
Molecular and Cellular Mechanisms of Metformin in Cervical Cancer



Abstract
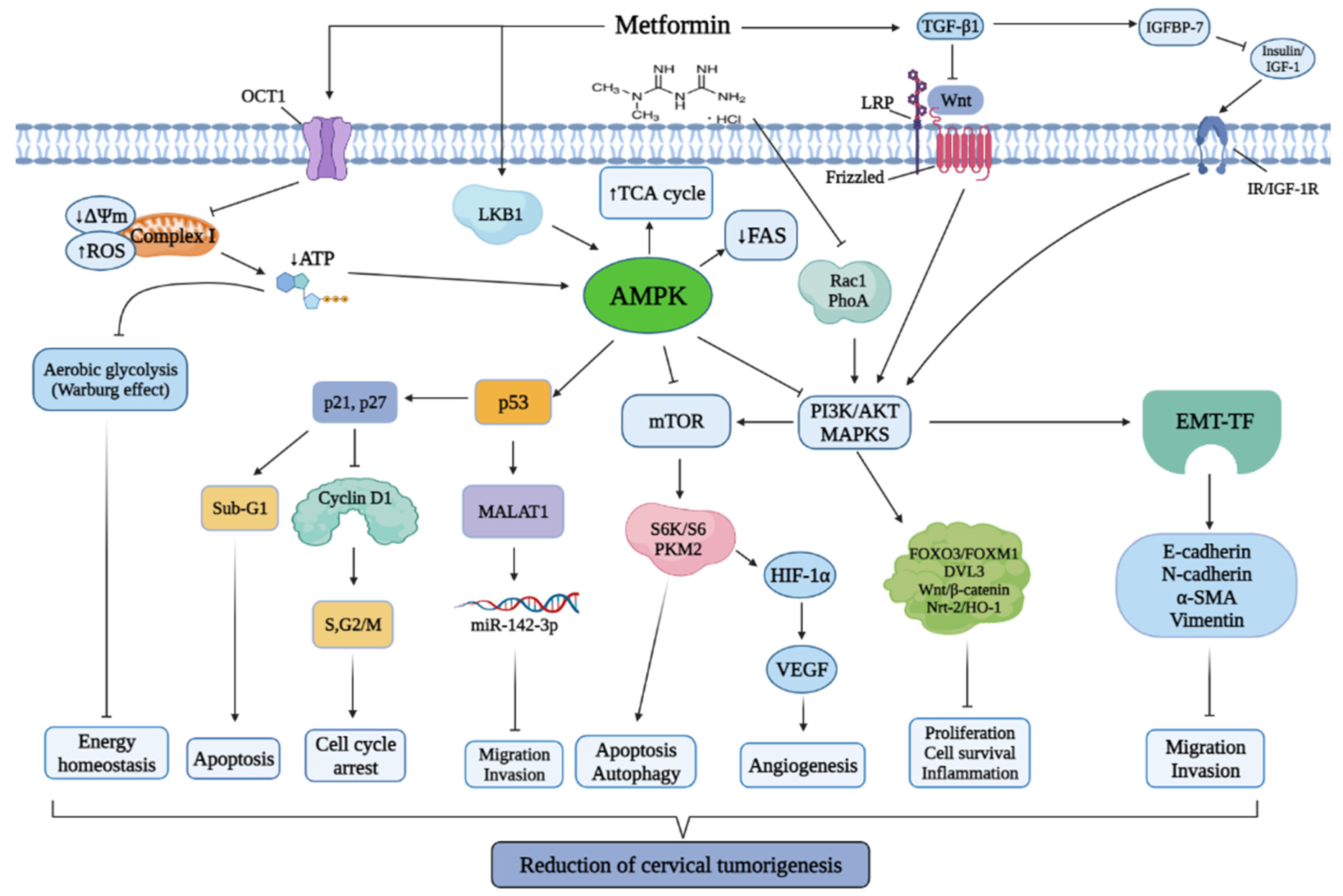
:Simple Summary
Abstract
1. Introduction
2. Mechanism of Metformin in Cervical Cancer
Effects of Metformin on Cervical Cancer Cells
3. Combination of Metformin with Therapeutic Agents Induced Anti-Cervical Cancer Effects
3.1. Metformin Combined with Carboplatin
3.2. Metformin Combined with Caffeic Acid
3.3. Metformin Combined with Caffeic Acid and Cisplatin
3.4. Metformin Combined with Nelfinavir
3.5. Clinical Trials of Metformin in Cervical Cancer
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AMPK | AMP-activated protein kinase |
| CaMKKβ | Ca2+ /calmodulin-dependent protein kinase kinase β |
| EMT | epithelial-mesenchymal transition |
| HIF-1α | hypoxia-inducible factor-1α |
| HO-1 | heme oxygenase-1 |
| IGF-1 | insulin-like growth factor 1 |
| IGFBP7 | Insulin Like Growth Factor Binding Protein 7 |
| LKB1 | liver kinase B1 |
| MALAT1 | metastasis associated lung adenocarcinoma transcript 1 |
| MAPK | mitogen-activated protein kinase |
| OCT-1 | organic cation transporter-1 |
| ROS | reactive oxygen species |
| TGF-β1 | transforming growth factor beta 1 |
| TORC2 | rapamycin complex 2 |
| VEGF | vascular endothelial growth factor |
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: Globocan estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naga, C.P.; Gurram, L.; Chopra, S.; Mahantshetty, U. The management of locally advanced cervical cancer. Curr. Opin. Oncol. 2018, 30, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Rena, G.; Hardie, D.G.; Pearson, E.R. The mechanisms of action of metformin. Diabetologia 2017, 60, 1577–1585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flory, J.; Lipska, K. Metformin in 2019. JAMA 2019, 321, 1926–1927. [Google Scholar] [CrossRef]
- Crowley, M.J.; Diamantidis, C.J.; McDuffie, J.R.; Cameron, C.B.; Stanifer, J.W.; Mock, C.K.; Wang, X.; Tang, S.; Nagi, A.; Kosinski, A.S.; et al. Clinical outcomes of metformin use in populations with chronic kidney disease, congestive heart failure, or chronic liver disease: A systematic review. Ann. Intern Med. 2017, 166, 191–200. [Google Scholar] [CrossRef]
- Suissa, S.; Azoulay, L. Metformin and cancer: Mounting evidence against an association. Diabetes Care 2014, 37, 1786–1788. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.; Li, Y.; Guo, Z.; Zeng, Y.; Zhang, W.; Wang, H. Metformin: Current clinical applications in nondiabetic patients with cancer. Aging 2020, 12, 3993–4009. [Google Scholar] [CrossRef] [PubMed]
- Takiuchi, T.; Machida, H.; Hom, M.S.; Mostofizadeh, S.; Frimer, M.; Brunette, L.L.; Matsuo, K. Association of metformin use and survival outcome in women with cervical cancer. Int. J. Gynecol. Cancer 2017, 27, 1455–1463. [Google Scholar] [CrossRef]
- Deng, D.; Yang, Y.; Tang, X.; Skrip, L.; Qiu, J.; Wang, Y.; Zhang, F. Association between metformin therapy and incidence, recurrence and mortality of prostate cancer: Evidence from a meta-analysis. Diabetes Metab. Res. Rev. 2015, 31, 595–602. [Google Scholar] [CrossRef]
- Hwang, I.C.; Park, S.M.; Shin, D.; Ahn, H.Y.; Rieken, M.; Shariat, S.F. Metformin association with lower prostate cancer recurrence in type 2 diabetes: A systematic review and meta-analysis. Asian Pac. J. Cancer Prev. 2015, 16, 595–600. [Google Scholar] [CrossRef] [Green Version]
- Ahn, H.K.; Lee, Y.H.; Koo, K.C. Current status and application of metformin for prostate cancer: A comprehensive review. Int. J. Mol. Sci. 2020, 21, 8540. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, G.; Tong, D.; Parmar, H.; Hasenmayer, D.; Yuan, W.; Zhang, D.; Jiang, J. Metformin represses androgen-dependent and androgen-independent prostate cancers by targeting androgen receptor. Prostate 2015, 75, 1187–1196. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Zheng, Z.; Li, X.; Li, B.; Lai, X.; Li, N.; Lei, S. Metformin attenuates hypoxia-induced endothelial cell injury by activating the ampk pathway. J. Cardiovasc. Pharmacol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.C.; Chiang, J.H.; Tsai, F.J.; Hsu, Y.M.; Juan, Y.N.; Yang, J.S.; Chiu, H.Y. Metformin triggers the intrinsic apoptotic response in human ags gastric adenocarcinoma cells by activating ampk and suppressing mtor/akt signaling. Int. J. Oncol. 2019, 54, 1271–1281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saraei, P.; Asadi, I.; Kakar, M.A.; Moradi-Kor, N. The beneficial effects of metformin on cancer prevention and therapy: A comprehensive review of recent advances. Cancer Manag. Res. 2019, 11, 3295–3313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noto, H.; Goto, A.; Tsujimoto, T.; Osame, K.; Noda, M. Latest insights into the risk of cancer in diabetes. J. Diabetes Investig. 2013, 4, 225–232. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Tao, M.; Zhao, L.; Zhang, X. The association between diabetes/hyperglycemia and the prognosis of cervical cancer patients: A systematic review and meta-analysis. Medicine 2017, 96, e7981. [Google Scholar] [CrossRef]
- Hanprasertpong, J.; Jiamset, I.; Geater, A.; Peerawong, T.; Hemman, W.; Kornsilp, S. The effect of metformin on oncological outcomes in patients with cervical cancer with type 2 diabetes mellitus. Int. J. Gynecol. Cancer 2017, 27, 131–137. [Google Scholar] [CrossRef]
- Anastasi, E.; Filardi, T.; Tartaglione, S.; Lenzi, A.; Angeloni, A.; Morano, S. Linking type 2 diabetes and gynecological cancer: An introductory overview. Clin. Chem. Lab. Med. 2018, 56, 1413–1425. [Google Scholar] [CrossRef]
- Evans, J.M.; Donnelly, L.A.; Emslie-Smith, A.M.; Alessi, D.R.; Morris, A.D. Metformin and reduced risk of cancer in diabetic patients. BMJ 2005, 330, 1304–1305. [Google Scholar] [CrossRef] [Green Version]
- Thakkar, B.; Aronis, K.N.; Vamvini, M.T.; Shields, K.; Mantzoros, C.S. Metformin and sulfonylureas in relation to cancer risk in type ii diabetes patients: A meta-analysis using primary data of published studies. Metabolism 2013, 62, 922–934. [Google Scholar] [CrossRef] [PubMed]
- Han, K.; Pintilie, M.; Lipscombe, L.L.; Lega, I.C.; Milosevic, M.F.; Fyles, A.W. Association between metformin use and mortality after cervical cancer in older women with diabetes. Cancer Epidemiol. Biomark. Prev. 2016, 25, 507–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, Q.; Zhao, Z.; Wen, J.; Zhou, J.; Wu, J.; Lei, S.; Miao, Y. The association between metformin therapy and risk of gynecological cancer in patients: Two meta-analyses. Eur. J. Obstet. Gynecol. Reprod. Biol. 2019, 237, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Mannucci, E.; Monami, M.; Balzi, D.; Cresci, B.; Pala, L.; Melani, C.; Lamanna, C.; Bracali, I.; Bigiarini, M.; Barchielli, A.; et al. Doses of insulin and its analogues and cancer occurrence in insulin-treated type 2 diabetic patients. Diabetes Care 2010, 33, 1997–2003. [Google Scholar] [CrossRef] [Green Version]
- Weinstein, D.; Simon, M.; Yehezkel, E.; Laron, Z.; Werner, H. Insulin analogues display igf-i-like mitogenic and anti-apoptotic activities in cultured cancer cells. Diabetes Metab. Res. Rev. 2009, 25, 41–49. [Google Scholar] [CrossRef]
- Stocks, T.; Rapp, K.; Bjørge, T.; Manjer, J.; Ulmer, H.; Selmer, R.; Lukanova, A.; Johansen, D.; Concin, H.; Tretli, S.; et al. Blood glucose and risk of incident and fatal cancer in the metabolic syndrome and cancer project (me-can): Analysis of six prospective cohorts. PLoS Med. 2009, 6, e1000201. [Google Scholar] [CrossRef]
- Barclay, A.W.; Petocz, P.; McMillan-Price, J.; Flood, V.M.; Prvan, T.; Mitchell, P.; Brand-Miller, J.C. Glycemic index, glycemic load, and chronic disease risk--a meta-analysis of observational studies. Am. J. Clin. Nutr. 2008, 87, 627–637. [Google Scholar] [CrossRef] [PubMed]
- Imai, A.; Ichigo, S.; Matsunami, K.; Takagi, H.; Yasuda, K. Clinical benefits of metformin in gynecologic oncology. Oncol. Lett. 2015, 10, 577–582. [Google Scholar] [CrossRef] [Green Version]
- Rehman, G.; Shehzad, A.; Khan, A.L.; Hamayun, M. Role of amp-activated protein kinase in cancer therapy. Arch. Pharm. 2014, 347, 457–468. [Google Scholar] [CrossRef]
- Kheirandish, M.; Mahboobi, H.; Yazdanparast, M.; Kamal, W.; Kamal, M.A. Anti-cancer effects of metformin: Recent evidences for its role in prevention and treatment of cancer. Curr. Drug Metab. 2018, 19, 793–797. [Google Scholar] [CrossRef]
- Dowling, R.J.; Niraula, S.; Stambolic, V.; Goodwin, P.J. Metformin in cancer: Translational challenges. J. Mol. Endocrinol. 2012, 48, R31–R43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Podhorecka, M.; Ibanez, B.; Dmoszyńska, A. Metformin-its potential anti-cancer and anti-aging effects. Postepy Hig. Med. Dosw. 2017, 71, 170–175. [Google Scholar] [CrossRef] [PubMed]
- Sui, X.; Xu, Y.; Wang, X.; Han, W.; Pan, H.; Xiao, M. Metformin: A novel but controversial drug in cancer prevention and treatment. Mol. Pharm. 2015, 12, 3783–3791. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.Y.; Xiao, D.; Wang, L.; Dong, L.H.; Yan, Z.X.; Shen, Z.X.; Chen, S.J.; Chen, Y.; Zhao, W.L. Therapeutic metformin/ampk activation blocked lymphoma cell growth via inhibition of mtor pathway and induction of autophagy. Cell Death Dis. 2012, 3, e275. [Google Scholar] [CrossRef] [PubMed]
- Vallianou, N.G.; Evangelopoulos, A.; Kazazis, C. Metformin and cancer. Rev. Diabet. Stud. 2013, 10, 228–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.H.; Yang, S.F.; Yang, C.K.; Tsai, H.D.; Chen, T.H.; Chou, M.C.; Hsiao, Y.H. Metformin induces apoptosis and inhibits migration by activating the ampk/p53 axis and suppressing pi3k/akt signaling in human cervical cancer cells. Mol. Med. Rep. 2021, 23, 88. [Google Scholar] [CrossRef]
- Xia, C.; Liu, C.; He, Z.; Cai, Y.; Chen, J. Metformin inhibits cervical cancer cell proliferation by modulating pi3k/akt-induced major histocompatibility complex class i-related chain a gene expression. J. Exp. Clin. Cancer Res. 2020, 39, 127. [Google Scholar] [CrossRef]
- Xiao, X.; He, Q.; Lu, C.; Werle, K.D.; Zhao, R.X.; Chen, J.; Davis, B.C.; Cui, R.; Liang, J.; Xu, Z.X. Metformin impairs the growth of liver kinase b1-intact cervical cancer cells. Gynecol. Oncol. 2012, 127, 249–255. [Google Scholar] [CrossRef] [Green Version]
- Hakimee, H.; Hutamekalin, P.; Tanasawet, S.; Chonpathompikunlert, P.; Tipmanee, V.; Sukketsiri, W. Metformin inhibit cervical cancer migration by suppressing the fak/akt signaling pathway. Asian Pac. J. Cancer Prev. 2019, 20, 3539–3545. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.O.; Kang, M.J.; Byun, W.S.; Kim, S.A.; Seo, I.H.; Han, J.A.; Moon, J.W.; Kim, J.H.; Kim, S.J.; Lee, E.J.; et al. Metformin overcomes resistance to cisplatin in triple-negative breast cancer (tnbc) cells by targeting rad51. Breast Cancer Res. 2019, 21, 115. [Google Scholar] [CrossRef] [Green Version]
- Xia, C.; Liang, S.; He, Z.; Zhu, X.; Chen, R.; Chen, J. Metformin, a first-line drug for type 2 diabetes mellitus, disrupts the malat1/mir-142-3p sponge to decrease invasion and migration in cervical cancer cells. Eur. J. Pharmacol. 2018, 830, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Yung, M.M.; Chan, D.W.; Liu, V.W.; Yao, K.M.; Ngan, H.Y. Activation of ampk inhibits cervical cancer cell growth through akt/foxo3a/foxm1 signaling cascade. BMC Cancer 2013, 13, 327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwan, H.T.; Chan, D.W.; Cai, P.C.; Mak, C.S.; Yung, M.M.; Leung, T.H.; Wong, O.G.; Cheung, A.N.; Ngan, H.Y. Ampk activators suppress cervical cancer cell growth through inhibition of dvl3 mediated wnt/β-catenin signaling activity. PLoS ONE 2013, 8, e53597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, K.; Hao, M. Metformin inhibits tgf-β1-induced epithelial-to-mesenchymal transition via pkm2 relative-mtor/p70s6k signaling pathway in cervical carcinoma cells. Int. J. Mol. Sci. 2016, 17, 2000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Do, M.T.; Kim, H.G.; Khanal, T.; Choi, J.H.; Kim, D.H.; Jeong, T.C.; Jeong, H.G. Metformin inhibits heme oxygenase-1 expression in cancer cells through inactivation of raf-erk-nrf2 signaling and ampk-independent pathways. Toxicol. Appl. Pharmacol. 2013, 271, 229–238. [Google Scholar] [CrossRef]
- Xia, C.; Yang, F.; He, Z.; Cai, Y. Itraq-based quantitative proteomic analysis of the inhibition of cervical cancer cell invasion and migration by metformin. Biomed. Pharmacother. 2020, 123, 109762. [Google Scholar] [CrossRef]
- Yudhani, R.D.; Astuti, I.; Mustofa, M.; Indarto, D.; Muthmainah, M. Metformin modulates cyclin d1 and p53 expression to inhibit cell proliferation and to induce apoptosis in cervical cancer cell lines. Asian Pac. J. Cancer Prev. 2019, 20, 1667–1673. [Google Scholar] [CrossRef] [Green Version]
- Yu, S.Y.; Chan, D.W.; Liu, V.W.; Ngan, H.Y. Inhibition of cervical cancer cell growth through activation of upstream kinases of amp-activated protein kinase. Tumour Biol. 2009, 30, 80–85. [Google Scholar] [CrossRef]
- Lin, C.C.; Yeh, H.H.; Huang, W.L.; Yan, J.J.; Lai, W.W.; Su, W.P.; Chen, H.H.; Su, W.C. Metformin enhances cisplatin cytotoxicity by suppressing signal transducer and activator of transcription-3 activity independently of the liver kinase b1-amp-activated protein kinase pathway. Am. J. Respir. Cell Mol. Biol. 2013, 49, 241–250. [Google Scholar] [CrossRef]
- Rocha, G.Z.; Dias, M.M.; Ropelle, E.R.; Osório-Costa, F.; Rossato, F.A.; Vercesi, A.E.; Saad, M.J.; Carvalheira, J.B. Metformin amplifies chemotherapy-induced ampk activation and antitumoral growth. Clin. Cancer Res. 2011, 17, 3993–4005. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, L.S.M.; Liu, S.T.; Chang, Y.L.; Ho, C.L.; Huang, S.M. Metformin causes cancer cell death through downregulation of p53-dependent differentiated embryo chondrocyte 1. J. Biomed. Sci. 2018, 25, 81. [Google Scholar] [CrossRef] [Green Version]
- Irie, H.; Banno, K.; Yanokura, M.; Iida, M.; Adachi, M.; Nakamura, K.; Umene, K.; Nogami, Y.; Masuda, K.; Kobayashi, Y.; et al. Metformin: A candidate for the treatment of gynecological tumors based on drug repositioning. Oncol. Lett. 2016, 11, 1287–1293. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.Y.; Kim, Y.S.; Kim, M.; Choi, M.Y.; Roh, G.S.; Lee, D.H.; Kim, H.J.; Kang, S.S.; Cho, G.J.; Shin, J.K.; et al. Metformin inhibits cervical cancer cell proliferation via decreased ampk o-glcnacylation. Anim. Cells Syst. 2019, 23, 302–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, Q.; Zhang, L.; Liu, X.; Zhou, L.; Wang, W.; Han, Z.; Sui, H.; Tang, Y.; Wang, Y.; Liu, N.; et al. Long non-coding rna malat1 promotes tumour growth and metastasis in colorectal cancer through binding to sfpq and releasing oncogene ptbp2 from sfpq/ptbp2 complex. Br. J. Cancer 2014, 111, 736–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, S.; Wang, F.; Shen, J.; Sun, Y.; Xu, W.; Lu, J.; Wei, M.; Xu, C.; Wu, C.; Zhang, Z.; et al. Long non-coding rna metastasis associated in lung adenocarcinoma transcript 1 derived minirna as a novel plasma-based biomarker for diagnosing prostate cancer. Eur. J. Cancer 2013, 49, 2949–2959. [Google Scholar] [CrossRef] [PubMed]
- Deng, B.; Zhang, Y.; Zhang, S.; Wen, F.; Miao, Y.; Guo, K. Microrna-142-3p inhibits cell proliferation and invasion of cervical cancer cells by targeting fzd7. Tumour Biol. 2015, 36, 8065–8073. [Google Scholar] [CrossRef]
- Li, M.; Li, B.Y.; Xia, H.; Jiang, L.L. Expression of microrna-142-3p in cervical cancer and its correlation with prognosis. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 2346–2350. [Google Scholar] [PubMed]
- Wierstra, I.; Alves, J. Foxm1, a typical proliferation-associated transcription factor. Biol. Chem. 2007, 388, 1257–1274. [Google Scholar] [CrossRef] [PubMed]
- Thiery, J.P.; Acloque, H.; Huang, R.Y.; Nieto, M.A. Epithelial-mesenchymal transitions in development and disease. Cell 2009, 139, 871–890. [Google Scholar] [CrossRef] [PubMed]
- Thomson, S.; Buck, E.; Petti, F.; Griffin, G.; Brown, E.; Ramnarine, N.; Iwata, K.K.; Gibson, N.; Haley, J.D. Epithelial to mesenchymal transition is a determinant of sensitivity of non-small-cell lung carcinoma cell lines and xenografts to epidermal growth factor receptor inhibition. Cancer Res. 2005, 65, 9455–9462. [Google Scholar] [CrossRef] [Green Version]
- Akagi, R.; Takahashi, T.; Sassa, S. Cytoprotective effects of heme oxygenase in acute renal failure. Contrib. Nephrol. 2005, 148, 70–85. [Google Scholar] [PubMed]
- Wagener, F.A.; Eggert, A.; Boerman, O.C.; Oyen, W.J.; Verhofstad, A.; Abraham, N.G.; Adema, G.; van Kooyk, Y.; de Witte, T.; Figdor, C.G. Heme is a potent inducer of inflammation in mice and is counteracted by heme oxygenase. Blood 2001, 98, 1802–1811. [Google Scholar] [CrossRef]
- Sass, G.; Leukel, P.; Schmitz, V.; Raskopf, E.; Ocker, M.; Neureiter, D.; Meissnitzer, M.; Tasika, E.; Tannapfel, A.; Tiegs, G. Inhibition of heme oxygenase 1 expression by small interfering rna decreases orthotopic tumor growth in livers of mice. Int. J. Cancer 2008, 123, 1269–1277. [Google Scholar] [CrossRef] [PubMed]
- Miyake, M.; Fujimoto, K.; Anai, S.; Ohnishi, S.; Kuwada, M.; Nakai, Y.; Inoue, T.; Matsumura, Y.; Tomioka, A.; Ikeda, T.; et al. Heme oxygenase-1 promotes angiogenesis in urothelial carcinoma of the urinary bladder. Oncol. Rep. 2011, 25, 653–660. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.M.; Chen, G.G.; Ng, E.K.; Leung, W.K.; Sung, J.J.; Chung, S.C. Upregulation of heme oxygenase-1 and p21 confers resistance to apoptosis in human gastric cancer cells. Oncogene 2004, 23, 503–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alaoui-Jamali, M.A.; Bismar, T.A.; Gupta, A.; Szarek, W.A.; Su, J.; Song, W.; Xu, Y.; Xu, B.; Liu, G.; Vlahakis, J.Z.; et al. A novel experimental heme oxygenase-1-targeted therapy for hormone-refractory prostate cancer. Cancer Res. 2009, 69, 8017–8024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berberat, P.O.; Dambrauskas, Z.; Gulbinas, A.; Giese, T.; Giese, N.; Künzli, B.; Autschbach, F.; Meuer, S.; Büchler, M.W.; Friess, H. Inhibition of heme oxygenase-1 increases responsiveness of pancreatic cancer cells to anticancer treatment. Clin. Cancer Res. 2005, 11, 3790–3798. [Google Scholar] [CrossRef] [Green Version]
- Hemminki, A.; Markie, D.; Tomlinson, I.; Avizienyte, E.; Roth, S.; Loukola, A.; Bignell, G.; Warren, W.; Aminoff, M.; Höglund, P.; et al. A serine/threonine kinase gene defective in peutz-jeghers syndrome. Nature 1998, 391, 184–187. [Google Scholar] [CrossRef]
- Herrmann, J.L.; Byekova, Y.; Elmets, C.A.; Athar, M. Liver kinase b1 (lkb1) in the pathogenesis of epithelial cancers. Cancer Lett. 2011, 306, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Zhu, H.; Dai, X.; Wang, C.; Ding, Y.; Song, P.; Zou, M.H. Macrophage liver kinase b1 inhibits foam cell formation and atherosclerosis. Circ. Res. 2017, 121, 1047–1057. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, L.; Yang, Y.; Li, Y. Liver kinase b1 (lkb1) regulates proliferation and apoptosis of non-small cell lung cancer a549 cells via targeting erk signaling pathway. Cancer Manag. Res. 2021, 13, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Guo, K.Y.; Han, L.; Li, X.; Yang, A.V.; Lu, J.; Guan, S.; Li, H.; Yu, Y.; Zhao, Y.; Yang, J.; et al. Novel proteasome inhibitor delanzomib sensitizes cervical cancer cells to doxorubicin-induced apoptosis via stabilizing tumor suppressor proteins in the p53 pathway. Oncotarget 2017, 8, 114123–114135. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Yang, X.; Chen, F.; Chen, D. Combined application of doxorubicin and naringin enhances the antitumor efficiency and attenuates the toxicity of doxorubicin in hela cervical cancer cells. Int. J. Clin. Exp. Pathol. 2017, 10, 7303–7311. [Google Scholar]
- Yuniarti, L.; Mustofa, M.; Aryandono, T.; Haryana, S.M. Synergistic action of 1,2-epoxy-3 (3- (3,4-dimethoxyphenyl)- 4h-1-benzopiyran-4-on) propane with doxorubicin and cisplatin through increasing of p53, timp-3, and microrna-34a in cervical cancer cell line (hela). Asian Pac. J. Cancer Prev. 2018, 19, 2955–2962. [Google Scholar] [PubMed]
- Marini, C.; Salani, B.; Massollo, M.; Amaro, A.; Esposito, A.I.; Orengo, A.M.; Capitanio, S.; Emionite, L.; Riondato, M.; Bottoni, G.; et al. Direct inhibition of hexokinase activity by metformin at least partially impairs glucose metabolism and tumor growth in experimental breast cancer. Cell Cycle 2013, 12, 3490–3499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrzejewski, S.; Gravel, S.P.; Pollak, M.; St-Pierre, J. Metformin directly acts on mitochondria to alter cellular bioenergetics. Cancer Metab. 2014, 2, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marini, C.; Ravera, S.; Buschiazzo, A.; Bianchi, G.; Orengo, A.M.; Bruno, S.; Bottoni, G.; Emionite, L.; Pastorino, F.; Monteverde, E.; et al. Discovery of a novel glucose metabolism in cancer: The role of endoplasmic reticulum beyond glycolysis and pentose phosphate shunt. Sci. Rep. 2016, 6, 25092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, J.; Wang, K.; Zheng, N.; Qiu, Y.; Xie, G.; Su, M.; Jia, W.; Li, H. Metformin suppressed the proliferation of lovo cells and induced a time-dependent metabolic and transcriptional alteration. Sci. Rep. 2015, 5, 17423. [Google Scholar] [CrossRef] [Green Version]
- Tang, Z.Y.; Sheng, M.J.; Qi, Y.X.; Wang, L.Y.; He, D.Y. Metformin enhances inhibitive effects of carboplatin on hela cell proliferation and increases sensitivity to carboplatin by activating mitochondrial associated apoptosis signaling pathway. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 8104–8112. [Google Scholar]
- Tyszka-Czochara, M.; Konieczny, P.; Majka, M. Caffeic acid expands anti-tumor effect of metformin in human metastatic cervical carcinoma htb-34 cells: Implications of ampk activation and impairment of fatty acids de novo biosynthesis. Int. J. Mol. Sci. 2017, 18, 462. [Google Scholar] [CrossRef] [Green Version]
- Tyszka-Czochara, M.; Bukowska-Strakova, K.; Kocemba-Pilarczyk, K.A.; Majka, M. Caffeic acid targets ampk signaling and regulates tricarboxylic acid cycle anaplerosis while metformin downregulates hif-1α-induced glycolytic enzymes in human cervical squamous cell carcinoma lines. Nutrients 2018, 10, 841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyszka-Czochara, M.; Lasota, M.; Majka, M. Caffeic acid and metformin inhibit invasive phenotype induced by tgf-β1 in c-4i and htb-35/siha human cervical squamous carcinoma cells by acting on different molecular targets. Int. J. Mol. Sci. 2018, 19, 266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyszka-Czochara, M.; Bukowska-Strakova, K.; Majka, M. Metformin and caffeic acid regulate metabolic reprogramming in human cervical carcinoma siha/htb-35 cells and augment anticancer activity of cisplatin via cell cycle regulation. Food Chem. Toxicol. 2017, 106, 260–272. [Google Scholar] [CrossRef] [PubMed]
- Xia, C.; Chen, R.; Chen, J.; Qi, Q.; Pan, Y.; Du, L.; Xiao, G.; Jiang, S. Combining metformin and nelfinavir exhibits synergistic effects against the growth of human cervical cancer cells and xenograft in nude mice. Sci. Rep. 2017, 7, 43373. [Google Scholar] [CrossRef] [Green Version]
- Muggia, F.M.; Gill, I. Role of carboplatin in endometrial and cervical carcinomas. Semin. Oncol. 1992, 19, 90–93. [Google Scholar]
- Muderspach, L.I.; Curtin, J.P.; Roman, L.D.; Gebhardt, J.A.; Klement, V.; Qian, D.; Morrow, C.P.; Felix, J.C.; Formenti, S.C.; Muggia, F.M. Carboplatin as a radiation sensitizer in locally advanced cervical cancer: A pilot study. Gynecol. Oncol. 1997, 65, 336–342. [Google Scholar] [CrossRef]
- Macdonald, M.C.; Tidy, J.A. Can we be less radical with surgery for early cervical cancer? Curr. Oncol. Rep. 2016, 18, 16. [Google Scholar] [CrossRef] [PubMed]
- Markman, M. Chemoradiation in the management of cervix cancer: Current status and future directions. Oncology 2013, 84, 246–250. [Google Scholar] [CrossRef]
- Yang, L.; Douple, E.B.; O’Hara, J.A.; Crabtree, R.A.; Eastman, A. Enhanced radiation-induced cell killing by carboplatin in cells of repair-proficient and repair-deficient cell lines. Radiat. Res. 1995, 144, 230–236. [Google Scholar] [CrossRef] [PubMed]
- Muggia, F.M. Overview of carboplatin: Replacing, complementing, and extending the therapeutic horizons of cisplatin. Semin. Oncol. 1989, 16, 7–13. [Google Scholar]
- Perez, R.P. Cellular and molecular determinants of cisplatin resistance. Eur. J. Cancer 1998, 34, 1535–1542. [Google Scholar] [CrossRef]
- Yuwen, D.L.; Sheng, B.B.; Liu, J.; Wenyu, W.; Shu, Y.Q. Mir-146a-5p level in serum exosomes predicts therapeutic effect of cisplatin in non-small cell lung cancer. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 2650–2658. [Google Scholar] [PubMed]
- Dziedzic, A.; Kubina, R.; Kabała-Dzik, A.; Wojtyczka, R.D.; Morawiec, T.; Bułdak, R.J. Caffeic acid reduces the viability and migration rate of oral carcinoma cells (scc-25) exposed to low concentrations of ethanol. Int. J. Mol. Sci. 2014, 15, 18725–18741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, C.L.; Chen, R.F.; Chen, J.Y.; Chu, Y.C.; Wang, H.M.; Chou, H.L.; Chang, W.C.; Fong, Y.; Chang, W.T.; Wu, C.Y.; et al. Protective effect of caffeic acid on paclitaxel induced anti-proliferation and apoptosis of lung cancer cells involves nf-κb pathway. Int. J. Mol. Sci. 2012, 13, 6236–6245. [Google Scholar] [CrossRef] [Green Version]
- Murad, L.D.; Soares Nda, C.; Brand, C.; Monteiro, M.C.; Teodoro, A.J. Effects of caffeic and 5-caffeoylquinic acids on cell viability and cellular uptake in human colon adenocarcinoma cells. Nutr. Cancer 2015, 67, 532–542. [Google Scholar] [CrossRef]
- Zhu, H.; Luo, H.; Zhang, W.; Shen, Z.; Hu, X.; Zhu, X. Molecular mechanisms of cisplatin resistance in cervical cancer. Drug Des. Dev. Ther. 2016, 10, 1885–1895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, D.W.; Pouliot, L.M.; Hall, M.D.; Gottesman, M.M. Cisplatin resistance: A cellular self-defense mechanism resulting from multiple epigenetic and genetic changes. Pharmacol. Rev. 2012, 64, 706–721. [Google Scholar] [CrossRef] [Green Version]
- Lorusso, D.; Petrelli, F.; Coinu, A.; Raspagliesi, F.; Barni, S. A systematic review comparing cisplatin and carboplatin plus paclitaxel-based chemotherapy for recurrent or metastatic cervical cancer. Gynecol. Oncol. 2014, 133, 117–123. [Google Scholar] [CrossRef]
- Subeha, M.R.; Telleria, C.M. The anti-cancer properties of the hiv protease inhibitor nelfinavir. Cancers 2020, 12, 3437. [Google Scholar] [CrossRef]
- Yang, Y.; Ikezoe, T.; Nishioka, C.; Bandobashi, K.; Takeuchi, T.; Adachi, Y.; Kobayashi, M.; Takeuchi, S.; Koeffler, H.P.; Taguchi, H. Nfv, an hiv-1 protease inhibitor, induces growth arrest, reduced akt signalling, apoptosis and docetaxel sensitisation in nsclc cell lines. Br. J. Cancer 2006, 95, 1653–1662. [Google Scholar] [CrossRef] [Green Version]
- Jiang, W.; Mikochik, P.J.; Ra, J.H.; Lei, H.; Flaherty, K.T.; Winkler, J.D.; Spitz, F.R. Hiv protease inhibitor nelfinavir inhibits growth of human melanoma cells by induction of cell cycle arrest. Cancer Res. 2007, 67, 1221–1227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiang, T.; Du, L.; Pham, P.; Zhu, B.; Jiang, S. Nelfinavir, an hiv protease inhibitor, induces apoptosis and cell cycle arrest in human cervical cancer cells via the ros-dependent mitochondrial pathway. Cancer Lett. 2015, 364, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Chou, T.C. Theoretical basis, experimental design, and computerized simulation of synergism and antagonism in drug combination studies. Pharmacol. Rev. 2006, 58, 621–681. [Google Scholar] [CrossRef] [PubMed]
- Koltai, T. Nelfinavir and other protease inhibitors in cancer: Mechanisms involved in anticancer activity. F1000Res 2015, 4, 9. [Google Scholar] [CrossRef] [PubMed]
- Kalender, A.; Selvaraj, A.; Kim, S.Y.; Gulati, P.; Brûlé, S.; Viollet, B.; Kemp, B.E.; Bardeesy, N.; Dennis, P.; Schlager, J.J.; et al. Metformin, independent of ampk, inhibits mtorc1 in a rag gtpase-dependent manner. Cell Metab. 2010, 11, 390–401. [Google Scholar] [CrossRef] [Green Version]
- Banno, K.; Iida, M. Drug repositioning for gynecologic tumors: A new therapeutic strategy for cancer. Sci. World J. 2015, 2015, 341362. [Google Scholar] [CrossRef] [Green Version]
- Febbraro, T.; Lengyel, E.; Romero, I.L. Old drug, new trick: Repurposing metformin for gynecologic cancers? Gynecol. Oncol. 2014, 135, 614–621. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
| Cell Lines/Animal Models | Anti-Cancer Effects | Mechanisms | Reference |
|---|---|---|---|
| CaSki, C33A, HeLa | ↑ apoptosis ↓ migration | ↑ AMPK/p53 ↓ PI3K/AKT | Chen et al. [36] |
| SiHa, HeLa BALB/c nude mice | ↓ proliferation ↓ tumor growth in xenografts | ↑ p53, MICA and NK cell cytotoxicity ↓ PI3K/AKT | Xia et al. [37] |
| C33A, Me180, CaSki, HeLa, HT-3, MS751 | ↑apoptosis ↑autophagy | ↑ LKB1-AMPK-mTOR | Xiao et al. [38] |
| HeLa | ↓ migration | ↑ FAK/Akt ↓ Rac1 and PhoA protein expression | Hakimee et al. [39] |
| HeLa | ↓ proliferation ↑apoptosis ↑ sub-G1 arrest | ↓ AMPK O-GlcNAcylation ↑ p21 and p27 levels | Kim et al. [40] |
| SiHa, HeLa BALB/c nude mice | ↓ invasion and migration ↓ tumor growth in xenografts | ↓ MALAT1 expression ↑ miR-142-3p expression | Xia et al. [41] |
| HeLa, CaSki, C33A, SiHa | ↓ proliferation | ↑ AMPK ↓ AKT/FOXO3a/FOXM1 | Yung et al. [42] |
| HeLa, SiHa, C33A, CaSki, C41 | ↓ proliferation | ↓ DVL3 protein synthesis ↑ AMPK activation ↓ Wnt/β-catenin signaling | Kwan et al. [43] |
| HeLa, SiHa | ↑ apoptosis ↓ proliferation ↓ migration | ↓ TGF-β1-induced EMT effects ↓ mTOR/p70s6k/PKM2 | Cheng et al. [44] |
| HeLa | ↓ proliferation | ↓ HO-1 protein expression ↓ Raf-ERK-Nrf2 ↑ AMPK-independent mechanisms | Do et al. [45] |
| HeLa, SiHa BALB/c nude mice | ↓ proliferation ↓ invasion and migration ↓ tumor growth in xenografts | ↑ IGFBP7 protein expression Regulates the insulin signaling pathway | Xiao et al. [46] |
| HeLa | ↑ apoptosis ↓ proliferation ↑ S and G2/M arrests | ↓ Cyclin D1 expression ↑ p53 expression | Yudhani et al. [47] |
| Combined Therapeutic Agent | Cell Lines/Animal Models | Anti-Cancer Effects | Mechanisms | Reference |
|---|---|---|---|---|
| Carboplatin | HeLa | ↑ apoptosis ↓ proliferation | ↑ Nuclear fragments formation ↓ Mitochondrial membrane potential | Tang et al. [79] |
| Caffeic acid | HTB-34 | ↑ apoptosis ↓ proliferation ↑ S and G2/M arrests | ↑ AMPK ↑ mitochondrial ROS ↑ TCA cycle ↑ Fatty acids de novo synthesis | Tyszka-Czochara et al. [80] |
| Caffeic acid | C-4I, HTB-35/SiHa | ↑apoptosis ↓ proliferation ↓ energy homeostasis | ↓ c-Myc, BAX and cyclin-D1 expression ↓ HIF-1α and Warburg effect ↑ mitochondrial ROS ↑ AMPK regulating oxidative metabolism/glycolysis | Tyszka-Czochara et al. [81] |
| Caffeic acid | C-4I, HTB-35/SiHa | ↓ proliferation ↓ migration | ↑ Epithelial adhesive markers ↓ Mesenchymal transcription factors regulating EMT | Tyszka-Czochara et al. [82] |
| Caffeic acid | HTB-35/SiHa | ↑ apoptosis ↑ cisplatin anti-cancer action | ↑ AMPK ↑ mitochondrial ROS ↑ TCA cycle ↓ novo unsaturated fatty acid synthesis | Tyszka-Czochara et al. [83] |
| Nelfinavir | HeLa, SiHa, CaSki BALB/c nude mice | ↑ apoptosis ↓ proliferation ↓ invasion and migration ↓ tumor growth in xenografts | ↑ ROS, p53 and p21 expression ↓ PI3K (p110α) | Xia et al. [84] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.-H.; Wang, P.-H.; Chen, P.-N.; Yang, S.-F.; Hsiao, Y.-H. Molecular and Cellular Mechanisms of Metformin in Cervical Cancer. Cancers 2021, 13, 2545. https://doi.org/10.3390/cancers13112545
Chen Y-H, Wang P-H, Chen P-N, Yang S-F, Hsiao Y-H. Molecular and Cellular Mechanisms of Metformin in Cervical Cancer. Cancers. 2021; 13(11):2545. https://doi.org/10.3390/cancers13112545
Chicago/Turabian StyleChen, Ya-Hui, Po-Hui Wang, Pei-Ni Chen, Shun-Fa Yang, and Yi-Hsuan Hsiao. 2021. "Molecular and Cellular Mechanisms of Metformin in Cervical Cancer" Cancers 13, no. 11: 2545. https://doi.org/10.3390/cancers13112545