Blood Monocyte Subsets with Activation Markers in Relation with Macrophages in Non-Small Cell Lung Cancer

 ,
, 
 ,
,
Abstract
Simple Summary
Abstract
1. Introduction
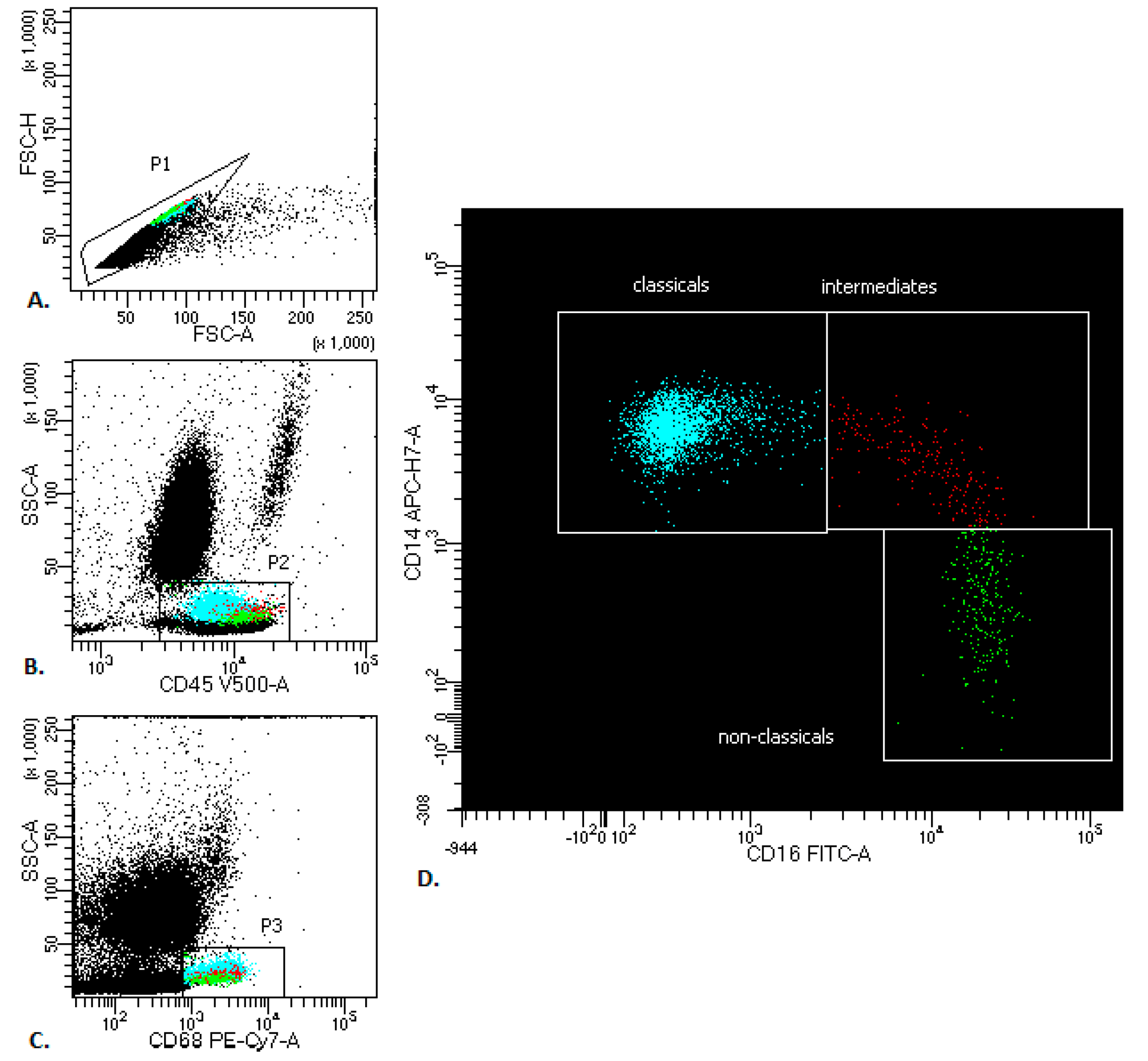
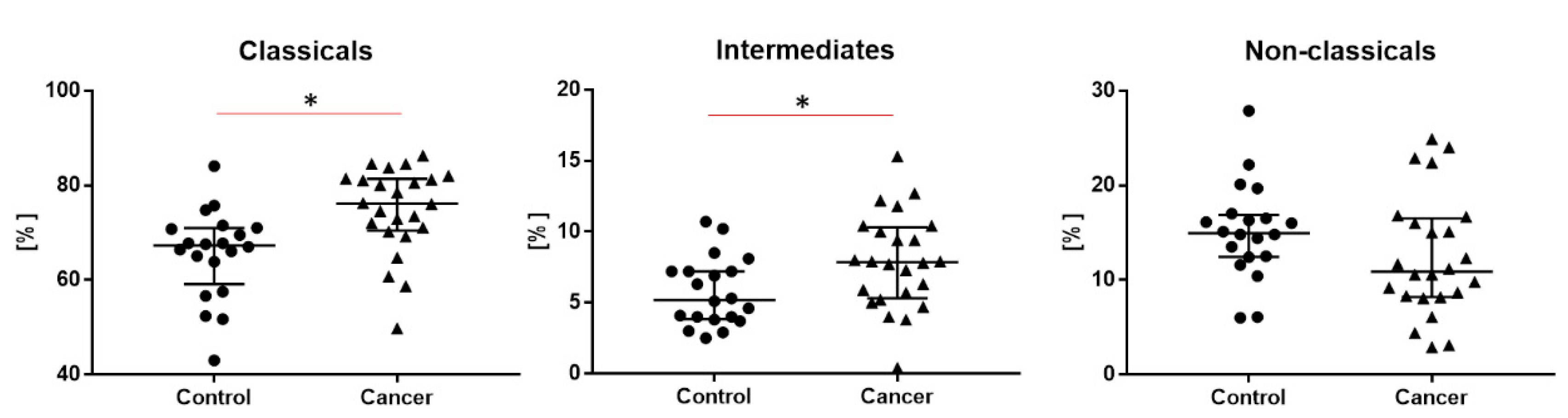
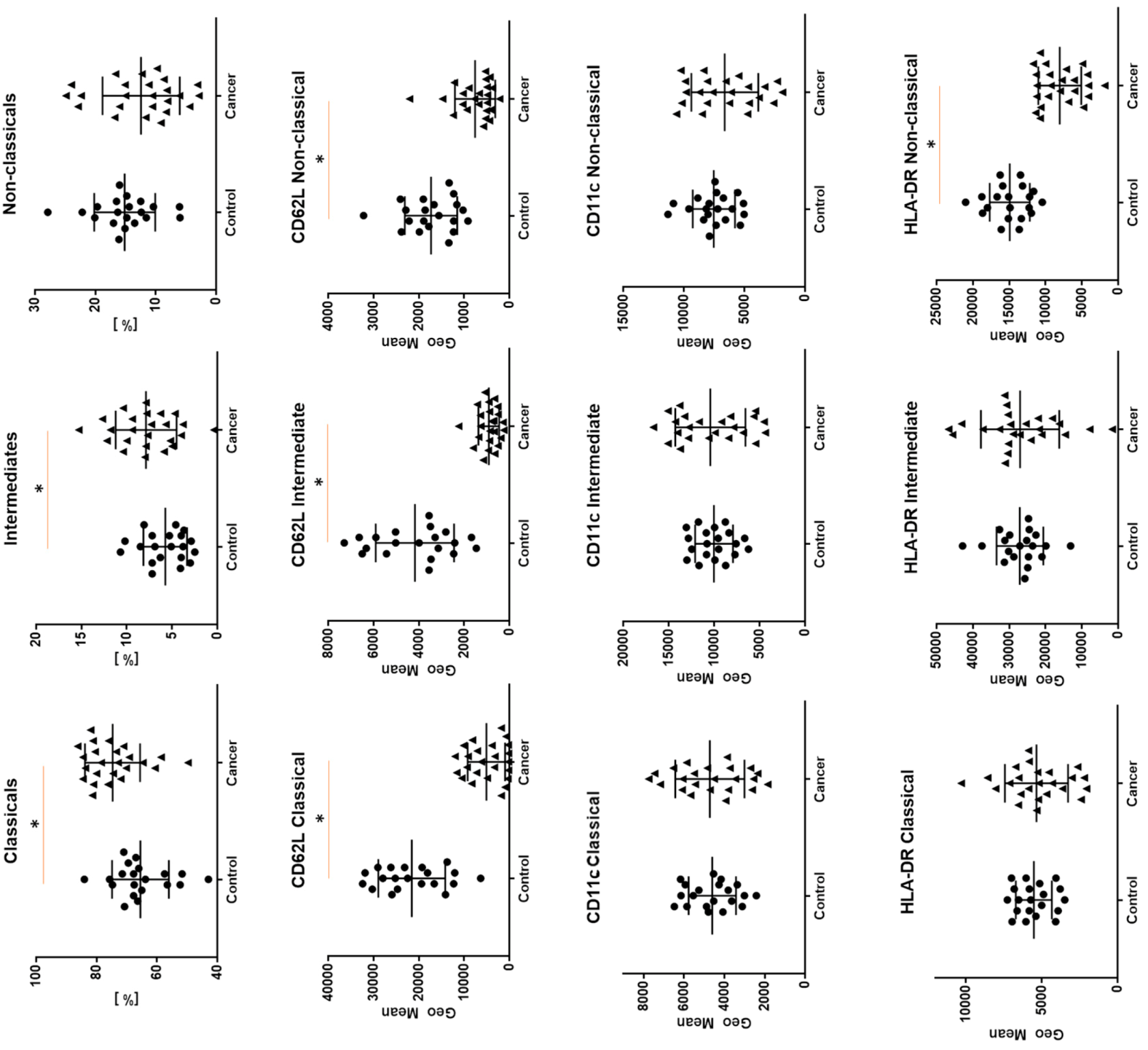
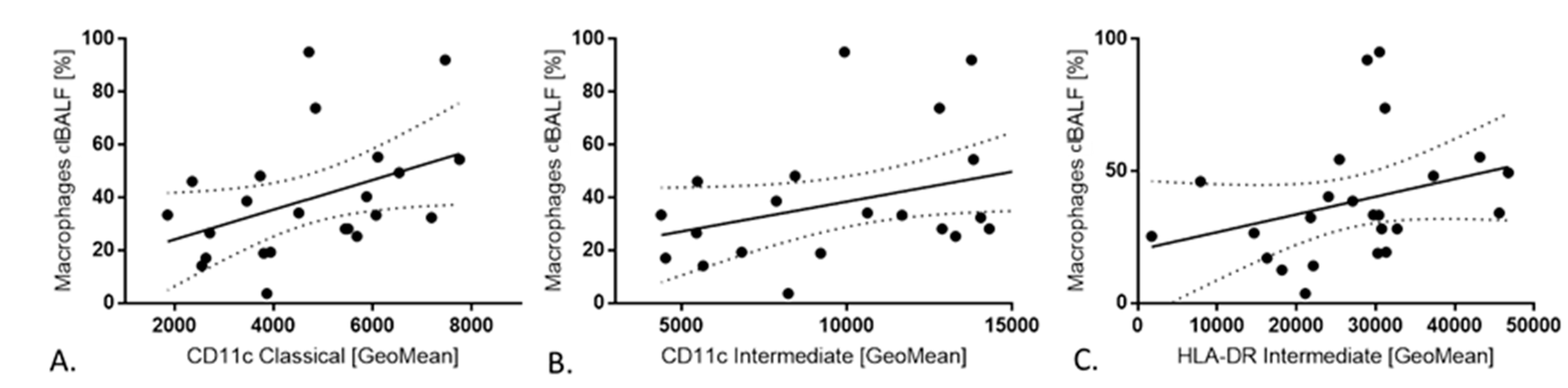
2. Results
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. Material
4.3. Flow Cytometry Analysis
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Olingy, C.E.; Dinh, H.Q.; Hedrick, C.C. Monocyte heterogeneity and functions in cancer. J. Leukoc. Biol. 2019, 106, 309–322. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A. Macrophage diversity and polarization: In vivo veritas. Blood 2006, 108, 408–409. [Google Scholar] [CrossRef]
- Shapouri-Moghaddam, A.; Mohammadian, S.; Vazini, H.; Taghadosi, M.; Esmaeili, S.A.; Mardani, F.; Seifi, B.; Mohammadi, A.; Afshari, J.T.; Sahebkar, A. Macrophage plasticity, polarization, and function in health and disease. J. Cell. Physiol. 2018, 233, 6425–6440. [Google Scholar] [CrossRef] [PubMed]
- Vuk-Pavlovic, S.; Bulur, P.A.; Lin, Y.; Qin, R.; Szumlanski, C.L.; Zhao, X.; Dietz, A.B. Immunosuppressive CD14+HLA-DRlow/- monocytes in prostate cancer. Prostate 2010, 70, 443–455. [Google Scholar] [CrossRef] [PubMed]
- Brooks, N.; Stojanovska, L.; Grant, P.; Apostolopoulos, V.; McDonald, C.F.; Pouniotis, D.S. Characterization of blood monocyte phenotype in patients with endometrial cancer. Int. J. Gynecol. Cancer Off. J. Int. Gynecol. Cancer Soc. 2012, 22, 1500–1508. [Google Scholar] [CrossRef]
- Mariotta, S.; Aquilini, M.; Ricci, A.; Papale, M.; Pabani, R.; Sposato, B.; Mannino, F. Changes in monocyte phagocyting activity after multi-agent chemotherapy in non-small cell lung cancer. Eur. Rev. Med Pharmacol. Sci. 2002, 6, 67–73. [Google Scholar]
- Yang, J.; Zhang, L.; Yu, C.; Yang, X.F.; Wang, H. Monocyte and macrophage differentiation: Circulation inflammatory monocyte as biomarker for inflammatory diseases. Biomark. Res. 2014, 2, 1. [Google Scholar] [CrossRef]
- Stansfield, B.K.; Ingram, D.A. Clinical significance of monocyte heterogeneity. Clin. Transl. Med. 2015, 4, 5. [Google Scholar] [CrossRef]
- Marimuthu, R.; Francis, H.; Dervish, S.; Li, S.C.H.; Medbury, H.; Williams, H. Characterization of human monocyte subsets by whole blood flow cytometry analysis. J. Vis. Exp. 2018. [Google Scholar] [CrossRef]
- Nicholson, I.C. CD62L (L-selectin). J. Biol. Regul. Homeost. Agents 2002, 16, 144–146. [Google Scholar]
- Adams, D.H.; Shaw, S. Leucocyte-endothelial interactions and regulation of leucocyte migration. Lancet 1994, 343, 831–836. [Google Scholar] [CrossRef]
- Randolph, G.J.; Beaulieu, S.; Lebecque, S.; Steinman, R.M.; Muller, W.A. Differentiation of monocytes into dendritic cells in a model of transendothelial trafficking. Science 1998, 282, 480–483. [Google Scholar] [CrossRef] [PubMed]
- Geissmann, F.; Jung, S.; Littman, D.R. Blood monocytes consist of two principal subsets with distinct migratory properties. Immunity 2003, 19, 71–82. [Google Scholar] [CrossRef]
- Sandor, N.; Lukacsi, S.; Ungai-Salanki, R.; Orgovan, N.; Szabo, B.; Horvath, R.; Erdei, A.; Bajtay, Z. CD11c/CD18 dominates adhesion of human monocytes, macrophages and dendritic cells over CD11b/CD18. PLoS ONE 2016, 11, e0163120. [Google Scholar] [CrossRef]
- Arnaout, M.A. Structure and function of the leukocyte adhesion molecules CD11/CD18. Blood 1990, 75, 1037–1050. [Google Scholar] [CrossRef]
- Schittenhelm, L.; Hilkens, C.M.; Morrison, V.L. β2 Integrins as regulators of dendritic cell, monocyte, and macrophage function. Front. Immunol. 2017, 8, 1866. [Google Scholar] [CrossRef]
- Tan, S.M. The leucocyte β2 (CD18) integrins: The structure, functional regulation and signalling properties. Biosci. Rep. 2012, 32, 241–269. [Google Scholar] [CrossRef]
- Schmidt, S.; Moser, M.; Sperandio, M. The molecular basis of leukocyte recruitment and its deficiencies. Mol. Immunol. 2013, 55, 49–58. [Google Scholar] [CrossRef]
- Parham, P.; Ohta, T. Population biology of antigen presentation by MHC class I molecules. Science 1996, 272, 67–74. [Google Scholar] [CrossRef]
- Frazier, W.J.; Hall, M.W. Immunoparalysis and adverse outcomes from critical illness. Pediatr. Clin. N. Am. 2008, 55, 647–668. [Google Scholar] [CrossRef]
- Hai, Y.; Chen, N.; Wu, W.; Wang, Z.; Lin, F.; Guo, C.; Liu, C.; Li, W.; Liu, L. High postoperative monocyte indicates inferior Clinicopathological characteristics and worse prognosis in lung adenocarcinoma or squamous cell carcinoma after lobectomy. BMC Cancer 2018, 18, 1011. [Google Scholar] [CrossRef] [PubMed]
- Zawada, A.M.; Rogacev, K.S.; Schirmer, S.H.; Sester, M.; Bohm, M.; Fliser, D.; Heine, G.H. Monocyte heterogeneity in human cardiovascular disease. Immunobiology 2012, 217, 1273–1284. [Google Scholar] [CrossRef] [PubMed]
- Rivas-Fuentes, S.; Iglesias, A.H.; Trejo, A.G.; Castro, D.Y.C.; Figueroa, N.I.; Perez, T.A.; Saldana, R.B.; Rosete, P.G. Restoration of peripheral intermediate and classical monocytes expressing HLA-DR in patients with lung adenocarcinoma after platinum-based chemotherapy. Technol. Cancer Res. Treat. 2018, 17. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.L.; Tai, J.J.; Wong, W.C.; Han, H.; Sem, X.; Yeap, W.H.; Kourilsky, P.; Wong, S.C. Gene expression profiling reveals the defining features of the classical, intermediate, and nonclassical human monocyte subsets. Blood 2011, 118, e16–e31. [Google Scholar] [CrossRef] [PubMed]
- Gren, S.T.; Rasmussen, T.B.; Janciauskiene, S.; Hakansson, K.; Gerwien, J.G.; Grip, O. A single-cell gene-expression profile reveals inter-cellular heterogeneity within human monocyte subsets. PLoS ONE 2015, 10, e0144351. [Google Scholar] [CrossRef] [PubMed]
- Hanna, R.N.; Cekic, C.; Sag, D.; Tacke, R.; Thomas, G.D.; Nowyhed, H.; Herrley, E.; Rasquinha, N.; McArdle, S.; Wu, R.; et al. Patrolling monocytes control tumor metastasis to the lung. Science 2015, 350, 985–990. [Google Scholar] [CrossRef]
- Menezes, S.; Melandri, D.; Anselmi, G.; Perchet, T.; Loschko, J.; Dubrot, J.; Patel, R.; Gautier, E.L.; Hugues, S.; Longhi, M.P.; et al. The heterogeneity of Ly6C(hi) monocytes controls their differentiation into iNOS(+) macrophages or monocyte-derived dendritic cells. Immunity 2016, 45, 1205–1218. [Google Scholar] [CrossRef]
- Lee, J.; Tam, H.; Adler, L.; Ilstad-Minnihan, A.; Macaubas, C.; Mellins, E.D. The MHC class II antigen presentation pathway in human monocytes differs by subset and is regulated by cytokines. PLoS ONE 2017, 12, e0183594. [Google Scholar] [CrossRef]
- Fingerle, G.; Pforte, A.; Passlick, B.; Blumenstein, M.; Strobel, M.; Ziegler-Heitbrock, H.W. The novel subset of CD14+/CD16+ blood monocytes is expanded in sepsis patients. Blood 1993, 82, 3170–3176. [Google Scholar] [CrossRef]
- Nockher, W.A.; Scherberich, J.E. Expanded CD14+ CD16+ monocyte subpopulation in patients with acute and chronic infections undergoing hemodialysis. Infect. Immun. 1998, 66, 2782–2790. [Google Scholar] [CrossRef]
- Skrzeczynska-Moncznik, J.; Bzowska, M.; Loseke, S.; Grage-Griebenow, E.; Zembala, M.; Pryjma, J. Peripheral blood CD14high CD16+ monocytes are main producers of IL-10. Scand. J. Immunol. 2008, 67, 152–159. [Google Scholar] [CrossRef]
- Abeles, R.D.; McPhail, M.J.; Sowter, D.; Antoniades, C.G.; Vergis, N.; Vijay, G.K.; Xystrakis, E.; Khamri, W.; Shawcross, D.L.; Ma, Y.; et al. CD14, CD16 and HLA-DR reliably identifies human monocytes and their subsets in the context of pathologically reduced HLA-DR expression by CD14(hi)/CD16(neg) monocytes: Expansion of CD14(hi) /CD16(pos) and contraction of CD14(lo) /CD16(pos) monocytes in acute liver failure. Cytom. Part A J. Int. Soc. Anal. Cytol. 2012, 81, 823–834. [Google Scholar] [CrossRef]
- Mukherjee, R.; Kanti Barman, P.; Kumar Thatoi, P.; Tripathy, R.; Kumar Das, B.; Ravindran, B. Non-classical monocytes display inflammatory features: Validation in Sepsis and Systemic Lupus Erythematous. Sci. Rep. 2015, 5, 13886. [Google Scholar] [CrossRef] [PubMed]
- Thomas, G.D.; Hamers, A.A.J.; Nakao, C.; Marcovecchio, P.; Taylor, A.M.; McSkimming, C.; Nguyen, A.T.; McNamara, C.A.; Hedrick, C.C. Human blood monocyte subsets: A new gating strategy defined using cell surface markers identified by mass cytometry. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 1548–1558. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Manivannan, A.; Crane, I.; Dawson, R.; Liversidge, J. Critical but divergent roles for CD62L and CD44 in directing blood monocyte trafficking in vivo during inflammation. Blood 2008, 112, 1166–1174. [Google Scholar] [CrossRef] [PubMed]
- Almatroodi, S.A.; McDonald, C.F.; Collins, A.L.; Darby, I.A.; Pouniotis, D.S. Blood classical monocytes phenotype is not altered in primary non-small cell lung cancer. World J. Clin. Oncol. 2014, 5, 1078–1087. [Google Scholar] [CrossRef]
- Chikamatsu, K.; Sakakura, K.; Toyoda, M.; Takahashi, K.; Yamamoto, T.; Masuyama, K. Immunosuppressive activity of CD14+ HLA-DR- cells in squamous cell carcinoma of the head and neck. Cancer Sci. 2012, 103, 976–983. [Google Scholar] [CrossRef]
- Xiu, B.; Lin, Y.; Grote, D.M.; Ziesmer, S.C.; Gustafson, M.P.; Maas, M.L.; Zhang, Z.; Dietz, A.B.; Porrata, L.F.; Novak, A.J.; et al. IL-10 induces the development of immunosuppressive CD14(+)HLA-DR(low/-) monocytes in B-cell non-Hodgkin lymphoma. Blood Cancer J. 2015, 5, e328. [Google Scholar] [CrossRef]
- Huang, A.; Zhang, B.; Wang, B.; Zhang, F.; Fan, K.X.; Guo, Y.J. Increased CD14(+)HLA-DR (-/low) myeloid-derived suppressor cells correlate with extrathoracic metastasis and poor response to chemotherapy in non-small cell lung cancer patients. Cancer Immunol. Immunother. CII 2013, 62, 1439–1451. [Google Scholar] [CrossRef]
- Tian, T.; Gu, X.; Zhang, B.; Liu, Y.; Yuan, C.; Shao, L.; Guo, Y.; Fan, K. Increased circulating CD14(+)HLA-DR-/low myeloid-derived suppressor cells are associated with poor prognosis in patients with small-cell lung cancer. Cancer Biomark. Sect. A Dis. Markers 2015, 15, 425–432. [Google Scholar] [CrossRef]
- Franklin, R.A.; Liao, W.; Sarkar, A.; Kim, M.V.; Bivona, M.R.; Liu, K.; Pamer, E.G.; Li, M.O. The cellular and molecular origin of tumor-associated macrophages. Science 2014, 344, 921–925. [Google Scholar] [CrossRef] [PubMed]
- Sidibe, A.; Ropraz, P.; Jemelin, S.; Emre, Y.; Poittevin, M.; Pocard, M.; Bradfield, P.F.; Imhof, B.A. Angiogenic factor-driven inflammation promotes extravasation of human proangiogenic monocytes to tumours. Nat. Commun. 2018, 9, 355. [Google Scholar] [CrossRef] [PubMed]
- Detterbeck, F.C. The eighth edition TNM stage classification for lung cancer: What does it mean on main street? J. Thorac. Cardiovasc. Surg. 2018, 155, 356–359. [Google Scholar] [CrossRef] [PubMed]
- Chcialowski, A.; Chorostowska-Wynimko, J.; Fal, A.; Pawlowicz, R.; Domagala-Kulawik, J. Recommendation of the Polish Respiratory Society for bronchoalveolar lavage (BAL) sampling, processing and analysis methods. Pneumonol. Alergol. Pol. 2011, 79, 75–89. [Google Scholar] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | n | Age (yr) Mean ± SD | Sex M/F | Stage I/II/III/IV | Subtypes AD/SQCC/LCC/AS |
|---|---|---|---|---|---|
| Cancer | 24 | 69.3 ± 6.4 | 9/15 | 15/6/3/0 | 16/5/2/1 |
| Excluded from the study: | |||||
| - no cancer cells | 10 | 71.4 ± 8.3 | 6/4 | - | - |
| - metastatic cells | 4 | 60.5 ± 10.8 | 1/3 | - | - |
| Control group | 20 | 51.9 ± 9.8 | 1/10 | - | - |
| Cells | Study Group PB % Median (Q1–Q3) | Control Group PB % Median (Q1–Q3) | Study Group cBALF % Median (Q1–Q3) | Study Group hBALF % Median (Q1–Q3) |
|---|---|---|---|---|
| All Monocytes | 7.1 (5.9–8.6) | 6.4 (4.4–7.5) | - | - |
| Classical monocytes * CD14++CD16- | 76.2 (70.7–81.3) * | 67.3 (60.7–70.9) * | - | - |
| Intermediate monocytes * CD14+CD16+ | 7.9 (5.5–10.2) * | 5.2 (3.9–0.2) * | - | - |
| Non classical monocytes CD14-/+CD16++ | 10.9 (8.3–16.4) | 14.9 (12.5–16.8) | - | - |
| Macrophages * CD68++ | - | - | 33.4 (22.3–48.8) * | 22.4 (11.5–30.9) * |
| Expression of: | Classical Monocytes a Geometric Mean (Median (Q1–Q3)) % (Median (Q1–Q3)) | Intermediate Monocytes b Geometric Mean (Median (Q1–Q3)) % (Median (Q1–Q3)) | Non-classical Monocytes c Geometric Mean (Median (Q1–Q3)) % (Median (Q1–Q3)) | Kruskal–Wallis Test p < 0.05 * |
|---|---|---|---|---|
| CD62L | 4425.5 (879.5–8793.5) | 883.5 (509–1220) | 556.5 (494.5–983.5) | 0.0007 *a–b, a–c |
| 95.0 (80.2–98.6) | 43.4 (26–65.7) | 26.7 (14.1–50.2) | 0.0001 *a–b, a–c | |
| CD11c | 4712 (3460–6069) | 10623 (6820–13828) | 6541 (4637-8732) | 0.0000 *a–b, b–c |
| 98 (96–99) | 100 (99.1–100) | 95.1 (87.9–98.1) | - | |
| CD18 | 1923 (1572.5–2407.5) | 2497.5 (1926.5–2993.5) | 1440 (1178–1831.5) | 0.0000 *a–c, b–c |
| 80.6 (48.7–94.3) | 96.9 (84.9–99.5) | 44.9 (34.7–62) | 0.0000 *a–b, b–c | |
| HLA-DR | 5314 (3977.5–6540) | 29320 (21456–31255) | 8654.5 (5483.5–10761.5) | 0.0000 *a–b, b–c |
| 96.4 (97.5–99.6) | 99.8 (97.9–100) | 90.6 (83–98) | - |
| Expression of: | Classical Monocytes a Geometric Mean (Median (Q1–Q3)) % (Median (Q1–Q3)) | Intermediate Monocytes b Geometric Mean (Median (Q1–Q3)) % (Median (Q1–Q3)) | Non classical Monocytes c Geometric Mean (Median (Q1–Q3)) % (Median (Q1–Q3)) | Kruskal–Wallis Test p < 0.05 * |
|---|---|---|---|---|
| CD62L | 22850.5 (15397.5–27104) | 3556. 5 (2983–5670) | 1727 (1237.5–2104.5) | 0.0000 * a–b–c |
| 95.6 (90.9–97.4) | 35.4 (28.3–50.9) | 16.2 (10.4–24.0) | 0.0001 * a–b–c | |
| CD11c | 4536 (3727–5703) | 9954 (8576–11728.5) | 7396.5 (6297.5–8275) | 0.0000 * a–b–c |
| 82.9 (70.9–92.9) | 88 (82.1–97.6) | 85.6 (76.9–89.1) | - | |
| CD18 | 1730 (1460–2350.5) | 2284 (1899–2891) | 1499 (1210–1922) | 0.0000 *a–c, b–c |
| 81.4 (50.1–96.1) | 95.8 (85.5–99.1) | 41.9 (37.1–69.2) | 0.0000 *a–b, b–c | |
| HLA–DR | 5777.5 (4495–6582) | 25483.5 (24065–30712.5) | 14999.5 (12732.5–16474) | 0.0000 * a–b–c |
| 69.3 (57.2–77.1) | 93.3 (85.1–96.45) | 77.8 (70.6–84.6) | 0.0001 * a–b, b–c |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kwiecień, I.; Rutkowska, E.; Polubiec-Kownacka, M.; Raniszewska, A.; Rzepecki, P.; Domagała-Kulawik, J. Blood Monocyte Subsets with Activation Markers in Relation with Macrophages in Non-Small Cell Lung Cancer. Cancers 2020, 12, 2513. https://doi.org/10.3390/cancers12092513
Kwiecień I, Rutkowska E, Polubiec-Kownacka M, Raniszewska A, Rzepecki P, Domagała-Kulawik J. Blood Monocyte Subsets with Activation Markers in Relation with Macrophages in Non-Small Cell Lung Cancer. Cancers. 2020; 12(9):2513. https://doi.org/10.3390/cancers12092513
Chicago/Turabian StyleKwiecień, Iwona, Elżbieta Rutkowska, Małgorzata Polubiec-Kownacka, Agata Raniszewska, Piotr Rzepecki, and Joanna Domagała-Kulawik. 2020. "Blood Monocyte Subsets with Activation Markers in Relation with Macrophages in Non-Small Cell Lung Cancer" Cancers 12, no. 9: 2513. https://doi.org/10.3390/cancers12092513
APA StyleKwiecień, I., Rutkowska, E., Polubiec-Kownacka, M., Raniszewska, A., Rzepecki, P., & Domagała-Kulawik, J. (2020). Blood Monocyte Subsets with Activation Markers in Relation with Macrophages in Non-Small Cell Lung Cancer. Cancers, 12(9), 2513. https://doi.org/10.3390/cancers12092513