. [Google Scholar] [CrossRef]
- Petrini, S.; Borghi, R.; D’Oria, V.; Restaldi, F.; Moreno, S.; Novelli, A.; Bertini, E.; Compagnucci, C. Aged induced pluripotent stem cell (iPSCs) as a new cellular model for studying premature aging. Aging 2017, 9, 1453–1466. [Google Scholar] [CrossRef] [Green Version]
- Sehgal, P.; Chaturvedi, P.; Kumaran, R.I.; Kumar, S.; Parnaik, V.K. Lamin A/C Haploinsufficiency Modulates the Differentiation Potential of Mouse Embryonic Stem Cells. PLoS ONE 2013, 8, e57891. [Google Scholar] [CrossRef] [Green Version]
- Ho, C.Y.; Lammerding, J. Lamins at a glance. J. Cell Sci. 2012, 125, 2087–2093. [Google Scholar] [CrossRef] [Green Version]
- Méjat, A.; Misteli, T. LINC complexes in health and disease. Nucleus 2010, 1, 40–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irianto, J.; Pfeifer, C.R.; Ivanovska, I.L.; Swift, J.; Discher, D.E. Nuclear Lamins in Cancer. Cell. Mol. Bioeng. 2016, 9, 258–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, S.; Comai, L. Lamin A, farnesylation and aging. Exp. Cell Res. 2012, 318, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, J.T.; Muchir, A.; Nagy, P.L.; Worman, H.J. LMNA cardiomyopathy: Cell biology and genetics meet clinical medicine. Dis. Model. Mech. 2011, 4, 562–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mercuri, E.; Poppe, M.; Quinlivan, R.; Messina, S.; Kinali, M.; Demay, L.; Bourke, J.; Richard, P.; Sewry, C.; Pike, M.; et al. Extreme variability of phenotype in patients with an identical missense mutation in the lamin A/C gene: From congenital onset with severe phenotype to milder classic Emery-Dreifuss variant. Arch. Neurol. 2004, 61, 690–694. [Google Scholar] [CrossRef] [Green Version]
- Scaffidi, P.; Misteli, T. Lamin A-dependent misregulation of adult stem cells associated with accelerated ageing. Nat. Cell Biol. 2008, 10, 452–459. [Google Scholar] [CrossRef] [Green Version]
- Coffeen, C.M. Genetic localization of an autosomal dominant leukodystrophy mimicking chronic progressive multiple sclerosis to chromosome 5q31. Hum. Mol. Genet. 2000, 9, 787–793. [Google Scholar] [CrossRef]
- Padiath, Q.S.; Saigoh, K.; Schiffmann, R.; Asahara, H.; Yamada, T.; Koeppen, A.; Hogan, K.; Ptáček, L.J.; Fu, Y.-H. Lamin B1 duplications cause autosomal dominant leukodystrophy. Nat. Genet. 2006, 38, 1114–1123. [Google Scholar] [CrossRef]
- Gao, J.; Li, Y.; Fu, X.; Luo, X. A Chinese patient with acquired partial lipodystrophy caused by a novel mutation with LMNB2 gene. J. Pediatr. Endocrinol. Metab. 2012, 25, 375–377. [Google Scholar] [CrossRef]
- Hegele, R.A.; Cao, H.; Liu, R.M.; Costain, G.A.; Charlton-Menys, V.; Rodger, N.W.; Durrington, P.N. Sequencing of the Reannotated LMNB2 Gene Reveals Novel Mutations in Patients with Acquired Partial Lipodystrophy. Am. J. Hum. Genet. 2006, 79, 383–389. [Google Scholar] [CrossRef] [Green Version]
- Helfand, B.T.; Wang, Y.; Pfleghaar, K.; Shimi, T.; Taimen, P.; Shumaker, D.K. Chromosomal regions associated with prostate cancer risk localize to lamin B-deficient microdomains and exhibit reduced gene transcription. J. Pathol. 2012, 226, 735–745. [Google Scholar] [CrossRef] [PubMed]
- Kaspi, E.; Frankel, D.; Guinde, J.; Perrin, S.; Laroumagne, S.; Robaglia-Schlupp, A.; Ostacolo, K.; Harhouri, K.; Tazi-Mezalek, R.; Micallef, J.; et al. Low lamin A expression in lung adenocarcinoma cells from pleural effusions is a pejorative factor associated with high number of metastatic sites and poor Performance status. PLoS ONE 2017, 12, e0183136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, L.; Schäfer, G.; Bu, H.; Zhang, Y.; Zhang, Y.; Klocker, H. Lamin A/C protein is overexpressed in tissue-invading prostate cancer and promotes prostate cancer cell growth, migration and invasion through the PI3K/AKT/PTEN pathway. Carcinogenesis 2012, 33, 751–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, S.O.; Park, S.-J.; Kim, W.; Park, S.G.; Kim, H.-J.; Kim, Y.-I.; Sohn, T.-S.; Noh, J.-H.; Jung, G. Proteome Analysis of Hepatocellular Carcinoma. Biochem. Biophys. Res. Commun. 2002, 291, 1031–1037. [Google Scholar] [CrossRef]
- Wazir, U.; Ahmed, M.H.; Bridger, J.M.; Harvey, A.; Jiang, W.G.; Sharma, A.K.; Mokbel, K. The clinicopathological significance of lamin A/C, lamin B1 and lamin B receptor mRNA expression in human breast cancer. Cell. Mol. Boil. Lett. 2013, 18, 595–611. [Google Scholar] [CrossRef]
- Ishikawa, Y.; Miller, R.W.; Machinami, R.; Sugano, H.; Goto, M. Atypical Osteosarcomas in Werner Syndrome (Adult Progeria). Jpn. J. Cancer Res. 2000, 91, 1345–1349. [Google Scholar] [CrossRef]
- King, C.R.; Lemmer, J.; Campbell, J.R.; Atkins, A.R. Osteosarcoma in a patient with Hutchinson-Gilford progeria. J. Med Genet. 1978, 15, 481–484. [Google Scholar] [CrossRef]
- Okamoto, N.; Satomura, K.; Hatsukawa, Y.; Hayashida, M.; Saijo, K.; Ohno, T.; Goto, M. Premature aging syndrome with osteosarcoma, cataracts, diabetes mellitus, osteoporosis, erythroid macrocytosis, severe growth and developmental deficiency. Am. J. Med Genet. 1997, 69, 169–170. [Google Scholar] [CrossRef]
- Shalev, S.A.; De Sandre-Giovannoli, A.; Shani, A.A.; Lévy, N. An association of Hutchinson–Gilford progeria and malignancy. Am. J. Med Genet. Part A 2007, 143, 1821–1826. [Google Scholar] [CrossRef]
- Damron, T.A.; Ward, W.G.; Stewart, A. Osteosarcoma, chondrosarcoma, and Ewing’s sarcoma: National Cancer Data Base Report. Clin. Orthop. Relat Res. 2007, 459, 40–47. [Google Scholar] [CrossRef]
- Sweetnam, R. Osteosarcoma. Br. J. Hosp. Med. 1982, 28, 116–121. [Google Scholar]
- Dunn, D.; Dehner, L.P. Metastatic osteosarcoma to lung. A clinicopathologic study of surgical biopsies and resections. Cancer 1977, 40, 3054–3064. [Google Scholar] [CrossRef]
- Kansara, M.; Thomas, D.M. Molecular Pathogenesis of Osteosarcoma. DNA Cell Boil. 2007, 26, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Ritter, J.; Bielack, S.S. Osteosarcoma. Ann. Oncol. 2010, 21, vii320–vii325. [Google Scholar] [CrossRef] [PubMed]
- Bermeo, S.; Vidal, C.; Zhou, H.; Duque, G. Lamin A/C Acts as an Essential Factor in Mesenchymal Stem Cell Differentiation Through the Regulation of the Dynamics of the Wnt/beta-Catenin Pathway. J. Cell Biochem. 2015, 116, 2344–2353. [Google Scholar] [CrossRef] [PubMed]
- Swift, J.; Ivanovska, I.L.; Buxboim, A.; Harada, T.; Dingal, P.C.D.P.; Pinter, J.; Pajerowski, J.D.; Spinler, K.R.; Shin, J.-W.; Tewari, M.; et al. Nuclear Lamin-A Scales with Tissue Stiffness and Enhances Matrix-Directed Differentiation. Science 2013, 341, 1240104. [Google Scholar] [CrossRef] [Green Version]
- Mohseny, A.B.; Machado, I.; Cai, Y.; Schaefer, K.-L.; Serra, M.; Hogendoorn, P.C.W.; Llombart-Bosch, A.; Cleton-Jansen, A.-M. Functional characterization of osteosarcoma cell lines provides representative models to study the human disease. Lab. Investig. 2011, 91, 1195–1205. [Google Scholar] [CrossRef]
- Urciuoli, E.; Giorda, E.; Scarsella, M.; Petrini, S.; Peruzzi, B. Osteosarcoma-derived extracellular vesicles induce a tumor-like phenotype in normal recipient cells. J. Cell. Physiol. 2018, 233, 6158–6172. [Google Scholar] [CrossRef]
- Parmacek, M.S. Myocardin-related transcription factors: Critical coactivators regulating cardiovascular development and adaptation. Circ. Res. 2007, 100, 633–644. [Google Scholar] [CrossRef]
- Ho, C.Y.; Jaalouk, D.E.; Vartiainen, M.K.; Lammerding, J. Lamin A/C and emerin regulate MKL1–SRF activity by modulating actin dynamics. Nature 2013, 497, 507–511. [Google Scholar] [CrossRef] [Green Version]
- Willer, M.K.; Carroll, C.W. Substrate stiffness-dependent regulation of the SRF-Mkl1 co-activator complex requires the inner nuclear membrane protein Emerin. J. Cell Sci. 2017, 130, 2111–2118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melcon, G.; Kozlov, S.; Hernandez, L.; Mitchell, S.; Nader, G.; Bakay, M.; Cutler, D.A.; Sullivan, T.; Zhao, P.; Rottman, J.N.; et al. Loss of emerin at the nuclear envelope disrupts the Rb1/E2F and MyoD pathways during muscle regeneration. Hum. Mol. Genet. 2006, 15, 637–651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, B.R.; Nitta, R.T.; Frock, R.L.; Mounkes, L.; Barbie, D.A.; Stewart, C.L.; Harlow, E.; Kennedy, B.K. A-type lamins regulate retinoblastoma protein function by promoting subnuclear localization and preventing proteasomal degradation. Proc. Natl. Acad. Sci. USA 2004, 101, 9677–9682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swift, J.; Discher, D.E. The nuclear lamina is mechano-responsive to ECM elasticity in mature tissue. J. Cell Sci. 2014, 127, 3005–3015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Romaih, K.; Bayani, J.; Vorobyova, J.; Karaskova, J.; Park, P.; Zielenska, M.; Squire, J. Chromosomal instability in osteosarcoma and its association with centrosome abnormalities. Cancer Genet. Cytogenet. 2003, 144, 91–99. [Google Scholar] [CrossRef]
- Martin, J.W.; Squire, J.A.; Zielenska, M. The Genetics of Osteosarcoma. Sarcoma 2012, 2012, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Weatherby, R.P.; Dahlin, D.C.; Ivins, J.C. Postradiation sarcoma of bone: Review of 78 Mayo Clinic cases. Mayo Clin. Proc. 1981, 56, 294–306. [Google Scholar]
- Therizols, P.; Fairhead, C.; Cabal, G.G.; Genovesio, A.; Olivo-Marin, J.C.; Dujon, B.; Fabre, E. Telomere tethering at the nuclear periphery is essential for efficient DNA double strand break repair in subtelomeric region. J. Cell Biol. 2006, 172, 189–199. [Google Scholar] [CrossRef]
- Campisi, J. Cellular senescence as a tumor-suppressor mechanism. Trends Cell Boil. 2001, 11, 27. [Google Scholar] [CrossRef] [Green Version]
- Hoeijmakers, J.H.J. Genome maintenance mechanisms for preventing cancer. Nature 2001, 411, 366–374. [Google Scholar] [CrossRef]
- Meng, E.; Mitra, A.; Tripathi, K.; Finan, M.A.; Scalici, J.; McClellan, S.; Da Silva, L.M.; Reed, I.; Shevde, L.A.; Palle, K.; et al. ALDH1A1 Maintains Ovarian Cancer Stem Cell-Like Properties by Altered Regulation of Cell Cycle Checkpoint and DNA Repair Network Signaling. PLoS ONE 2014, 9, e107142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakthivel, K.M.; Sehgal, P. A Novel Role of Lamins from Genetic Disease to Cancer Biomarkers. Oncol. Rev. 2016, 10, 309. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, K.; Mani, C.; Barnett, R.; Nalluri, S.; Bachaboina, L.; Rocconi, R.P.; Athar, M.; Owen, L.B.; Palle, K. Gli1 Regulates S-phase Checkpoint in Tumor Cells via Bid and its Inhibition Sensitizes to DNA Topoisomerase 1 Inhibitors. J. Biol. Chem. 2014, 289, 31513–31525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Booth-Gauthier, E.A.; Du, V.; Ghibaudo, M.; Rape, A.D.; Dahl, K.N.; Ladoux, B. Hutchinson–Gilford progeria syndrome alters nuclear shape and reduces cell motility in three dimensional model substrates. Integr. Boil. 2013, 5, 569. [Google Scholar] [CrossRef] [PubMed]
- Alhudiri, I.M.; Nolan, C.C.; Ellis, I.O.; Elzagheid, A.; Rakha, E.A.; Green, A.R.; Chapman, C.J. Expression of Lamin A/C in early-stage breast cancer and its prognostic value. Breast Cancer Res. Treat. 2019, 174, 661–668. [Google Scholar] [CrossRef] [PubMed]
- Guinde, J.; Frankel, D.; Perrin, S.; Delecourt, V.; Lévy, N.; Barlesi, F.; Astoul, P.; Roll, P.; Kaspi, E. Lamins in Lung Cancer: Biomarkers and Key Factors for Disease Progression through miR-9 Regulation? Cells 2018, 7, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Z.; Wu, L.; Weng, D.; Xu, D.; Geng, J.; Zhao, F. Reduced expression of lamin A/C correlates with poor histological differentiation and prognosis in primary gastric carcinoma. J. Exp. Clin. Cancer Res. 2009, 28, 8. [Google Scholar] [CrossRef] [Green Version]
- Finch-Edmondson, M.; Sudol, M. Framework to function: Mechanosensitive regulators of gene transcription. Cell. Mol. Boil. Lett. 2016, 21, 28. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age | Cases/Total | % |
|---|---|---|
| ≤18 years | 25/50 | 50% |
| >18 years | 25/50 | 50% |
| Gender | ||
| Male | 37/50 | 74% |
| Female | 13/50 | 26% |
| Type of Osteosarcoma | ||
| Conventional osteosarcoma: | 29/50 | 58% |
| Osteoblastic type | 20/29 | 69% |
| Chondroblastic type | 8/29 | 28% |
| Fibroblastic type | 1/29 | 3% |
| Telangiectatic osteosarcoma | 1/50 | 2% |
| Extraskeletal osteosarcoma | 2/50 | 4% |
| Metastatic osteosarcoma, chondroblastic type | 1/50 | 2% |
| Synovial sarcoma | 1/50 | 2% |
| Granulation tissue | 1/50 | 2% |
| Radiation-induced sarcoma | 1/50 | 2% |
| Osteosarcoma (not defined) | 15/50 | 30% |
| Site of Osteosarcoma | ||
| Bone, femur | 23/50 | 46% |
| Bone, tibia | 7/50 | 14% |
| Bone, fibula | 2/50 | 4% |
| Bone, humerus | 3/50 | 6% |
| Bone, pelvis | 2/50 | 4% |
| Bone, ilium | 2/50 | 4% |
| Bone, mandible | 2/50 | 4% |
| Bone, tibia and fibula | 2/50 | 4% |
| Bone, calcaneus | 1/50 | 2% |
| Mediastinum | 1/50 | 2% |
| Knee | 1/50 | 2% |
| Hip | 1/50 | 2% |
| Leg | 1/50 | 2% |
| Soft tissue | 2/50 | 4% |
| Overall Survival | ||
| Alive (beyond 5 year-follow up) | 29/50 | 48.30% |
| Dead (within 5 year-follow up) | 29/50 | 48.30% |
| Gene | Sequences | Product Size |
|---|---|---|
| Human LMNA | left: CTACACCAGCCAACCCAGAT | 126 bp |
| right: GGTCGAAGGACAGAGACTGC | ||
| Human LMNB | left: AACGAGACCAGAAGGAAGCA | 117 bp |
| right: GGCATCATGTTGCTCTCTCA | ||
| EMERIN | left: CAGTGCCTACCAGAGCATCA | 232 bp |
| right: AAAGACCAGGAAAAGCAGCA | ||
| GAPDH | left: CGACCACTTTGTCAAGCTCA | 228 bp |
| right: AGGGGTCTACATGGCAACTG |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Urciuoli, E.; Petrini, S.; D’Oria, V.; Leopizzi, M.; Della Rocca, C.; Peruzzi, B. Nuclear Lamins and Emerin Are Differentially Expressed in Osteosarcoma Cells and Scale with Tumor Aggressiveness. Cancers 2020, 12, 443. https://doi.org/10.3390/cancers12020443
Urciuoli E, Petrini S, D’Oria V, Leopizzi M, Della Rocca C, Peruzzi B. Nuclear Lamins and Emerin Are Differentially Expressed in Osteosarcoma Cells and Scale with Tumor Aggressiveness. Cancers. 2020; 12(2):443. https://doi.org/10.3390/cancers12020443
Chicago/Turabian StyleUrciuoli, Enrica, Stefania Petrini, Valentina D’Oria, Martina Leopizzi, Carlo Della Rocca, and Barbara Peruzzi. 2020. "Nuclear Lamins and Emerin Are Differentially Expressed in Osteosarcoma Cells and Scale with Tumor Aggressiveness" Cancers 12, no. 2: 443. https://doi.org/10.3390/cancers12020443
APA StyleUrciuoli, E., Petrini, S., D’Oria, V., Leopizzi, M., Della Rocca, C., & Peruzzi, B. (2020). Nuclear Lamins and Emerin Are Differentially Expressed in Osteosarcoma Cells and Scale with Tumor Aggressiveness. Cancers, 12(2), 443. https://doi.org/10.3390/cancers12020443