Connecting the Missing Dots: ncRNAs as Critical Regulators of Therapeutic Susceptibility in Breast Cancer




Simple Summary
Abstract
1. Introduction
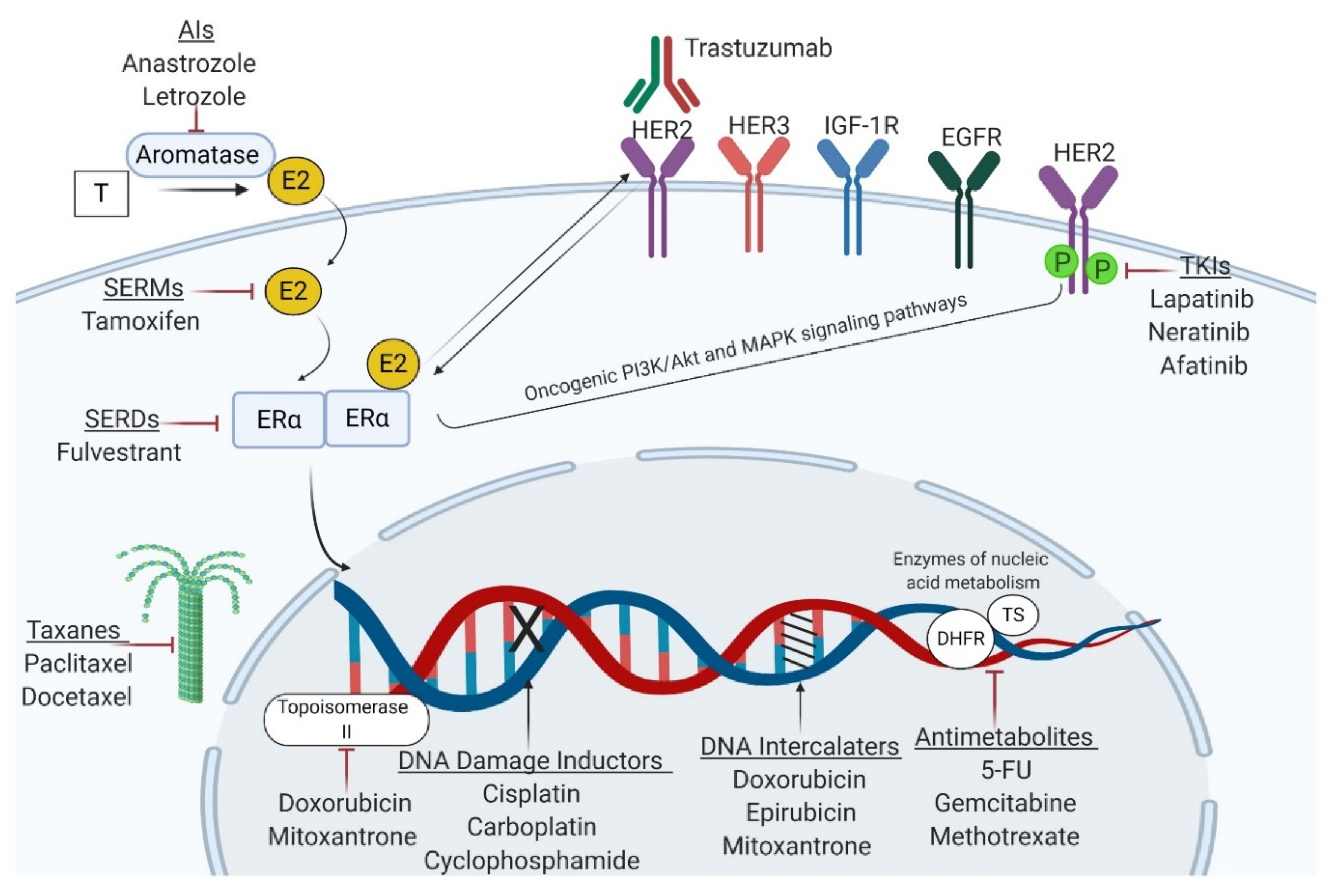
2. Current Therapeutic Strategies and Associated Drug Resistance Profiles in BC
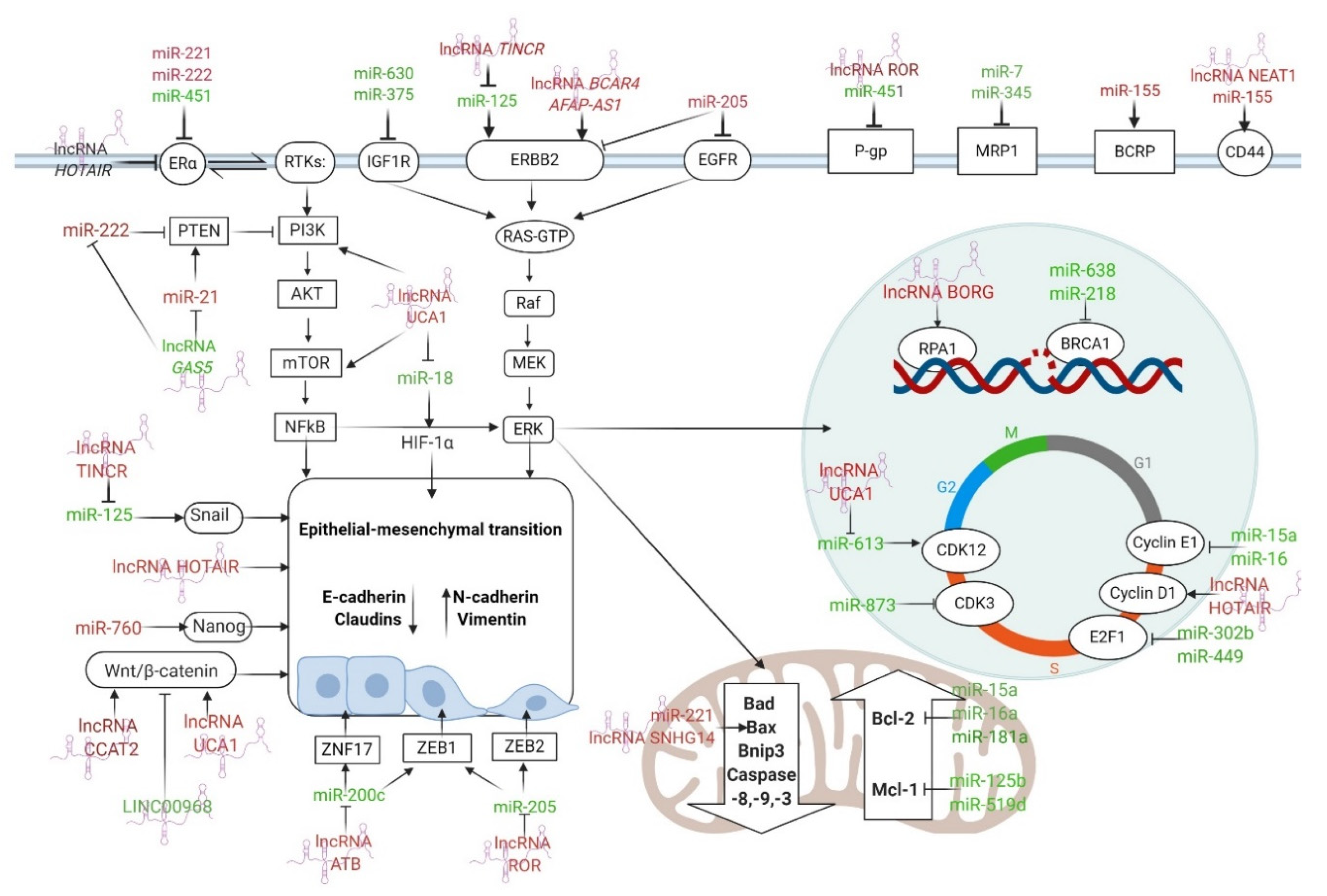
3. miRNAs and BC Drug Resistance
3.1. miRNAs Involved in BC Resistance to Endocrine Therapies
3.2. miRNAs Involved in BC Resistance to Targeted Therapies
3.3. miRNAs Involved in BC Chemoresistance
4. lncRNAs in BC Drug Resistance
4.1. lncRNAs and BC Resistance to Endocrine Therapy
4.2. lncRNAs and Resistance to Anti-HER2 Drugs
4.3. lncRNAs and BC Chemoresistance
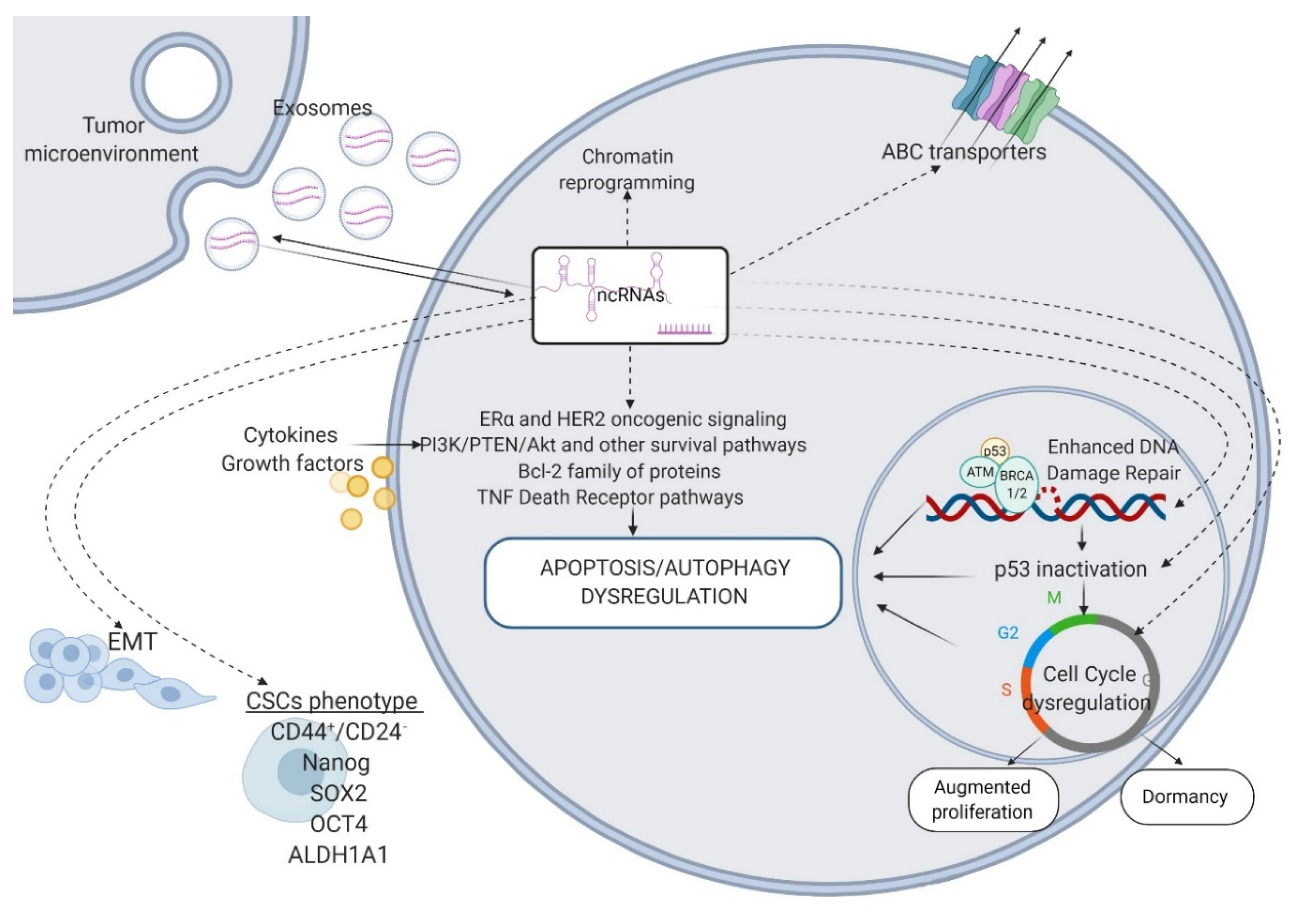
5. ncRNAs and their Predictive Value in BC
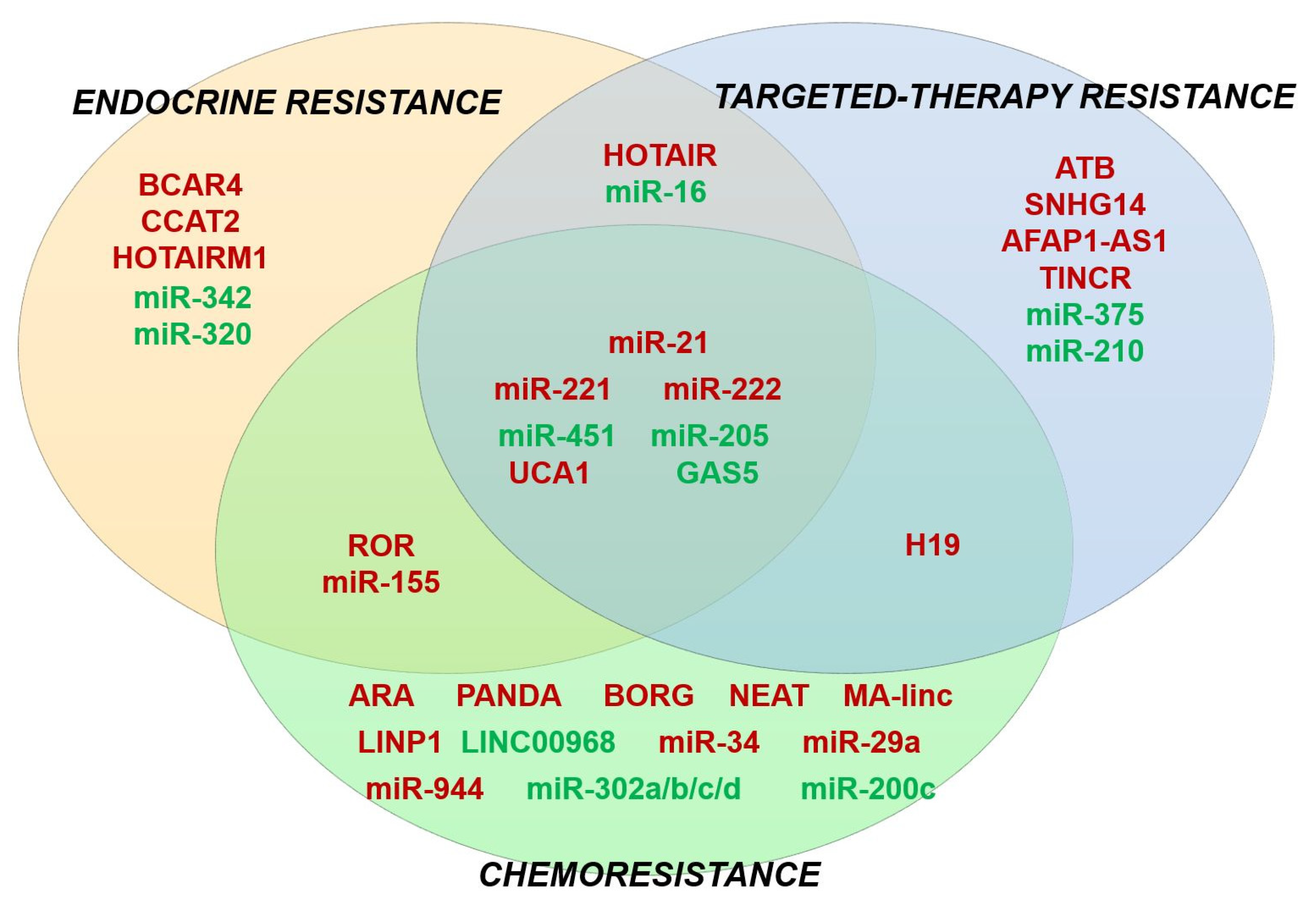
6. Discussion
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Klinge, C.M. Non-Coding RNAs in Breast Cancer: Intracellular and Intercellular Communication. Non-Coding RNA 2018, 4, 40. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed]
- Caplan, L. Delay in breast cancer: Implications for stage at diagnosis and survival. Front. Public Health 2014, 2, 87. [Google Scholar] [CrossRef]
- Guo, H.; Treude, F.; Krämer, O.H.; Lüscher, B.; Hartkamp, J. PAR-4 overcomes chemo-resistance in breast cancer cells by antagonizing cIAP1. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef]
- Prat, A.; Perou, C.M. Deconstructing the molecular portraits of breast cancer. Mol. Oncol. 2011, 5, 5–23. [Google Scholar] [CrossRef] [PubMed]
- Holliday, D.L.; Speirs, V. Choosing the right cell line for breast cancer research. Breast Cancer Res. 2011, 13, 215. [Google Scholar] [CrossRef]
- Malhotra, A.; Jain, M.; Prakash, H.; Vasquez, K.M.; Jain, A. The regulatory roles of long non-coding RNAs in the development of chemoresistance in breast cancer. Oncotarget 2017, 8, 110671–110684. [Google Scholar] [CrossRef]
- Wang, R.; Zhu, Y.; Liu, X.; Liao, X.; He, J.; Niu, L. The Clinicopathological features and survival outcomes of patients with different metastatic sites in stage IV breast cancer. BMC Cancer 2019, 19, 1091. [Google Scholar] [CrossRef]
- Survival Rates for Breast Cancer. Available online: https://www.cancer.org/cancer/breast-cancer/understanding-a-breast-cancer-diagnosis/breast-cancer-survival-rates.html (accessed on 27 July 2020).
- Gonzalez-Angulo, A.M.; Morales-Vasquez, F.; Hortobagyi, G.N. Overview of resistance to systemic therapy in patients with breast cancer. Adv. Exp. Med. Biol. 2007, 608, 1–22. [Google Scholar] [CrossRef]
- Masoud, V.; Pagès, G. Targeted therapies in breast cancer: New challenges to fight against resistance. World J. Clin. Oncol. 2017, 8, 120–134. [Google Scholar] [CrossRef]
- Szostakowska, M.; Trębińska-Stryjewska, A.; Grzybowska, E.A.; Fabisiewicz, A. Resistance to endocrine therapy in breast cancer: Molecular mechanisms and future goals. Breast Cancer Res. Treat. 2019, 173, 489–497. [Google Scholar] [CrossRef] [PubMed]
- Kyrochristos, I.D.; Ziogas, D.E.; Roukos, D.H. Dynamic genome and transcriptional network-based biomarkers and drugs: Precision in breast cancer therapy. Med. Res. Rev. 2019, 39, 1205–1227. [Google Scholar] [CrossRef] [PubMed]
- Lander, E.S.; Linton, L.M.; Birren, B.; Nusbaum, C.; Zody, M.C.; Baldwin, J.; Devon, K.; Dewar, K.; Doyle, M.; FitzHugh, W.; et al. Initial sequencing and analysis of the human genome. Nature 2001, 409, 860–921. [Google Scholar] [CrossRef]
- Gomes, A.Q.; Nolasco, S.; Soares, H. Non-coding RNAs: Multi-tasking molecules in the cell. Int. J. Mol. Sci. 2013, 14, 16010–16039. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Zhang, L.; Zhang, C. Long noncoding RNAs and tumorigenesis: Genetic associations, molecular mechanisms, and therapeutic strategies. Tumor Biol. 2016, 37, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Tran, A.M.; Chalbatani, G.M.; Berland, L.; Cruz De los Santos, M.; Raj, P.; Jalali, S.A.; Gharagouzloo, E.; Ivan, C.; Dragomir, M.P.; Calin, G.A. A New World of Biomarkers and Therapeutics for Female Reproductive System and Breast Cancers: Circular RNAs. Front. Cell Dev. Biol. 2020, 8, 50. [Google Scholar] [CrossRef]
- Hansen, T.B.; Jensen, T.I.; Clausen, B.H.; Bramsen, J.B.; Finsen, B.; Damgaard, C.K.; Kjems, J. Natural RNA circles function as efficient microRNA sponges. Nature 2013, 495, 384–388. [Google Scholar] [CrossRef]
- Memczak, S.; Jens, M.; Elefsinioti, A.; Torti, F.; Krueger, J.; Rybak, A.; Maier, L.; Mackowiak, S.D.; Gregersen, L.H.; Munschauer, M.; et al. Circular RNAs are a large class of animal RNAs with regulatory potency. Nature 2013, 495, 333–338. [Google Scholar] [CrossRef]
- Gupta, R.A.; Shah, N.; Wang, K.C.; Kim, J.; Horlings, H.M.; Wong, D.J.; Tsai, M.-C.; Hung, T.; Argani, P.; Rinn, J.L.; et al. Long non-coding RNA HOTAIR reprograms chromatin state to promote cancer metastasis. Nature 2010, 464, 1071–1076. [Google Scholar] [CrossRef]
- Han, M.; Wang, Y.; Liu, M.; Bi, X.; Bao, J.; Zeng, N.; Zhu, Z.; Mo, Z.; Wu, C.; Chen, X. MiR-21 regulates epithelial-mesenchymal transition phenotype and hypoxia-inducible factor-1α expression in third-sphere forming breast cancer stem cell-like cells. Cancer Sci. 2012, 103, 1058–1064. [Google Scholar] [CrossRef]
- Shin, V.Y.; Chen, J.; Cheuk, I.W.Y.; Siu, M.T.; Ho, C.W.; Wang, X.; Jin, H.; Kwong, A. Long non-coding RNA NEAT1 confers oncogenic role in triple-negative breast cancer through modulating chemoresistance and cancer stemness. Cell Death Dis. 2019, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Han, M.; Gu, Y.; Lu, P.; Li, J.; Cao, H.; Li, X.; Qian, X.; Yu, C.; Yang, Y.; Yang, X.; et al. Exosome-mediated lncRNA AFAP1-AS1 promotes trastuzumab resistance through binding with AUF1 and activating ERBB2 translation. Mol. Cancer 2020, 19, 26. [Google Scholar] [CrossRef] [PubMed]
- De Cola, A.; Volpe, S.; Budani, M.C.; Ferracin, M.; Lattanzio, R.; Turdo, A.; D’Agostino, D.; Capone, E.; Stassi, G.; Todaro, M.; et al. miR-205-5p-mediated downregulation of ErbB/HER receptors in breast cancer stem cells results in targeted therapy resistance. Cell Death Dis. 2015, 6, e1823. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Wang, Y.; Jiang, L.; He, M.; Bai, X.; Yu, L.; Wei, M. MIR-302a/b/c/d cooperatively sensitizes breast cancer cells to adriamycin via suppressing P-glycoprotein(P-gp) by targeting MAP/ERK kinase kinase 1 (MEKK1). J. Exp. Clin. Cancer Res. 2016, 35, 25. [Google Scholar] [CrossRef] [PubMed]
- Cataldo, A.; Piovan, C.; Plantamura, I.; D’Ippolito, E.; Camelliti, S.; Casalini, P.; Giussani, M.; Déas, O.; Cairo, S.; Judde, J.G.; et al. MiR-205 as predictive biomarker and adjuvant therapeutic tool in combination with trastuzumab. Oncotarget 2018, 9, 27920–27928. [Google Scholar] [CrossRef]
- Hu, Y.; Qiu, Y.; Yagüe, E.; Ji, W.; Liu, J.; Zhang, J. MiRNA-205 targets VEGFA and FGF2 and regulates resistance to chemotherapeutics in breast cancer. Cell Death Dis. 2016, 7, e2291. [Google Scholar] [CrossRef]
- Soni, M.; Patel, Y.; Markoutsa, E.; Jie, C.; Liu, S.; Xu, P.; Chen, H. Autophagy, Cell Viability, and Chemoresistance Are Regulated By miR-489 in Breast Cancer. Mol. Cancer Res. 2018, 16, 1348–1360. [Google Scholar] [CrossRef]
- Cheng, S.; Huang, Y.; Lou, C.; He, Y.; Zhang, Y.; Zhang, Q. FSTL1 enhances chemoresistance and maintains stemness in breast cancer cells via integrin β3/Wnt signaling under miR-137 regulation. Cancer Biol. Ther. 2019, 20, 328–337. [Google Scholar] [CrossRef]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef]
- Steinbichler, T.B.; Dudás, J.; Skvortsov, S.; Ganswindt, U.; Riechelmann, H.; Skvortsova, I.I. Therapy resistance mediated by exosomes. Mol. Cancer 2019, 18, 1–11. [Google Scholar] [CrossRef]
- Guzel, E.; Okyay, T.M.; Yalcinkaya, B.; Karacaoglu, S.; Gocmen, M.; Akcakuyu, M.H. Tumor suppressor and oncogenic role of long non-coding RNAs in cancer. North. Clin. Istanb. 2019, 7, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Santos, J.C.; Ribeiro, M.L.; Sarian, L.O.; Ortega, M.M.; Derchain, S.F. Exosomes-mediate microRNAs transfer in breast cancer chemoresistance regulation. Am. J. Cancer Res. 2016, 6, 2129–2139. [Google Scholar] [PubMed]
- Ariazi, E.A.; Ariazi, J.L.; Cordera, F.; Jordan, V.C. Estrogen receptors as therapeutic targets in breast cancer. Curr. Top. Med. Chem. 2006, 6, 181–202. [Google Scholar] [CrossRef]
- Campos-Parra, A.D.; Mitznahuatl, G.C.; Pedroza-Torres, A.; Romo, R.V.; Reyes, F.I.P.; López-Urrutia, E.; Pérez-Plasencia, C. Micro-RNAs as Potential Predictors of Response to Breast Cancer Systemic Therapy: Future Clinical Implications. Int. J. Mol. Sci. 2017, 18, 1182. [Google Scholar] [CrossRef]
- Ali, S.; Rasool, M.; Chaoudhry, H.; Pushparaj, P.N.; Jha, P.; Hafiz, A.; Mahfooz, M.; Abdus Sami, G.; Azhar Kamal, M.; Bashir, S.; et al. Molecular mechanisms and mode of tamoxifen resistance in breast cancer. Bioinformation 2016, 12, 135–139. [Google Scholar] [CrossRef] [PubMed]
- Van Hellemond, I.E.G.; Geurts, S.M.E.; Tjan-Heijnen, V.C.G. Current Status of Extended Adjuvant Endocrine Therapy in Early Stage Breast Cancer. Curr. Treat. Options Oncol. 2018, 19, 26. [Google Scholar] [CrossRef] [PubMed]
- Schwartzberg, L.S.; Wang, G.; Somer, B.G.; Blakely, L.J.; Wheeler, B.M.; Walker, M.S.; Stepanski, E.J.; Houts, A.C. Phase II trial of fulvestrant with metronomic capecitabine for postmenopausal women with hormone receptor-positive, HER2-negative metastatic breast cancer. Clin. Breast Cancer 2014, 14, 13–19. [Google Scholar] [CrossRef]
- Pernas, S.; Tolaney, S.M.; Winer, E.P.; Goel, S. CDK4/6 inhibition in breast cancer: Current practice and future directions. Ther. Adv. Med. Oncol. 2018, 10, 1758835918786451. [Google Scholar] [CrossRef]
- Tripathy, D.; Im, S.-A.; Colleoni, M.; Franke, F.; Bardia, A.; Harbeck, N.; Hurvitz, S.A.; Chow, L.; Sohn, J.; Lee, K.S.; et al. Ribociclib plus endocrine therapy for premenopausal women with hormone-receptor-positive, advanced breast cancer (MONALEESA-7): A randomised phase 3 trial. Lancet Oncol. 2018, 19, 904–915. [Google Scholar] [CrossRef]
- Rugo, H.S.; Finn, R.S.; Diéras, V.; Ettl, J.; Lipatov, O.; Joy, A.A.; Harbeck, N.; Castrellon, A.; Iyer, S.; Lu, D.R.; et al. Palbociclib plus letrozole as first-line therapy in estrogen receptor-positive/human epidermal growth factor receptor 2-negative advanced breast cancer with extended follow-up. Breast Cancer Res. Treat. 2019, 174, 719–729. [Google Scholar] [CrossRef]
- Sledge, G.W.J.; Toi, M.; Neven, P.; Sohn, J.; Inoue, K.; Pivot, X.; Burdaeva, O.; Okera, M.; Masuda, N.; Kaufman, P.A.; et al. The Effect of Abemaciclib Plus Fulvestrant on Overall Survival in Hormone Receptor-Positive, ERBB2-Negative Breast Cancer That Progressed on Endocrine Therapy-MONARCH 2: A Randomized Clinical Trial. JAMA Oncol. 2019, 6, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Pandey, K.; An, H.-J.; Kim, S.K.; Lee, S.A.; Kim, S.; Lim, S.M.; Kim, G.M.; Sohn, J.; Moon, Y.W. Molecular mechanisms of resistance to CDK4/6 inhibitors in breast cancer: A review. Int. J. Cancer 2019, 145, 1179–1188. [Google Scholar] [CrossRef] [PubMed]
- Eggemann, H.; Ignatov, T.; Burger, E.; Kantelhardt, E.J.; Fettke, F.; Thomssen, C.; Costa, S.D.; Ignatov, A. Moderate HER2 expression as a prognostic factor in hormone receptor positive breast cancer. Endocr. Relat. Cancer 2015, 22, 725–733. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Tang, P. Genomic pathology and biomarkers in breast cancer. Crit. Rev. Oncog. 2017, 22, 411–426. [Google Scholar] [CrossRef] [PubMed]
- Pernas, S.; Tolaney, S.M. HER2-positive breast cancer: New therapeutic frontiers and overcoming resistance. Ther. Adv. Med. Oncol. 2019, 11, 1758835919833519. [Google Scholar] [CrossRef] [PubMed]
- Campos-Parra, A.; López-Urrutia, E.; Orozco Moreno, L.; López-Camarillo, C.; Meza-Menchaca, T.; Figueroa González, G.; Bustamante Montes, L.; Pérez-Plasencia, C. Long Non-Coding RNAs as New Master Regulators of Resistance to Systemic Treatments in Breast Cancer. Int. J. Mol. Sci. 2018, 19, 2711. [Google Scholar] [CrossRef] [PubMed]
- Martin, H.L.; Smith, L.; Tomlinson, D.C. Multidrug-resistant breast cancer: Current perspectives. Breast Cancer 2014, 6, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Teven, C.M.; Schmid, D.B.; Sisco, M.; Ward, J.; Howard, M.A. Systemic Therapy for Early-Stage Breast Cancer: What the Plastic Surgeon Should Know. Eplasty 2017, 17, e7. [Google Scholar] [PubMed]
- Haque, M.M.; Desai, K.V. Pathways to Endocrine Therapy Resistance in Breast Cancer. Front. Endocrinol. 2019, 10, 573. [Google Scholar] [CrossRef] [PubMed]
- Godoy-Ortiz, A.; Sanchez-Muñoz, A.; Parrado, M.R.C.; Álvarez, M.; Ribelles, N.; Dominguez, A.R.; Alba, E. Deciphering her2 breast cancer disease: Biological and clinical implications. Front. Oncol. 2019, 9, 1124. [Google Scholar] [CrossRef] [PubMed]
- Nedeljković, M.; Damjanović, A. Mechanisms of Chemotherapy Resistance in Triple-Negative Breast Cancer-How We Can Rise to the Challenge. Cells 2019, 8, 957. [Google Scholar] [CrossRef] [PubMed]
- Pogribny, I.P.; Filkowski, J.N.; Tryndyak, V.P.; Golubov, A.; Shpyleva, S.I.; Kovalchuk, O. Alterations of microRNAs and their targets are associated with acquired resistance of MCF-7 breast cancer cells to cisplatin. Int. J. Cancer 2010, 127, 1785–1794. [Google Scholar] [CrossRef] [PubMed]
- Kastl, L.; Brown, I.; Schofield, A.C. miRNA-34a is associated with docetaxel resistance in human breast cancer cells. Breast Cancer Res. Treat. 2012, 131, 445–454. [Google Scholar] [CrossRef]
- Wei, Y.; Lai, X.; Yu, S.; Chen, S.; Ma, Y.; Zhang, Y.; Li, H.; Zhu, X.; Yao, L.; Zhang, J. Exosomal miR-221/222 enhances tamoxifen resistance in recipient ER-positive breast cancer cells. Breast Cancer Res. Treat. 2014, 147, 423–431. [Google Scholar] [CrossRef]
- Shi, S.-J.; Wang, L.-J.; Yu, B.; Li, Y.-H.; Jin, Y.; Bai, X.-Z. LncRNA-ATB promotes trastuzumab resistance and invasion-metastasis cascade in breast cancer. Oncotarget 2015, 6, 11652–11663. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Xu, K.; Yagüe, E. miR-218 targets survivin and regulates resistance to chemotherapeutics in breast cancer. Breast Cancer Res. Treat. 2015, 151, 269–280. [Google Scholar] [CrossRef]
- Zhu, Q.N.; Wang, G.; Guo, Y.; Peng, Y.; Zhang, R.; Deng, J.L.; Li, Z.X.; Zhu, Y.S. LncRNA H19 is a major mediator of doxorubicin chemoresistance in breast cancer cells through a cullin4A-MDR1 pathway. Oncotarget 2017, 8, 91990–92003. [Google Scholar] [CrossRef]
- Chen, D.; Si, W.; Shen, J.; Du, C.; Lou, W.; Bao, C.; Zheng, H.; Pan, J.; Zhong, G.; Xu, L.; et al. MiR-27b-3p inhibits proliferation and potentially reverses multi-chemoresistance by targeting CBLB/GRB2 in breast cancer cells. Cell Death Dis. 2018, 9, 1–13. [Google Scholar] [CrossRef]
- Sevignani, C.; Calin, G.A.; Siracusa, L.D.; Croce, C.M. Mammalian microRNAs: A small world for fine-tuning gene expression. Mamm. Genome 2006, 17, 189–202. [Google Scholar] [CrossRef]
- Aisina, D.; Niyazova, R.; Atambayeva, S.; Ivashchenko, A. Prediction of clusters of miRNA binding sites in mRNA candidate genes of breast cancer subtypes. PeerJ 2019, 7, e8049. [Google Scholar] [CrossRef]
- Kazimierczyk, M.; Kasprowicz, M.K.; Kasprzyk, M.E.; Wrzesinski, J. Human Long Noncoding RNA Interactome: Detection, Characterization and Function. Int. J. Mol. Sci. 2020, 21, 1027. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Wang, D. The Pattern of microRNA Binding Site Distribution. Genes 2017, 8, 296. [Google Scholar] [CrossRef] [PubMed]
- Stavast, C.J.; Erkeland, S.J. The Non-Canonical Aspects of MicroRNAs: Many Roads to Gene Regulation. Cells 2019, 8, 1465. [Google Scholar] [CrossRef] [PubMed]
- Treiber, T.; Treiber, N.; Meister, G. Regulation of microRNA biogenesis and its crosstalk with other cellular pathways. Nat. Rev. Mol. Cell Biol. 2019, 20, 5–20. [Google Scholar] [CrossRef]
- Martinez, I.; Hayes, K.E.; Barr, J.A.; Harold, A.D.; Xie, M.; Bukhari, S.I.A.; Vasudevan, S.; Steitz, J.A.; DiMaio, D. An Exportin-1-dependent microRNA biogenesis pathway during human cell quiescence. Proc. Natl. Acad. Sci. USA 2017, 114, E4961–E4970. [Google Scholar] [CrossRef]
- Patterson, D.G.; Roberts, J.T.; King, V.M.; Houserova, D.; Barnhill, E.C.; Crucello, A.; Polska, C.J.; Brantley, L.W.; Kaufman, G.C.; Nguyen, M.; et al. Human snoRNA-93 is processed into a microRNA-like RNA that promotes breast cancer cell invasion. NPJ Breast Cancer 2017, 3, 25. [Google Scholar] [CrossRef]
- Maute, R.L.; Schneider, C.; Sumazin, P.; Holmes, A.; Califano, A.; Basso, K.; Dalla-Favera, R. tRNA-derived microRNA modulates proliferation and the DNA damage response and is down-regulated in B cell lymphoma. Proc. Natl. Acad. Sci. USA 2013, 110, 1404–1409. [Google Scholar] [CrossRef]
- Guo, H.; Ingolia, N.T.; Weissman, J.S.; Bartel, D.P. Mammalian microRNAs predominantly act to decrease target mRNA levels. Nature 2010, 466, 835–840. [Google Scholar] [CrossRef]
- Gasparini, P.; Cascione, L.; Fassan, M.; Lovat, F.; Guler, G.; Balci, S.; Irkkan, C.; Morrison, C.; Croce, C.M.; Shapiro, C.L.; et al. microRNA expression profiling identifies a four microRNA signature as a novel diagnostic and prognostic biomarker in triple negative breast cancers. Oncotarget 2014, 5, 1174–1184. [Google Scholar] [CrossRef]
- Liang, F.; Yang, M.; Tong, N.; Fang, J.; Pan, Y.; Li, J.; Zhang, X. Identification of six key miRNAs associated with breast cancer through screening large-scale microarray data. Oncol. Lett. 2018, 16, 4159–4168. [Google Scholar] [CrossRef]
- Cai, J.; Guan, H.; Fang, L.; Yang, Y.; Zhu, X.; Yuan, J.; Wu, J.; Li, M. MicroRNA-374a activates Wnt/β-catenin signaling to promote breast cancer metastasis. J. Clin. Investig. 2013, 123, 566–579. [Google Scholar] [CrossRef]
- Shen, F.; Cai, W.-S.; Feng, Z.; Li, J.-L.; Chen, J.-W.; Cao, J.; Xu, B. MiR-492 contributes to cell proliferation and cell cycle of human breast cancer cells by suppressing SOX7 expression. Tumour Biol. J. Int. Soc. Oncodev. Biol. Med. 2015, 36, 1913–1921. [Google Scholar] [CrossRef] [PubMed]
- Santos, J.C.; Lima, N.D.S.; Sarian, L.O.; Matheu, A.; Ribeiro, M.L.; Derchain, S.F.M. Exosome-mediated breast cancer chemoresistance via miR-155 transfer. Sci. Rep. 2018, 8, 829. [Google Scholar] [CrossRef] [PubMed]
- Calin, G.A.; Sevignani, C.; Dumitru, C.D.; Hyslop, T.; Noch, E.; Yendamuri, S.; Shimizu, M.; Rattan, S.; Bullrich, F.; Negrini, M.; et al. Human microRNA genes are frequently located at fragile sites and genomic regions involved in cancers. Proc. Natl. Acad. Sci. USA 2004, 101, 2999–3004. [Google Scholar] [CrossRef] [PubMed]
- Ye, P.; Fang, C.; Zeng, H.; Shi, Y.; Pan, Z.; An, N.; He, K.; Zhang, L.; Long, X. Differential microRNA expression profiles in tamoxifen-resistant human breast cancer cell lines induced by two methods. Oncol. Lett. 2018, 15, 3532–3539. [Google Scholar] [CrossRef]
- Zhou, Q.; Zeng, H.; Ye, P.; Shi, Y.; Guo, J.; Long, X. Differential microRNA profiles between fulvestrant-resistant and tamoxifen-resistant human breast cancer cells. Anticancer Drugs 2018, 29, 539–548. [Google Scholar] [CrossRef]
- Cittelly, D.M.; Das, P.M.; Spoelstra, N.S.; Edgerton, S.M.; Richer, J.K.; Thor, A.D.; Jones, F.E. Downregulation of miR-342 is associated with tamoxifen resistant breast tumors. Mol. Cancer 2010, 9, 317. [Google Scholar] [CrossRef]
- Lü, M.; Ding, K.; Zhang, G.; Yin, M.; Yao, G.; Tian, H.; Lian, J.; Liu, L.; Liang, M.; Zhu, T.; et al. MicroRNA-320a sensitizes tamoxifen-resistant breast cancer cells to tamoxifen by targeting ARPP-19 and ERRγ∗. Sci. Rep. 2015, 5, 1–10. [Google Scholar] [CrossRef]
- Bergamaschi, A.; Katzenellenbogen, B.S. Tamoxifen downregulation of miR-451 increases 14-3-3ζ and promotes breast cancer cell survival and endocrine resistance. Oncogene 2012, 31, 39–47. [Google Scholar] [CrossRef]
- Chu, J.; Zhu, Y.; Liu, Y.; Sun, L.; Lv, X.; Wu, Y.; Hu, P.; Su, F.; Gong, C.; Song, E.; et al. E2F7 overexpression leads to tamoxifen resistance in breast cancer cells by competing with E2F1 at miR-15a/16 promoter. Oncotarget 2015, 6, 31944–31957. [Google Scholar] [CrossRef]
- Yu, X.; Li, R.; Shi, W.; Jiang, T.; Wang, Y.; Li, C.; Qu, X. Silencing of MicroRNA-21 confers the sensitivity to tamoxifen and fulvestrant by enhancing autophagic cell death through inhibition of the PI3K-AKT-mTOR pathway in breast cancer cells. Biomed. Pharmacother. 2016, 77, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.-H.; Tao, Z.-H.; Zhang, J.; Li, T.; Ni, C.; Xie, J.; Zhang, J.-F.; Hu, X.-C. MiRNA-21 induces epithelial to mesenchymal transition and gemcitabine resistance via the PTEN/AKT pathway in breast cancer. Tumour Biol. J. Int. Soc. Oncodev. Biol. Med. 2016, 37, 7245–7254. [Google Scholar] [CrossRef]
- Gong, C.; Yao, Y.; Wang, Y.; Liu, B.; Wu, W.; Chen, J.; Su, F.; Yao, H.; Song, E. Up-regulation of miR-21 mediates resistance to trastuzumab therapy for breast cancer. J. Biol. Chem. 2011, 286, 19127–19137. [Google Scholar] [CrossRef] [PubMed]
- Rao, X.; Di Leva, G.; Li, M.; Fang, F.; Devlin, C.; Hartman-Frey, C.; Burow, M.E.; Ivan, M.; Croce, C.M.; Nephew, K.P. MicroRNA-221/222 confers breast cancer fulvestrant resistance by regulating multiple signaling pathways. Oncogene 2011, 30, 1082–1097. [Google Scholar] [CrossRef]
- Sansone, P.; Berishaj, M.; Rajasekhar, V.K.; Ceccarelli, C.; Chang, Q.; Strillacci, A.; Savini, C.; Shapiro, L.; Bowman, R.L.; Mastroleo, C.; et al. Evolution of Cancer Stem-like Cells in Endocrine-Resistant Metastatic Breast Cancers Is Mediated by Stromal Microvesicles. Cancer Res. 2017, 77, 1927–1941. [Google Scholar] [CrossRef]
- Shen, R.; Wang, Y.; Wang, C.-X.; Yin, M.; Liu, H.-L.; Chen, J.-P.; Han, J.-Q.; Wang, W.-B. MiRNA-155 mediates TAM resistance by modulating SOCS6-STAT3 signalling pathway in breast cancer. Am. J. Transl. Res. 2015, 7, 2115–2126. [Google Scholar] [PubMed]
- Miller, T.E.; Ghoshal, K.; Ramaswamy, B.; Roy, S.; Datta, J.; Shapiro, C.L.; Jacob, S.; Majumder, S. MicroRNA-221/222 confers tamoxifen resistance in breast cancer by targeting p27Kip1. J. Biol. Chem. 2008, 283, 29897–29903. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Zhang, W.; Liu, C.; Li, G. miR-200 affects tamoxifen resistance in breast cancer cells through regulation of MYB. Sci. Rep. 2019, 9, 1–6. [Google Scholar] [CrossRef]
- Cui, J.; Yang, Y.; Li, H.; Leng, Y.; Qian, K.; Huang, Q.; Zhang, C.; Lu, Z.; Chen, J.; Sun, T.; et al. MiR-873 regulates ERα transcriptional activity and tamoxifen resistance via targeting CDK3 in breast cancer cells. Oncogene 2015, 34, 3895–3907. [Google Scholar] [CrossRef]
- Ahmad, A.; Ginnebaugh, K.R.; Yin, S.; Bollig-Fischer, A.; Reddy, K.B.; Sarkar, F.H. Functional role of miR-10b in tamoxifen resistance of ER-positive breast cancer cells through down-regulation of HDAC4. BMC Cancer 2015, 15, 540. [Google Scholar] [CrossRef]
- Chen, M.-J.; Cheng, Y.-M.; Chen, C.-C.; Chen, Y.-C.; Shen, C.-J. MiR-148a and miR-152 reduce tamoxifen resistance in ER+ breast cancer via downregulating ALCAM. Biochem. Biophys. Res. Commun. 2017, 483, 840–846. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Zou, Z.; Nie, P.; Kou, X.; Wu, B.; Wang, S.; Song, Z.; He, J. Downregulation of microRNA-27b-3p enhances tamoxifen resistance in breast cancer by increasing NR5A2 and CREB1 expression. Cell Death Dis. 2016, 7, e2454. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wu, Y.; Liu, A.; Tang, X. MiR-27b is epigenetically downregulated in tamoxifen resistant breast cancer cells due to promoter methylation and regulates tamoxifen sensitivity by targeting HMGB3. Biochem. Biophys. Res. Commun. 2016, 477, 768–773. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Luo, A.; Liu, Y.; Wang, S.; Li, Y.; Shi, W.; Liu, Z.; Qu, X. MiR-214 increases the sensitivity of breast cancer cells to tamoxifen and fulvestrant through inhibition of autophagy. Mol. Cancer 2015, 14, 208. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Fu, Z.; Yang, M.; Sun, C.; Li, Y.; Chu, J.; Zhang, Y.; Li, W.; Huang, X.; Li, J.; et al. Expression pattern of microRNAs related with response to trastuzumab in breast cancer. J. Cell. Physiol. 2019, 234, 16102–16113. [Google Scholar] [CrossRef]
- Li, H.; Liu, J.; Chen, J.; Wang, H.; Yang, L.; Chen, F.; Fan, S.; Wang, J.; Shao, B.; Yin, D.; et al. A serum microRNA signature predicts trastuzumab benefit in HER2-positive metastatic breast cancer patients. Nat. Commun. 2018, 9. [Google Scholar] [CrossRef]
- Di Cosimo, S.; Appierto, V.; Pizzamiglio, S.; Silvestri, M.; Baselga, J.; Piccart, M.; Huober, J.; Izquierdo, M.; de la Pena, L.; Hilbers, F.S.; et al. Early Modulation of Circulating MicroRNAs Levels in HER2-Positive Breast Cancer Patients Treated with Trastuzumab-Based Neoadjuvant Therapy. Int. J. Mol. Sci. 2020, 21, 1386. [Google Scholar] [CrossRef]
- Ye, X.-M.; Zhu, H.-Y.; Bai, W.-D.; Wang, T.; Wang, L.; Chen, Y.; Yang, A.-G.; Jia, L.-T. Epigenetic silencing of miR-375 induces trastuzumab resistance in HER2-positive breast cancer by targeting IGF1R. BMC Cancer 2014, 14, 134. [Google Scholar] [CrossRef]
- Corcoran, C.; Rani, S.; Breslin, S.; Gogarty, M.; Ghobrial, I.M.; Crown, J.; O’Driscoll, L. MiR-630 targets IGF1R to regulate response to HER-targeting drugs and overall cancer cell progression in HER2 over-expressing breast cancer. Mol. Cancer 2014, 13, 71. [Google Scholar] [CrossRef]
- Crosby, M.E.; Kulshreshtha, R.; Ivan, M.; Glazer, P.M. MicroRNA regulation of DNA repair gene expression in hypoxic stress. Cancer Res. 2009, 69, 1221–1229. [Google Scholar] [CrossRef]
- Jung, E.-J.; Santarpia, L.; Kim, J.; Esteva, F.J.; Moretti, E.; Buzdar, A.U.; Di Leo, A.; Le, X.-F.; Bast, R.C.J.; Park, S.-T.; et al. Plasma microRNA 210 levels correlate with sensitivity to trastuzumab and tumor presence in breast cancer patients. Cancer 2012, 118, 2603–2614. [Google Scholar] [CrossRef] [PubMed]
- De Mattos-Arruda, L.; Bottai, G.; Nuciforo, P.G.; Di Tommaso, L.; Giovannetti, E.; Peg, V.; Losurdo, A.; Pérez-Garcia, J.; Masci, G.; Corsi, F.; et al. MicroRNA-21 links epithelial-to-mesenchymal transition and inflammatory signals to confer resistance to neoadjuvant trastuzumab and chemotherapy in HER2-positive breast cancer patients. Oncotarget 2015, 6, 37269–37280. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.; Bai, W.; Zhu, H.; Zhang, X.; Chen, Y.; Wang, L.; Yang, A.; Zhao, J.; Jia, L. MiR-221 promotes trastuzumab-resistance and metastasis in HER2-positive breast cancers by targeting PTEN. BMB Rep. 2014, 47, 268–273. [Google Scholar] [CrossRef] [PubMed]
- Venturutti, L.; Cordo Russo, R.I.; Rivas, M.A.; Mercogliano, M.F.; Izzo, F.; Oakley, R.H.; Pereyra, M.G.; De Martino, M.; Proietti, C.J.; Yankilevich, P.; et al. MiR-16 mediates trastuzumab and lapatinib response in ErbB-2-positive breast and gastric cancer via its novel targets CCNJ and FUBP1. Oncogene 2016, 35, 6189–6202. [Google Scholar] [CrossRef]
- Han, M.; Hu, J.; Lu, P.; Cao, H.; Yu, C.; Li, X.; Qian, X.; Yang, X.; Yang, Y.; Han, N.; et al. Exosome-transmitted miR-567 reverses trastuzumab resistance by inhibiting ATG5 in breast cancer. Cell Death Dis. 2020, 11, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Bai, W.-D.; Ye, X.-M.; Zhang, M.-Y.; Zhu, H.-Y.; Xi, W.-J.; Huang, X.; Zhao, J.; Gu, B.; Zheng, G.-X.; Yang, A.-G.; et al. MiR-200c suppresses TGF-β signaling and counteracts trastuzumab resistance and metastasis by targeting ZNF217 and ZEB1 in breast cancer. Int. J. Cancer 2014, 135, 1356–1368. [Google Scholar] [CrossRef]
- Huynh, F.C.; Jones, F.E. MicroRNA-7 Inhibits Multiple Oncogenic Pathways to Suppress HER2Δ16 Mediated Breast Tumorigenesis and Reverse Trastuzumab Resistance. PLoS ONE 2014, 9, e114419. [Google Scholar] [CrossRef]
- Wind, N.S.; Holen, I. Multidrug resistance in breast cancer: From in vitro models to clinical studies. Int. J. Breast Cancer 2011, 2011, 967419. [Google Scholar] [CrossRef]
- Tiwari, A.K.; Sodani, K.; Dai, C.-L.; Ashby, C.R.J.; Chen, Z.-S. Revisiting the ABCs of multidrug resistance in cancer chemotherapy. Curr. Pharm. Biotechnol. 2011, 12, 570–594. [Google Scholar] [CrossRef]
- Kovalchuk, O.; Filkowski, J.; Meservy, J.; Ilnytskyy, Y.; Tryndyak, V.P.; Chekhun, V.F.; Pogribny, I.P. Involvement of microRNA-451 in resistance of the MCF-7 breast cancer cells to chemotherapeutic drug doxorubicin. Mol. Cancer Ther. 2008, 7, 2152–2159. [Google Scholar] [CrossRef]
- Cataldo, A.; Cheung, D.G.; Balsari, A.; Tagliabue, E.; Coppola, V.; Iorio, M.V.; Palmieri, D.; Croce, C.M. miR-302b enhances breast cancer cell sensitivity to cisplatin by regulating E2F1 and the cellular DNA damage response. Oncotarget 2016, 7, 786–797. [Google Scholar] [CrossRef] [PubMed]
- Tormo, E.; Ballester, S.; Adam-Artigues, A.; Burgués, O.; Alonso, E.; Bermejo, B.; Menéndez, S.; Zazo, S.; Madoz-Gúrpide, J.; Rovira, A.; et al. The miRNA-449 family mediates doxorubicin resistance in triple-negative breast cancer by regulating cell cycle factors. Sci. Rep. 2019, 9, 5316. [Google Scholar] [CrossRef] [PubMed]
- Tan, X.; Peng, J.; Fu, Y.; An, S.; Rezaei, K.; Tabbara, S.; Teal, C.B.; Man, Y.G.; Brem, R.F.; Fu, S.W. miR-638 mediated regulation of BRCA1 affects DNA repair and sensitivity to UV and cisplatin in triple-negative breast cancer. Breast Cancer Res. 2014, 16, 435. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Xiao, X.; Dong, L.; Wan, N.; Zhou, Z.; Deng, H.; Zhang, X. MiR-218 regulates cisplatin chemosensitivity in breast cancer by targeting BRCA1. Tumour Biol. J. Int. Soc. Oncodev. Biol. Med. 2015, 36, 2065–2075. [Google Scholar] [CrossRef]
- Zhong, S.; Li, W.; Chen, Z.; Xu, J.; Zhao, J. MiR-222 and miR-29a contribute to the drug-resistance of breast cancer cells. Gene 2013, 531, 8–14. [Google Scholar] [CrossRef]
- Zhang, B.; Zhao, R.; He, Y.; Fu, X.; Fu, L.; Zhu, Z.; Fu, L.; Dong, J.-T. MicroRNA 100 sensitizes luminal A breast cancer cells to paclitaxel treatment in part by targeting mTOR. Oncotarget 2016, 7, 5702–5714. [Google Scholar] [CrossRef]
- Lyu, H.; Wang, S.; Huang, J.; Wang, B.; He, Z.; Liu, B. Survivin-targeting miR-542-3p overcomes HER3 signaling-induced chemoresistance and enhances the antitumor activity of paclitaxel against HER2-overexpressing breast cancer. Cancer Lett. 2018, 420, 97–108. [Google Scholar] [CrossRef]
- Gu, X.; Li, J.-Y.; Guo, J.; Li, P.-S.; Zhang, W.-H. Influence of MiR-451 on Drug Resistances of Paclitaxel-Resistant Breast Cancer Cell Line. Med. Sci. Monit. 2015, 21, 3291–3297. [Google Scholar] [CrossRef]
- Zhu, Y.; Wu, J.; Li, S.; Ma, R.; Cao, H.; Ji, M.; Jing, C.; Tang, J. The function role of miR-181a in chemosensitivity to adriamycin by targeting Bcl-2 in low-invasive breast cancer cells. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2013, 32, 1225–1237. [Google Scholar] [CrossRef]
- Rodriguez-Barrueco, R.; Nekritz, E.A.; Bertucci, F.; Yu, J.; Sanchez-Garcia, F.; Zeleke, T.Z.; Gorbatenko, A.; Birnbaum, D.; Ezhkova, E.; Cordon-Cardo, C.; et al. miR-424(322)/503 is a breast cancer tumor suppressor whose loss promotes resistance to chemotherapy. Genes Dev. 2017, 31, 553–566. [Google Scholar] [CrossRef]
- Ye, Z.; Hao, R.; Cai, Y.; Wang, X.; Huang, G. Knockdown of miR-221 promotes the cisplatin-inducing apoptosis by targeting the BIM-Bax/Bak axis in breast cancer. Tumor Biol. 2016, 37, 4509–4515. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Tian, W.; Chen, H.; Jiang, K. MiR-944 functions as a novel oncogene and regulates the chemoresistance in breast cancer. Tumour Biol. J. Int. Soc. Oncodev. Biol. Med. 2016, 37, 1599–1607. [Google Scholar] [CrossRef]
- Körner, C.; Keklikoglou, I.; Bender, C.; Wörner, A.; Münstermann, E.; Wiemann, S. MicroRNA-31 sensitizes human breast cells to apoptosis by direct targeting of protein kinase C epsilon (PKCepsilon). J. Biol. Chem. 2013, 288, 8750–8761. [Google Scholar] [CrossRef] [PubMed]
- Tsang, W.P.; Kwok, T.T. Let-7a microRNA suppresses therapeutics-induced cancer cell death by targeting caspase-3. Apoptosis 2008, 13, 1215–1222. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wang, N.; Liu, P.; Chen, Q.; Situ, H.; Xie, T.; Zhang, J.; Peng, C.; Lin, Y.; Chen, J. MicroRNA-25 regulates chemoresistance-associated autophagy in breast cancer cells, a process modulated by the natural autophagy inducer isoliquiritigenin. Oncotarget 2014, 5, 7013–7026. [Google Scholar] [CrossRef]
- Wu, D.; Zhang, J.; Lu, Y.; Bo, S.; Li, L.; Wang, L.; Zhang, Q.; Mao, J. miR-140-5p inhibits the proliferation and enhances the efficacy of doxorubicin to breast cancer stem cells by targeting Wnt1. Cancer Gene Ther. 2019, 26, 74–82. [Google Scholar] [CrossRef]
- Zuo, J.; Yu, Y.; Zhu, M.; Jing, W.; Yu, M.; Chai, H.; Liang, C.; Tu, J. Inhibition of miR-155, a therapeutic target for breast cancer, prevented in cancer stem cell formation. Cancer Biomark. 2018, 21, 383–392. [Google Scholar] [CrossRef]
- Xie, Q.; Wang, S.; Zhao, Y.; Zhang, Z.; Qin, C.; Yang, X. MiR-519d impedes cisplatin-resistance in breast cancer stem cells by down-regulating the expression of MCL-1. Oncotarget 2017, 8, 22003–22013. [Google Scholar] [CrossRef]
- Hu, W.; Tan, C.; He, Y.; Zhang, G.; Xu, Y.; Tang, J. Functional miRNAs in breast cancer drug resistance. OncoTargets Ther. 2018, 11, 1529–1541. [Google Scholar] [CrossRef]
- Bockhorn, J.; Dalton, R.; Nwachukwu, C.; Huang, S.; Prat, A.; Yee, K.; Chang, Y.F.; Huo, D.; Wen, Y.; Swanson, K.E.; et al. MicroRNA-30c inhibits human breast tumour chemotherapy resistance by regulating TWF1 and IL-11. Nat. Commun. 2013, 4, 1–14. [Google Scholar] [CrossRef]
- Ozawa, P.M.M.; Alkhilaiwi, F.; Cavalli, I.J.; Malheiros, D.; de Souza Fonseca Ribeiro, E.M.; Cavalli, L.R. Extracellular vesicles from triple-negative breast cancer cells promote proliferation and drug resistance in non-tumorigenic breast cells. Breast Cancer Res. Treat. 2018, 172, 713–723. [Google Scholar] [CrossRef] [PubMed]
- Shimono, Y.; Zabala, M.; Cho, R.W.; Lobo, N.; Dalerba, P.; Qian, D.; Diehn, M.; Liu, H.; Panula, S.P.; Chiao, E.; et al. Downregulation of miRNA-200c links breast cancer stem cells with normal stem cells. Cell 2009, 138, 592–603. [Google Scholar] [CrossRef] [PubMed]
- Knezevic, J.; Pfefferle, A.D.; Petrovic, I.; Greene, S.B.; Perou, C.M.; Rosen, J.M. Expression of miR-200c in claudin-low breast cancer alters stem cell functionality, enhances chemosensitivity and reduces metastatic potential. Oncogene 2015, 34, 5997–6006. [Google Scholar] [CrossRef]
- Yu, D.-D.; Wu, Y.; Zhang, X.-H.; Lv, M.-M.; Chen, W.-X.; Chen, X.; Yang, S.-J.; Shen, H.; Zhong, S.-L.; Tang, J.-H.; et al. Exosomes from adriamycin-resistant breast cancer cells transmit drug resistance partly by delivering miR-222. Tumour Biol. J. Int. Soc. Oncodev. Biol. Med. 2016, 37, 3227–3235. [Google Scholar] [CrossRef]
- Li, X.J.; Ren, Z.J.; Tang, J.H.; Yu, Q. Exosomal MicroRNA MiR-1246 Promotes Cell Proliferation, Invasion and Drug Resistance by Targeting CCNG2 in Breast Cancer. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2017, 44, 1741–1748. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liang, Y.; Sang, Y.; Song, X.; Zhang, H.; Liu, Y.; Jiang, L.; Yang, Q. MiR-770 suppresses the chemo-resistance and metastasis of triple negative breast cancer via direct targeting of STMN1. Cell Death Dis. 2018, 9, 14. [Google Scholar] [CrossRef]
- Rameshwar, P. Breast cancer cell dormancy in bone marrow: Potential therapeutic targets within the marrow microenvironment. Expert Rev. Anticancer Ther. 2010, 10, 129–132. [Google Scholar] [CrossRef]
- Sueta, A.; Yamamoto, Y.; Iwase, H. The role of exosomal microRNAs; focus on clinical applications in breast cancer. Cancer Drug Resist. 2019. [Google Scholar] [CrossRef]
- Ono, M.; Kosaka, N.; Tominaga, N.; Yoshioka, Y.; Takeshita, F.; Takahashi, R.; Yoshida, M.; Tsuda, H.; Tamura, K.; Ochiya, T. Exosomes from bone marrow mesenchymal stem cells contain a microRNA that promotes dormancy in metastatic breast cancer cells. Sci. Signal. 2014, 7, ra63. [Google Scholar] [CrossRef]
- Bliss, S.A.; Sinha, G.; Sandiford, O.A.; Williams, L.M.; Engelberth, D.J.; Guiro, K.; Isenalumhe, L.L.; Greco, S.J.; Ayer, S.; Bryan, M.; et al. Mesenchymal Stem Cell-Derived Exosomes Stimulate Cycling Quiescence and Early Breast Cancer Dormancy in Bone Marrow. Cancer Res. 2016, 76, 5832–5844. [Google Scholar] [CrossRef]
- Wang, W.; Zhang, L.; Wang, Y.; Ding, Y.; Chen, T.; Wang, Y.; Wang, H.; Li, Y.; Duan, K.; Chen, S.; et al. Involvement of miR-451 in resistance to paclitaxel by regulating YWHAZ in breast cancer. Cell Death Dis. 2017, 8, e3071. [Google Scholar] [CrossRef]
- Bao, L.; Hazari, S.; Mehra, S.; Kaushal, D.; Moroz, K.; Dash, S. Increased expression of P-glycoprotein and doxorubicin chemoresistance of metastatic breast cancer is regulated by miR-298. Am. J. Pathol. 2012, 180, 2490–2503. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.; Wu, H.; Xia, J.; Li, Y.; Zhang, Y.; Huang, K.; Wagar, N.; Yoon, Y.; Cho, H.T.; Scala, S.; et al. Involvement of miR-326 in chemotherapy resistance of breast cancer through modulating expression of multidrug resistance-associated protein 1. Biochem. Pharmacol. 2010, 79, 817–824. [Google Scholar] [CrossRef] [PubMed]
- He, D.-X.; Gu, X.-T.; Jiang, L.; Jin, J.; Ma, X. A methylation-based regulatory network for microRNA 320a in chemoresistant breast cancer. Mol. Pharmacol. 2014, 86, 536–547. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Miao, L.; Liu, M.; Li, C.; Yu, C.; Yan, H.; Yin, Y.; Wang, Y.; Qi, X.; Ren, J. miR-145 sensitizes breast cancer to doxorubicin by targeting multidrug resistance-associated protein-1. Oncotarget 2016, 7, 59714–59726. [Google Scholar] [CrossRef]
- Qian, P.X.; Banerjee, A.; Wu, Z.S.; Zhang, X.; Wang, H.; Pandey, V.; Zhang, W.J.; Lv, X.F.; Tan, S.; Lobie, P.E.; et al. Loss of SNAIL regulated miR-128-2 on chromosome 3p22.3 targets multiple stem cell factors to promote transformation of mammary epithelial cells. Cancer Res. 2012, 72, 6036–6050. [Google Scholar] [CrossRef] [PubMed]
- Ma, M.-T.; He, M.; Wang, Y.; Jiao, X.-Y.; Zhao, L.; Bai, X.-F.; Yu, Z.-J.; Wu, H.-Z.; Sun, M.-L.; Song, Z.-G.; et al. MiR-487a resensitizes mitoxantrone (MX)-resistant breast cancer cells (MCF-7/MX) to MX by targeting breast cancer resistance protein (BCRP/ABCG2). Cancer Lett. 2013, 339, 107–115. [Google Scholar] [CrossRef]
- Zhu, W.-J.; Chen, X.; Wang, Y.-W.; Liu, H.-T.; Ma, R.-R.; Gao, P. MiR-1268b confers chemosensitivity in breast cancer by targeting ERBB2-mediated PI3K-AKT pathway. Oncotarget 2017, 8, 89631–89642. [Google Scholar] [CrossRef]
- Li, X.; Ji, M.; Zhong, S.; Zha, Q.; Xu, J.; Zhao, J.; Tang, J. MicroRNA-34a modulates chemosensitivity of breast cancer cells to adriamycin by targeting Notch1. Arch. Med. Res. 2012, 43, 514–521. [Google Scholar] [CrossRef]
- Li, Z.-H.; Weng, X.; Xiong, Q.-Y.; Tu, J.-H.; Xiao, A.; Qiu, W.; Gong, Y.; Hu, E.-W.; Huang, S.; Cao, Y.-L. miR-34a expression in human breast cancer is associated with drug resistance. Oncotarget 2017, 8, 106270–106282. [Google Scholar] [CrossRef]
- Wu, M.-Y.; Fu, J.; Xiao, X.; Wu, J.; Wu, R.-C. MiR-34a regulates therapy resistance by targeting HDAC1 and HDAC7 in breast cancer. Cancer Lett. 2014, 354, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Li, L.; Yang, S.; Wang, D.; Zhong, S.; Zhao, J.; Tang, J. MicroRNA-29a contributes to drug-resistance of breast cancer cells to adriamycin through PTEN/AKT/GSK3β signaling pathway. Gene 2016, 593, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Dai, H.; Xu, L.-Y.; Qian, Q.; Zhu, Q.-W.; Chen, W.-X. MicroRNA-222 promotes drug resistance to doxorubicin in breast cancer via regulation of miR-222/bim pathway. Biosci. Rep. 2019, 39, BSR20190650. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Yang, S.; Chen, X.; Shen, H.-Y.; Luo, L.; Zhang, X.; Zhong, S.; Zhao, J.; Tang, J. miR-222 induces Adriamycin resistance in breast cancer through PTEN/Akt/p27kip1 pathway. Tumor Biol. 2016, 37, 15315–15324. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-W.; Guan, W.; Han, S.; Hong, D.-K.; Kim, L.-S.; Kim, H. MicroRNA-708-3p mediates metastasis and chemoresistance through inhibition of epithelial-to-mesenchymal transition in breast cancer. Cancer Sci. 2018, 109, 1404–1413. [Google Scholar] [CrossRef] [PubMed]
- Lv, J.; Xia, K.; Xu, P.; Sun, E.; Ma, J.; Gao, S.; Zhou, Q.; Zhang, M.; Wang, F.; Chen, F.; et al. miRNA expression patterns in chemoresistant breast cancer tissues. Biomed. Pharmacother. 2014, 68, 935–942. [Google Scholar] [CrossRef]
- Zhang, X.; Zhong, S.; Xu, Y.; Yu, D.; Ma, T.; Chen, L.; Zhao, Y.; Chen, X.; Yang, S.; Wu, Y.; et al. MicroRNA-3646 Contributes to Docetaxel Resistance in Human Breast Cancer Cells by GSK-3β/β-Catenin Signaling Pathway. PLoS ONE 2016, 11, e0153194. [Google Scholar] [CrossRef]
- Feng, X.; Zhao, L.; Gao, S.; Song, X.; Dong, W.; Zhao, Y.; Zhou, H.; Cheng, L.; Miao, X.; Jia, L. Increased fucosylation has a pivotal role in multidrug resistance of breast cancer cells through miR-224-3p targeting FUT4. Gene 2016, 578, 232–241. [Google Scholar] [CrossRef]
- Long, J.; Ji, Z.; Jiang, K.; Wang, Z.; Meng, G. miR-193b Modulates Resistance to Doxorubicin in Human Breast Cancer Cells by Downregulating MCL-1. Biomed. Res. Int. 2015, 2015, 373574. [Google Scholar] [CrossRef]
- Wang, Z.-X.; Lu, B.-B.; Wang, H.; Cheng, Z.-X.; Yin, Y.-M. MicroRNA-21 modulates chemosensitivity of breast cancer cells to doxorubicin by targeting PTEN. Arch. Med. Res. 2011, 42, 281–290. [Google Scholar] [CrossRef]
- Yuan, Y.; Yao, Y.F.; Hu, S.N.; Gao, J.; Zhang, L.-L. MiR-133a Is Functionally Involved in Doxorubicin-Resistance in Breast Cancer Cells MCF-7 via Its Regulation of the Expression of Uncoupling Protein 2. PLoS ONE 2015, 10, e0129843. [Google Scholar] [CrossRef]
- Yu, S.-J.; Yang, L.; Hong, Q.; Kuang, X.-Y.; Di, G.-H.; Shao, Z.-M. MicroRNA-200a confers chemoresistance by antagonizing TP53INP1 and YAP1 in human breast cancer. BMC Cancer 2018, 18, 74. [Google Scholar] [CrossRef] [PubMed]
- He, D.-X.; Gu, X.-T.; Li, Y.-R.; Jiang, L.; Jin, J.; Ma, X. Methylation-regulated miR-149 modulates chemoresistance by targeting GlcNAc N -deacetylase/N -sulfotransferase-1 in human breast cancer. FEBS J. 2014, 281, 4718–4730. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.-S.; Qiu, W.-S.; Yao, R.-Y.; Zhang, Q.; Zhuang, L.-K.; Zhou, F.; Sun, L.-B.; Yue, L. miR-141 confers docetaxel chemoresistance of breast cancer cells via regulation of EIF4E expression. Oncol. Rep. 2015, 33, 2504–2512. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Hu, Y.; Xu, L.; Fu, Y.; Tu, J.; Zhao, H.; Zhang, S.; Hong, R.; Gu, X. The role of miR-125b-mitochondria-caspase-3 pathway in doxorubicin resistance and therapy in human breast cancer. Tumor Biol. 2015, 36, 7185–7194. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Wang, Y.; Lu, X.; Zhao, Z.; Zhu, L.; Chen, S.; Wu, Q.; Chen, C.; Wang, Z. MiR-125b regulates epithelial-mesenchymal transition via targeting Sema4C in paclitaxel-resistant breast cancer cells. Oncotarget 2015, 6, 3268–3279. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Liu, Z.; Zhao, Y.; Ding, Y.; Liu, H.; Xi, Y.; Xiong, W.; Li, G.; Lu, J.; Fodstad, O.; et al. MicroRNA-125b confers the resistance of breast cancer cells to paclitaxel through suppression of pro-apoptotic Bcl-2 antagonist killer 1 (Bak1) expression. J. Biol. Chem. 2010, 285, 21496–21507. [Google Scholar] [CrossRef]
- Hu, G.; Zhao, X.; Wang, J.; Lv, L.; Wang, C.; Feng, L.; Shen, L.; Ren, W. miR-125b regulates the drug-resistance of breast cancer cells to doxorubicin by targeting HAX-1. Oncol. Lett. 2018, 15, 1621–1629. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, Y.; Wei, Y.; Li, M.; Yu, S.; Ye, M.; Zhang, H.; Chen, S.; Liu, W.; Zhang, J. MiR-129-3p promotes docetaxel resistance of breast cancer cells via CP110 inhibition. Sci. Rep. 2015, 5, 1–12. [Google Scholar] [CrossRef]
- Zhang, H.; Sun, D.; Mao, L.; Zhang, J.; Jiang, L.; Li, J.; Wu, Y.; Ji, H.; Chen, W.; Wang, J.; et al. MiR-139-5p inhibits the biological function of breast cancer cells by targeting Notch1 and mediates chemosensitivity to docetaxel. Biochem. Biophys. Res. Commun. 2015, 465, 702–713. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.-H.; Wang, C.-H.; Huang, Z.-J.; Liu, F.; Xu, C.-W.; Li, X.-L.; Chen, G.-Q. miR-760 mediates chemoresistance through inhibition of epithelial mesenchymal transition in breast cancer cells. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 5002–5008. [Google Scholar] [PubMed]
- Ye, F.G.; Song, C.G.; Cao, Z.G.; Xia, C.; Chen, D.N.; Chen, L.; Li, S.; Qiao, F.; Ling, H.; Yao, L.; et al. Cytidine deaminase axis modulated by mir-484 differentially regulates cell proliferation and chemoresistance in breast cancer. Cancer Res. 2015, 75, 1504–1515. [Google Scholar] [CrossRef]
- Wang, X.; Wang, X.; Gu, J.; Zhou, M.; He, Z.; Wang, X.; Ferrone, S. Overexpression of miR-489 enhances efficacy of 5-fluorouracil-based treatment in breast cancer stem cells by targeting XIAP. Oncotarget 2017, 8, 113837–113846. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wang, Y.-W.; Xing, A.-Y.; Xiang, S.; Shi, D.-B.; Liu, L.; Li, Y.-X.; Gao, P. Suppression of SPIN1-mediated PI3K-Akt pathway by miR-489 increases chemosensitivity in breast cancer. J. Pathol. 2016, 239, 459–472. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; He, D.; Yang, D.; Chen, Z.; Pan, Q.; Mao, A.; Cai, Y.; Li, X.; Xing, H.; Shi, M.; et al. MiR-489 regulates chemoresistance in breast cancer via epithelial mesenchymal transition pathway. FEBS Lett. 2014, 588, 2009–2015. [Google Scholar] [CrossRef]
- Hu, H.; Li, S.; Cui, X.; Lv, X.; Jiao, Y.; Yu, F.; Yao, H.; Song, E.; Chen, Y.; Wang, M.; et al. The overexpression of hypomethylated miR-663 induces chemotherapy resistance in human breast cancer cells by targeting heparin sulfate proteoglycan 2 (HSPG2). J. Biol. Chem. 2013, 288, 10973–10985. [Google Scholar] [CrossRef]
- Kopp, F.; Oak, P.S.; Wagner, E.; Roidl, A. miR-200c Sensitizes Breast Cancer Cells to Doxorubicin Treatment by Decreasing TrkB and Bmi1 Expression. PLoS ONE 2012, 7, e50469. [Google Scholar] [CrossRef]
- Li, H.-Y.; Liang, J.-L.; Kuo, Y.-L.; Lee, H.-H.; Calkins, M.J.; Chang, H.-T.; Lin, F.-C.; Chen, Y.-C.; Hsu, T.-I.; Hsiao, M.; et al. miR-105/93-3p promotes chemoresistance and circulating miR-105/93-3p acts as a diagnostic biomarker for triple negative breast cancer. Breast Cancer Res. 2017, 19, 133. [Google Scholar] [CrossRef]
- Roscigno, G.; Puoti, I.; Giordano, I.; Donnarumma, E.; Russo, V.; Affinito, A.; Adamo, A.; Quintavalle, C.; Todaro, M.; dM Vivanco, M.; et al. MiR-24 induces chemotherapy resistance and hypoxic advantage in breast cancer. Oncotarget 2017, 8, 19507–19521. [Google Scholar] [CrossRef]
- Si, W.; Shen, J.; Du, C.; Chen, D.; Gu, X.; Li, C.; Yao, M.; Pan, J.; Cheng, J.; Jiang, D.; et al. A miR-20a/MAPK1/c-Myc regulatory feedback loop regulates breast carcinogenesis and chemoresistance. Cell Death Differ. 2018, 25, 406–420. [Google Scholar] [CrossRef]
- Jiao, X.; Zhao, L.; Ma, M.; Bai, X.; He, M.; Yan, Y.; Wang, Y.; Chen, Q.; Zhao, X.; Zhou, M.; et al. MiR-181a enhances drug sensitivity in mitoxantone-resistant breast cancer cells by targeting breast cancer resistance protein (BCRP/ABCG2). Breast Cancer Res. Treat. 2013, 139, 717–730. [Google Scholar] [CrossRef]
- Niu, J.; Xue, A.; Chi, Y.; Xue, J.; Wang, W.; Zhao, Z.; Fan, M.; Yang, C.H.; Shao, Z.M.; Pfeffer, L.M.; et al. Induction of miRNA-181a by genotoxic treatments promotes chemotherapeutic resistance and metastasis in breast cancer. Oncogene 2016, 35, 1302–1313. [Google Scholar] [CrossRef]
- Su, C.-M.; Wang, M.-Y.; Hong, C.-C.; Chen, H.-A.; Su, Y.-H.; Wu, C.-H.; Huang, M.-T.; Chang, Y.-W.; Jiang, S.-S.; Sung, S.-Y.; et al. miR-520h is crucial for DAPK2 regulation and breast cancer progression. Oncogene 2016, 35, 1134–1142. [Google Scholar] [CrossRef]
- Sha, L.-Y.; Zhang, Y.; Wang, W.; Sui, X.; Liu, S.-K.; Wang, T.; Zhang, H. MiR-18a upregulation decreases Dicer expression and confers paclitaxel resistance in triple negative breast cancer. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 2201–2208. [Google Scholar] [PubMed]
- Liu, X.; Tang, H.; Chen, J.; Song, C.; Yang, L.; Liu, P.; Wang, N.; Xie, X.; Lin, X.; Xie, X. MicroRNA-101 inhibits cell progression and increases paclitaxel sensitivity by suppressing MCL-1 expression in human triple-negative breast cancer. Oncotarget 2015, 6, 20070–20083. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Zhou, S.; Zheng, M.; Deng, X.; Yi, Y.; Huang, T. MiR-199a-3p enhances breast cancer cell sensitivity to cisplatin by downregulating TFAM (TFAM). Biomed. Pharmacother. 2017, 88, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Rinn, J.L.; Chang, H.Y. Genome regulation by long noncoding RNAs. Annu. Rev. Biochem. 2012, 81, 145–166. [Google Scholar] [CrossRef]
- Flynn, R.A.; Chang, H.Y. Long noncoding RNAs in cell-fate programming and reprogramming. Cell Stem Cell 2014, 14, 752–761. [Google Scholar] [CrossRef]
- Jiang, C.; Li, Y.; Zhao, Z.; Lu, J.; Chen, H.; Ding, N.; Wang, G.; Xu, J.; Li, X. Identifying and functionally characterizing tissue-specific and ubiquitously expressed human lncRNAs. Oncotarget 2016, 7, 7120–7133. [Google Scholar] [CrossRef]
- Mattioli, K.; Volders, P.J.; Gerhardinger, C.; Lee, J.C.; Maass, P.G.; Melé, M.; Rinn, J.L. High-throughput functional analysis of lncRNA core promoters elucidates rules governing tissue specificity. Genome Res. 2019, 29, 344–355. [Google Scholar] [CrossRef]
- Richard, J.L.C.; Eichhorn, P.J.A. Deciphering the roles of lncRNAs in breast development and disease. Oncotarget 2018, 9, 20179–20212. [Google Scholar] [CrossRef] [PubMed]
- Ponting, C.P.; Oliver, P.L.; Reik, W. Evolution and functions of long noncoding RNAs. Cell 2009, 136, 629–641. [Google Scholar] [CrossRef] [PubMed]
- Rinn, J.L.; Kertesz, M.; Wang, J.K.; Squazzo, S.L.; Xu, X.; Brugmann, S.A.; Goodnough, L.H.; Helms, J.A.; Farnham, P.J.; Segal, E.; et al. Functional demarcation of active and silent chromatin domains in human HOX loci by noncoding RNAs. Cell 2007, 129, 1311–1323. [Google Scholar] [CrossRef]
- Wang, K.C.; Chang, H.Y. Molecular mechanisms of long noncoding RNAs. Mol. Cell 2011, 43, 904–914. [Google Scholar] [CrossRef] [PubMed]
- Yao, R.-W.; Wang, Y.; Chen, L.-L. Cellular functions of long noncoding RNAs. Nat. Cell Biol. 2019, 21, 542–551. [Google Scholar] [CrossRef]
- Flicek, P.; Amode, M.R.; Barrell, D.; Beal, K.; Billis, K.; Brent, S.; Carvalho-Silva, D.; Clapham, P.; Coates, G.; Fitzgerald, S.; et al. Ensembl 2014. Nucleic Acids Res. 2014, 42, D749–D755. [Google Scholar] [CrossRef]
- Fatica, A.; Bozzoni, I. Long non-coding RNAs: New players in cell differentiation and development. Nat. Rev. Genet. 2014, 15, 7–21. [Google Scholar] [CrossRef]
- Gibb, E.A.; Brown, C.J.; Lam, W.L. The functional role of long non-coding RNA in human carcinomas. Mol. Cancer 2011, 10, 1–17. [Google Scholar] [CrossRef]
- Chen, S.; Zhu, J.; Wang, F.; Guan, Z.; Ge, Y.; Yang, X.; Cai, J. LncRNAs and their role in cancer stem cells. Oncotarget 2017, 8, 110685–110692. [Google Scholar] [CrossRef]
- Chou, J.; Wang, B.; Zheng, T.; Li, X.; Zheng, L.; Hu, J.; Zhang, Y.; Xing, Y.; Xi, T. MALAT1 induced migration and invasion of human breast cancer cells by competitively binding miR-1 with cdc42. Biochem. Biophys. Res. Commun. 2016, 472, 262–269. [Google Scholar] [CrossRef]
- Zhang, H.-Y.; Liang, F.; Zhang, J.-W.; Wang, F.; Wang, L.; Kang, X.-G. Effects of long noncoding RNA-ROR on tamoxifen resistance of breast cancer cells by regulating microRNA-205. Cancer Chemother. Pharmacol. 2017, 79, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Song, Y.; Wang, Z.; Zhang, Z.; Lu, M.; Wang, Y. Long Non-coding RNA LINC01787 Drives Breast Cancer Progression via Disrupting miR-125b Generation. Front. Oncol. 2019, 9, 1140. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Li, J.; Xiong, Y.-F.; Zeng, Z.; Zhang, X.; Li, H.-Y. A Potential Prognostic Long Noncoding RNA Signature to Predict Recurrence among ER-positive Breast Cancer Patients Treated with Tamoxifen. Sci. Rep. 2018, 8, 3179. [Google Scholar] [CrossRef]
- Xue, X.; Yang, Y.A.; Zhang, A.; Fong, K.-W.; Kim, J.; Song, B.; Li, S.; Zhao, J.C.; Yu, J. LncRNA HOTAIR enhances ER signaling and confers tamoxifen resistance in breast cancer. Oncogene 2016, 35, 2746–2755. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.Y.; Oh, J.H.; Lee, J.-Y.; Kim, M.H. The LncRNA HOTAIRM1 Promotes Tamoxifen Resistance by Mediating HOXA1 Expression in ER+ Breast Cancer Cells. J. Cancer 2020, 11, 3416–3423. [Google Scholar] [CrossRef]
- Godinho, M.F.E.; Sieuwerts, A.M.; Look, M.P.; Meijer, D.; Foekens, J.A.; Dorssers, L.C.J.; van Agthoven, T. Relevance of BCAR4 in tamoxifen resistance and tumour aggressiveness of human breast cancer. Br. J. Cancer 2010, 103, 1284–1291. [Google Scholar] [CrossRef]
- Godinho, M.F.E.; Wulfkuhle, J.D.; Look, M.P.; Sieuwerts, A.M.; Sleijfer, S.; Foekens, J.A.; Petricoin, E.F.; Dorssers, L.C.J.; van Agthoven, T. BCAR4 induces antioestrogen resistance but sensitises breast cancer to lapatinib. Br. J. Cancer 2012, 107, 947–955. [Google Scholar] [CrossRef]
- Cai, Y.; He, J.; Zhang, D. Long noncoding RNA CCAT2 promotes breast tumor growth by regulating the Wnt signaling pathway. OncoTargets Ther. 2015, 8, 2657–2664. [Google Scholar] [CrossRef]
- Wu, Z.-J.; Li, Y.; Wu, Y.-Z.; Wang, Y.; Nian, W.-Q.; Wang, L.-L.; Li, L.-C.; Luo, H.-L.; Wang, D.-L. Long non-coding RNA CCAT2 promotes the breast cancer growth and metastasis by regulating TGF-β signaling pathway. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 706–714. [Google Scholar]
- Cai, Y.; He, J.; Zhang, D. Suppression of long non-coding RNA CCAT2 improves tamoxifen-resistant breast cancer cells’ response to tamoxifen]. Mol. Biol. 2016, 50, 821–827. [Google Scholar] [CrossRef]
- Peng, W.-X.; Huang, J.-G.; Yang, L.; Gong, A.-H.; Mo, Y.-Y. Linc-RoR promotes MAPK/ERK signaling and confers estrogen-independent growth of breast cancer. Mol. Cancer 2017, 16, 161. [Google Scholar] [CrossRef]
- Li, Z.; Yu, D.; Li, H.; Lv, Y.; Li, S. Long non-coding RNA UCA1 confers tamoxifen resistance in breast cancer endocrinotherapy through regulation of the EZH2/p21 axis and the PI3K/AKT signaling pathway. Int. J. Oncol. 2019, 54, 1033–1042. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Luo, J. Long Non-Coding RNA (lncRNA) Urothelial Carcinoma-Associated 1 (UCA1) Enhances Tamoxifen Resistance in Breast Cancer Cells via Inhibiting mTOR Signaling Pathway. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2016, 22, 3860–3867. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wang, G.; Yang, L.; Qu, J.; Yang, Z.; Zhou, X. Knockdown of Long Non-Coding RNA UCA1 Increases the Tamoxifen Sensitivity of Breast Cancer Cells through Inhibition of Wnt/β-Catenin Pathway. PLoS ONE 2016, 11, e0168406. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wu, Y.; Liu, A.; Tang, X. Long non-coding RNA UCA1 enhances tamoxifen resistance in breast cancer cells through a miR-18a-HIF1α feedback regulatory loop. Tumour Biol. J. Int. Soc. Oncodev. Biol. Med. 2016, 37, 14733–14743. [Google Scholar] [CrossRef]
- Xu, C.-G.; Yang, M.-F.; Ren, Y.-Q.; Wu, C.-H.; Wang, L.-Q. Exosomes mediated transfer of lncRNA UCA1 results in increased tamoxifen resistance in breast cancer cells. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 4362–4368. [Google Scholar]
- Gu, J.; Wang, Y.; Wang, X.; Zhou, D.; Shao, C.; Zhou, M.; He, Z. Downregulation of lncRNA GAS5 confers tamoxifen resistance by activating miR-222 in breast cancer. Cancer Lett. 2018, 434, 1–10. [Google Scholar] [CrossRef]
- Gregory, P.A.; Bracken, C.P.; Smith, E.; Bert, A.G.; Wright, J.A.; Roslan, S.; Morris, M.; Wyatt, L.; Farshid, G.; Lim, Y.-Y.; et al. An autocrine TGF-beta/ZEB/miR-200 signaling network regulates establishment and maintenance of epithelial-mesenchymal transition. Mol. Biol. Cell 2011, 22, 1686–1698. [Google Scholar] [CrossRef]
- Dong, H.; Wang, W.; Chen, R.; Zhang, Y.; Zou, K.; Ye, M.; He, X.; Zhang, F.; Han, J. Exosome-mediated transfer of lncRNA-SNHG14 promotes trastuzumab chemoresistance in breast cancer. Int. J. Oncol. 2018, 53, 1013–1026. [Google Scholar] [CrossRef]
- Sun, Z.; Zhang, C.; Wang, T.; Shi, P.; Tian, X.; Guo, Y. Correlation between long non-coding RNAs (lncRNAs) H19 expression and trastuzumab resistance in breast cancer. J. Cancer Res. Ther. 2019, 15, 933–940. [Google Scholar] [CrossRef]
- Zhu, H.-Y.; Bai, W.-D.; Ye, X.-M.; Yang, A.-G.; Jia, L.-T. Long non-coding RNA UCA1 desensitizes breast cancer cells to trastuzumab by impeding miR-18a repression of Yes-associated protein 1. Biochem. Biophys. Res. Commun. 2018, 496, 1308–1313. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; Hu, J.; Zou, K.; Ye, M.; Chen, Y.; Wu, C.; Chen, X.; Han, M. Activation of LncRNA TINCR by H3K27 acetylation promotes Trastuzumab resistance and epithelial-mesenchymal transition by targeting MicroRNA-125b in breast Cancer. Mol. Cancer 2019, 18, 3. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Liu, Z.; Zeng, W.; Huang, T. Down-regulation of long non-coding RNA HOTAIR sensitizes breast cancer to trastuzumab. Sci. Rep. 2019, 9, 19881. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Zhai, L.; Wang, H.; Liu, C.; Zhang, J.; Chen, W.; Wei, Q. Downregulation of LncRNA GAS5 causes trastuzumab resistance in breast cancer. Oncotarget 2016, 7, 27778–27786. [Google Scholar] [CrossRef]
- Hung, T.; Wang, Y.; Lin, M.F.; Koegel, A.K.; Kotake, Y.; Grant, G.D.; Horlings, H.M.; Shah, N.; Umbricht, C.; Wang, P.; et al. Extensive and coordinated transcription of noncoding RNAs within cell-cycle promoters. Nat. Genet. 2011, 43, 621–629. [Google Scholar] [CrossRef]
- Jiang, M.; Huang, O.; Xie, Z.; Wu, S.; Zhang, X.; Shen, A.; Liu, H.; Chen, X.; Wu, J.; Lou, Y.; et al. A novel long non-coding RNA-ARA: Adriamycin Resistance Associated. Biochem. Pharmacol. 2014, 87, 254–283. [Google Scholar] [CrossRef]
- Gu, M.; Zheng, W.; Zhang, M.; Dong, X.; Zhao, Y.; Wang, S.; Jiang, H.; Zheng, X. LncRNA NONHSAT141924 promotes paclitaxel chemotherapy resistance through p-CREB/Bcl-2 apoptosis signaling pathway in breast cancer. J. Cancer 2020, 11, 3645–3654. [Google Scholar] [CrossRef]
- Si, X.; Zang, R.; Zhang, E.; Liu, Y.; Shi, X.; Zhang, E.; Shao, L.; Li, A.; Yang, N.; Han, X.; et al. LncRNA H19 confers chemoresistance in ERα-positive breast cancer through epigenetic silencing of the pro-apoptotic gene BIK. Oncotarget 2016, 7, 81452–81462. [Google Scholar] [CrossRef]
- Gooding, A.J.; Zhang, B.; Gunawardane, L.; Beard, A.; Valadkhan, S.; Schiemann, W.P. The lncRNA BORG facilitates the survival and chemoresistance of triple-negative breast cancers. Oncogene 2019, 38, 2020–2041. [Google Scholar] [CrossRef]
- Xiu, D.-H.; Liu, G.-F.; Yu, S.-N.; Li, L.-Y.; Zhao, G.-Q.; Liu, L.; Li, X.-F. Long non-coding RNA LINC00968 attenuates drug resistance of breast cancer cells through inhibiting the Wnt2/β-catenin signaling pathway by regulating WNT2. J. Exp. Clin. Cancer Res. 2019, 38, 94. [Google Scholar] [CrossRef]
- Chen, Z.; Pan, T.; Jiang, D.; Jin, L.; Geng, Y.; Feng, X.; Shen, A.; Zhang, L. The lncRNA-GAS5/miR-221-3p/DKK2 Axis Modulates ABCB1-Mediated Adriamycin Resistance of Breast Cancer via the Wnt/β-Catenin Signaling Pathway. Mol. Ther. Nucleic Acids 2020, 19, 1434–1448. [Google Scholar] [CrossRef]
- Bida, O.; Gidoni, M.; Ideses, D.; Efroni, S.; Ginsberg, D. A novel mitosis-associated lncRNA, MA-linc1, is required for cell cycle progression and sensitizes cancer cells to Paclitaxel. Oncotarget 2015, 6, 27880–27890. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Jiang, F.; Zhang, X.; Xu, X. Long Non-Coding RNA UCA1 Modulates Paclitaxel Resistance in Breast Cancer via miR-613/CDK12 Axis. Cancer Manag. Res. 2020, 12, 2777–2788. [Google Scholar] [CrossRef] [PubMed]
- He, D.-X.; Zhang, G.-Y.; Gu, X.-T.; Mao, A.-Q.; Lu, C.-X.; Jin, J.; Liu, D.-Q.; Ma, X. Genome-wide profiling of long non-coding RNA expression patterns in anthracycline-resistant breast cancer cells. Int. J. Oncol. 2016, 49, 1695–1703. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Li, Y.; Song, X.; Zhang, N.; Sang, Y.; Zhang, H.; Liu, Y.; Chen, B.; Zhao, W.; Wang, L.; et al. Long noncoding RNA LINP1 acts as an oncogene and promotes chemoresistance in breast cancer. Cancer Biol. Ther. 2018, 19, 120–131. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Yuan, Y.; Zhang, L.; Yu, S.; Lu, J.; Feng, J.; Hu, S. Inhibition of ZEB1-AS1 confers cisplatin sensitivity in breast cancer by promoting microRNA-129-5p-dependent ZEB1 downregulation. Cancer Cell Int. 2020, 20, 90. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-M.; Liu, Y.; Wei, H.-Y.; Lv, K.-Z.; Fu, P. Linc-ROR induces epithelial-mesenchymal transition and contributes to drug resistance and invasion of breast cancer cells. Tumour Biol. J. Int. Soc. Oncodev. Biol. Med. 2016, 37, 10861–10870. [Google Scholar] [CrossRef]
- Fultang, N.; Illendula, A.; Lin, J.; Pandey, M.K.; Klase, Z.; Peethambaran, B. ROR1 regulates chemoresistance in Breast Cancer via modulation of drug efflux pump ABCB1. Sci. Rep. 2020, 10, 1821. [Google Scholar] [CrossRef]
- Wang, G.; Chen, X.; Liang, Y.; Wang, W.; Shen, K. A Long Noncoding RNA Signature That Predicts Pathological Complete Remission Rate Sensitively in Neoadjuvant Treatment of Breast Cancer. Transl. Oncol. 2017, 10, 988–997. [Google Scholar] [CrossRef]
- Zeng, Y.; Wang, G.; Zhou, C.-F.; Zhang, H.-B.; Sun, H.; Zhang, W.; Zhou, H.-H.; Liu, R.; Zhu, Y.-S. LncRNA Profile Study Reveals a Three-LncRNA Signature Associated With the Pathological Complete Response Following Neoadjuvant Chemotherapy in Breast Cancer. Front. Pharmacol. 2019, 10, 574. [Google Scholar] [CrossRef]
- Zhang, Y.; He, Q.; Hu, Z.; Feng, Y.; Fan, L.; Tang, Z.; Yuan, J.; Shan, W.; Li, C.; Hu, X.; et al. Long noncoding RNA LINP1 regulates repair of DNA double-strand breaks in triple-negative breast cancer. Nat. Struct. Mol. Biol. 2016, 23, 522–530. [Google Scholar] [CrossRef]
- Hu, X.-L.; Wang, J.; He, W.; Zhao, P.; Wu, W.-Q. Down-regulation of lncRNA Linc00152 suppressed cell viability, invasion, migration, and epithelial to mesenchymal transition, and reversed chemo-resistance in breast cancer cells. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 3074–3084. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Zeng, L.; Chu, J.; Xu, P.; Lv, M.; Xu, J.; Wen, J.; Li, W.; Wang, L.; Wu, X.; et al. Chemoresistance-related long non-coding RNA expression profiles in human breast cancer cells. Mol. Med. Rep. 2018, 18, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Song, X.; Li, Y.; Sang, Y.; Zhang, N.; Zhang, H.; Liu, Y.; Duan, Y.; Chen, B.; Guo, R.; et al. A novel long non-coding RNA-PRLB acts as a tumor promoter through regulating miR-4766-5p/SIRT1 axis in breast cancer. Cell Death Dis. 2018, 9, 563. [Google Scholar] [CrossRef] [PubMed]
- Balachandran, V.P.; Gonen, M.; Smith, J.J.; DeMatteo, R.P. Nomograms in oncology: More than meets the eye. Lancet. Oncol. 2015, 16, e173–e180. [Google Scholar] [CrossRef]
- Lai, J.; Wang, H.; Pan, Z.; Su, F. A novel six-microRNA-based model to improve prognosis prediction of breast cancer. Aging (Albany NY) 2019, 11, 649–662. [Google Scholar] [CrossRef]
- Rodríguez-González, F.G.; Sieuwerts, A.M.; Smid, M.; Look, M.P.; Meijer-Van Gelder, M.E.; De Weerd, V.; Sleijfer, S.; Martens, J.W.M.; Foekens, J.A. MicroRNA-30c expression level is an independent predictor of clinical benefit of endocrine therapy in advanced estrogen receptor positive breast cancer. Breast Cancer Res. Treat. 2011, 127, 43–51. [Google Scholar] [CrossRef]
- Uhr, K.; Sieuwerts, A.M.; De Weerd, V.; Smid, M.; Hammerl, D.; Foekens, J.A.; Martens, J.W.M. Association of microRNA-7 and its binding partner CDR1-AS with the prognosis and prediction of 1st-line tamoxifen therapy in breast cancer. Sci. Rep. 2018, 8, 1–14. [Google Scholar] [CrossRef]
- Wu, X.; Somlo, G.; Yu, Y.; Palomares, M.R.; Li, A.X.; Zhou, W.; Chow, A.; Yen, Y.; Rossi, J.J.; Gao, H.; et al. De novo sequencing of circulating miRNAs identifies novel markers predicting clinical outcome of locally advanced breast cancer. J. Transl. Med. 2012, 10, 42. [Google Scholar] [CrossRef]
- Falkenberg, N.; Anastasov, N.; Rappl, K.; Braselmann, H.; Auer, G.; Walch, A.; Huber, M.; Höfig, I.; Schmitt, M.; Höfler, H.; et al. MiR-221/-222 differentiate prognostic groups in advanced breast cancers and influence cell invasion. Br. J. Cancer 2013, 109, 2714–2723. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, L.; Dong, Z.; Xu, H.; Yan, L.; Yang, S.; Chen, T.; Chen, S.; Wang, W.; Yang, Q.; et al. MicroRNA-155-5p promotes tumor progression and contributes paclitaxel resistance to chemotherapy via TP53INP1 in human Breast Cancer. Research Square 2020. [Google Scholar] [CrossRef]
- Chandra Gupta, S.; Nandan Tripathi, Y. Potential of long non-coding RNAs in cancer patients: From biomarkers to therapeutic targets. Int. J. Cancer 2017, 140, 1955–1967. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Cui, Q.; Zhang, D.; Kong, D.; Liao, X.; Ren, J.; Gong, Y.; Wu, G. A prognostic eight-lncRNA expression signature in predicting recurrence of ER-positive breast cancer receiving endocrine therapy. J. Cell. Physiol. 2020, 235, 4746–4755. [Google Scholar] [CrossRef]
- Jiang, Y.-Z.; Liu, Y.-R.; Xu, X.-E.; Jin, X.; Hu, X.; Yu, K.-D.; Shao, Z.-M. Transcriptome Analysis of Triple-Negative Breast Cancer Reveals an Integrated mRNA-lncRNA Signature with Predictive and Prognostic Value. Cancer Res. 2016, 76, 2105–2114. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.; Xie, G.; Zhang, Z.; Wang, Z.; Wang, Y.; Wang, Y.; Qiu, Y.; Li, L.; Bu, H.; Li, J.; et al. MiR-4653-3p and its target gene FRS2 are prognostic biomarkers for hormone receptor positive breast cancer patients receiving tamoxifen as adjuvant endocrine therapy. Oncotarget 2016, 7, 61166–61182. [Google Scholar] [CrossRef]
- Amorim, M.; Lobo, J.; Fontes-Sousa, M.; Estevão-Pereira, H.; Salta, S.; Lopes, P.; Coimbra, N.; Antunes, L.; Palma de Sousa, S.; Henrique, R.; et al. Predictive and Prognostic Value of Selected MicroRNAs in Luminal Breast Cancer. Front. Genet. 2019, 10, 815. [Google Scholar] [CrossRef]
- Li, W.; Jin, X.; Zhang, Q.; Zhang, G.; Deng, X.; Ma, L. Decreased expression of miR-204 is associated with poor prognosis in patients with breast cancer. Int. J. Clin. Exp. Pathol. 2014, 7, 3287–3292. [Google Scholar]
- Bleckmann, A.; Leha, A.; Artmann, S.; Menck, K.; Salinas-Riester, G.; Binder, C.; Pukrop, T.; Beissbarth, T.; Klemm, F. Integrated miRNA and mRNA profiling of tumor-educated macrophages identifies prognostic subgroups in estrogen receptor-positive breast cancer. Mol. Oncol. 2015, 9, 155–166. [Google Scholar] [CrossRef]
- Rothé, F.; Ignatiadis, M.; Chaboteaux, C.; Haibe-Kains, B.; Kheddoumi, N.; Majjaj, S.; Badran, B.; Fayyad-Kazan, H.; Desmedt, C.; Harris, A.L.; et al. Global microRNA expression profiling identifies MiR-210 associated with tumor proliferation, invasion and poor clinical outcome in breast cancer. PLoS ONE 2011, 6, e20980. [Google Scholar] [CrossRef]
- Camps, C.; Buffa, F.M.; Colella, S.; Moore, J.; Sotiriou, C.; Sheldon, H.; Harris, A.L.; Gleadle, J.M.; Ragoussis, J. hsa-miR-210 Is induced by hypoxia and is an independent prognostic factor in breast cancer. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2008, 14, 1340–1348. [Google Scholar] [CrossRef]
- Vilquin, P.; Donini, C.F.; Villedieu, M.; Grisard, E.; Corbo, L.; Bachelot, T.; Vendrell, J.A.; Cohen, P.A. MicroRNA-125b upregulation confers aromatase inhibitor resistance and is a novel marker of poor prognosis in breast cancer. Breast Cancer Res. 2015, 17, 13. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Cao, X.; Zhang, L.; Zhang, X.; Sheng, H.; Tao, K. UCA1 overexpression predicts clinical outcome of patients with ovarian cancer receiving adjuvant chemotherapy. Cancer Chemother. Pharmacol. 2016, 77, 629–634. [Google Scholar] [CrossRef]
- Chen, X.; Lu, P.; Wang, D.-D.; Yang, S.-J.; Wu, Y.; Shen, H.-Y.; Zhong, S.-L.; Zhao, J.-H.; Tang, J.-H. The role of miRNAs in drug resistance and prognosis of breast cancer formalin-fixed paraffin-embedded tissues. Gene 2016, 595, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Su, F.; Chen, M.; Li, Y.; Qi, X.; Xiao, J.; Li, X.; Liu, X.; Liang, W.; Zhang, Y.; et al. Serum miR-21 and miR-125b as markers predicting neoadjuvant chemotherapy response and prognosis in stage II/III breast cancer. Hum. Pathol. 2017, 64, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yang, B.; Ma, B. The UCA1/miR-204/Sirt1 axis modulates docetaxel sensitivity of prostate cancer cells. Cancer Chemother. Pharmacol. 2016, 78, 1025–1031. [Google Scholar] [CrossRef]
- Bian, Z.; Jin, L.; Zhang, J.; Yin, Y.; Quan, C.; Hu, Y.; Feng, Y.; Liu, H.; Fei, B.; Mao, Y.; et al. LncRNA—UCA1 enhances cell proliferation and 5-fluorouracil resistance in colorectal cancer by inhibiting MIR-204-5p. Sci. Rep. 2016, 6, 1–12. [Google Scholar] [CrossRef]
- Xiao, Y.; Jiao, C.; Lin, Y.; Chen, M.; Zhang, J.; Wang, J.; Zhang, Z. lncRNA UCA1 Contributes to Imatinib Resistance by Acting as a ceRNA Against miR-16 in Chronic Myeloid Leukemia Cells. DNA Cell Biol. 2017, 36, 18–25. [Google Scholar] [CrossRef]
- Pan, J.; Li, X.; Wu, W.; Xue, M.; Hou, H.; Zhai, W.; Chen, W. Long non-coding RNA UCA1 promotes cisplatin/gemcitabine resistance through CREB modulating miR-196a-5p in bladder cancer cells. Cancer Lett. 2016, 382, 64–76. [Google Scholar] [CrossRef]
- Fang, Q.; Chen, X.; Zhi, X. Long Non-Coding RNA (LncRNA) Urothelial Carcinoma Associated 1 (UCA1) Increases Multi-Drug Resistance of Gastric Cancer via Downregulating miR-27b. Med. Sci. Monit. 2016, 22, 3506–3513. [Google Scholar] [CrossRef]
- Yu, W.-C.; Chen, H.-H.; Qu, Y.-Y.; Xu, C.-W.; Yang, C.; Liu, Y. MicroRNA-221 promotes cisplatin resistance in osteosarcoma cells by targeting PPP2R2A. Biosci. Rep. 2019, 39. [Google Scholar] [CrossRef]
- Zhao, L.; Zou, D.; Wei, X.; Wang, L.; Zhang, Y.; Liu, S.; Si, Y.; Zhao, H.; Wang, F.; Yu, J.; et al. MiRNA-221-3p desensitizes pancreatic cancer cells to 5-fluorouracil by targeting RB1. Tumor Biol. 2016, 37, 16053–16063. [Google Scholar] [CrossRef]
- Cao, L.; Chen, J.; Ou, B.; Liu, C.; Zou, Y.; Chen, Q. GAS5 knockdown reduces the chemo-sensitivity of non-small cell lung cancer (NSCLC) cell to cisplatin (DDP) through regulating miR-21/PTEN axis. Biomed. Pharmacother. 2017, 93, 570–579. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.-Q.; Wang, J.; Chen, D.-H.; Ma, X.-S.; Yang, W.; Zhe, T.; Dang, X.-W. Long non-coding RNA GAS5 antagonizes the chemoresistance of pancreatic cancer cells through down-regulation of miR-181c-5p. Biomed. Pharmacother. 2018, 97, 809–817. [Google Scholar] [CrossRef] [PubMed]
- Pickard, M.R.; Mourtada-Maarabouni, M.; Williams, G.T. Long non-coding RNA GAS5 regulates apoptosis in prostate cancer cell lines. Biochim. Biophys. Acta 2013, 1832, 1613–1623. [Google Scholar] [CrossRef] [PubMed]
- Cilek, E.E.; Ozturk, H.; Gur Dedeoglu, B. Construction of miRNA-miRNA networks revealing the complexity of miRNA-mediated mechanisms in trastuzumab treated breast cancer cell lines. PLoS ONE 2017, 12, e0185558. [Google Scholar] [CrossRef] [PubMed]
- Xie, M.; Ma, L.; Xu, T.; Pan, Y.; Wang, Q.; Wei, Y.; Shu, Y. Potential Regulatory Roles of MicroRNAs and Long Noncoding RNAs in Anticancer Therapies. Mol. Ther. Nucleic Acids 2018, 13, 233–243. [Google Scholar] [CrossRef]
- Aggarwal, V.; Priyanka, K.; Tuli, H.S. Emergence of Circulating MicroRNAs in Breast Cancer as Diagnostic and Therapeutic Efficacy Biomarkers. Mol. Diagn. Ther. 2020, 24, 153–173. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ncRNAs | Drug | Status | Impact on Drug Resistance | Target(s) | Mechanism(s) | Experimental System | References |
|---|---|---|---|---|---|---|---|
| miR-342 | Tamoxifen | d | + | BMP7, GEMIN4 | Apoptosis evasion | MCF-7 | [78] |
| miR-320 | Tamoxifen | d | + | ARPP-19, ERRγ | Cell proliferation | MCF-7, T47D | [79] |
| miR-451a | Tamoxifen | u | − | 14-3-3ζ, ERα | Erα signaling and cell survival | MCF-7 | [80] |
| miR-15a/16 | Tamoxifen | u | - | Cyclin E1, Bcl-2 | Cell cycle regulation, apoptosis evasion | MCF-7 | [81] |
| miR-21 | Tamoxifen fulvestrant | u | + | PTEN | PI3K/Akt activation, autophagy | MCF-7 | [82] |
| miR-155-5p | Tamoxifen | u | + | SOCS6 | STAT3 pathway | MCF-7, SKBR3 | [87] |
| miR-221/222 | Tamoxifen fulvestrant | u | + | p27, ERα | Erα signaling, cell cycle regulation, EMT | MCF-7, BT474 | [55,88] |
| miR-200 | Tamoxifen | d | + | MYB | EMT | MCF-7 | [89] |
| miR-873 | Tamoxifen | d | + | CDK3 | Erα signaling and cell cycle progression | MCF-7 | [90] |
| miR-10b | Tamoxifen | u | + | HDAC | TGF-β1-induced EMT | MCF-7, T47D | [91] |
| miR-148a miR152 | Tamoxifen | u | − | ALCAM | Tumor growth and survival | MCF-7 | [92] |
| miR-27b-3p | Tamoxifen | d | + | NR5A2/CREB1 | Cell proliferation | MCF-7, T47D | [93] |
| miR-27b | Tamoxifen | d | + | HMGB3 | EMT | MCF-7 | [94] |
| miR-214 | Tamoxifen fulvestrant | u | − | UCP-2 | Autophagy | MCF-7 | [95] |
| ncRNAs | Drug | Status | Impact on Drug Resistance | Target(s) | Mechanism(s) | Experimental System | References |
|---|---|---|---|---|---|---|---|
| miR-205-5p | Trastuzumab | u | + | ERBB2, EGFR | Cell survival | BCSCs | [24] |
| miR-205 | Trastuzumab | u | − | NA | Akt impairment | SKBR3 | [26] |
| mir-375 | Trastuzumab | u | − | IGF1R | Cell survival | SKBR3 | [99] |
| miR-630 | Lapatinib neratinib afatinib | u | − | IGF1R | Cell survival | SKBR3, HCC1954 | [100] |
| miR-210 | Trastuzumab | u | − | E2F3, RAD52 | DNA damage repair and cell proliferation | BT474, patient serum | [101,102] |
| miR-21 | Trastuzumab | u | + | PTEN, PDCD4 | PI3K/Akt activation, EMT and inflammatory signals | SKBR3, BC patient tissues | [103] |
| miR-221 | Trastuzumab | u | + | PTEN | PI3K/Akt/mTOR pathway | SKBR3 | [104] |
| miR-16 | Trastuzumab lapatinib | u | − | CCNJ, FUBP1 | Cell proliferation and survival | BT474, SKBR3, HCC-1569, MDA-MB-453 | [105] |
| miR-567 | Trastuzumab | d | + | ATG5 | Autophagy | SKBR-3, BT474 | [106] |
| miR-200c | Trastuzumab | u | − | ZNF217, ZEB1 | TGF-β signaling-induced EMT | SKBR3 | [107] |
| miR-7 | Trastuzumab | u | - | EGFR, Src kinase | ERBB2-driven proliferation | MCF-7 | [108] |
| ncRNA(s) | Drug | Status | Impact on Drug Resistance | Target(s) | Mechanism(s) | Experimental System | References |
|---|---|---|---|---|---|---|---|
| miR-302a-3p miR-302b-3p miR-302c-3p miR-302d-3p | Doxorubicin | u u u u | − − − − | MEKK1/P-gp | Drug efflux | MCF-7 | [25] |
| miR-451 | Doxorubicin | d | + | P-gp (ABCB1/MDR-1) | Drug efflux | MCF7 | [111] |
| Paclitaxel | u | − | YWHAZ | Cell cycle control, apoptosis and invasion | MCF-7, SKBR3 | [142] | |
| miR-298 | Doxorubicin | u | − | P-gp | Drug efflux | MDA-MB-231 | [143] |
| miR-326 | Doxorubicin VP-16 | d | + | ABCC1 (MRP-1) | Drug efflux | MCF-7 | [144] |
| miR-320a | Doxorubicin paclitaxel | d | + | TRPC5, NFATC3, ETS1 | P-gp-mediated drug efflux | MCF-7 | [145] |
| miR-145-5p | Doxorubicin | u | − | ABCC1 | Drug efflux | MCF-7 | [146] |
| miR-128 | Doxorubicin | d | + | Bmi1, ABCC5 | Drug efflux, stemness | SK-3RD and MCF-7 mammosphere | [147] |
| miR-25a | Adriamycin | u | + | Unc51 | ABCG2 regulation | MCF-7 | [126] |
| miR-487a | Mitoxantrone | d | + | ABCG2 (BCRP) | Drug efflux | MCF-7 | [148] |
| miR-449 | Doxorubicin | u | − | E2F1, E2F3, CDK2 | Cell cycle regulation | MDA-MB-231 | [113] |
| miR-1268b | Doxorubicin | u | − | ERBB2 | PI3K/Akt pathway regulation | MCF-7 | [149] |
| miR-222 miR-29a | Doxorubicin docetaxel | u u | + + | PTEN | PI3K/Akt pathway activation | MCF-7 | [116] |
| miR-34a | MDR | d | + | Bcl-2, CCND1, Notch | Survival pathways, cell cycle progression | MCF-7, MDA-MB-231 | [54,150,151] |
| d | + | HDAC1, HDAC7 | Cell survival and autophagy | MCF-7, MDA-MB-231 | [152] | ||
| miR-29a | Doxorubicin | u | + | PTEN | PI3K/Akt/GSK3β signaling regulation | MCF-7 | [153] |
| miR-31 | Doxorubicin | u | − | PKCepsilon/Bcl2 axis | Cell survival | MCF10A, MDA-MB-231 | [124] |
| miR-222 | Doxorubicin | u | + | Bim | Apoptosis/survival pathways | MCF-7, patients’ serum | [154] |
| Doxorubicin | u | + | PTEN/Akt/p27 kip1 | Survival pathway | MCF-7 | [155] | |
| miR-708 | Doxorubicin | d | + | ZEB1, CDH2, vimentin | EMT | MCF-7, MDA-MB-468 | [156] |
| miR-218 | Doxorubicin taxol | u | − | Survivin | Apoptosis | MCF-7, Cal51, mouse tumor xenografts | [57] |
| miR-141, miR-200c, miR-31 let-7e, miR-576-3p, miR-125b-1, miR-370, miR-145, miR-765, miR-760 | Doxorubicin | u d | + + | Genes involved in MAPK signaling pathway, regulation of the actin cytoskeleton, cytokine–cytokine receptor interaction | MCF-7 | [157] | |
| miR-302b | Cisplatin | u | − | E2F1, ATM signaling | Cell cycle progression, DNA damage repair | MDA-MB-231 | [112] |
| miR-221 | Cisplatin | d | − | Bim-Bax/Bak | Apoptosis | MDA-MB-231 | [122] |
| miR-30c | Doxorubicin paclitaxel | d | + | TWF1/IL-11 | EMT | T47D, MCF-7, MDA-MB-231, BT-20, HCC-70, HCC-38 | [131] |
| miR-3646 | Docetaxel | u | + | GSK-3β/β-catenin | Cell survival and proliferation | MDA-MB-231, MCF-7 | [158] |
| miR-224-3p | MDR | d | + | FUT4 | Cell growth and survival | T47D | [159] |
| miR-193b | Doxorubicin | d | + | MCL-1 | Apoptosis | MCF-7 | [160] |
| miR-21-5p | Docetaxel | u | + | PTEN | Cell survival | MCF-7 | [161] |
| miR-133a | Doxorubicin | d | + | UCP-2 | Cell metabolism alteration | MCF-7, mouse xenograft models | [162] |
| miR-200a | Cisplatin gemcitabine paclitaxel | u | + | TP53INP1, YAP1 | EMT | ZR-75-30, MDA-MB-231 | [163] |
| miR-149 | Doxorubicin | d | + | NDST1 | Tumor growth and angiogenesis | MCF-7 | [164] |
| miR-141 | Docetaxel | u | + | EIF4E | Apoptosis | MCF-7, MDA-MB-231 | [165] |
| miR-125b | Doxorubicin | u | − | Mcl-1 | Apoptosis | MCF-7 | [166] |
| Paclitaxel | u | − | Sema4C | EMT | MCF7, SKBR3 | [167] | |
| Paclitaxel | u | + | Bak1 | Apoptosis | MDA-MB-435, MDA-MB-436, SKBR3 | [168] | |
| Doxorubicin | u | − | HAX1 | Apoptosis | MCF-7 | [169] | |
| miR-129-3p | Docetaxel | u | + | CP110 | Cell cycle control and apoptosis | MDA-MB-231, MCF-7, nude xenograft model | [170] |
| miR-542-3p | Paclitaxel | u | − | Survivin | Apoptosis | SKBR3, BT474, MDA-MB-453, HCC1954 | [118] |
| miR-25-3p | Epirubicin | d | − | ULK1 | Autophagy | MCF-7 | [126] |
| miR-139-5p | Docetaxel | d | + | Notch-1 | Cell cycle control and survival, invasion | MCF-7 | [171] |
| miR-760 | Doxorubicin | d | + | Nanog | EMT | MCF-7 | [172] |
| miR-484 | Gemcitabine | u | − | CDA | Proliferation and cell cycle control | MDA-MB-231 | [173] |
| miR-489 | Doxorubicin | u | − | ULK1, LAPTM4B | Autophagy and cell viability | T47D, MDA-MB-231 | [28] |
| 5-FU | u | − | XIAP | Survival pathways | T47D, SKBR3 | [174] | |
| Doxorubicin | u | − | SPIN1 Smad3 | PI3K/Akt modulation, EMT | MCF-7 | [175] [176] | |
| miR-100 | Paclitaxel | u | − | mTOR | Apoptosis, cell cycle control | MCF-7, MDA-MB-231 | [117] |
| miR-663a | Doxorubicin | u | + | HSPG2 | Antiapoptotic response | MDA-MB-231 | [177] |
| miR-200c | Doxorubicin | u | − | TrkB, Bmi1 | Cell survival and EMT | BT474 | [178] |
| Carboplatin doxycycline | u | − | EZH2, Bmi1 | EMT | p53-null claudin-low tumor model | [133,134] | |
| miR-105 miR-93-3p | Cisplatin | u | + | SFRP1/Wnt/β-catenin | Stemness and EMT induction | HCC70, MDA-MB-231, BT-549, HCC1937 | [179] |
| miR-24 | Cisplatin | u | + | BimL, FIH1-HIF1α | Cell survival and EMT | T47D, MDA-MB-231, MCF-7, BT-549 | [180] |
| miR-20a | MDR | u | − | MAPK1/c-Myc/P-gp | Survival pathways, drug efflux | BCap37, Bads-200, Bats-72 | [181] |
| miR-424(322) miR-503 | Paclitaxel | d | + | BCL-2, IGF1R | Apoptosis/survival pathways | Mouse model | [121] |
| miR-638 | Cisplatin | u | - | BRCA1 | DNA damage repair | MCF-7, MDA-MB-231, T47D, Hs 578T | [114] |
| miR-519d | Cisplatin | u | − | MCL-1 | Apoptosis | T47D, T47D-CSCs | [129] |
| miR-181a | Doxorubicin | d | + | Bcl-2 | Apoptosis | MCF-7 | [120] |
| Mitoxantrone | u | − | ABCG2 (BCRP) | Drug efflux | MCF-7 | [182] | |
| Doxorubicin | u | + | Bax | Apoptosis | MDA-MB-231 | [183] | |
| miR-27b-3p | MDR | d | + | CBLB/GRB2 | PI3K/Akt and MAPK/Erk signaling pathways regulation | Bcap37, Bads-200, MCF-7, MDA-MB-231 | [59] |
| miR-520h | Paclitaxel | u | + | DAPK2 | Apoptosis evasion | [184] | |
| miR-18a | Paclitaxel | u | + | Dicer | Apoptosis evasion | MDA-MB-231 | [185] |
| miR-101 | Paclitaxel | u | + | MCL-1 | Apoptosis evasion | MDA-MB-435 | [186] |
| miR-345 miR-7 | Cisplatin | d d | + + | MRP-1 | Drug efflux | MCF-7 | [53] |
| miR-944 | Cisplatin | d | − | BNIP-3 | Apoptosis | MCF-7 | [123] |
| miR-199a-3p | Cisplatin | u | − | TFAM | Apoptosis | MDA-MB-231 | [187] |
| miR-218 | Cisplatin | u | − | BRCA1 | DNA damage repair | MCF-7 | [115] |
| miR-21 | Gemcitabine | u | + | PTEN/Akt pathway | EMT | MDA-MB-231 | [83] |
| miR-205 | MDR | u | − | VEGFA, FGF2 | Cell growth and angiogenesis | MCF-7, Cal51 | [27] |
| miR-137 | MDR | u | + | FSTL1 | Wnt/β-catenin related cellular stemness | MDA-MB-231 | [29] |
| miR-140 | MDR | u | − | Wnt/β-catenin signaling | Cellular stemness | MCF-7, MDA-MB-231 | [127] |
| miR-155 | Doxorubicinol | u | + | CD44, CD90, ABCG2 | Cellular stemness and drug efflux | MDA-MB-231 | [128] |
| ncRNA(s) | Drug | Status | Impact on Drug Resistance | Target(s) | Mechanism(s) | Experimental System | References |
|---|---|---|---|---|---|---|---|
| ROR | Tamoxifen | u | + | miR-205/ZEB1, ZEB2 | TGF-β-induced EMT | MCF-7, MDA-MB-231 | [202] |
| HOTAIR | Tamoxifen | u | + | ER signaling | Cell proliferation | MCF-7 | [205] |
| BCAR4 | Tamoxifen | u | + | ERBB2 and ERBB3 signaling | Cell survival and proliferation | ZR-75-1 | [207] |
| CCAT2 | Tamoxifen | u | + | Wnt/β-catenin pathway | Cell survival and viability | MCF-7, T47D | [209,211] |
| UCA1 | Tamoxifen | u | + | EZH2/p21 | Akt/mTOR signaling | MCF-7 T47D | [213,214] |
| u | + | Wnt/β-catenin pathway | Cell survival and EMT | MCF-7, T47D | [215] | ||
| u | + | miR-18a/HIF1α axis | Cell cycle regulation | BT474 | [216] | ||
| GAS5 | Tamoxifen | d | + | miR-222/PTEN | PI3K/Akt/mTOR signaling | MCF-7 | [218] |
| ncRNA(s) | Drug | Status | Impact on Drug Resistance | Target(s) | Mechanism(s) | Experimental System | References |
|---|---|---|---|---|---|---|---|
| AFAP-AS1 | Trastuzumab | u | + | AUF1/ERBB2 | Cell survival | SKBR3, BT474 | [23] |
| ATB | Trastuzumab | u | + | miR-200c/ZEB1, ZNF-217 | EMT | SKBR3, BC patient tissues | [56] |
| SNHG14 | Trastuzumab | u | + | Bcl2/Bax | Apoptosis | SKBR3, BT474 | [220] |
| H19 | Trastuzumab | u | + | NA | NA | BC patients | [221] |
| UCA1 | Trastuzumab | u | + | miR-18a/YAP1 | Cell viability | SKBR3 | [222] |
| TINCR | Trastuzumab | u | + | miR-125b/HER2, Snail | EMT | SKBR3, BT474 | [223] |
| HOTAIR | Trastuzumab | u | + | TGF-β, Snail, Vimentin, p-AKT, CyclinD1, E-cadherin, PTEN, P27 | Cell cycle and proliferation, EMT | SKBR3 | [224] |
| GAS5 | Trastuzumab, lapatinib | u | − | miR-21/PTEN | AKT/mTOR modulation | SKBR3 | [225] |
| ncRNA(s) | Drug | Status | Impact on Drug Resistance | Target (s) | Mechanism(s) | Experimental System | References |
|---|---|---|---|---|---|---|---|
| NEAT | Cisplatin taxol | u | + | CD44, CD24, SOX2 | Cellular stemness | MDA-MB-231 | [22] |
| H19 | Antracyclines | u | + | CUL4A-ABCB1/MDR1 | Drug efflux | MCF-7 | [58] |
| Paclitaxel | u | + | BIK, NOXA | Apoptosis | MCF-7, ZR-75-1 | [229] | |
| PANDA | Doxorubicin | u | + | NF-YA/APAF-1, BKI, FAS and LRDD | Apoptosis | Primary breast tumors | [226] |
| ARA | Doxorubicin | u | + | NA | MAPK and PPAR signaling, metabolic signaling pathways, cell cycle and focal adhesion | MCF-7 | [227] |
| NONHSAT141924 | Paclitaxel | u | + | p-CREB/Bcl2 | Apoptosis | MCF-7 | [228] |
| BORG | Doxorubicin | u | + | RPA-1 | Cell survival and DNA damage repair | SKBR3, BT474 | [230] |
| LINC00968 | Adriamycin | u | − | Wnt2 | Wnt/β-catenin survival pathway | MCF-7, KPL-4 | [231] |
| GAS5 | Adriamycin | d | + | miR-221/DKK2 and Wnt/β-catenin survival | Drug efflux | MCF-7 | [232] |
| MA-linc1 | Paclitaxel | u | + | Purα | Cell cycle regulation | U2OS | [233] |
| UCA1 | Paclitaxel | u | + | miR-613/CDK12 | Cell viability and apoptosis | MCF-7 | [234] |
| NONHSAT028712 NONHSAT057282 NONHSAG023333 | Doxorubicin | u u u | + + − | CDK2 ELF1, E2F1, SOCS3, BRAC2 | Cell cycle regulation and survival | MCF-7 | [235] |
| LINP1 | 5-FU doxorubicin | u u | + + | N/E-cadherin, vimentin Ku80 | EMT DNA Damage Repair | MDA-MB-231 | [236] [242] |
| ZEB1-AS1 | Cisplatin | u | + | miR-129-5p/ZEB1 | EMT | MCF-7 | [237] |
| ROR | 5-FU paclitaxel | u | + | N/E-cadherin, vimentin | EMT | BC tissues and lines | [238] |
| MDR | u | + | ABCB1 | Drug efflux | MDA-MB-231, Sum159PT | [239] | |
| Linc00152 | Doxorubicin | u | + | N-cadherin, E-cadherin, vimentin | EMT | MDA-MB-231, MCF-7 | [243] |
| AX747207 | Doxorubicin | d | + | RUNX3 | PI3K/Akt, Hippo and ErbB oncogenic pathways | MCF-7 | [244] |
| PRLB | 5-FU | u | + | miR-4766-5p/SIRT1 | Tumor growth and metastasis | MDA-MB-231/468 | [245] |
| Clinical Outcome | ncRNA(s) | Biological Samples | Association | Reference(s) |
|---|---|---|---|---|
| Survival | ↑miR-4653-3p | FFPE BC tissues | Increased DSF in ER-positive BC patients receiving TAM adjuvant therapy | [256] |
| ↑miR-30c-5p, ↑miR-↑30b-5p, ↓miR-182-5p, ↑miR-200b-3p | Fresh frozen BC tissues | Improved ERFS in endocrine therapy-treated BC patients | [257] | |
| ↑miR-204 | FFPE tissues | Improved DSF and OS in patients treated with chemotherapy; inverse correlation with TNM stage and metastasis | [258] | |
| ↑miR-375 | BC tissues and serum | Improved OS for stage II–III HER2-positive BC patients who underwent chemotherapy | [250] | |
| ↑miR-30c | BC tissues | Response to TAM and longer PFS in ER-positive BC patients | [248] | |
| ↑PINK1.AS, ↑KLF3.AS1, ↑LINC00339, ↑LINC00472, ↑RP11.351I21.11, ↑PKD1P6.NPIPP1, ↑PDCD4.AS1, ↑RP11.69E11.4, ↓RP11.259N19.1, ↓KB.1460A1.5, ↓PP14571 | BC tissues | Longer RSF in ER positive BC patients treated with tamoxifen | [204] | |
| Relapse | ↑miR-7 | BC cell lines and fresh frozen tissues | Poor PFS and post-relapse OS in TAM-treated BC patients | [249] |
| ↑miR-210 | BC cell tissues; BC cells an TEMs in coculture | Reduced DSF and OS in tamoxifen treated ER-positive BC patients | [259,260,261] | |
| ↑miR-21 | Frozen tissues | Poor patient outcome in trastuzumab-treated patients | [84] | |
| ↑miR-454 | FFPE BC tissues | Worse DFS in anthracycline-treated TNBC patients | [262] | |
| ↑miR-221/222 | FFPE and frozen BC tissues, cell lines | Poor prognosis related to lymph node metastasis in BC patients undergoing chemotherapy | [251,263] | |
| ↑miR-95-3p | FFPE BC tissues | Decreased OS and RFS in patients treated with anthracycline-based chemotherapy | [264] | |
| ↑miR-222, ↑miR-29a, ↑miR-140, ↑miR-574, ↑miR-6780b, ↑miR-7107, ↑miR-744 | FFPE BC tissues | Associated with decreased PFS and OS in chemotherapy-treated BC patients | [263] | |
| ↑miR-125, ↑miR-21 | Serum | Decreased DSF in patients receiving neoadjuvant chemotherapy | [265] | |
| ↑miR-122 | BC tissues and serum | Predicts metastatic recurrence in stage II–III HER2-positive BC patients who underwent chemotherapy | [250] | |
| ↑miR-155 | BC tissues | Associated with tumor grade and lymph node metastasis in paclitaxel-resistant BC patients | [252] | |
| ↑HOTAIR | BC tissues | Metastatic disease and poor OS | [20] | |
| ↑HIF1A-AS2, ↑AK124454 | BC tissues | Poor outcome in paclitaxel-resistant TNBC patients | [255] | |
| ↑FAM224A, ↑LINC00987, ↑MCM3AP-AS1, ↑RP11–351I21.11, ↑SNHG17, ↓CTA-228A9.4, ↓EGOT, ↓ HAND2-AS1 | BC tissues | Poor OS in ER-positive patients receiving endocrine therapy | [254] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dobre, E.-G.; Dinescu, S.; Costache, M. Connecting the Missing Dots: ncRNAs as Critical Regulators of Therapeutic Susceptibility in Breast Cancer. Cancers 2020, 12, 2698. https://doi.org/10.3390/cancers12092698
Dobre E-G, Dinescu S, Costache M. Connecting the Missing Dots: ncRNAs as Critical Regulators of Therapeutic Susceptibility in Breast Cancer. Cancers. 2020; 12(9):2698. https://doi.org/10.3390/cancers12092698
Chicago/Turabian StyleDobre, Elena-Georgiana, Sorina Dinescu, and Marieta Costache. 2020. "Connecting the Missing Dots: ncRNAs as Critical Regulators of Therapeutic Susceptibility in Breast Cancer" Cancers 12, no. 9: 2698. https://doi.org/10.3390/cancers12092698
APA StyleDobre, E.-G., Dinescu, S., & Costache, M. (2020). Connecting the Missing Dots: ncRNAs as Critical Regulators of Therapeutic Susceptibility in Breast Cancer. Cancers, 12(9), 2698. https://doi.org/10.3390/cancers12092698