
Neuro-Signals from Gut Microbiota: Perspectives for Brain Glioma



 ,
,  ,
,
Abstract
Simple Summary
Abstract

1. Introduction
2. Gut Microbiota Influences the CNS: The Gut-Brain Axis
2.1. Gut Microbiota and Brain Disorders
2.2. Neuro-Signals from Gut Microbiota
2.3. Modulation of Neurotransmitter Levels by Gut Microbiota
3. Neurotransmitters beyond the Neuronal Function
3.1. Neurotransmitters Influence Physiological Cell Proliferation in Adult Brain
3.1.1. Dopamine
3.1.2. Serotonin
3.1.3. Norepinephrine
3.1.4. GABA
3.1.5. Glutamate

{kind=link}
{kind=link}
{kind=link}
| Neurotransmitters | Receptors | Effects | References |
|---|---|---|---|
| dopamine | n.d. | Increases NPC proliferation in SVZ | [115,116,117] |
| D2-like | Promotes entry in S phase | [120] | |
| D2-like | Decreases NPC proliferation | [118,119] | |
| D1-like | Inhibits the entry in S phase | [121] | |
| D3 | Inhibits the maturation of OPC | [122] | |
| serotonin | n.d. | Depletion decreases proliferation in DG and SVZ | [123] |
| 5-HT1A | Blockade of receptors decreases proliferation in DG | [124] | |
| n.d. | Increases NPC proliferation in DG | [125,126] | |
| n.d. | Blocks OPC maturation | [127] | |
| 5-HT | Increases levels of trophic factors | [128,129,130,131] | |
| norepinephrine | n.d. | Depletion decreases NPC proliferation but not differentiation in DG | [132] |
| β3-AR | Increases proliferation of NPC in DG | [133] | |
| a2-AR/β-AR | Balance in receptor activity regulates NPC activity in DG | [134] | |
| n.d. | Reduces NPC proliferation in SVZ | [135] | |
| gaba | extra synaptic GABA-A | Inhibits NPC proliferation in SVZ | [136] |
| extra synaptic GABA-A | Regulates NPC production and migration | [137,138,139] | |
| extra synaptic GABA-A | Maintains NPC quiescence in DG | [140] | |
| synaptic GABA-A | Promotes NPC maturation in DG | [140] | |
| glutamate | n.d. | Regulates adult neurogenesis | [114,142] |
| mGluR5 | Promotes Neurogenesis in DG | [142,149] | |
| NMDA | Promotes commitment and diferentiation in DG | [145,146,147,149] | |
| NR2B | Increases dendritic arborization and contributes LTP induced neurogenesis | [148] |
3.2. Neurotransmitters Influence Glioma
3.2.1. Monoamines
3.2.2. Dopamine
3.2.3. Serotonin
3.2.4. Norepinephrine
3.2.5. GABA
3.2.6. Glutamate
| Neurotransmitters | Receptors | Effects | References |
|---|---|---|---|
| dopamine | D2 | Regulates survival and cell proliferation | [155,156] |
| D2 | Increases cancer stem cell-enriched spheroid proliferation | [158,159,160] | |
| serotonin | 5-HT1, 5-HT2 | Increases cell proliferation | [172,173] |
| 5-HT2A | Increases cell proliferation and migration | [174] | |
| n.d. | Induces cell differentiation | [175] | |
| 5-HT2A | Increases the release of neurotrophic factor GDNF | [176] | |
| norepinephrine | β | Increases proliferation | [180] |
| β2 | Inhibits proliferation | [181] | |
| n.d. | Inhibits migration and invasion | [182] | |
| gaba | GABA-A | Reduces cell proliferation sustaining cell quiescence | [186] |
| glutamate | mGluR2/3 | Regulates cell growth | [193,194] |
| GluR1/GluR4 | Increase cell proliferation and migration | [195] | |
| AMPA | Promotes perivascular invasion | [196] | |
| NMDA | Increases proliferation | [197] | |
| n.d. | Drives tumor growth and invasion | [198,199,200,201] |
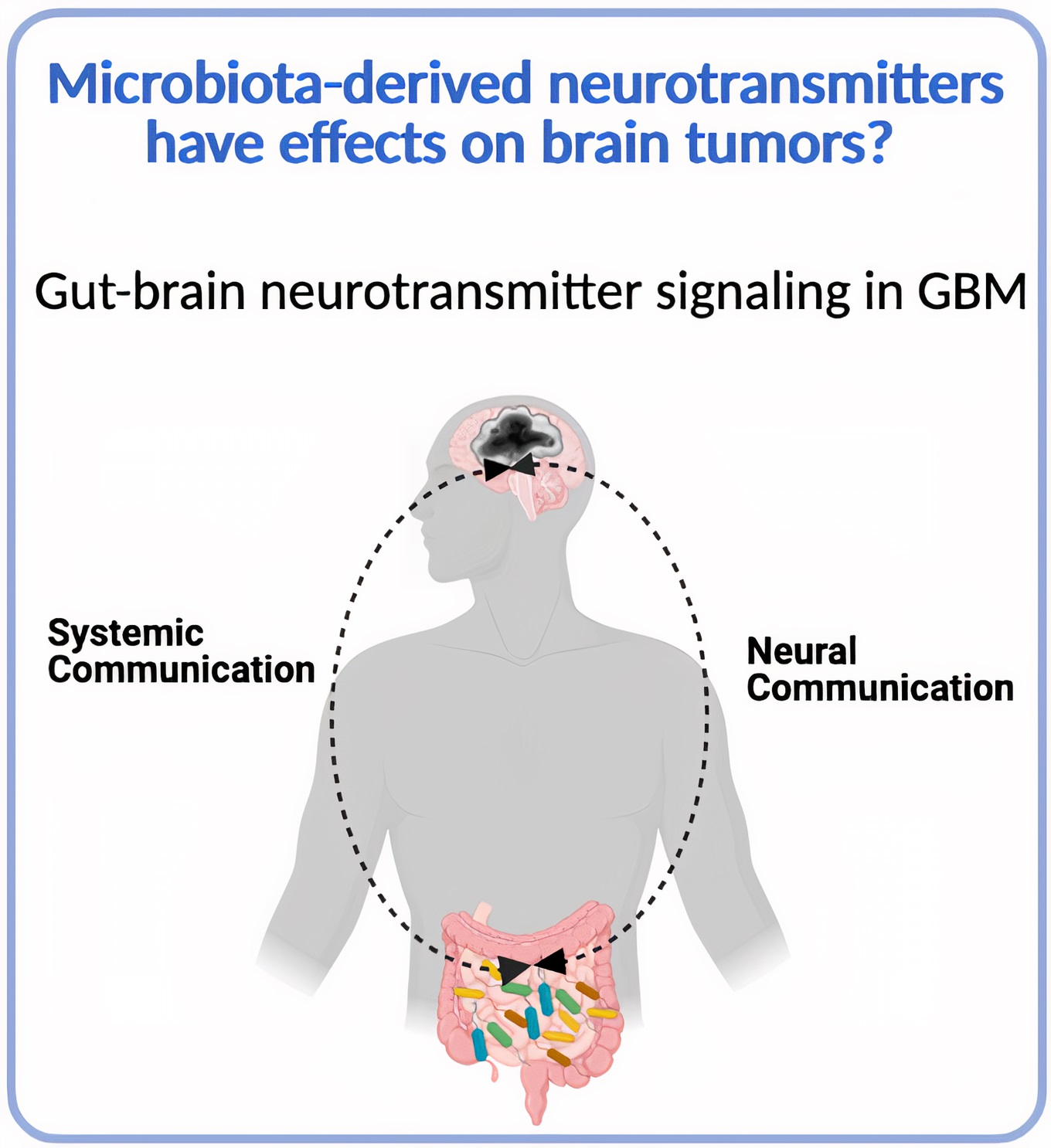
4. Do Gut Microbiota-Derived Neurotransmitters Affect Glioma Development?
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| GBM | Glioblastoma |
| GF | Germ Free |
| GABA | Gamma-aminobutyric acid |
| CNS | Central nervous system |
| 5-HT | 5-hydroxytryptamine |
| GI | Gastrointestinal |
| SCFAs | Short-chain fatty acids |
| BDNF | Brain derived neurotrophic factor |
| ASD | Autism spectrum disorder |
| PD | Parkinson’s disease |
| AD | Alzheimer’s disease |
| MS | Multiple sclerosis |
| MIA | Maternal immune activation |
| ASO | α-synuclein overexpressing |
| Aβ | Amiloid–β |
| EAE | Experimental autoimmune encephalomyelitis |
| MCAO | Middle cerebral artery occlusion |
| EECs | Enteroendocrine cells |
| ANS | Autonomic nervous system |
| ENS | Enteric nervous system |
| LPS | Lipopolysaccharide |
| GLP | Glucagon like peptide |
| LBP | LPS binding protein |
| BBB | Blood-brain barrier |
| MCTs | Monocarboxylate transporters |
| HPA | Hypothalamic-pituitary-adrenal |
| CCK | Cholecystokinin |
| NO | Nitric oxide |
| BH4 | Tetrahydrobiopterin |
| TH | Tyrosine hydroxylase |
| 5-HIAA | 5-hydroxyindoleacetic acid |
| Kyn | Kynurine |
| NMDA | N-methyl-D-aspartate |
| AMPA | Alpha-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid |
| SZ | Subventricular zone |
| OPC | Oligodendrocyte progenitor cells |
| DG | Dentate Gyrus |
| β3AR | β3 adrenergic receptor |
| GFAP | Glial fibrillary acidic protein |
| NPCs | Neuronal precursor cells |
| mGluR5 | metabotropic glutamate 5 receptor |
| LTP | Long-term potentiation |
| MAO-A | Monoamine oxidase A |
| DRD-2 | Dopamine receptor D2 |
| EGFR | Epidermal growth factor receptor |
| NK | Natural killer |
| SSRI | Selective serotonin reuptake inhibitor |
| GDNF | Glial cell line-derived neurotrophic factor |
| MMP | Matrix metalloproteinase |
| GAD | Glutamate decarboxylase |
| ABAT | 4-aminobutyrate aminotransferase |
| NLGN3 | Neuroligin-3 |
References
- De Robles, P.; Fiest, K.M.; Frolkis, A.D. The worldwide incidence and prevalence of primary brain tumors: A systematic review and meta-analysis. Neuro Oncol. 2015, 17, 776–783. [Google Scholar] [CrossRef] [PubMed]
- Koshy, M.; Villano, J.L.; Dolecek, T.A.; Howard, A.; Mahmood, U.; Chmura, S.J.; Weichselbaum, R.R.; McCarthy, B.J. Improved survival time trends for glioblastoma using the SEER 17 population-based registries. J. Neuro Oncol. 2012, 107, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Wu, X. Brain Neurotransmitter Modulation by Gut Microbiota in Anxiety and Depression. Front. Cell Dev. Biol. 2021, 9, 649103. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.R.; Yu, J.B.; Fu, J.; Pan, L.B.; Yu, H.; Han, P.; Zhang, Z.W.; Peng, R.; Xu, H.; Wang, Y. Determination and Application of Nineteen Monoamines in the Gut Microbiota Targeting Phenylalanine, Tryptophan, and Glutamic Acid Metabolic Pathways. Molecules 2021, 26, 1377. [Google Scholar] [CrossRef] [PubMed]
- Fung, T.C.; Vuong, H.E.; Luna, C.; Pronovost, G.N.; Aleksandrova, A.A.; Riley, N.G.; Vavilina, A.; McGinn, J.; Rendon, T.; Forrest, L.R.; et al. Intestinal serotonin and fluoxetine exposure modulate bacterial colonization in the gut. Nat. Microbiol. 2019, 4, 2064–2073. [Google Scholar] [CrossRef]
- Mayer, E.A.; Tillisch, K.; Gupta, A. Gut/brain axis and the microbiota. J. Clin. Investig. 2015, 125, 926–938. [Google Scholar] [CrossRef] [PubMed]
- Deng, P.; Swanson, K.S. Gut microbiota of humans, dogs and cats: Current knowledge and future opportunities and challenges. Br. J. Nutr. 2015, 113 (Suppl. S1), S6–S17. [Google Scholar] [CrossRef]
- Sender, R.; Fuchs, S.; Milo, R. Revised Estimates for the Number of Human and Bacteria Cells in the Body. PLoS Biol. 2016, 14, e1002533. [Google Scholar] [CrossRef] [PubMed]
- Ferreiro, A.; Crook, N.; Gasparrini, A.J.; Dantas, G. Multiscale Evolutionary Dynamics of Host-Associated Microbiomes. Cell 2018, 172, 1216–1227. [Google Scholar] [CrossRef]
- Lynch, S.V.; Pedersen, O. The Human Intestinal Microbiome in Health and Disease. N. Engl. J. Med. 2016, 375, 2369–2379. [Google Scholar] [CrossRef] [PubMed]
- Ogbonnaya, E.S.; Clarke, G.; Shanahan, F.; Dinan, T.G.; Cryan, J.F.; O’Leary, O.F. Adult Hippocampal Neurogenesis Is Regulated by the Microbiome. Biol. Psychiatry 2015, 78, e7–e9. [Google Scholar] [CrossRef]
- Diaz Heijtz, R.; Wang, S.; Anuar, F.; Qian, Y.; Björkholm, B.; Samuelsson, A.; Hibberd, M.L.; Forssberg, H.; Pettersson, S. Normal gut microbiota modulates brain development and behavior. Proc. Natl. Acad. Sci. USA 2011, 108, 3047–3052. [Google Scholar] [CrossRef]
- Fung, T.C.; Olson, C.A.; Hsiao, E.Y. Interactions between the microbiota, immune and nervous systems in health and disease. Nat. Neurosci. 2017, 20, 145–155. [Google Scholar] [CrossRef]
- Rhee, S.H.; Pothoulakis, C.; Mayer, E.A. Principles and clinical implications of the brain-gut-enteric microbiota axis. Nat. Rev. Gastroenterol. Hepatol. 2009, 6, 306–314. [Google Scholar] [CrossRef] [PubMed]
- Furness, J.B.; Callaghan, B.P.; Rivera, L.R.; Cho, H.J. The enteric nervous system and gastrointestinal innervation: Integrated local and central control. Adv. Exp. Med. Biol. 2014, 817, 39–71. [Google Scholar] [PubMed]
- Macia, L.; Tan, J.; Vieira, A.T.; Leach, K.; Stanley, D.; Luong, S.; Maruya, M.; McKenzie, C.I.; Hijikata, A.; Wong, C.; et al. Metabolite-sensing receptors GPR43 and GPR109A facilitate dietary fibre-induced gut homeostasis through regulation of the inflammasome. Nat. Comun. 2015, 6, 6734. [Google Scholar] [CrossRef]
- Kim, C.H.; Park, J.; Kim, M. Gut microbiota-derived short-chain fatty acids, T cells, and inflammation. Immune Netw. 2014, 14, 277–288. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.-M.; Lee, H.-J.; Jang, S.E.; Han, M.J.; Kim, D.-H. Evidence for interplay among antibacterial-induced gut microbiota disturbance, neuro-inflammation, and anxiety in mice. Mucosal Immunol. 2018, 11, 1386–1397. [Google Scholar] [CrossRef] [PubMed]
- Bailey, M.T.; Coe, C.L. Maternal separation disrupts the integrity of the intestinal microflora in infant rhesus monkeys. Dev. Psychobiol. 1999, 35, 146–155. [Google Scholar] [CrossRef]
- Osadchiy, V.; Martin, C.R.; Mayer, E.A. Gut Microbiome and Modulation of CNS Function. Compr. Physiol. 2019, 10, 57–72. [Google Scholar]
- Sudo, N.; Chida, Y.; Aiba, Y.; Sonoda, J.; Oyama, N.; Yu, X.N.; Kubo, C.; Koga, Y. Postnatal microbial colonization programs the hypothalamic-pituitary-adrenal system for stress response in mice. J. Physiol. 2004, 558, 263–275. [Google Scholar] [CrossRef] [PubMed]
- Neufeld, K.M.; Kang, N.; Bienenstock, J.; Foster, J.A. Reduced anxiety-like behavior and central neurochemical change in germ-free mice. Neurogastroenterol. Motil. 2011, 23, 255-e119. [Google Scholar] [CrossRef]
- Nishino, R.; Mikami, K.; Takahashi, H.; Tomonaga, S.; Furuse, M.; Hiramoto, T.; Aiba, Y.; Koga, Y.; Sudo, N. Commensal microbiota modulate murine behaviors in a strictly contamination-free environment confirmed by culture-based methods. Neurogastroenterol. Motil. 2013, 25, 521–528. [Google Scholar] [CrossRef] [PubMed]
- Davis, D.J.; Bryda, E.C.; Gillespie, C.H.; Ericsson, A.C. 16S rRNA amplicon sequencing dataset for conventionalized and conventionally raised zebrafish larvae. Data Brief. 2016, 8, 938–943. [Google Scholar] [CrossRef]
- Mangiola, F.; Ianiro, G.; Franceschi, F.; Fagiuoli, S.; Gasbarrini, G.; Gasbarrini, A. Gut microbiota in autism and mood disorders. World J. Gastroenterol. 2016, 22, 361–368. [Google Scholar] [CrossRef]
- Hsiao, E.Y.; McBride, S.W.; Hsien, S.; Sharon, G.; Hyde, E.R.; McCue, T.; Codelli, J.A.; Chow, J.; Reisman, S.E.; Petrosino, J.F.; et al. Microbiota modulate behavioral and physiological abnormalities associated with neurodevelopmental disorders. Cell 2013, 155, 1451–1463. [Google Scholar] [CrossRef]
- Sampson, T.R.; Debelius, J.W.; Thron, T.; Janssen, S.; Shastri, G.G.; Ilhan, Z.E.; Challis, C.; Schretter, C.E.; Rocha, S.; Gradinaru, V.; et al. Gut Microbiota Regulate Motor Deficits and Neuroinflammation in a Model of Parkinson’s Disease. Cell 2016, 167, 1469–1480.e12. [Google Scholar] [CrossRef] [PubMed]
- Minter, M.R.; Zhang, C.; Leone, V.; Ringus, D.L.; Zhang, X.; Oyler-Castrillo, P.; Musch, M.W.; Liao, F.; Ward, J.F.; Holtzman, D.M.; et al. Antibiotic-induced perturbations in gut microbial diversity influences neuro-inflammation and amyloidosis in a murine model of Alzheimer’s disease. Sci. Rep. 2016, 6, 30028. [Google Scholar] [CrossRef]
- Berer, K.; Gerdes, L.A.; Cekanaviciute, E.; Jia, X.; Xiao, L.; Xia, Z.; Liu, C.; Klotz, L.; Stauffer, U.; Baranzini, S.E.; et al. Gut microbiota from multiple sclerosis patients enables spontaneous autoimmune encephalomyelitis in mice. Proc. Natl. Acad. Sci. USA 2017, 114, 10719–10724. [Google Scholar] [CrossRef] [PubMed]
- Cekanaviciute, E.; Yoo, B.B.; Runia, T.F.; Debelius, J.W.; Singh, S.; Nelson, C.A.; Kanner, R.; Bencosme, Y.; Lee, Y.K.; Hauser, S.L.; et al. Gut bacteria from multiple sclerosis patients modulate human T cells and exacerbate symptoms in mouse models. Proc. Natl. Acad. Sci. USA 2017, 114, 10713–10718. [Google Scholar] [CrossRef]
- Benakis, C.; Brea, D.; Caballero, S.; Faraco, G.; Moore, J.; Murphy, M.; Sita, G.; Racchumi, G.; Ling, L.; Pamer, E.G.; et al. Commensal microbiota affects ischemic stroke outcome by regulating intestinal gammadelta T cells. Nat. Med. 2016, 22, 516–523. [Google Scholar] [CrossRef] [PubMed]
- Schneider, S.A.; Alcalay, R.N. Neuropathology of genetic synucleinopathies with parkinsonism: Review of the literature. Mov. Disord. 2017, 32, 1504–1523. [Google Scholar] [CrossRef] [PubMed]
- Alzheimer’s Association. Alzheimer’s disease facts and figures. J. Alzheimer’s Assoc. 2016, 12, 459–509. [Google Scholar]
- Harach, T.; Marungruang, N.; Duthilleul, N.; Cheatham, V.; Mc Coy, K.D.; Frisoni, G.; Neher, J.J.; Fåk, F.; Jucker, M.; Lasser, T.; et al. Reduction of Abeta amyloid pathology in APPPS1 transgenic mice in the absence of gut microbiota. Sci. Rep. 2017, 7, 41802. [Google Scholar] [CrossRef] [PubMed]
- Croxford, A.L.; Kurschus, F.C.; Waisman, A. Mouse models for multiple sclerosis: Historical facts and future implications. Biochim. Biophys. Acta 2011, 1812, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.K.; Menezes, J.S.; Umesaki, Y.; Mazmanian, S.K. Proinflammatory T-cell responses to gut microbiota promote experimental autoimmune encephalomyelitis. Proc. Natl. Acad. Sci. USA 2011, 108, 4615–4622. [Google Scholar] [CrossRef]
- Tankou, S.K.; Regev, K.; Healy, B.C.; Tjon, E.; Laghi, L.; Cox, L.M.; Kivisäkk, P.; Pierre, I.V.; Hrishikesh, L.; Gandhi, R. A probiotic modulates the microbiome and immunity in multiple sclerosis. Ann. Neurol. 2018, 83, 1147–1161. [Google Scholar] [CrossRef]
- D’Alessandro, G.; Antonangeli, F.; Marrocco, F.; Porzia, A.; Lauro, C.; Santoni, A.; Limatola, C. Gut microbiota alterations affect glioma growth and innate immune cells involved in tumor immunosurveillance in mice. Eur. J. Immunol. 2020, 50, 705–711. [Google Scholar] [CrossRef]
- Qiu, Q.; Lin, Y.; Ma, Y.; Li, X.; Liang, J.; Chen, Z.; Liu, K.; Huang, Y.; Luo, H.; Huang, R.; et al. Exploring the Emerging Role of the Gut Microbiota and Tumor Microenvironment in Cancer Immunotherapy. Front. Immunol. 2021, 11, 612202. [Google Scholar] [CrossRef] [PubMed]
- Cryan, J.F.; O’Riordan, K.J.; Cowan, C.S.M.; Sandhu, K.V.; Bastiaanssen, T.F.S.; Boehme, M.; Codagnone, M.G.; Cussotto, S.; Fulling, C.; Golubeva, A.V.; et al. The Microbiota-Gut-Brain Axis. Physiol. Rev. 2019, 99, 1877–2013. [Google Scholar] [CrossRef] [PubMed]
- Olivares, M.; Schüppel, V.; Hassan, A.M.; Beaumont, M.; Neyrinck, A.M.; Bindels, L.B.; Benítez-Páez, A.; Sanz, Y.; Haller, D.; Holzer, P.; et al. The Potential Role of the Dipeptidyl Peptidase-4-Like Activity From the Gut Microbiota on the Host Health. Front. Microbiol. 2018, 9, 1900. [Google Scholar] [CrossRef]
- Holzer, P.; Farzi, A. Neuropeptides and the microbiota-gut-brain axis. Adv. Exp. Med. Biol. 2014, 817, 195–219. [Google Scholar]
- Cani, P.D.; Knauf, C. How gut microbes talk to organs: The role of endocrine and nervous routes. Mol. Metab. 2016, 5, 743–752. [Google Scholar] [CrossRef]
- Neuman, H.; Debelius, J.W.; Knight, R.; Koren, O. Microbial endocrinology: The interplay between the microbiota and the endocrine system. FEMS Microbiol. Rev. 2015, 39, 509–521. [Google Scholar] [CrossRef]
- Iyer, L.M.; Aravind, L.; Coon, S.L.; Klein, D.C.; Koonin, E.V. Evolution of cell-cell signaling in animals: Did late horizontal gene transfer from bacteria have a role? Trends Genet. 2004, 20, 292–299. [Google Scholar] [CrossRef] [PubMed]
- Bogunovic, M.; Davé, S.H.; Tilstra, J.S.; Chang, D.T.; Harpaz, N.; Xiong, H.; Mayer, L.F.; Plevy, S.E. Enteroendocrine cells express functional Toll-like receptors. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 292, G1770–G1783. [Google Scholar] [CrossRef]
- Tolhurst, G.; Heffron, H.; Lam, Y.S.; Parker, H.E.; Habib, A.M.; Diakogiannaki, E.; Cameron, J.; Grosse, J.; Reimann, F.; Gribble, F.M. Short-chain fatty acids stimulate glucagon-like peptide-1 secretion via the G-protein-coupled receptor FFAR2. Diabetes 2012, 61, 364–371. [Google Scholar] [CrossRef] [PubMed]
- Chimerel, C.; Emery, E.; Summers, D.K.; Keyser, U.; Gribble, F.M.; Reimann, F. Bacterial metabolite indole modulates incretin secretion from intestinal enteroendocrine L cells. Cell Rep. 2014, 20, 1202–1208. [Google Scholar] [CrossRef] [PubMed]
- Tsavkelova, E.A.; Botvinko, I.V.; Kudrin, V.S.; Oleskin, A.V. Detection of neurotransmitter amines in microorganisms with the use of high-performance liquid chromatography. Dokl. Biochem. 2000, 372, 115–117. [Google Scholar] [PubMed]
- Shishov, V.A.; Kirovskaia, T.A.; Kudrin, V.S.; Oleskin, A.V. Amine neuromediators, their precursors, and oxidation products in the culture of Escherichia coli K-12. Prikl. Biokhim. Mikrobiol. 2009, 45, 550–554. [Google Scholar] [CrossRef] [PubMed]
- Ozogul, F. Production of biogenic amines by Morganella morganii, Klebsiella pneumoniae and Hafnia alvei using a rapid HPLC method. Eur. Food Res. Technol. 2004, 219, 465–469. [Google Scholar] [CrossRef]
- Reigstad, C.S.; Salmonson, C.E.; Rainey, J.F.; Szurszewski, J.H.; Linden, D.R.; Sonnenburg, J.L.; Farrugia, G.; Kashyap, P.C. Gut microbes promote colonic serotonin production through an effect of short-chain fatty acids on enterochromaffin cells. FASEB J. 2015, 29, 1395–1403. [Google Scholar] [CrossRef]
- Barrett, E.; Ross, R.P.; O’Toole, P.W.; Fitzgerald, G.F.; Stanton, C. γ-Aminobutyric acid production by culturable bacteria from the human intestine. J. Appl. Microbiol. 2012, 113, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Pokusaeva, K.; Johnson, C.; Luk, B.; Uribe, G.; Fu, Y.; Oezguen, N.; Matsunami, R.K.; Lugo, M.; Major, A.; Mori-Akiyama, Y. GABA-producing Bifidobacterium dentium modulates visceral sensitivity in the intestine. Neurogastroenterol. Motil. 2017, 29, e12904. [Google Scholar] [CrossRef]
- Komatsuzaki, N.; Shima, J.; Kawamoto, S.; Momose, H.; Kimura, T. Production of g-aminobutyric acid (GABA) by Lactobacillus paracasei isolated from traditional fermented foods. Food Microbiol. 2005, 22, 497–504. [Google Scholar] [CrossRef]
- Yang, S.Y.; Lü, F.X.; Lu, Z.X.; Bie, X.M.; Jiao, Y.; Sun, L.J.; Yu, B. Production of gamma-aminobutyric acid by Streptococcus salivarius subsp. thermophilus Y2 under submerged fermentation. Amino Acids 2008, 34, 473–478. [Google Scholar] [CrossRef] [PubMed]
- Stanaszek, P.M.; Snell, J.F.; O’Neill, J.J. Isolation, extraction, and measurement of acetylcholine from Lactobacillus plantarum. Appl. Environ. Microbiol. 1977, 34, 237–239. [Google Scholar] [CrossRef] [PubMed]
- Prentiss, P.G.; Rosen, H.; Brown, N.; Horowitz, R.E.; Malm, O.J.; Levenson, S.M. The metabolism of choline by the germfree rat. Arch. Biochem. Biophys. 1961, 94, 424–429. [Google Scholar] [CrossRef]
- Wang, Z.; Klipfell, E.; Bennett, B.J.; Koeth, R.; Levison, B.S.; Dugar, B.; Feldstein, A.E.; Britt, E.B.; Fu, X.; Chung, Y.-M.; et al. Gut flora metabolism of phosphatidylcholine promotes cardiovascular disease. Nature 2011, 472, 57–63. [Google Scholar] [CrossRef]
- Kim, S.H.; Ben-Gigirey, B.; Barros-Velázquez, J.; Price, R.J.; An, H. Histamine and biogenic amine production by Morganella morganii isolated from temperature-abused albacore. J. Food Prot. 2000, 63, 244–251. [Google Scholar] [CrossRef] [PubMed]
- Landete, J.M.; de Las Rivas, B.; Marcobal, A.; Muñoz, R. Molecular methods for the detection of biogenic amine-producing bacteria on foods. Int. J. Food Microbiol. 2007, 117, 258–269. [Google Scholar] [CrossRef]
- Li, G.; Young, K.D. Indole production by the tryptophanase TnaA in Escherichia coli is determined by the amount of exogenous tryptophan. Microbiology 2013, 159, 402–410. [Google Scholar] [CrossRef] [PubMed]
- Wikoff, W.R.; Anfora, A.T.; Liu, J.; Schultz, P.G.; Lesley, S.A.; Peters, E.C.; Siuzdak, G. Metabolomics analysis reveals large effects of gut microflora on mammalian blood metabolites. Proc. Natl. Acad. Sci. USA 2009, 106, 3698–3703. [Google Scholar] [CrossRef] [PubMed]
- Whitfield, C.; Trent, M.S. Biosynthesis and export of bacterial lipopolysaccharides. Annu. Rev. Biochem. 2014, 83, 99–128. [Google Scholar] [CrossRef]
- El Aidy, S.; Kunze, W.; Bienenstock, J.; Kleerebezem, M. The microbiota and the gut-brain axis: Insights from the temporal and spatial mucosal alterations during colonisation of the germfree mouse intestine. Benef. Microb. 2012, 3, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Nøhr, M.K.; Pedersen, M.H.; Gille, A.; Egerod, K.L.; Engelstoft, M.S.; Husted, A.S.; Sichlau, R.M.; Grunddal, K.V.; Poulsen, S.S.; Han, S. GPR41/FFAR3 and GPR43/FFAR2 as cosensors for short-chain fatty acids in enteroendocrine cells vs FFAR3 in enteric neurons and FFAR2 in enteric leukocytes. Endocrinology 2013, 154, 3552–3564. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, R.W.; On, N.H.; Del Bigio, M.R.; Miller, D.W.; Hatch, G.M. Fatty acid transport protein expression in human brain and potential role in fatty acid transport across human brain microvessel endothelial cells. J. Neurochem. 2011, 117, 735–746. [Google Scholar] [CrossRef] [PubMed]
- Vijay, N.; Morris, M.E. Role of monocarboxylate transporters in drug delivery to the brain. Curr. Pharm. Des. 2014, 20, 1487–1498. [Google Scholar] [CrossRef] [PubMed]
- Bonaz, B.; Bazin, T.; Pellissier, S. The Vagus Nerve at the Interface of the Microbiota-Gut-Brain Axis. Front. Neurosci. 2018, 12, 49. [Google Scholar] [CrossRef] [PubMed]
- Bravo, J.A.; Forsythe, P.; Chew, M.V.; Escaravage, E.; Savignac, H.M.; Dinan, T.G.; Bienenstock, J.; Cryan, J.F. Ingestion of Lactobacillus strain regulates emotional behavior and central GABA receptor expression in a mouse via the vagus nerve. Proc. Natl. Acad. Sci. USA 2011, 108, 16050–16055. [Google Scholar] [CrossRef] [PubMed]
- Erny, D.; Hrabě de Angelis, A.L.; Jaitin, D.; Wieghofer, P.; Staszewski, O.; David, E.; Keren-Shaul, H.; Mahlakoiv, T.; Jakobshagen, K.; Buch, T. Host microbiota constantly control maturation and function of microglia in the CNS. Nat. Neurosci. 2015, 18, 965–977. [Google Scholar] [CrossRef]
- Braniste, V.; Al-Asmakh, M.; Kowal, C.; Anuar, F.; Abbaspour, A.; Tóth, M.; Korecka, A.; Bakocevic, N.; Ng, L.G.; Kundu, P. The gut microbiota influences blood-brain barrier permeability in mice. Sci. Transl. Med. 2014, 6, 263ra158. [Google Scholar] [CrossRef]
- Asano, Y.; Hiramoto, T.; Nishino, R.; Aiba, Y.; Kimura, T.; Yoshihara, K.; Koga, Y.; Sudo, N. Critical role of gut microbiota in the production of biologically active, free catecholamines in the gut lumen of mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 303, G1288–G1295. [Google Scholar] [CrossRef]
- Hegstrand, L.R.; Hine, R.J. Variations of brain histamine levels in germ-free and nephrectomized rats. Neurochem. Res. 1986, 11, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Oleskin, A.V.; Shenderov, B.A. Neuromodulatory effects and targets of the SCFAs and gasotransmitters produced by the human symbiotic microbiota. Microb. Ecol. Health Dis. 2016, 27, 30971. [Google Scholar] [CrossRef] [PubMed]
- Carabotti, M.; Scirocco, A.; Maselli, M.A.; Severi, C. The gut-brain axis: Interactions between enteric microbiota, central and enteric nervous systems. Ann. Gastroenterol. 2015, 28, 203–209. [Google Scholar] [PubMed]
- Divyashri, G.; Krishna, G.; Muralidhara; Prapulla, S.G. Probiotic attributes, antioxidant, anti-inflammatory and neuromodulatory effects of Enterococcus faecium CFR 3003: In vitro and in vivo evidence. J. Med. Microbiol. 2015, 64, 1527–1540. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tong, Q.; Ma, S.R.; Zhao, Z.X.; Pan, L.B.; Cong, L.; Han, P.; Peng, R.; Yu, H.; Lin, Y.; et al. Oral berberine improves brain dopa/dopamine levels to ameliorate Parkinson’s disease by regulating gut microbiota. Signal Transduct. Target. Ther. 2021, 6, 77. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.-H.; Chuang, H.-L.; Huang, Y.-T.; Wu, C.-C.; Chou, G.-T.; Wang, S.; Tsai, Y.-C. Alteration of behavior and monoamine levels attributable to Lactobacillus plantarum PS128 in germ-free mice. Behav. Brain Res. 2016, 298, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Clarke, G.; Grenham, S.; Scully, P.; Fitzgerald, P.; Moloney, R.D.; Shanahan, F.; Dinan, T.G.; Cryan, J.F. The microbiome-gut-brain axis during early life regulates the hippocampal serotonergic system in a sex-dependent manner. Mol. Psychiatry 2013, 18, 666–673. [Google Scholar] [CrossRef]
- Matsumoto, M.; Kibe, R.; Ooga, T.; Aiba, Y.; Sawaki, E.; Koga, Y.; Benno, Y. Cerebral low-molecular metabolites influenced by intestinal microbiota: A pilot study. Front. Syst. Neurosci. 2013, 7, 9. [Google Scholar] [CrossRef]
- Janik, R.; Thomason, L.A.M.; Stanisz, A.M.; Forsythe, P.; Bienenstock, J.; Stanisz, G.J. Magnetic resonance spectroscopy reveals oral Lactobacillus promotion of increases in brain GABA, N-acetyl aspartate and glutamate. Neuroimage 2016, 125, 988–995. [Google Scholar] [CrossRef] [PubMed]
- Mazzoli, R.; Pessione, E. The Neuro-endocrinological Role of Microbial Glutamate and GABA Signaling. Front. Microbiol. 2016, 7, 1934. [Google Scholar] [CrossRef] [PubMed]
- Agus, A.; Planchais, J.; Sokol, H. Gut Microbiota Regulation of Tryptophan Metabolism in Health and Disease. Cell. Host Microbe 2018, 23, 716–724. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Xu, K.; Liu, H.; Liu, G.; Bai, M.; Peng, C.; Li, T.; Yin, Y. Impact of the Gut Microbiota on Intestinal Immunity Mediated by Tryptophan Metabolism. Front. Cell. Infect. Microbiol. 2018, 8, 13. [Google Scholar] [CrossRef] [PubMed]
- Kaszaki, J.; Erces, D.; Varga, G.; Szabó, A.; Vécsei, L.; Boros, M. Kynurenines and intestinal neurotransmission: The role of N-methyl-D-aspartate receptors. J. Neural. Transm. 2012, 119, 211–223. [Google Scholar] [CrossRef]
- Miladinovic, T.; Nashed, M.G.; Singh, G. Overview of Glutamatergic Dysregulation in Central Pathologies. Biomolecules 2015, 5, 3112–3141. [Google Scholar] [CrossRef]
- Snyder, S.H. Neurotransmitters, receptors, and second messengers galore in 40 years. J. Neurosci. 2009, 29, 12717–12721. [Google Scholar] [CrossRef] [PubMed]
- Benitez-Diaz, P.; Miranda-Contreras, L.; Mendoza-Briceno, R.V.; Pena-Contreras, Z.; Palacios-Pru, E. Prenatal and postnatal contents of amino acid neurotransmitters in mouse parietal cortex. Dev. Neurosci. 2003, 25, 366–374. [Google Scholar] [CrossRef]
- Ascenzi, M.; Bony, G. The building of the neocortex with non-hyperpolarizing neurotransmitters. Dev. Neurobiol. 2017, 77, 1023–1037. [Google Scholar] [CrossRef]
- Cameron, H.A.; Hazel, T.G.; Mckay, R.D. Regulation of neurogenesis by growth factors and neurotransmitters. J. Neurobiol. 1998, 36, 287–306. [Google Scholar] [CrossRef]
- Nguyen, L.; Rigo, J.M.; Rocher, V.; Belachew, S.; Malgrange, B.; Rogister, B.; Leprince, P.; Moonen, G. Neurotransmitters as early signals for central nervous system development. Cell Tissue Res. 2001, 305, 187–202. [Google Scholar] [CrossRef]
- Ojeda, J.; Avila, A. Early actions of neurotransmitters during cortex development and maturation of reprogrammed neurons. Front. Synaptic Neurosci. 2019, 11, 33. [Google Scholar] [CrossRef]
- Haydar, T.F.; Wang, F.; Schwartz, M.L.; Rakic, P. Differential modulation of proliferation in the neocortical ventricular and subventricular zones. J. Neurosci. 2000, 20, 5764–5774. [Google Scholar] [CrossRef]
- Young, S.Z.; Taylor, M.M.; Wu, S.; Ikeda-Matsuo, Y.; Kubera, C.; Bordey, A. NKCC1 knockdown decreases neuron production through GABA(A)-regulated neural progenitor proliferation and delays dendrite development. J. Neurosci. 2012, 32, 13630–13638. [Google Scholar] [CrossRef] [PubMed]
- Salazar, P.; Velasco-Velazquez, M.A.; Velasco, I. GABA effects during neuronal differentiation of stem cells. Neurochem. Res. 2008, 33, 1546–1557. [Google Scholar] [CrossRef]
- Brustein, E.; Cote, S.; Ghislain, J.; Drapeau, P. Spontaneous glycine-induced calcium transients in spinal cord progenitors promote neurogenesis. Dev. Neurobiol. 2013, 73, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Komuro, H.; Rakic, P. Modulation of neuronal migration by NMDAreceptors. Science 1993, 260, 95–97. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Miranda, L.R.; Parnavelas, J.G.; Chiara, F. Molecules and mechanisms involved in the generation and migration of cortical interneurons. ASN Neuro 2010, 2, e00031. [Google Scholar] [CrossRef]
- Murthy, S.; Niquille, M.; Hurni, N.; Limoni, G.; Frazer, S.; Chameau, P.; van Hooft, J.A.; Vitalis, T.; Dayer, A. Serotonin receptor 3A controls interneuron migration into the neocortex. Nat. Commun. 2014, 5, 5524. [Google Scholar] [CrossRef]
- Fu, Y.; Wu, X.; Lu, J.; Huang, Z.J. Presynaptic GABA(B) receptor regulates activity-dependent maturation and patterning of inhibitory synapses through dynamic allocation of synaptic vesicles. Front. Cell Neurosci. 2012, 6, 57. [Google Scholar]
- Sernagor, E.; Chabrol, F.; Bony, G.; Cancedda, L. GABAergic control of neurite outgrowth and remodeling during development and adult neurogenesis: General rules and differences in diverse systems. Front. Cell Neurosci 2010, 4, 11. [Google Scholar] [CrossRef]
- Fiorentino, H.; Kuczewski, N.; Diabira, D.; Ferrand, N.; Pangalos, M.N.; Porcher, C.; Gaiarsa, J.-L. GABA(B) receptor activation triggers BDNF release and promotes the maturation of GABAergic synapses. J. Neurosci. 2009, 29, 11650–11661. [Google Scholar] [CrossRef] [PubMed]
- Anelli, T.; Cardarelli, S.; Ori, M.; Nardi, I.; Biagioni, S.; Poiana, G. 5-Hydroxytryptamine 1A and 2B serotonin receptors in neurite outgrowth: Involvement of early growth response protein 1. Dev. Neurosci. 2013, 35, 450–460. [Google Scholar] [CrossRef] [PubMed]
- Cellot, G.; Cherubinik, E. Functional role of ambient GABA in refining neuronal circuits early in postnatal development. Front. Neural Circuits 2013, 7, 136. [Google Scholar] [CrossRef]
- Ikonomidou, C.; Bittigau, P.; Koch, C.; Genz, K.; Hoerster, F.; Felderhoff-Mueser, U.; Tenkova, T.; Dikranian, K.; Olney, J.W. Neurotransmitters and apoptosis in the developing brain. Biochem. Pharmacol. 2001, 62, 401–405. [Google Scholar] [CrossRef]
- Zhang, Y.; Bhavnani, B.R. Glutamate-induced apoptosis in neuronal cells is mediated via caspase-dependent and independent mechanisms involving calpain and caspase-3 proteases as well as apoptosis inducing factor (AIF) and this process is inhibited by equine estrogen s. BMC Neurosci. 2006, 7, 49. [Google Scholar] [CrossRef] [PubMed]
- Blanquie, O.; Liebmann, L.; Hubner, C.A.; Luhmann, H.J.; Sinning, A. NKCC1-mediated GABAergic signaling promotes postnatal cell death in neocortical Cajal-Retzius cells. Cereb. Cortex 2017, 27, 1644–1659. [Google Scholar] [CrossRef] [PubMed]
- LoTurco, J.J.; Owens, D.F.; Heath, M.J.S.; Davis, M.B.E.; Kriegstein, A.R. GABA and glutamate depolarize cortical progenitor cells and inhibit DNA synthesis. Neuron 1995, 15, 1287–1298. [Google Scholar] [CrossRef]
- Luk, K.C.; Kennedy, T.E.; Sadikot, A.F. Glutamate promotes proliferation of striatal neuronal progenitors by an NMDA receptor-mediated mechanism. J. Neurosci. 2003, 23, 2239–2250. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Represa, A.; Ben-Ari, Y. Trophic actions of GABA on neuronaldevelopment. Trends Neurosci. 2005, 28, 278–283. [Google Scholar] [CrossRef] [PubMed]
- Côté, F.; Fligny, C.; Bayard, E.; Launay, J.M.; Gershon, M.D.; Mallet, J.; Vodjdani, G. Maternal serotonin is crucial for murine embryonic development. Proc. Natl. Acad. Sci. USA 2007, 104, 329–334. [Google Scholar] [CrossRef] [PubMed]
- Xing, L.; Huttner, W.B. Neurotransmitters as Modulators of Neural Progenitor Cell Proliferation During Mammalian Neocortex Development. Front. Cell Dev. Biol. 2020, 8, 515. [Google Scholar] [CrossRef]
- Jansson, L.C.; Åkerman, K.E. The role of glutamate and its receptors in the proliferation, migration, differentiation and survival of neural progenitor cells. J. Neural Transm. 2014, 121, 819–836. [Google Scholar] [CrossRef]
- Baker, S.A.; Baker, K.A.; Hagg, T. Dopaminergic nigrostriatal projections regulate neural precursor proliferation in the adult mouse subventricular zone. Eur. J. Neurosci. 2004, 20, 575–579. [Google Scholar] [CrossRef] [PubMed]
- Höglinger, G.U.; Rizk, P.; Muriel, M.P.; Duyckaerts, C.; Oertel, W.H.; Caille, I.; Hirsch, E.C. Dopamine depletion impairs precursor cell proliferation in Parkinson disease. Nat. Neurosci. 2004, 7, 726–735. [Google Scholar] [CrossRef] [PubMed]
- Van Kampen, J.M.; Hagg, T.; Robertson, H.A. Induction of neurogenesis in the adult rat subventricular zone and neostriatum following dopamine D3 receptor stimulation. Eur. J. Neurosci. 2004, 19, 2377–2387. [Google Scholar] [CrossRef] [PubMed]
- Kippin, T.E.; Kapur, S.; van der Kooy, D. Dopamine specifically inhibits forebrain neural stem cell proliferation, suggesting a novel effect of antipsychotic drugs. J. Neurosci. 2005, 25, 5815–5823. [Google Scholar] [CrossRef] [PubMed]
- Hedlund, E.; Belnoue, L.; Theofilopoulos, S.; Salto, C.; Bye, C.; Parish, C.; Deng, Q.; Kadkhodaei, B.; Ericson, J.; Arenas, E.; et al. Dopamine Receptor Antagonists Enhance Proliferation and Neurogenesis of Midbrain Lmx1a-expressing Progenitors. Sci. Rep. 2016, 6, 26448. [Google Scholar] [CrossRef] [PubMed]
- Ohtani, N.; Goto, T.; Waeber, C.; Bhide, P.G. Dopamine modulates cell cycle in the lateral ganglionic eminence. J. Neurosci. 2003, 23, 2840–2850. [Google Scholar] [CrossRef]
- Popolo, M.; McCarthy, D.M.; Bhide, P.G. Influence of dopamine on precursor cell proliferation and differentiation in the embryonic mouse telencephalon. Dev. Neurosci. 2004, 26, 229–244. [Google Scholar] [CrossRef]
- Bongarzone, E.R.; Howard, S.G.; Schonmann, V.; Campagnoni, A.T. Identification of the dopamine D3 receptor in oligodendrocyte precursors: Potential role in regulating differentiation and myelin formation. J. Neurosci. 1998, 18, 5344–5353. [Google Scholar] [CrossRef] [PubMed]
- Brezun, J.M.; Daszuta, A. Depletion in serotonin decreases neurogenesis in the dentate gyrus and the subventricular zone of adult rats. Neuroscience 1999, 89, 999–1002. [Google Scholar] [CrossRef]
- Radley, J.J.; Jacobs, B.L. 5-HT1A receptor antagonist administration decreases cell proliferation in the dentate gyrus. Brain Res. 2002, 955, 264–267. [Google Scholar] [CrossRef]
- Encinas, J.M.; Vaahtokari, A.; Enikolopov, G. Fluoxetine targets early progenitor cells in the adult brain. Proc. Natl. Acad. Sci. USA 2006, 103, 8233–8238. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, A.; Benninghoff, J.; Moessner, R.; Rizzi, M.; Paizanis, E.; Doenitz, C.; Gross, S.; Hermann, M.; Gritti, A.; Lanfumey, L.; et al. Adult neurogenesis in serotonin transporter deficient mice. J. Neural Transm. 2007, 114, 1107–1119. [Google Scholar] [CrossRef]
- Kimoto, S.; Okuda, A.; Toritsuka, M.; Yamauchi, T.; Makinodan, M.; Okuda, H.; Tatsumi, K.; Nakamura, Y.; Wanaka, A.; Kishimoto, T. Olanzapine stimulates proliferation but inhibits differentiation in rat oligodendrocyte precursor cell cultures. Prog. Neuropsychopharmacol. Biol. Psychiatry 2011, 35, 1950–1956. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Dowlatshahi, D.; MacQueen, G.M.; Wang, J.; Young, L.T. Increased hippocampal BDNF immunoreactivity in subjects treated with antidepressant medication. Biol. Psych. 2001, 50, 260–265. [Google Scholar] [CrossRef]
- Maragnoli, M.E.; Fumagalli, F.; Gennarelli, M.; Racagni, G.; Riva, M.A. Fluoxetine and olanzapine have synergistic effects in the modulation of fibroblast growth factor 2 expression within the rat brain. Biol. Psych. 2004, 55, 1095–1102. [Google Scholar] [CrossRef]
- Cowen, D.S. Serotonin and neuronal growth factors—A convergence of signaling pathways. J. Neurochem. 2007, 101, 1161–1171. [Google Scholar] [CrossRef] [PubMed]
- Khawaja, X.; Xu, J.; Liang, J.J.; Barrett, J.E. Proteomic analysis of protein changes developing in rat hippocampus after chronic antidepressant treatment: Implications for depressive disorders and future therapies. J. Neurosci. Res. 2004, 75, 451–460. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, V.A.; Jha, S.; Vaidya, V.A. Depletion of norepinephrine decreases the proliferation, but does not influence the survival and differentiation, of granule cell progenitors in the adult rat hippocampus. Eur. J. Neurosci. 2002, 16, 2008–2012. [Google Scholar] [CrossRef] [PubMed]
- Jhaveri, D.J.; Mackay, E.W.; Hamlin, A.S.; Marathe, S.W.; Nandam, S.L.; Vaidya, V.A.; Bartlett, P.F. Norepinephrine directly activates adult hippocampal precursors via beta3-adrenergic receptors. J. Neurosci. 2010, 30, 2795–2806. [Google Scholar] [CrossRef] [PubMed]
- Jhaveri, D.J.; Nanavaty, I.; Prosper, B.W.; Marathe, S.; Husain, B.F.; Kernie, S.G.; Bartlett, P.F.; Vaidya, V.A. Opposing effects of alpha2- and beta-adrenergic receptor stimulation on quiescent neural precursor cell activity and adult hippocampal neurogenesis. PLoS ONE 2014, 9, e98736. [Google Scholar] [CrossRef] [PubMed]
- Weselek, G.; Keiner, S.; Fauser, M.; Wagenführ, L.; Müller, J.; Kaltschmidt, B.; Gerlach, M.; Redecker, C.; Hermann, A.; Alexander Storch, A. Norepinephrine is a negative regulator of the adult periventricular neural stem cell niche. Stem Cells 2020, 3, 1188–1201. [Google Scholar] [CrossRef]
- Liu, X.; Wang, Q.; Haydar, T.F.; Bordey, A. Nonsynaptic GABA signaling in postnatal subventricular zone controls proliferation of GFAP-expressing progenitors. Nat. Neurosci. 2005, 8, 1179–1187. [Google Scholar] [CrossRef] [PubMed]
- Platel, J.C.; Lacar, B.; Bordey, A. GABA and glutamate signaling: Homeostatic control of adult forebrain neurogenesis. J. Mol. Histol. 2007, 38, 602–610. [Google Scholar] [CrossRef] [PubMed]
- Platel, J.C.; Dave, K.A.; Bordey, A. Control of neuroblast production and migration by converging GABA and glutamate signals in the postnatal forebrain. J. Physiol. 2008, 586, 3739–3743. [Google Scholar] [CrossRef]
- Platel, J.C.; Dave, K.A.; Gordon, V.; Lacar, B.; Rubio, M.E.; Bordey, A. NMDA receptors activated by subventricular zone astrocytic glutamate are critical for neuroblast survival prior to entering a synaptic network. Neuron 2010, 65, 859–872. [Google Scholar] [CrossRef]
- Catavero, C.; Bao, H.; Song, J. Neural mechanisms underlying GABAergic regulation of adult hippocampal neurogenesis. Cell Tissue Res. 2018, 371, 33–46. [Google Scholar] [CrossRef] [PubMed]
- Schlett, K. Glutamate as a modulator of embryonic and adult neurogenesis. Curr. Top. Med. Chem. 2006, 6, 949–960. [Google Scholar] [CrossRef]
- Di Giorgi-Gerevini, V.; Melchiorri, D.; Battaglia, G.; Ricci-Vitiani, L.; Ciceroni, C.; Busceti, C.L.; Biagioni, F.; Iacovelli, L.; Canudas, A.M.; Parati, E.; et al. Endogenous activation of metabotropic glutamate receptors supports the proliferation and survival of neural progenitor cells. Cell Death Differ. 2005, 12, 1124–1133. [Google Scholar] [CrossRef][Green Version]
- Di Giorgi-Gerevini, V.D.; Caruso, A.; Cappuccio, I.; Ricci-Vitiani, L.; Romeo, S.; Della Rocca, C.; Gradini, R.; Melchiorri, D.; Nicoletti, F. The mGlu5 metabotropic glutamate receptor is expressed in zones of active neurogenesis of the embryonic and postnatal brain. Brain Res. Dev. Brain Res. 2004, 150, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Kitayama, T.; Yoneyama, M.; Tamaki, K.; Yoneda, Y. Regulation of neuronal differentiation by N-methyl-D-aspartate receptors expressed in neural progenitor cells isolated from adult mouse hippocampus. J. Neurosci. Res. 2004, 76, 599–612. [Google Scholar] [CrossRef]
- Cameron, H.A.; McEwen, B.S.; Gould, E. Regulation of adult neurogenesis by excitatory input and NMDA receptor activation in the dentate gyrus. J. Neurosci. 1995, 15, 4687–4692. [Google Scholar] [CrossRef]
- Nacher, J.; Rosell, D.; Alonso-Llosa, G.; McEwen, B. NMDA receptor antagonist treatment induces a long-lasting increase in the number of proliferating cells, PSA-NCAM-immunoreactive granule neurons and radial glia in the adult rat dentate gyrus. Eur. J. Neurosci. 2002, 13, 512–520. [Google Scholar] [CrossRef]
- Joo, J.Y.; Kim, B.W.; Lee, J.S.; Park, J.Y.; Kim, S.; Yun, Y.J.; Son, H. Activation of NMDA receptors increases proliferation and differentiation of hippocampal neural progenitor cells. J. Cell Sci. 2007, 120, 1358–1370. [Google Scholar] [CrossRef] [PubMed]
- Kheirbek, M.A.; Tannenholz, L.; Hen, R. NR2B-dependent plasticity of adult-born granule cells is necessary for context discrimination. J. Neurosci. 2012, 32, 8696–8702. [Google Scholar] [CrossRef] [PubMed]
- Thakker-Varia, S.; Behnke, J.; Doobin, D.; Dalal, V.; Thakkar, K.; Khadim, F.; Wilson, E.; Palmieri, A.; Antila, H.; Rantamaki, T.; et al. VGF (TLQP-62)-induced neurogenesis targets early phase neural progenitor cells in the adult hippocampus and requires glutamate and BDNF signaling. Stem Cell Res. 2014, 12, 762–777. [Google Scholar] [CrossRef] [PubMed]
- Ligon, K.L.; Huillard, E.; Mehta, S.; Kesari, S.; Liu, H.; Alberta, J.A.; Bachoo, R.M.; Kane, M.; Louis, D.N.; Depinho, R.A.; et al. Olig2-regulated lineage-restricted pathway controls replication competence in neural stem cells and malignant glioma. Neuron 2007, 53, 503–517. [Google Scholar] [CrossRef] [PubMed]
- Caragher, S.P.; Hall, R.R.; Ahsan, R.; Ahmed, A.U. Monoamines in glioblastoma: Complex biology with therapeutic potential. Neuro Oncol. 2018, 20, 1014–1025. [Google Scholar] [CrossRef] [PubMed]
- Sjöberg, R.L.; Wu, W.Y.; Dahlin, A.M.; Tsavachidis, S.; Bondy, M.L.; Melin, B.; Gliogene Group. Role of monoamine-oxidase-A-gene variation in the development of glioblastoma in males: A case control study. J. Neuro Oncol. 2019, 145, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Ostrom, Q.T.; Gittleman, H.; Xu, J.; Kromer, C.; Wolinsky, Y.; Kruchko, C.; Barnholtz-Sloan, J.S. CBTRUS Statistical Report: Primary Brain and Other Central Nervous System Tumors Diagnosed in the United States in 2009–2013. Neuro Oncol. 2016, 18 (Suppl. 5), v1–v75. [Google Scholar] [CrossRef]
- Kushal, S.; Wang, W.; Vaikari, V.P.; Kota, R.; Chen, K.; Yeh, T.S.; Jhaveri, N.; Groshen, S.L.; Olenyuk, B.Z.; Chen, T.C.; et al. Monoamine oxidase A (MAO A) inhibitors decrease glioma progression. Oncotarget 2016, 7, 13842–13853. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhu, S.; Kozono, D.; Ng, K.; Futalan, D.; Shen, Y.; Akers, J.C.; Steed, T.; Kushwaha, D.; Schlabach, M.; et al. Genome-wide shRNA screen revealed integrated mitogenic signaling between dopamine receptor D2 (DRD2) and epidermal growth factor receptor (EGFR) in glioblastoma. Oncotarget 2014, 5, 882–893. [Google Scholar] [CrossRef]
- Bartek, J.; Hodny, Z. Dopamine signaling: Target in glioblastoma. Oncotarget 2014, 5, 1116–1117. [Google Scholar] [CrossRef] [PubMed]
- Prabhu, V.V.; Morrow, S.; Kawakibi, A.R.; Zhou, L.; Ralff, M.; Ray, J.; Jhaveri, A.; Ferrarini, I.; Lee, Y.; Parker, C.; et al. ONC201 and imipridones: Anti-cancer compounds with clinical efficacy. Neoplasia 2020, 22, 725–744. [Google Scholar] [CrossRef]
- Prabhu, V.V.; Lulla, A.R.; Madhukar, N.S.; Ralff, M.D.; Zhao, D.; Kline, C.L.B.; Van den Heuvel, A.P.J.; Lev, A.; Garnett, M.J.; McDermott, U.; et al. Cancer stem cell-related gene expression as a potential biomarker of response for first-in-class imipridone ONC201 in solid tumors. PLoS ONE 2017, 12, e0180541. [Google Scholar] [CrossRef] [PubMed]
- Karpel-Massler, G.; Bâ, M.; Shu, C.; Halatsch, M.-E.; Westhoff, M.-A.; Bruce, J.N.; Peter Canoll, P.; Siegelin, M.D. TIC10/ONC201 synergizes with Bcl-2/Bcl-xL inhibition in glioblastoma by suppression of Mcl-1 and its binding partners in vitro and in vivo. Oncotarget 2015, 6, 36456–36471. [Google Scholar] [CrossRef]
- Weissenrieder, J.S.; Reed, J.L.; Green, M.V.; Moldovan, G.L.; Koubek, E.J.; Neighbors, J.D.; Hohl, R.J. The Dopamine D2 Receptor Contributes to the Spheroid Formation Behavior of U87 Glioblastoma Cells. Pharmacology 2020, 105, 19–27. [Google Scholar] [CrossRef]
- Qin, T.; Wang, C.; Chen, X.; Duan, C.; Zhang, X.; Zhang, J.; Chai, H.; Tang, T.; Chen, H.; Yue, J.; et al. Dopamine induces growth inhibition and vascular normalization through reprogramming M2-polarized macrophages in rat C6 glioma. Toxicol. Appl. Pharmacol. 2015, 286, 112–123. [Google Scholar] [CrossRef]
- Huck, J.H.; Freyer, D.; Bottcher, C.; Mladinov, M.; Muselmann-Genschow, C.; Thielke, M.; Gladow, N.; Bloomquist, D.; Mergenthaler, P.; Priller, J. De novo expression of dopamine D2 receptors on microglia after stroke. J. Cereb. Blood Flow Metab. 2015, 35, 1804–1811. [Google Scholar] [CrossRef] [PubMed]
- Arce-Sillas, A.; Sevilla-Reyes, E.; Alvarez-Luquin, D.D.; Guevara-Salinas, A.; Boll, M.C.; Perez-Correa, C.A.; Vivas-Almazan, A.V.; Rodríguez-Ortiz, U.; Castellanos Barba, C.; Hernandez, M.; et al. Expression of Dopamine Receptors in Immune Regulatory Cells. Neuroimmunomodulation 2019, 26, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Tracey, K.J. Reflex control of immunity. Nat. Rev. Immunol. 2009, 9, 418–428. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, R. Targeting dopamine receptor D3 signalling in inflammation. Oncotarget 2017, 8, 7224–7225. [Google Scholar] [CrossRef]
- Vidal, P.M.; Pacheco, R. Targeting the Dopaminergic System in Autoimmunity. J. Neuroimmune Pharmacol. 2020, 15, 57–73. [Google Scholar] [CrossRef]
- Liu, K.-H.; Yang, S.-T.; Lin, Y.-K.; Lin, J.-W.; Lee, Y.-H.; Wang, J.-Y.; Hu, C.-J.; Lin, E.-Y.; Chen, S.-M.; Then, C.-K.; et al. Fluoxetine, an antidepressant, suppresses glioblastoma by evoking AMPAR-mediated calcium-dependent apoptosis. Oncotarget 2015, 6, 5088–5101. [Google Scholar] [CrossRef] [PubMed]
- Chen, V.C.; Hsieh, Y.; Chen, L.-J.; Hsu, T.-C.; Tzang, B.-S. Escitalopram oxalate induces apoptosis in U-87MG cells and autophagy in GBM8401 cells. J. Cell. Mol. Med. 2017, 22, 1167–1178. [Google Scholar] [CrossRef]
- Wang, Y.X.; Xu, S.Q.; Chen, X.H.; Liu, R.S.; Liang, Z.Q. Autophagy involvement in olanzapine-mediated cytotoxic effects in human glioma cells. Asian Pac. J. Cancer Prev. 2014, 15, 8107–8113. [Google Scholar] [CrossRef]
- Karpel-Massler, G.; Kast, R.E.; Westhoff, M.A.; Dwucet, A.; Welscher, N.; Nonnenmacher, L.; Hlavac, M.; Siegelin, M.D.; Wirtz, C.R.; Debatin, K.M.; et al. Olanzapine inhibits proliferation, migration and anchorage-independent growth in human glioblastoma cell lines and enhances temozolomide’s antiproliferative effect. J. Neuro Oncol. 2015, 122, 21–33. [Google Scholar] [CrossRef]
- Otto-Meyer, S.; DeFaccio, R.; Dussold, C.; Ladomersky, E.; Zhai, L.; Lauing, K.L.; Bollu, L.R.; Amidei, C.; Lukas, R.V.; Scholtens, D.M.; et al. A retrospective survival analysis of Glioblastoma patients treated with selective serotonin reuptake inhibitors. Brain Behav. Immun. Health 2020, 2, 100025. [Google Scholar] [CrossRef] [PubMed]
- Sarrouilhe, D.; Clarhaut, J.; Defamie, N.; Mesnil, M. Serotonin and cancer: What is the link? Curr. Mol. Med. 2015, 15, 62–77. [Google Scholar] [CrossRef]
- Merzak, A.; Koochekpour, S.; Fillion, M.P.; Fillion, G.; Pilkington, G.J. Expression of serotonin receptors in human fetal astrocytes and glioma cell lines: A possible role in glioma cell proliferation and migration. Brain Res. Mol. Brain Res. 1996, 41, 1–7. [Google Scholar] [CrossRef]
- Siddiqui, E.J.; Thompson, C.S.; Mikhailidis, D.P.; Mumtaz, F.H. The role of serotonin in tumour growth. Oncol. Rep. 2005, 14, 1593–1597. [Google Scholar]
- Morita, K.; Arimochi, H.; Itoh, H.; Her, S. Possible involvement of 5alpha-reduced neurosteroids in adrenergic and serotonergic stimulation of GFAP gene expression in rat C6 glioma cells. Brain Res. 2006, 1085, 49–56. [Google Scholar] [CrossRef]
- Hisaoka, K.; Nishida, A.; Takebayashi, M.; Koda, T.; Yamawaki, S.; Nakata, Y. Serotonin increases glial cell line-derived neutrophic factor release in rat C6 glioblastoma cells. Brain Res 2004, 1002, 167–170. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.Y.; Leung, Y.M.; Cheung, C.W.; Chen, Y.R.; Wong, K.L. Glial cell line-derived neurotrophic factor induces cell migration and matrix metalloproteinase-13 expression in glioma cells. Biochem. Pharmacol. 2010, 80, 1201–1209. [Google Scholar] [CrossRef] [PubMed]
- Prenner, L.; Sieben, A.; Zeller, K.; Weiser, D.; Häberlein, H. Reduction of high-affinity beta2-adrenergic receptor binding by hyperforin and hyperoside on rat C6 glioblastoma cells measured by fluorescence correlation spectroscopy. Biochemistry 2007, 46, 5106–5113. [Google Scholar] [CrossRef]
- Conroy, W.G.; Peoples, R.W.; Isom, G.E. Identification of functional beta-adrenergic receptors on AC glioma cells. Biochem. Pharmacol. 1989, 38, 3175–3178. [Google Scholar] [CrossRef]
- He, J.; Zhang, W.; Liu, S. Activation of β-adrenergic receptor promotes cellular proliferation in human glioblastoma. Oncol. Lett. 2017, 14, 3846–3852. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Toll, L.; Jimenez, L.; Waleh, N.; Jozwiak, K.; Woo, A.Y.; Xiao, R.P.; Bernier, M.; Wainer, I.W. β2-adrenergic receptor agonists inhibit the proliferation of 1321N1 astrocytoma cells. J. Pharmacol. Exp. Ther. 2011, 336, 524–532. [Google Scholar] [CrossRef]
- Zhong, J.; Shan, W.; Zuo, Z. Norepinephrine inhibits migration and invasion of human glioblastoma cell cultures possibly via MMP-11 inhibition. Brain Res. 2021, 1756, 147280. [Google Scholar] [CrossRef] [PubMed]
- Johansen, M.D.; Urup, T.; Holst, C.B.; Christensen, I.J.; Grunnet, K.; Lassen, U.; Friis, S.; Poulsen, H.S. Outcome of bevacizumab therapy in patients with recurrent glioblastoma treated with angiotensin system inhibitors. Cancer Investig. 2018, 36, 512–519. [Google Scholar] [CrossRef] [PubMed]
- Nagaraja, A.S.; Armaiz-Pena, G.N.; Lutgendorf, S.K.; Sood, A.K. Why stress is BAD for cancer patients. J. Clin. Investig. 2013, 123, 558–560. [Google Scholar] [CrossRef]
- Jussofie, A.; Reinhardt, V.; Kalff, R. GABA binding sites: Their density, their affinity to muscimol and their behaviour against neuroactive steroids in human gliomas of different degrees of malignancy. J. Neural Transm. Gen. Sect. 1994, 96, 233–241. [Google Scholar] [CrossRef]
- Blanchart, A.; Fernando, R.; Häring, M.; Assaife-Lopes, N.; Romanov, R.A.; Andäng, M.; Harkany, T.; Ernfors, P. Endogenous GABAA receptor activity suppresses glioma growth. Oncogene 2017, 36, 777–786. [Google Scholar] [CrossRef]
- Gong, T.; Zhang, X.; Wei, X.; Yuan, S.; Saleh, M.G.; Song, Y.; Edden, R.A.; Wang, G. GSH and GABA decreases in IDH1-mutated low-grade gliomas detected by HERMES spectral editing at 3 T in vivo. Neurochem. Int. 2020, 141, 104889. [Google Scholar] [CrossRef] [PubMed]
- Panosyan, E.H.; Lin, H.J.; Koster, J.; Lasky, J.L. In search of druggable targets for GBM amino acid metabolism. BMC Cancer 2017, 17, 162. [Google Scholar] [CrossRef] [PubMed]
- Corsi, L.; Mescola, A.; Alessandrini, A. Glutamate Receptors and Glioblastoma Multiforme: An Old “Route” for New Perspectives. Int. J. Mol. Sci. 2019, 20, 1796. [Google Scholar] [CrossRef] [PubMed]
- Buckingham, S.C.; Campbell, S.L.; Haas, B.R.; Montana, V.; Robel, S.; Ogunrinu, T.; Sontheimer, H. Glutamate release by primary brain tumors induces epileptic activity. Nat. Med. 2011, 17, 1269–1274. [Google Scholar] [CrossRef]
- Pereira, M.S.L.; Klamt, F.; Thomé, C.C.; Worm, P.V.; de Oliveira, D.L. Metabotropic glutamate receptors as a new therapeutic target for malignant gliomas. Oncotarget 2017, 8, 22279–22298. [Google Scholar] [CrossRef] [PubMed]
- Ciceroni, C.; Bonelli, M.; Mastrantoni, E.; Niccolini, C.; Laurenza, M.; Larocca, L.M.; Pallini, R.; Traficante, A.; Spinsanti, P.; Ricci-Vitiani, L.; et al. Type-3 metabotropic glutamate receptors regulate chemoresistance in glioma stem cells, and their levels are inversely related to survival in patients with malignant gliomas. Cell Death Differ. 2013, 20, 396–407. [Google Scholar] [CrossRef] [PubMed]
- Arcella, A.; Carpinelli, G.; Battaglia, G.; D’Onofrio, M.; Santoro, F.; Ngomba, R.T.; Bruno, V.; Casolini, P.; Giangaspero, F.; Nicoletti, F. Pharmacological blockade of group II metabotropic glutamate receptors reduces the growth of glioma cells in vivo. Neuro Oncol. 2005, 7, 236–245. [Google Scholar] [CrossRef] [PubMed]
- D’Onofrio, M.; Arcella, A.; Bruno, V.; Ngomba, R.T.; Battaglia, G.; Lombari, V.; Ragona, G.; Calogero, A.; Nicoletti, F. Pharmacological blockade of mGlu2/3 metabotropic glutamate receptors reduces cell proliferation in cultured human glioma cells. J. Neurochem. 2003, 84, 1288–1295. [Google Scholar] [CrossRef]
- Ishiuchi, S.; Tsuzuki, K.; Yoshida, Y.; Yamada, N.; Hagimura, N.; Okado, H.; Miwa, A.; Kurihara, H.; Nakazato, Y.; Tamura, M.; et al. Blockage of Ca2+-permeable AMPA receptors suppresses migration and induces apoptosis in human glioblastoma cells. Nat. Med. 2002, 8, 971–978. [Google Scholar] [CrossRef] [PubMed]
- Piao, Y.; Lu, L.; de Groot, J. AMPA receptors promote perivascular glioma invasion via β1 integrin–dependent adhesion to the extracellular matrix. Neuro Oncol. 2009, 11, 260–273. [Google Scholar] [CrossRef] [PubMed]
- Ramaswamy, P.; Aditi Devi, N.; Hurmath Fathima, K.; Dalavaikodihalli-Nanjaiah, N. Activation of NMDA receptor of glutamate influences MMP-2 activity and proliferation of glioma cells. Neurol. Sci. 2014, 35, 823–829. [Google Scholar] [CrossRef] [PubMed]
- Venkatesh, H.S.; Johung, T.B.; Caretti, V.; Noll, A.; Tang, Y.; Nagaraja, S.; Gibson, E.M.; Mount, C.W.; Polepalli, J.; Mitra, S.S.; et al. Neuronal Activity Promotes Glioma Growth through Neuroligin-3 Secretion. Cell 2015, 161, 803–816. [Google Scholar] [CrossRef] [PubMed]
- Venkatesh, H.S.; Tam, L.T.; Woo, P.J.; Lennon, J.; Nagaraja, S.; Gillespie, S.M.; Ni, J.; Duveau, D.Y.; Morris, P.J.; Zhao, J.J.; et al. Targeting neuronal activity-regulated neuroligin-3 dependency in high-grade glioma. Nature 2017, 549, 533–537. [Google Scholar] [CrossRef] [PubMed]
- Venkataramani, V.; Tanev, D.I.; Strahle, C.; Studier-Fischer, A.; Fankhauser, L.; Kessler, T.; Körber, C.; Kardorff, M.; Ratliff, M.; Xie, R.; et al. Glutamatergic synaptic input to glioma cells drives brain tumour progression. Nature 2019, 573, 532–538. [Google Scholar] [CrossRef] [PubMed]
- Venkatesh, H.S.; Morishita, W.; Geraghty, A.C.; Silverbush, D.; Gillespie, S.M.; Arzt, M.; Tam, L.T.; Espenel, C.; Ponnuswami, A.; Ni, L.; et al. Electrical and synaptic integration of glioma into neural circuits. Nature 2019, 573, 539–545. [Google Scholar] [CrossRef] [PubMed]
- Rajagopala, S.V.; Vashee, S.; Oldfield, L.M.; Suzuki, Y.; Venter, J.C.; Telenti, A.; Nelson, K.E. The Human Microbiome and Cancer. Cancer Prev. Res. 2017, 10, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Patrizz, A.; Dono, A.; Zorofchian, S.; Hines, G.; Takayasu, T.; Husein, N.; Otani, Y.; Arevalo, O.; Choi, H.A.; Savarraj, J.; et al. Glioma and temozolomide induced alterations in gut microbiome. Sci. Rep. 2020, 10, 21002. [Google Scholar] [CrossRef] [PubMed]
- Dono, A.; Patrizz, A.; McCormack, R.M.; Putluri, N.; Ganesh, B.P.; Kaur, B.; McCullough, L.D.; Ballester, L.Y.; Esquenazi, Y. Glioma induced alterations in fecal short-chain fatty acids and neurotransmitters. CNS Oncol. 2020, 9, CNS57. [Google Scholar] [CrossRef] [PubMed]


| Neurotransmitters | Microbes | Effects | References |
|---|---|---|---|
| dopamine | Enterococcus faecium and Lactobacillus rhamnosus | Increased levels in healthy mouse brain | [77] |
| n.d. | Increased turnover rate in the brain striatum of GF mice | [12] | |
| Lactobacillus plantarum | Increased levels in the brain striatum of GF mice | [79] | |
| Enterococcus faecalis, Enterococcus faecium | Increased levels in brain, blood, and feces of the pseudo GF mice | [78] | |
| Enterococcus faecalis, Enterococcus faecium, Proteus mirabilis, Lactobacillus acidophilus | Increase level in vitro | [78] | |
| Clostridium species | Restored levels in the lumen of GF mice | [73] | |
| serotonin | Lactobacillus plantarum | Increased levels in the brain striatum of GF mice | [79] |
| n.d. | Increased turnover rate in the brain striatum of GF mice | [12] | |
| n.d. | Increased levels in hippocampus of male GF mice | [80] | |
| norepinephrine | Clostridium species | Restore levels in the lumen of GF mice | [73] |
| n.d | Increased turnover rate in the brain striatum of GF mice | [12] | |
| gaba | n.d. | Reduced levels in gut lumen of GF mice | [81] |
| Lactobacillus rhamnosus | Modulated GABA receptor mRNA expression in healthy mice brains | [70] | |
| Enterococcus faecium and Lactobacillus rhamnosus | Increased levels in healthy mice brains | [77] | |
| Lactobacillus rhamnosus | Increased brain levels in healthy mice | [82] | |
| glutamate | Lactobacillus rhamnosus | Increased brain levels in healthy mice | [82,83] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
D’Alessandro, G.; Lauro, C.; Quaglio, D.; Ghirga, F.; Botta, B.; Trettel, F.; Limatola, C. Neuro-Signals from Gut Microbiota: Perspectives for Brain Glioma. Cancers 2021, 13, 2810. https://doi.org/10.3390/cancers13112810
D’Alessandro G, Lauro C, Quaglio D, Ghirga F, Botta B, Trettel F, Limatola C. Neuro-Signals from Gut Microbiota: Perspectives for Brain Glioma. Cancers. 2021; 13(11):2810. https://doi.org/10.3390/cancers13112810
Chicago/Turabian StyleD’Alessandro, Giuseppina, Clotilde Lauro, Deborah Quaglio, Francesca Ghirga, Bruno Botta, Flavia Trettel, and Cristina Limatola. 2021. "Neuro-Signals from Gut Microbiota: Perspectives for Brain Glioma" Cancers 13, no. 11: 2810. https://doi.org/10.3390/cancers13112810
APA StyleD’Alessandro, G., Lauro, C., Quaglio, D., Ghirga, F., Botta, B., Trettel, F., & Limatola, C. (2021). Neuro-Signals from Gut Microbiota: Perspectives for Brain Glioma. Cancers, 13(11), 2810. https://doi.org/10.3390/cancers13112810